Differential Early Performance of Two Underplanted Hardwood Tree Species Following Restoration Treatments in High-Graded Temperate Rainforests

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites
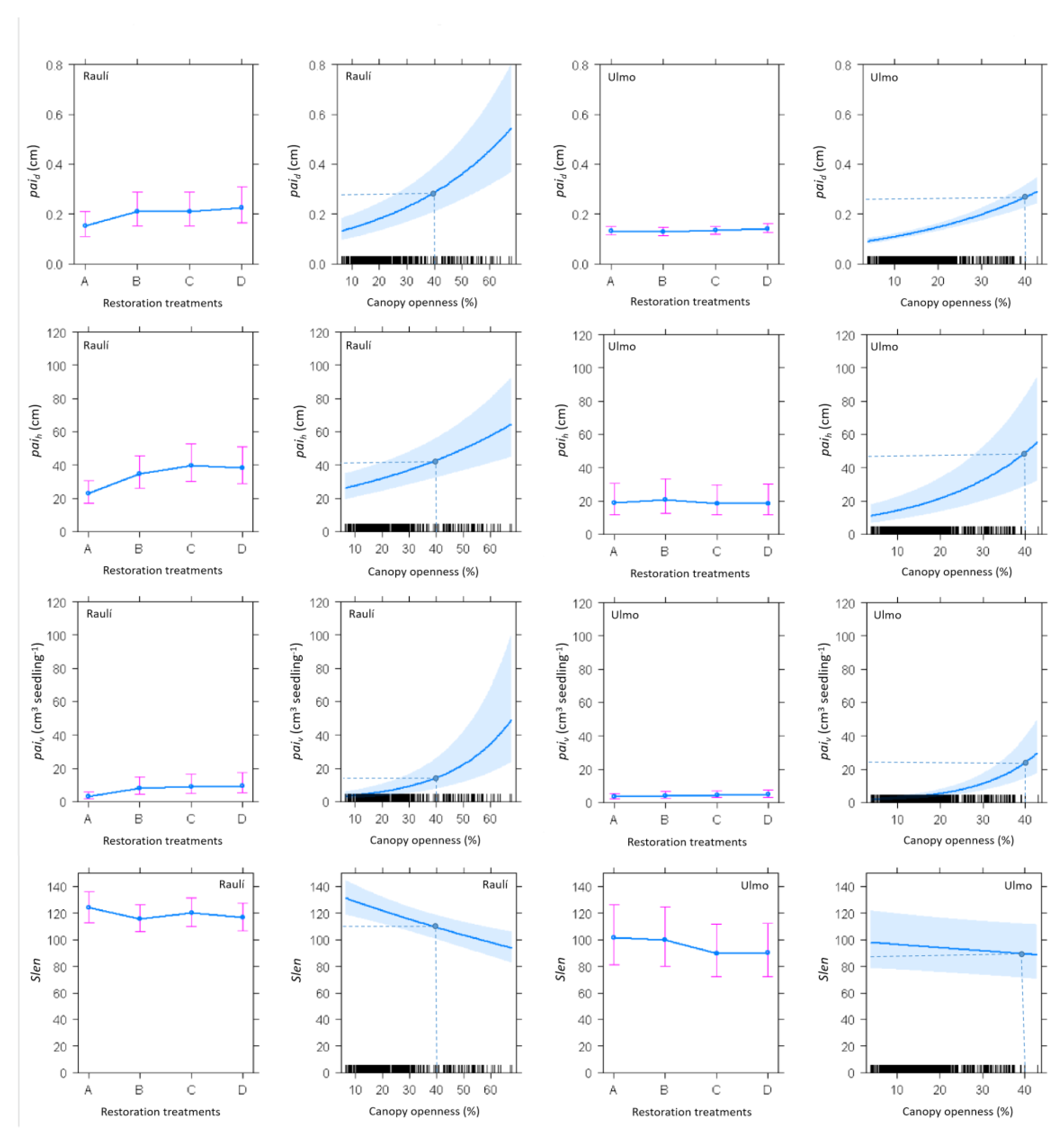
2.2. Study Design and Restoration Treatments
2.3. Plant Material
2.4. Measurements
2.5. Statistical Analyses
3. Results
4. Discussion
4.1. Growth Responses to Partial Light Conditions
4.2. Mixed Silvicultural Treatments to Favor the Development of Underplanted Seedlings in High-Graded Forest
4.3. Implications for Forests Restoration
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
Effects of the Structure Within the Random Effects

{kind=link}
{kind=link}
| Species | Response Variable | Random Variable | AIC | BIC | χ2 | p-value |
|---|---|---|---|---|---|---|
| Ulmo | paid | Site/P | −1313.7 | −1293.0 | - | - |
| Ulmo | paid | Site | −1302.4 | −1285.9 | 13.2 | <0.001 |
| Ulmo | paid | P | −1300.7 | −1284.2 | 14.9 | <0.001 |
| Ulmo | paih | Site/P | 3626.6 | 3647.3 | - | - |
| Ulmo | paih | Site | 3624.6 | 3641.1 | 52.636 | <0.001 |
| Ulmo | paih | P | 3677.3 | 3693.8 | 0 | 1 |
| Ulmo | paiv | Site/P | 2273.4 | 2306.3 | - | - |
| Ulmo | paiv | Site | 2271.4 | 2300.2 | 0 | 1 |
| Ulmo | paiv | P | 2306.9 | 2335.7 | 35.503 | <0.001 |
| Ulmo | Slen | Site/P | 4086.6 | 4115.0 | - | - |
| Ulmo | Slen | Site | 4086.1 | 4119.3 | 1.5307 | 0.216 |
| Ulmo | Slen | P | 4200.3 | 4229.1 | 115.6 | <0.001 |
| Rauli | paid | Site/P | −706.8 | −675.5 | - | - |
| Rauli | paid | Site | −697.3 | −669.9 | 11.4 | <0.001 |
| Rauli | paid | P | −678.4 | −650.9 | 30.4 | <0.001 |
| Rauli | paih | Site/P | 3136.2 | 3179.3 | - | - |
| Rauli | paih | Site | 3134.2 | 3173.4 | 0 | 1 |
| Rauli | paih | P | 3153.5 | 3192.7 | 19.313 | <0.001 |
| Rauli | paiv | Site/P | 2153.4 | 2184.7 | - | - |
| Rauli | paiv | Site | 2174.3 | 2201.7 | 22.937 | <0.001 |
| Rauli | paiv | P | 2216.3 | 2243.7 | 64.916 | <0.001 |
| Rauli | Slen | Site/P | 3469.3 | 3500.6 | - | - |
| Rauli | Slen | Site | 3468.3 | 3495.7 | 0.9499 | 0.329 |
| Rauli | Slen | P | 3470.5 | 3497.9 | 3.19 | 0.074 |
References
- Ghazoul, J.; Burilova, Z.; Garcia-Ulloa, J.; King, L.A. Conceptualizing forest degradation. Trends Ecol. 2015, 30, 622–632. [Google Scholar] [CrossRef] [PubMed]
- Vásquez-Grandón, A.; Donoso, P.J.; Gerding, V. Forest Degradation: When is a Forest Degraded? Forests 2018, 9, 726. [Google Scholar] [CrossRef] [Green Version]
- Soto, D.P.; Puettmann, K.J.; Fuentes, C.; Jacobs, D.F. Regeneration niches in Nothofagus-dominated old-growth forests after partial disturbance: Insights to overcome arrested succession. For. Ecol. Manag. 2019, 445, 26–36. [Google Scholar] [CrossRef]
- Mallik, A.U. Conifer regeneration problems in boreal and temperate forests with ericaceous understory: Role of disturbance, seedbed limitation, and keystone species change. Crit. Rev. Plant Sci. 2003, 22, 341–366. [Google Scholar] [CrossRef]
- Royo, A.A.; Carson, W.P. On the formation of dense understory layers in forests worldwide: Consequences and implications for forest dynamics, biodiversity, and succession. Can. J. For. Res. 2006, 36, 1345–1362. [Google Scholar] [CrossRef]
- Luken, J.O. Directing Ecological Succession; Chapman and Hall: New York, NY, USA, 1991. [Google Scholar]
- Paquette, A.; Bouchard, A.; Cogliastro, A. Survival and growth of underplanted trees: A meta-analysis across four biomes. Ecol. Appl. 2006, 16, 1575–1589. [Google Scholar] [CrossRef]
- Chazdon, R.L. Beyond deforestation: Restoring forests and ecosystem services on degraded lands. Science 2008, 320, 1458–1460. [Google Scholar] [CrossRef] [Green Version]
- Donoso, P.J.; Soto, D.P.; Fuentes, C. Differential growth rates through the seedling and sapling stages of two Nothofagus species underplanted at low-light environments in an Andean high-graded forest. New For. 2015, 46, 885–895. [Google Scholar] [CrossRef]
- Soto, D.P.; Donoso, P.J.; Salas, C.; Puettmann, K.J. Light availability and soil compaction influence the growth of underplanted Nothofagus following partial shelterwood harvest and soil scarification. Can. J. For. Res. 2015, 45, 998–1005. [Google Scholar] [CrossRef] [Green Version]
- Ninan, K.N.; Kontoleon, A. Valuing forest ecosystem services and disservices–Case study of a protected area in India. Ecosyst. Serv. 2016, 20, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Suding, K.N.; Gross, K.L. The Dynamics Nature of Ecological Systems: Multiple States and Restoration Trajectories. In Foundations of Restoration Ecology; Falk, D.A., Palmer, M.A., Zedler, J.B., Eds.; Island Press: Washington, DC, USA, 2006; pp. 190–209. [Google Scholar]
- Lamb, D.; Erskine, P.D.; Parrotta, J.A. Restoration of degraded tropical forest landscapes. Science 2005, 310, 1628–1632. [Google Scholar] [CrossRef] [Green Version]
- Baldini, A. CONAF, Por Un Chile Sustentable. Corporacion Nacional Forestal; Editorial Zigzag: Santiago, Chile, 2013. [Google Scholar]
- Lara, A.; Donoso, C.; Aravena, J.C. La Conservación Del Bosque Nativo En Chile: Problemas Y Desafíos. In Ecología De Los Bosques Nativos De Chile; Armesto, J., Villagran, C., Arroyo, M.K., Eds.; Editorial Universitaria: Santaigo, Chile, 1995; pp. 335–362. [Google Scholar]
- Otero, L. La Huella Del Fuego: Historia De Los Bosques Nativos: Poblamiento Y Cambios En El Paisaje Del Sur De Chile; Pehuén Editores Limitada: Providencia, Chile, 2006. [Google Scholar]
- Donoso, P.; Soto, D.P. Plantaciones con especies nativas en el centro-sur de Chile: Experiencias, desafíos y oportunidades. Revista Bosque Nativo 2010, 47, 10–17. [Google Scholar]
- Armesto, J.J.; Manuschevich, D.; Mora, A.; Smith-Ramirez, C.; Rozzi, R.; Abarzúa, A.M.; Marquet, P.A. From the Holocene to the Anthropocene: A historical framework for land cover change in southwestern South America in the past 15,000 years. Land Use Policy 2010, 27, 148–160. [Google Scholar] [CrossRef]
- Donoso, C. Las Especies Arbóreas de Los Bosques Templados de Chile y Argentina Autoecología; Marisa Cuneo Ediciones: Valdivia, Chile, 2006. [Google Scholar]
- Lindenmayer, D.B.; Laurance, W.F. The ecology, distribution, conservation and management of large old trees. Biol. Rev. 2017, 92, 1434–1458. [Google Scholar] [CrossRef]
- Donoso, C.; Deus, R.; Cockbaine, J.C.; Castillo, H. Variaciones estructurales del tipo forestal Coigue-Rauli-Tepa. Bosque 1986, 30, 17–35. [Google Scholar] [CrossRef]
- Donoso, C. Antecedentes básicos para la silvicultura del tipo forestal siempreverde. Bosque 1989, 29, 10. [Google Scholar]
- Donoso, C.; González, M.E.; Lara, A. Ecología Forestal: Bases Para el Manejo Sustentable y Conservación de los Bosques Nativos de Chile; Ediciones Universidad Austral de Chile: Valdivia, Chile, 2014. [Google Scholar]
- Glenn-Lewin, D.C.; Peet, R.K.; Veblen, T.T. Plant Succession: Theory and Prediction; Springer: Berlin/Heidelberg, Germany, 1992. [Google Scholar]
- Veblen, T.T.; Donoso, C.; Kitzberger, T.; Rebertus, A.J. Ecology of Southern Chilean and Argentinean Nothofagus Forests. In The Ecology and Biogeography of Nothofagus Forests; Hill, R.S., Read, J., Veblen, T.T., Eds.; Yale University Press: New Haven, CT, USA, 1996; pp. 93–353. [Google Scholar]
- Veblen, T.T. Forest development in tree-fall gaps in the temperature rain forests of Chile. Natl. Geogr. Res. 1985, 1, 162–183. [Google Scholar]
- Veblen, T.T.; Donoso, C.; Schlegel, F.M.; Escobar, B. Forest dynamics in south-central Chile. J. Biogeogr. 1981, 1, 211–247. [Google Scholar] [CrossRef]
- Soto, D.P.; Jacobs, D.F.; Salas, C.; Donoso, P.J.; Fuentes, C.; Puettmann, K.J. Light and nitrogen interact to influence regeneration in old-growth Nothofagus-dominated forests in south-central Chile. For. Ecol. Manag. 2017, 384, 303–313. [Google Scholar] [CrossRef]
- Donoso, P.J.; Nyland, R.D. Seedling density according to structure, dominance and understory cover in old-growth forest stands of the evergreen forest type in the coastal range of Chile. Rev. Chil. Hist. Nat. 2005, 78, 51–63. [Google Scholar] [CrossRef]
- González, M.E.; Veblen, T.T.; Donoso, C.; Valeria, L. Tree regeneration responses in lowland Nothofagus-dominated forest after bamboo dieback in South Central Chile. Plant Ecol. 2002, 161, 59–73. [Google Scholar] [CrossRef]
- Soto, D.P.; Puettmann, K.J. Topsoil removal through scarification improves natural regeneration in high-graded Nothofagus old-growth forests. J. Appl. Ecol. 2018, 55, 967–976. [Google Scholar] [CrossRef]
- Lara, A.; Soto, D.; Armesto, J.; Donoso, P.; Wernli, C. Componentes Científicos Clave Para Una Política Nacional Sobre Usos, Servicios y Conservación de Los Bosques Nativos Chilenos; ICM-UACh: St. John’s, NL, Canada, 2003. [Google Scholar]
- Donoso, P.J.; Otero, L.A. Hacia una definición de país forestal: Dónde se sitúa Chile? Bosque 2005, 26, 5–18. [Google Scholar] [CrossRef]
- Salas, C.; Donoso, P.J.; Vargas, R.; Arriagada, C.A.; Pedraza, R.; Soto, D.P. The forest sector in Chile: An overview and current challenges. J. For. 2016, 114, 562–571. [Google Scholar] [CrossRef] [Green Version]
- Soto, D.P.; Donoso, P.J.; Puettmann, K.J. Mortality in relation to growth rate and soil resistance varies by species for underplanted Nothofagus seedlings in scarified shelterwoods. New For. 2014, 45, 655–669. [Google Scholar] [CrossRef]
- González, M.E.; Donoso, C.; Fraver, S. Respuesta inicial de Eucryphia cordifolia Cav., Laurelia sempervirens R. et P. Tul. y Aextoxicon punctatum R. et P. en plantaciones mixtas en sectores recientemente florecidos con Chusquea quila Kunth en el centro-sur de Chile. Bosque 1997, 18, 53–59. [Google Scholar] [CrossRef]
- Uteau, D.; Donoso, P.J. Early individual growth of Eucryphia cordifolia and Laurelia sempervirens planted under different competition conditions in south-central Chile. Cienc. Investig. Agrar. 2009, 36, 85–96. [Google Scholar] [CrossRef] [Green Version]
- Bannister, J.R.; Vargas-Gaete, R.; Ovalle, J.F.; Acevedo, M.; Fuentes-Ramirez, A.; Donoso, P.J.; Smith-Ramírez, C. Major bottlenecks for the restoration of natural forests in Chile. Rest. Ecol. 2018, 26, 1039–1044. [Google Scholar] [CrossRef] [Green Version]
- Donoso, C. Tipos Forestales de Los Bosques Nativos de Chile; Doc. De trabajo No 38. Investigación y Desarrollo Forestal (CONAF, PNUD-FAO); FAO Chile: Santiago, Chile, 1981. [Google Scholar]
- Vásquez-Grandón, A.; Donoso, P.J.; Gerding, V. Degradación de los Bosques: Concepto, Proceso y Estado—Un Ejemplo de Aplicación en Bosques Adultos Nativos de Chile. In Silvicultura en Bosques Nativos. Experiencias en Silvicultura y Restauración en Chile, Argentina y el Oeste de Estados Unidos; Donoso, P., Promis, A., Soto, D., Eds.; OSU College of Forestry: Corvallis, OR, USA, 2018; pp. 175–196. [Google Scholar]
- CIREN. Estudio Agrológico X Región; Publicación CIREN N° 123; CIREN: Santiago, Chile, 2003. [Google Scholar]
- Dahlgren, R.; Saigusa, M.; Ugolini, F. The nature, properties and management of volcanic soils. Adv. Agron. 2004, 82, 113–182. [Google Scholar]
- Ponce, D.B.; Donoso, P.J.; Salas-Eljatib, C. Differentiating structural and compositional attributes across successional stages in Chilean temperate rainforests. Forests 2017, 8, 329. [Google Scholar] [CrossRef] [Green Version]
- Hajek, E.R.; di Castri, F. Bioclimatografía de Chile; Dirección de Investigación, Universidad Católica de Chile: Santiago, Chile, 1975. [Google Scholar]
- Donoso, P.J.; Soto, D.P.; Coopman, R.E.; Rodriguez-Bertos, S. Early performance of planted Nothofagus dombeyi and Nothofagus alpina in response to light availability and gap size in a high-graded forest in the south-central Andes of Chile. Bosque 2013, 33, 23–32. [Google Scholar] [CrossRef] [Green Version]
- Bustos, F.; González, M.E.; Donoso, P.; Gerding, V.; Donoso, C.; Escobar, B. Efectos de distintas dosis de fertilizante de liberación controlada (Osmocote®) en el desarrollo de plantas de coigüe, Rauli y ulmo. Bosque 2008, 29, 155–161. [Google Scholar] [CrossRef]
- Donoso, P.; Navarro, C.; Soto, D.; Gerding, V.; Thiers, O.; Pinares, J.; Escobar, B.; Sanhueza, M.J. Manual de Plantaciones de Rauli (Nothofagus alpina) y Coihue (Nothofagus dombeyi) en Chile; Universidad Católica de Temuco: Temuco, Chile, 2015. [Google Scholar]
- Rose, R.; Ketchum, J.S. Interaction of initial seedling diameter, fertilization and weed control on Douglas-fir growth over the first four years after planting. Ann. For. Sci. 2003, 60, 625–635. [Google Scholar] [CrossRef] [Green Version]
- Frazer, G.W.; Canham, C.D.; Lertzman, K.P. Gap Light Analyzer (GLA), Version 2.0: Imaging Software to Extract Canopy Structure and Gap Light Indices from True-Colour Fisheye Photographs; Simon Fraser University: Burnaby, BC, USA; Institute of Ecosystem Studies: Millbrook, NY, USA, 1999. [Google Scholar]
- Crawley, M.J. The R Book; John Wiley & Sons: Hoboken, NJ, USA, 2012. [Google Scholar]
- Salas, C.; Stage, A.R.; Robinson, A.P. Modeling effects of overstory density and competing vegetation on tree height growth. For. Sci. 2008, 54, 107–122. [Google Scholar]
- Salas-Eljatib, C.; Corvalán, P.; Pino, N.; Donoso, P.J.; Soto, D.P. Modelos de efectos mixtos de altura-diámetro para Drimys winteri en el sur (41–43° S) de Chile. Bosque 2019, 40, 71–80. [Google Scholar] [CrossRef] [Green Version]
- Zuur, A.; Ieno, E.N.; Walker, N.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Springer: Berlin/Heidelberg, Germany, 2009. [Google Scholar]
- Pinheiro, J.C.; Bates, D.M. Linear Mixed-Effects Models: Basic Concepts and Examples. Mix.-Effects Models S S-Plus 2000, 2000, 3–56. [Google Scholar]
- Fox, J.; Weisberg, S. Visualizing fit and lack of fit in complex regression models with predictor effect plots and partial residuals. J. Stat. Soft. 2018, 87, 9. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Escandón, A.B.; Paula, S.; Rojas, R.; Corcuera, L.A.; Coopman, R.E. Sprouting extends the regeneration niche in temperate rain forests: The case of the long-lived tree Eucryphia cordifolia. For. Ecol. Manag. 2013, 310, 321–336. [Google Scholar] [CrossRef]
- Read, J.; Hill, R.S. Photosynthetic responses to light of Australian and Chilean species of Nothofagus and their relevance to the rainforest dynamics. New Phytol. 1985, 101, 731–742. [Google Scholar] [CrossRef]
- Lusk, C.H.; Sendall, K.M.; Clarke, P.J. Seedling growth rates and light requirements of subtropical rainforest trees associated with basaltic and rhyolitic soils. Aust. J. Bot. 2014, 62, 48–55. [Google Scholar] [CrossRef] [Green Version]
- Valladares, F.; Martinez-Ferri, E.; Balaguer, L.; Perez-Corona, E.; Manrique, E. Low leaf-level response to light and nutrients in Mediterranean evergreen oaks: A conservative resource-use strategy? New Phytol. 2000, 148, 79–91. [Google Scholar] [CrossRef] [Green Version]
- Lambers, H.; Chapin, F.S., III; Pons, T.L. Plant Physiological Ecology; Springer: Berlin/Heidelberg, Germany, 2008. [Google Scholar]
- Lusk, C.H.; Contreras, O.; Figueroa, J. Growth, biomass allocation and plant nitrogen concentration in Chilean temperate rainforest tree seedlings: Effects of nutrient availability. Oecologia 1996, 109, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Valladares, F.; Niinemets, Ü. Shade tolerance, a key plant feature of complex nature and consequences. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 237–257. [Google Scholar] [CrossRef] [Green Version]
- Coates, K.D.; Burton, P.J. Growth of planted tree seedlings in response to ambient light levels in northwestern interior cedar-hemlock forests of British Columbia. Can. J. For. Res. 1999, 29, 1374–1382. [Google Scholar] [CrossRef]
- Donoso, P.J.; Muñoz, A.A.; Thiers, O.; Soto, D.P.; Donoso, C. Effects of aspect and type of competition on the early performance of Nothofagus dombeyi and Nothofagus nervosa in a mixed plantation. Can. J. For. Res. 2011, 41, 1075–1081. [Google Scholar] [CrossRef]


| Location | San Pablo de Tregua | Riñimahuida | Llancahue | Pumillahue |
|---|---|---|---|---|
| Sites Characteristics | ||||
| County | Panguipulli | Riñihue | Valdivia | Mariquina |
| Latitude | 39°36’21’’ S | 39°50’49’’ S | 39°50’56’’ S | 39°38’20’’ S |
| Longitude | 72°06’08’’ W | 72°22’11’’ W | 73°07’50’’ W | 72°45’01’’ W |
| Altitude (m a.s.l.) | 750 | 600 | 325 | 260 |
| Climate Characteristics (average) | ||||
| Annual precipitation (mm) | 3863 | 2279 | 2100 | 2500 |
| Mean annual temperature (°C) | 11 | 13 | 12 | 12 |
| Forest Characteristics (Mean ± Standard Deviation) | ||||
| Density (trees ha−1) | 914 ± 468 | 843 ± 191 | 1135 ± 306 | 775 ± 290 |
| Basal area (m2 ha−1) | 65.5 ± 10 | 25.4 ± 4 | 48.8 ± 10 | 64.7 ± 23 |
| Dg (cm) | 31.4 ± 12 | 18.8 ± 1 | 23.4 ± 13 | 32.6 ± 5 |
| Soil Characteristics | ||||
| Soil series, type and family | Liquiñe Recent volcanic ashes (Andisol) | Liquiñe Recent volcanic ashes (Andisol) | Los Ulmos Old volcanic ashes (Ultisol) | Correltúe Old volcanic ashes (Ultisol) |
| Soil thickness | <100 | <100 | <160 | <130 |
| Soil texture | Fine sandy-loam | Fine sandy-loam | clay-loam | silt-loam |
| pH (H2O) | 5.5–5.7 | 5.5–5.7 | 5.0–5.4 | 4.9–5.6 |
| Bulk density (range, g/cm3) | 0.51–0.79 | 0.51–0.79 | 0.83–1.11 | 0.67–0.84 |
| Soil water availability (range, %) | 10.6–24.7 | 10.6–24.7 | 14.0–18.8 | 13.0–22.0 |
| P retention (range, %) | 94–98 | 94–98 | 91–95 | 89–97 |
| Treatments | Description and Activities |
|---|---|
| 1. Control (untreated plots) | Underplanting without restoration treatments. |
| 2. Improvement cut | Underplanting plus improvement cut of trees with lowest quality and thinning among clumped groups of low-diameter trees with the goal to homogenize light penetration into the understory to stimulate growth of seedlings. |
| 3. Improvement cut and understory vegetation control | Same as treatment 2, but including understory control in the entire plot, which included manually cutting the shrubs and piling them outside of the plots. This treatment aimed to avoid competition of seedlings with understory vegetation. |
| 4. Improvement cut, understory vegetation control and soil scarification | Same as above (3), plus manual topsoil scarification with the goal to remove the litter layer, including small woody debris (<20 cm in diameter), which was piled out of the regeneration plots. |
| Response | Raulí | Ulmo | |||
|---|---|---|---|---|---|
| variable | Model | AIC | BIC | AIC | BIC |
| paid | 1 | −683.6 | −664.0 | −1297.7 | −1277.1 |
| paid | 2 | −637.0 | −609.6 | −1235.2 | −1206.3 |
| paid | 3 | −694.9 | −663.6 | −1294.2 | −1261.2 |
| paid | 4 | −690.0 | −646.9 | −1292.6 | −1247.3 |
| paih | 1 | 3162.5 | 3182.1 | 3544.3 | 3564.9 |
| paih | 2 | 3152.9 | 3180.3 | 3567.5 | 3596.3 |
| paih | 3 | 3124.9 | 3156.3 | 3548.3 | 3581.2 |
| paih | 4 | 3127.2 | 3170.3 | 3635.4 | 3680.7 |
| paiv | 1 | 2225.4 | 2244.9 | 2225.2 | 2258.2 |
| paiv | 2 | 2246.1 | 2273.5 | 2306.3 | 2335.2 |
| paiv | 3 | 2176.7 | 2208.0 | 2228.4 | 2248.0 |
| paiv | 4 | 2180.5 | 2223.6 | 2230.0 | 2275.3 |
| Slen | 1 | 3440.1 | 3459.7 | 4039.5 | 4060.1 |
| Slen | 2 | 3464.9 | 3492.3 | 4047.4 | 4076.2 |
| Slen | 3 | 3443.2 | 3474.6 | 4035.3 | 4068.2 |
| Slen | 4 | 3443.6 | 3486.7 | 4038.3 | 4083.7 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soto, D.P.; Donoso, P.J.; Vásquez-Grandón, A.; González-Chang, M.; Salas-Eljatib, C. Differential Early Performance of Two Underplanted Hardwood Tree Species Following Restoration Treatments in High-Graded Temperate Rainforests. Forests 2020, 11, 401. https://doi.org/10.3390/f11040401
Soto DP, Donoso PJ, Vásquez-Grandón A, González-Chang M, Salas-Eljatib C. Differential Early Performance of Two Underplanted Hardwood Tree Species Following Restoration Treatments in High-Graded Temperate Rainforests. Forests. 2020; 11(4):401. https://doi.org/10.3390/f11040401
Chicago/Turabian StyleSoto, Daniel P., Pablo J. Donoso, Angélica Vásquez-Grandón, Mauricio González-Chang, and Christian Salas-Eljatib. 2020. "Differential Early Performance of Two Underplanted Hardwood Tree Species Following Restoration Treatments in High-Graded Temperate Rainforests" Forests 11, no. 4: 401. https://doi.org/10.3390/f11040401