Patchy Fires Promote Regeneration of Longleaf Pine (Pinus palustris Mill.) in Pine Savannas


1
Tall Timbers Research Station, Tallahassee, FL 32312, USA
2
Department of Biological Sciences, Louisiana State University, Baton Rouge, LA 70803, USA
3
School of Forestry and Environmental Studies, Yale University, New Haven, CT 06511, USA
*
Author to whom correspondence should be addressed.
Forests 2019, 10(5), 367; https://doi.org/10.3390/f10050367
Submission received: 6 March 2019
/
Revised: 19 April 2019
/
Accepted: 24 April 2019
/
Published: 28 April 2019
(This article belongs to the Special Issue Longleaf Pine)
Abstract
:Research Highlights: Spatial patterns of fire spread and severity influence survival of juvenile pines in longleaf pine savannas. Small areas that do not burn during frequent fires facilitate formation of patches of even-aged longleaf pine juveniles. These regeneration patches are especially associated with inner portions of openings (gaps) and where canopy trees have died in recent decades. Patterns of prescribed fire can thus have an important influence on stand dynamics of the dominant tree in pine savannas. Background and Objectives: Savannas are characterized by bottlenecks to tree regeneration. In pine savannas, longleaf pine is noted for recruitment in discrete clusters located within gaps away from canopy trees. Various mechanisms promoting this pattern have been hypothesized: light limitations, soil moisture, soil nutrients, pine needle mulching, competition with canopy tree roots, and fire severity associated with pine needle litter. We tested the hypothesis that regeneration patches are associated with areas that remain unburned during some prescribed fires, as mediated by gaps in the canopy, especially inner portions of gaps, and areas re-opened by death of canopy trees. Materials and Methods: We mapped areas that were unburned during prescribed fires applied at 1–2 year intervals from 2005–2018 in an old-growth pine savanna in Georgia, USA. We compared the maps to locations of longleaf pine juveniles (<1.5 m height) measured in 2018 and canopy cover and canopy tree deaths using a long-term (40 year) tree census. Results: Logistic regression analysis showed juveniles to be associated with unburned areas, gaps, inner gaps, and areas where canopy trees died. Conclusions: Patterns of fire spread and severity limit survival of longleaf pine juveniles to patches away from canopy trees, especially where canopy trees have died in recent decades. These processes contribute to a buffering mechanism that maintains the savanna structure and prevents transition to closed canopy forest or open grassland communities.
1. Introduction
Worldwide, tree populations in savannas are constrained by bottlenecks to regeneration. Such limits are typically imposed on the period of the tree life cycle between seed dispersal and growth to a stage where juveniles can resist fire [1,2,3]. Mechanisms limiting tree regeneration during this period include granivory [4,5], competition with perennial grasses that dominate ground-layer vegetation [6,7,8,9,10], seasonal limitations on moisture and nutrients [11,12], and, most notably, effects of herbivory and fire [2,3,5,13,14,15,16,17,18,19]. Combinations of these mechanisms are hypothesized to constrain regeneration niches [20] to those allowing infrequent recruitment, primarily in small patches [21].
Canopy trees in savannas have been shown to influence spatial patterns of tree regeneration. Canopy cover can be facilitative, reducing drought stress and increasing nutrient availability through shading, as well as decreasing competition with grasses via litter deposition [22,23,24]. Alternatively, canopy trees can limit regeneration by removing soil moisture and nutrients [25]. Certain canopy tree species produce flammable litter [26,27,28] and impose particularly strong limits to regeneration beneath tree canopies [29,30,31,32]. This effect may be amplified by the diffuse canopies of savanna tree species, allowing persistence of flammable grasses beneath the canopy [33,34]. As a result, juvenile trees may not survive under or in the vicinity of canopy trees.
The role of fire in causing bottlenecks to tree regeneration is particularly important in mesic savannas. In savannas with relatively high precipitation and fertility, frequent fire is important for removing dense herbaceous vegetation and litter, preventing duff accumulation and allowing contact of seeds with mineral soil, as well as temporarily reducing competition with ground-layer vegetation [2,5,11,23,29,32]. High fire frequency is also necessary to prevent rapid state shifts to forests dominated by relatively fire-sensitive forest tree species, as has been observed under extended fire return intervals or fire exclusion [16,35,36,37]. Nonetheless, frequent fires threaten the survival of juveniles prior to their achieving a fire-tolerant growth stage [19,32,38]. Thus, conditions allowing tree regeneration likely include transient extensions of fire return intervals and/or reductions in fire severity [11,30,39,40,41].
Savannas of the North American Coastal Plain are well-known for spatial and temporal limits to regeneration of the dominant canopy tree, longleaf pine (Pinus palustris Mill.). These savannas have ground-layer vegetation dominated by warm-season grasses that facilitate surface fires (sensu [42]). These fires are thought to have historically occurred primarily at 1–3 year intervals, as interpreted from requirements to maintain savanna structure and associated biodiversity [43,44,45] and dendrochronological evidence [46,47,48]. Whereas most tree species in savanna systems worldwide are capable of surviving fire by resprouting [5,49], longleaf pine seedlings exhibit very restricted resprouting capability [39], requiring the entire vascular cambium and apical meristem to survive fires. Once longleaf pine seedlings reach the lifecycle stage known as the grass-stage (in which the plant resembles a grass tussock), fire tolerance provides a competitive advantage over broadleaf trees and shrubs, which are typically top-killed or completely killed in this fire-dominated landscape [32,50,51]. During the transition from the grass stage to sapling size, longleaf pines initiate rapid vertical growth, outgrowing broadleaf woody vegetation periodically top-killed by fire [50,52,53]. However, survival from seed germination to the relatively fire-tolerant grass stage is typically limited to dense, distinct clusters only within canopy gaps [29,30,38,54,55]. Although isolated juveniles also can survive at low densities, it is the regeneration patches that largely shape the savanna structure and dominate demographic processes over the long term through natural regeneration [29,56]. Frequency of reproduction is also limited by longleaf pine being a masting species, typically producing abundant seeds 2–5 time per decade [57,58,59]. The location of recruitment in gaps is attributable to pine needle litter being particularly flammable [32,50,60], thus limiting survival of longleaf seedlings and early juveniles to locations away from trees [30,39,61].
Although there is broad consensus that longleaf pine recruitment is limited to gaps, there is so far no consensus on why they are further limited to regeneration in small patches within larger gaps. Some conceptual models have linked longleaf pine seedling establishment and survival to light and nutrient availability within gaps [24,62,63,64], especially in locations farthest away from the edges of tree canopies [54]. Brockway and Outcalt [38] hypothesized that root competition with canopy trees limits seedlings to an exclusionary zone about 8 m or more from the canopy edge. Grace and Platt [65] also showed that seedlings widely distributed after mast years preferentially survived and grew at greater distances from crowns of overstory pines, both pre-fire and post-fire, with the latter attributed to decreasing fire severity with lower pine needle litter loads. The authors have observed that clusters of longleaf pine regeneration often occur where a large canopy tree has recently died. This pattern might be attributed to the thinning of ground-layer vegetation in the relatively high fire severity environment beneath large canopy trees [30,32,34,50,66] combined with the reduction in pine needle litter following the death of such trees. Thus, there are competing hypotheses regarding spatial patterns of longleaf pine regeneration.
We hypothesize that regeneration patches of longleaf pine in a naturally-functioning mesic pine savanna are primarily shaped by differential mortality of seedlings and juveniles during fires. We propose that frequent fires tend to kill seedlings except in areas that remain unburned in one or a few fires. Thus, whereas the occurrence of fire and fire effects are typical foci of demographic studies in fire-frequented ecosystems, we focus on unburned areas within the larger matrix of burned areas as a potential regeneration niche for longleaf pine. These areas are expected to be in gaps, and more specifically in the inner portion of gaps farther from adult trees, and in locations where canopy trees have died in recent decades. Using an old-growth stand of longleaf pine studied for >40 years (1978–2018), we mapped the extent of fires for the 14 most recent years (2005–2018). Then, in 2018, we mapped the locations of individual longleaf pine juveniles established in recent years. We compared locations of patches of juveniles with the patterns of tree canopy cover and canopy tree death recorded in the long-term study to explore patterns of recruitment relative to locations of unburned patches and mortality of overstory trees.
2. Materials and Methods
2.1. Study Site
We conducted our study on the privately-owned Wade Tract Preserve protected by a conservation easement on Arcadia Plantation (30°45′ N; 84°00′ W; Thomas County, GA, USA; Figure 1A). The site is located approximately 80 km north of the Gulf of Mexico on moderately dissected terrain 25–50 m above sea level on Pliocene sediments of the Miccosukee Formation [67,68]. Mean annual precipitation (recorded at Tall Timbers Research Station, 19 km to the south, 1930–2011) is 1380 mm with mean monthly temperatures ranging from 27.8 °C in July to 10.7 °C in January, with minimum daily temperatures frequently below freezing in December–February. The soils are in the Ultisol order (Typic and Arenic Kandiudults) characterized by sand or sandy loam A and E horizons and sandy clay loam Bt subhorizons [69]. The site contains 85 ha of old-growth pine savanna protected by a conservation easement held by Tall Timbers Land Conservancy and managed during the past century for northern bobwhite hunting and conservation using frequent (1–2 year interval) prescribed fires. Most trees are longleaf pine (average basal area 22 m2 ha−1) with occasional broadleaf trees, mostly Quercus spp. and Carya spp. Much of the ground layer vegetation is dominated by warm season cespitose grasses (especially Aristida beyrichiana Trinius & Ruprecht, Schizachyrium scoparium (Michaux) Nash, and Sorghastrum secundum (Elliott) Nash). There also are numerous forbs and broadleaf woody plants that are typically top-killed and resprout following fire. The plant community is species-rich, with >500 native plant species documented within the Wade Tract easement (Platt et al. unpublished data).
A road divides the easement into two similar-sized burn units (Figure 1A). Each unit has been burned in 11 prescribed fires during the 14 year period from 2005–2018 (average return interval 1.3 years), although units are burned at different times. These fires have been applied from February–June, with mean dates for both units in early-mid April with standard deviations of about one-two weeks.
2.2. Data Collection
A 50 ha mapped plot was established within the old-growth easement in 1978 (Figure 1B). All pines >1.5 m tall and all broadleaf woody plants ≥2 cm dbh were tagged, mapped, and measured for diameter at breast height (dbh, approximately 1.47 m) in 1978 [29,57]. Trees have been re-censused for growth, mortality, and recruitment measured every 3–4 years as part of the 40 year study. The most recent census was in 2017.
Beginning in 2005, we mapped sizes of areas that contained unburned vegetation within two weeks after each prescribed fire. We used a global positioning system (GPS) unit (Trimble Inc., Sunnyvale, CA, USA) with accuracies of approximately 1–5 m (ca. 2 m average). Unburned areas were limited to those greater than approximately 5 m in width. These maps provided conservative estimates of unburned areas because smaller unburned areas were somewhat common but not included.
In August and September of 2018, we located and mapped longleaf pine juveniles <1.5 m height (and therefore not yet included in the long-term census) within the 50 ha study area. Juveniles were restricted to pines in the grass stage, with secondary needles surrounding a prominent apical bud still within the ground-layer vegetation, or in initial height growth (<1.5 m tall and not yet tagged). No information was recorded on the size of juveniles, which can remain in the grass stage from one year to over a decade [39], and thus size does not necessarily reflect age. The two burn units had been burned most recently in March (west unit) and May (east unit), 2018.
2.3. Data Analysis
All data from the long-term tree census and prescribed fire mapping were converted to geodatabases in ArcMap 10.5 (ESRI, Redman, CA, USA) geographic information system (GIS) for analysis. We used these data to explore the questions posed at the onset of the study.
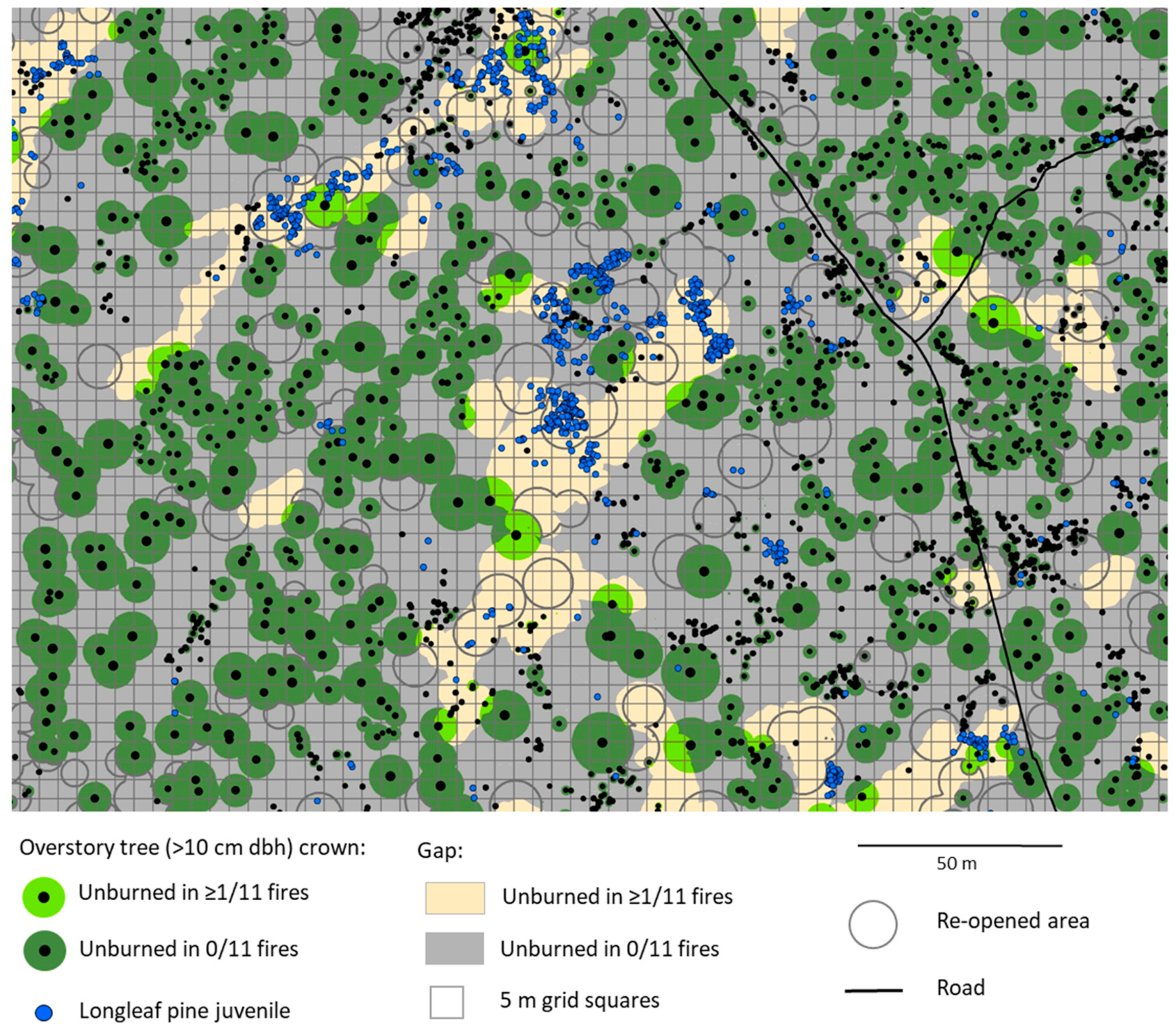
First, the study area was spatially separated into discrete land-cover categories for analysis. The entire study area, except for wetlands where pine juveniles rarely occur (1.3 ha out of the 50 ha study area), was classified as either having been unburned in at least one fire since 2005 or having been burned in every fire since 2005 (i.e., unburned in zero out of the 11 fires). The study area was also classified into areas with a tree canopy versus those without a canopy (gaps). We note that gaps in this community type are not discrete openings in a continuous canopy cover as in forests, but instead compose the matrix within which patches of trees form a canopy (Figure 2), as described for mature pine savannas in other studies [54,55,56]. Within gaps, we delineated areas that were previously under canopy trees but were re-opened by the death of one or more trees within the past four decades of tree census record, hereafter called re-opened areas (Figure 2). We also delineated areas called inner gaps covering a smaller portion of total gaps more distant from canopy trees (not shown in Figure 2) as described further below.
Maps of unburned areas following fires from 2005 to 2018 were merged into one layer to designate areas unburned in one or more fires from areas burned in all fires. Although it would be desirable to know in which years unburned areas contributed to specific instances of longleaf pine recruitment, the frequent overlap of unburned areas and inability to determine sapling age precluded such determination. In the merged layer, the polygons representing unburned areas were expanded slightly using a 2 m buffer to compensate for GPS error during mapping. Given that the expansion may result in some error of commission, juveniles mapped within these areas were interpreted as being within or along the edges of an area that was unburned during one or more fires.
Canopy areas were estimated based on the 2017 tree census. We designated canopy trees as those >10 cm dbh based on the transition from nearly pure apical dominance to lateral expansion of the canopy around this size and the capacity to produce significant amounts of pine litter fuel in a measurable area. Canopies included both pine and broadleaf trees, although >95% of trees >10 cm dbh were P. palustris. Canopy cover was estimated based on stem diameter using a modification of a previously determined allometric equation relating P. palustris stem diameter to crown area [70]. Given that trees in that study did not exceed 50 cm dbh and some trees on the Wade Tract exceed 90 cm dbh, we changed the original exponential form of the equation to a sigmoid form establishing a maximum potential crown area of approximately 200 m2 (8 m radius) based on field observations. The equation applied was r = 11.05/(1 + (d/51.74)−1.766), where r is the estimated radius of the crown in meters and d is the tree diameter in cm. The equation was used to create circular buffers around each stem which were merged into one layer indicating canopy vs. gap, although canopy cover in mature, frequently burned longleaf pine communities is diffuse and complex with actual cover around 50% [24,29,38,71]. Polygons representing inner gaps were created by buffering the canopy cover area an additional 8 m. A zone minimally this wide has been identified in other studies as having low longleaf pine recruitment and negative effects on recruits in comparison to greater distances from the canopy tree crowns [38,54,57,65,72].
A layer indicating areas re-opened by death of canopy trees was constructed from trees that were alive in the original 1978 census and were >30 cm dbh prior to death. Crown area of trees was estimated based on their stem diameter in the census before their death using the allometric equation for canopy area described above.
Analyses sought to determine the degree to which land cover categories predicted occurrence of unburned areas, and then to what degree the patches of longleaf pine juveniles were non-randomly located with regard to unburned areas and other land cover categories.
In the first analysis, we used logistic regression to test for the likelihood that areas unburned during at least one fire from 2005–2018 (as opposed to burned in each fire) were located within gaps, inner gaps, or re-opened areas. The study area was parceled into 5 m square grid cells, which was judged to be the most appropriate scale for characterizing the spatial heterogeneity of cover variables as well as typical regeneration clusters of longleaf juveniles. Each cell was assigned a binary value for each of the four cover variables (three independent and one response) according to the cover at the cell midpoint. We randomly selected 500 of the 5 m grid squares representing areas that were unburned during one or more fires, using the criterion that grid cells could not be adjacent or diagonal to each other to minimize spatial autocorrelation [73]. We randomly selected 500 additional 5 m grid squares (1000 total) outside of areas unburned in one or more fires for comparison. We used Systat 13.2 (Systat Software Inc., San Jose, CA, USA) to run backward stepwise logistic regression analyses where the full model consisted of each of the independent variables listed above and their two-way interactions. The final model retained was that with the highest (least negative) log-likelihood indicating the best fit model.
To identify potential effects of unburned areas and the other land cover categories on the location of longleaf juvenile patches, we ran a similar logistic regression analysis using 5 m grid cells. A cell was considered to represent a part of a longleaf juvenile regeneration patch if it contained one or more juveniles that were part of patch containing five or more juveniles each of which were within 3 m of another individual. Typically, such cells represented an entire individual patch, although there were exceptions where patches were exceptionally large or sinuous. These criteria were chosen to be somewhat inclusive of juveniles, given that many patches in the field have dozens of individuals spaced at higher densities. A subset of grid cells representing juvenile patches as described above was randomly selected for analysis using the same spatial criteria as described for the first analysis in order to minimize spatial autocorrelation of selected cells. The process resulted in selection of 132 cells representing longleaf juvenile patches. An equal number of cells was randomly chosen from non-juvenile patch cells within in the study area (excluding wetlands) and assigned the binary value of juveniles not present (264 cells total). A backward stepwise logistic regression analysis was run as described above.
Descriptive analyses also were conducted to assist interpretation of results of the logistic regression analyses. We reported the degree to which each land cover category overlapped with each other category. We also reported the distribution of individual longleaf pine juveniles among cover types and their combinations, providing the number, percent of total, and spatial density, as well as percentage of juveniles that were in patches as opposed to being more widely dispersed. We also compiled the distribution of longleaf juveniles with regard to the number of times their locations were unburned out of the 11 total prescribed fires from 2005 to 2018.
3. Results
3.1. Spatial Pattern of Unburned Areas
Areas that were unburned in one or more out of the 11 fires from 2005–2018 were positively associated with gaps, inner gaps, and, to a lesser extent, re-opened areas, as indicated by results of the logistic regression analysis (Table 1). The location of areas unburned in one or more fires was correctly identified 80.0% of the time by gaps, 90.2% of the time by inner gaps, and 85.2% of the time by re-opened areas (Appendix A Table A1). The degree of spatial overlap of each of the land cover categories is presented in Table 2.
3.2. Longleaf Pine Juveniles
The locations of 2565 longleaf pine juveniles were recorded using GPS within the 49.4 ha census area that excluded wetlands. Results from the logistic regression analysis showed positive associations of longleaf pine juveniles with areas unburned in one or more fires, gaps, inner gaps, and re-opened areas (Table 3). The interaction terms in the model were non-significant and so were not included in the final model. The overall model had modest accuracy, as indicated by the 63.6% rate of correct prediction of location of longleaf juvenile clusters, area under the Receiver Operator Characteristic (ROC) curve of 0.730 out of 1.000, and low pseudo-R2 values (Table 3). Thus, while longleaf juvenile patches were strongly distributed toward gaps, inner gaps, and re-opened areas, the relatively low accuracy of the model resulted from much of the area of these land cover categories not harboring longleaf juvenile patches.
The patterns of overall density of longleaf juveniles among land cover categories reflected the results of the logistic regression analysis (Table 4), showing apparently independent and additive effects of gap, inner gap, and re-opened areas on density of juveniles (inner gap and re-opened areas overlapping with gap by definition). There were progressively higher densities of juveniles in the order of areas under tree canopies, areas in gaps in general, inner gaps, re-opened areas, and portions of inner gaps within re-opened areas (IG/RO) (Table 4). This pattern was evident both within areas that were unburned in one or more fires and areas unburned in zero out of 11 fires (burned in each fire) (Table 4). More specifically, longleaf juvenile density within areas unburned during one or more fires ranged upward from 62.2 juveniles per hectare under tree canopies to 446.7 juveniles per hectare in re-opened areas within inner gaps, and in areas unburned in zero out 11 fires (burned in each fire) juvenile density ranged from 11.1 juveniles/hectare under tree canopies to 71.3 juveniles per hectare in re-opened areas within inner gaps (Table 4). The percentage of juveniles that were a part of regeneration patches as defined in this study was consistently higher in areas unburned in one or more fires, exceeding 90% in gap, inner-gap, re-opened, and inner-gap re-opened areas (Table 4).
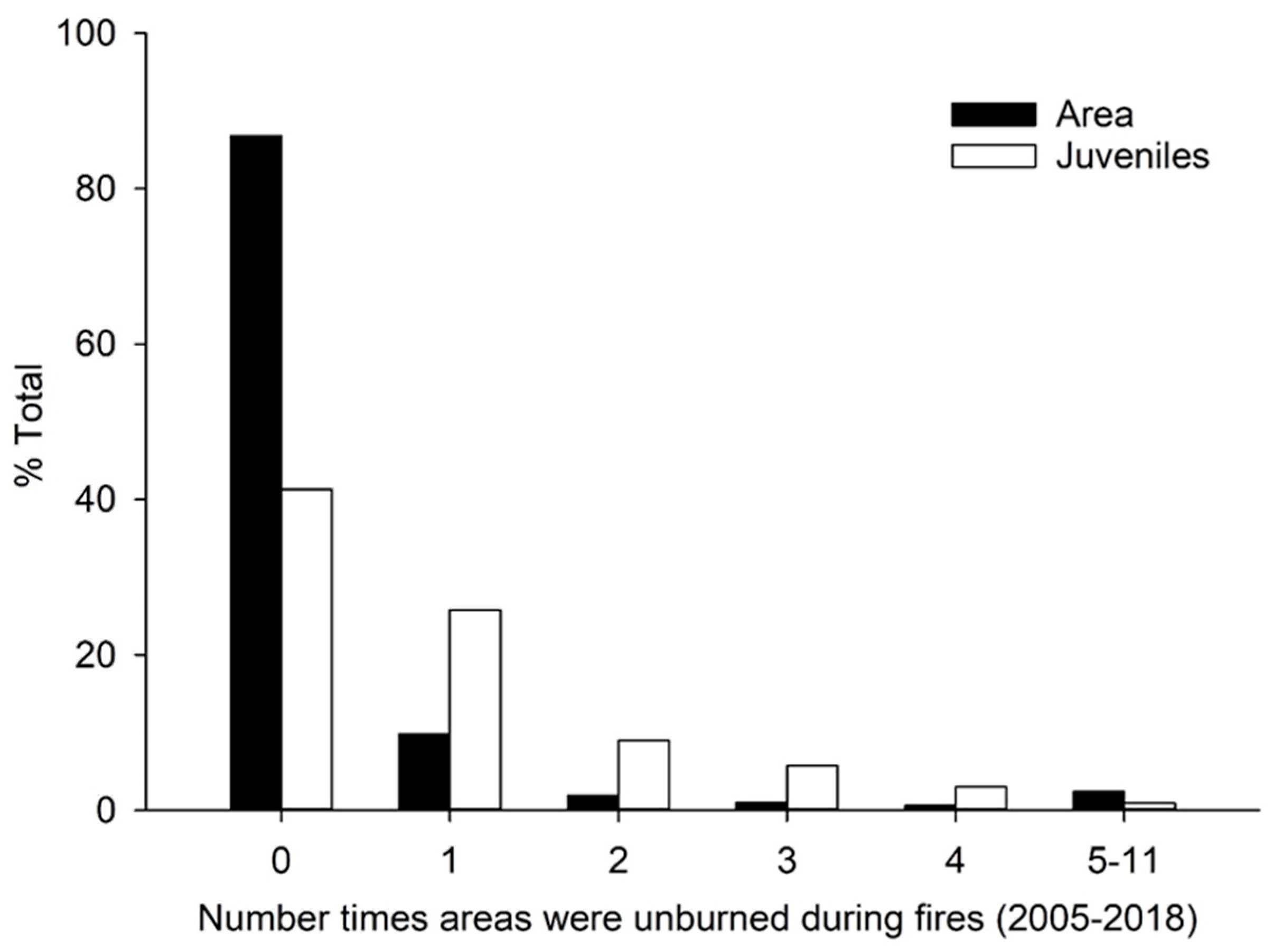
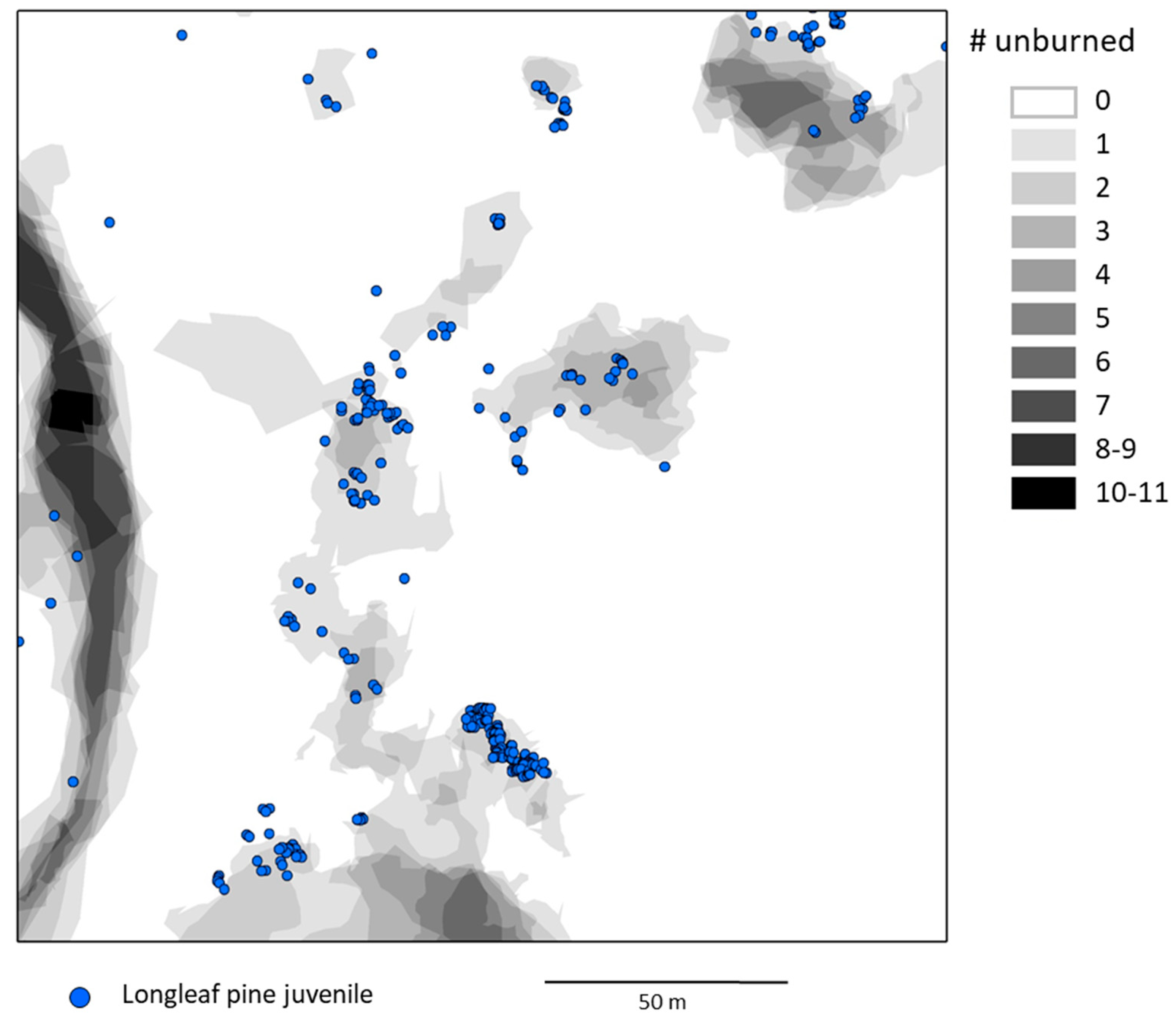
Most juveniles were in areas that were unburned during either one or a few of the total 11 fires from 2005–2018. Of the 58.7% of juveniles in areas unburned during one or more fires, about half were in areas that were unburned in only one fire, and progressively smaller percentages were in areas that were unburned in a larger number of fires (Figure 3). Very few juveniles were within areas that were unburned in more than four fires, even relative to the small total area in that category (Figure 3 and Figure 4).
4. Discussion
Results of our study support our hypothesis that recruitment of longleaf pine in patches within gaps is shaped by spatial patterns of fire. In the context of 1–2 year fire return intervals over the past 1.5 decades, most recruitment into the grass stage has occurred in areas that were unburned during one or more fires, but not in areas that were unburned during most fires. The locations and extent of periodically unburned areas and associated longleaf pine juveniles were spatially associated with gaps (absence of longleaf pine canopy cover), and especially inner gaps, consistent with previous observations of the distribution of juveniles [29,30,31,32,38]. Although many longleaf juveniles were in areas burned in each fire, juvenile clusters were concentrated in the areas that were unburned in one or more fires. Thus, the spatial distribution of burned versus unburned areas over the previous 1–2 decades provides an explanation as to why longleaf pine juveniles that enter tree life cycle stages are typically in discrete patches of closely-spaced individuals, as often observed in naturally regenerating longleaf pine savannas [29,30,57].
The positive and statistically independent effect of gaps on location of longleaf juveniles suggests that tree canopy cover influences seedlings prior to or in between exposure to fire. Abundance and distribution of longleaf pine seeds during mast years do not appear to be limiting [65,74]. However, after germination, canopy cover has a negative effect on seedling and juvenile growth rates, variously attributed to root competition with canopy trees, mulching by pine needles [24,38,57,63,64,65,75,76,77], and possibly light, although light levels are typically characterized by only a slight gradient or no gradient with distance from gap edge [24,38,62,71]. Unlike growth rates, survival of juveniles prior to fire appears not to be strongly influenced by canopy cover, or even may be positively influenced during droughts [24,63,64,75,76] (but see [66]). Evidence for effects of soil moisture and nutrients on juvenile growth and survival are mixed [24,34,62,64,77]. Although these local environmental conditions probably have an influence in the absence of fire, it is likely that their greatest influence is through predisposing juveniles to survive or die during subsequent fires. Past studies have shown that smaller sized juveniles (measured as root collar diameter, needle production, or height growth) prior to fires are less likely to survive fire [30,72,76]. Combined effects of these different environmental conditions on growth and survival of pre-grass stage juveniles are predicted to decrease with increasing distance from overstory trees, contributing to higher survival during fires away from canopy trees.
The positive association of longleaf juveniles with areas re-opened by death of canopy trees suggests legacy effects of canopy pines on the vegetation fuel structure and/or changes in fire behavior following tree death. While the tree is living, bunchgrasses and woody plants tend to be reduced somewhat by competition with canopy trees [24,77], mulching effects of pine needles [30,77,78], and damage from increased fire severity associated with deposition of pine needles, bark, branches, and cones [31,32,34,66,72]. Once the canopy tree has died, reduced ground layer fuels and a decrease in needle deposition likely reduce fire severity and spread, thereby increasing juvenile survival. Although survival and growth of longleaf juveniles in re-opened areas could also be ameliorated by decreased competition with ground-layer plants [57,63,75,77,78], artificial creation of gaps by tree felling has been shown to decrease longleaf juvenile survival, attributed to release of competing ground-layer plants [24,76]. It is possible that under the natural conditions of the Wade Tract (where dead trees are not cut for salvage), deposition of coarse woody debris following death of trees causes localized high severity combustion [31,79,80] resulting in small areas with reduced densities of ground-layer plants amenable to longleaf survival. Also, fallen tree trunks and large branches locally block fire spread and temporarily protect some juveniles from fire [81]. Thus, the effect of re-opened areas on location of longleaf juveniles appears to be both through the increased probability that the area will be unburned as mapped in this study and through smaller-scale patterns of fire not related the mapped unburned areas.
The influences of pine savanna canopy cover on fire spread and severity and associated patterns of pine regeneration appear to present an example of ecological buffering (sensu [16,25,82]). Effects of trees on fire spread and intensity may reduce chances of transition from mesic savanna to either closed-canopy forest or treeless grassland. Specifically, low levels of canopy cover promote unburned areas and opportunities for tree regeneration, while higher levels of canopy cover promote more complete burning with increased fire severity and reduced tree recruitment. It is well established that pine needle litter fuel significantly increases fire intensity, total heat release, and duration of heating relative to grasses alone [30,31,32,66].
Pine savannas of the North American Coastal Plain resemble savanna types worldwide in having bottlenecks to tree regeneration. Such bottlenecks are widely acknowledged to maintain savanna states via different combinations of fire and herbivory [3,17]. Nonetheless, the life cycle stages involved and the evolutionary responses of savanna trees to such bottlenecks varies among savannas dominated by different tree species. Generally, fire is the primary mechanism limiting recruitment of trees, such that only when juveniles escape the “fire trap” for some critical time does recruitment occur into tree life cycle stages [2,19,83]. However, longleaf pine differs from most savanna tree species in that they are unlikely to resprout after fire, such that new recruits only enter the tree life cycle stages if they avoid fires severe enough to kill above-ground tissues from germination onward. Although longleaf juveniles that reach the grass stage are exceptionally fire tolerant [53,72], they are well-known to require periods without fire, or considerably reduced fire severity, to survive their first few years [29,30,39,53,61,84,85]. In contrast to many other savannas, where savanna tree leaf litter has a neutral or negative effect on fire severity relative to that of surrounding grass-dominated fuel types (e.g., [42,86]), tree-stage longleaf pines have a positive effect on fire severity, greatly reducing the area suitable for regeneration. Our analysis indicates that fire regimes characterized by limited areas with no fire or low fire severity, mediated by spatial patterns of canopy cover and canopy tree death, widen the bottleneck, resulting in longleaf pine regeneration that occurs periodically and typically in small patches.
Our results have implications for fire management in longleaf pine ecosystems. Where natural longleaf pine regeneration is desired, applying prescribed fire in a manner that provides unburned areas during the first few years following a mast year is beneficial. Alternatively, fire could be excluded from the whole unit for such a period. However, heavy fuel accumulation, especially under canopy trees where successful regeneration is not needed nor expected to be successful, would likely result in undesirable levels of fire severity after the fire-free interval, and such wide-spread fire exclusion is counterproductive to most wildlife management goals [87]. Thus, frequent burning (1–2 year intervals) with patchy burns following mast years are recommended. Patchy burning also precludes the need for protecting longleaf regeneration patches using disking or other methods resulting in soil disturbance, which degrades native ground-layer plant communities [88]. The capacity for unburned patches to promote longleaf pine regeneration and thereafter increase fuel loads and fire severity should also reduce concerns about increased woody plant cover in temporarily unburned areas. Results of this study provide a concrete example of the application of "pyrodiversity" (sensu [89]) and patch mosaic burning [90] for accomplishing conservation and forest management goals, in light of criticism of their application in the absence of demonstrated efficacy [91].
5. Conclusions
Spatial patterns of fire spread and severely influence locations where juvenile longleaf pine become established as patches of grass-stage recruits. Specifically, regeneration patches tend to occur in small areas that remain unburned during one or a few fires, presumably reflecting mortality of most longleaf pine seedlings in burned areas following mast years. Both temporarily unburned areas and associated longleaf pine regeneration patches tend to be associated with gaps, particularly the inner portion further from the edges of tree crowns, and in areas where canopy trees have died in recent decades. These patterns are primarily attributable to variation in pine needle fuel loads and the associated likelihood of burning and having relatively high fire severity. Thus, spatial heterogeneity in the structure of pine populations and resulting fire effects have an important influence on longleaf pine population dynamics as well as the maintenance of the savanna state.
Author Contributions
Conceptualization, K.M.R., W.J.P.; methodology, K.M.R.; formal analysis, K.M.R.; investigation, C.E.F., K.M.R.; writing, K.M.R., W.J.P., C.E.F. The long-term research on which this study is based has been developed by W.J.P.
Funding
This research was funded by Tall Timbers Research, Inc. The long-term study that formed the basis for this current study has been funded by Tall Timbers Research, Inc., with different phases funded through the National Science Foundation by individual and collaborative research awards (BSR 8012090 & 8605318, W.P., PI; BSR-8718803 & 8718993, W.P. & J.H. PIs; DEB-8907138, W.P., PI; DEB 0950347 & 0950302, W.P. & B.B., PIs; NSF DEB 1557000 & 1556837, W.P. & B.S., PIs)
Acknowledgments
We thank Tom Ostertag, Jason Isbell, Angie Reid, Monica Rother, Michelle Smith, Cinnamon Dixon, and Allison Snyder for assistance with mapping unburned areas, and we thank three anonymous reviewers for their contribution to improving the manuscript. Many people over the past 40 years have assisted with various aspects of the long-term study, especially Maynard Hiss, Brian Beckage, Jim Hamrick and Ben Sikes. We thank the Wade Foundation and acknowledge the legacy of Jeptha Wade and Bruce Means in establishment of the Wade Tract easement.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table A1.
Overall results from logistic regression analysis presented in Table 3 testing the effects of Gap, Inner Gap, and Re-opened areas on areas unburned (UB) in ≥1 of 11 prescribed fires vs. unburned in 0 of 11 fires (burned in every fire) from 2005–2018 in separate analyses. Analyses are based 500 random points located in each of the burn categories.
Table A1.
Overall results from logistic regression analysis presented in Table 3 testing the effects of Gap, Inner Gap, and Re-opened areas on areas unburned (UB) in ≥1 of 11 prescribed fires vs. unburned in 0 of 11 fires (burned in every fire) from 2005–2018 in separate analyses. Analyses are based 500 random points located in each of the burn categories.
| Gap | |||
| Chi-square | 65.75, 1 d.f., p < 0.001 | UB in ≥1 fire % correct | 80.0 |
| Cox & Snell R-sq | 0.064 | UB in 0 fires % correct | 43.7 |
| Nagelkerke’s R-sq | 0.085 | Total % correct | 61.9 |
| ROC AUC | 0.618 | Log likelihood constant model | −692.5 |
| Log likelihood full model | −659.6 | ||
| Inner Gap | |||
| Chi-square | 69.6, 1 d.f., p < 0.001 | UB in ≥1 fire % correct | 90.2 |
| Cox & Snell R-sq | 0.067 | UB in 0 fires % correct | 30.6 |
| Nagelkerke’s R-sq | 0.090 | Total % correct | 60.4 |
| ROC AUC | 0.604 | Log likelihood constant model | −692.5 |
| Log likelihood full model | −657.7 | ||
| Re-Opened Area | |||
| Chi-square | 4.66.7, 1 d.f., p = 0.031 | UB in ≥1 fire % correct | 85.2 |
| Cox & Snell R-sq | 0.005 | UB in 0 fires % correct | 20.0 |
| Nagelkerke’s R-sq | 0.006 | Total % correct | 52.6 |
| ROC AUC | 0.526 | Log likelihood constant model | −692.5 |
| Log likelihood full model | −690.1 | ||
References
- Ratnam, J.; Bond, W.J.; Fensham, R.J.; Hoffmann, W.A.; Archibald, S.; Lehmann, C.E.R.; Anderson, M.T.; Higgins, S.I.; Sankaran, M. Where is a ’forest’ a savanna, and why does it matter. Glob. Ecol. Biogeogr. 2011, 20, 653–660. [Google Scholar] [CrossRef]
- Werner, P.A.; Prior, L.D. Demography and growth of subadult savanna trees: interactions of life history, size, fire season, and grassy understory. Ecol. Monogr. 2013, 83, 67–93. [Google Scholar] [CrossRef]
- Staal, A.; van Nes, E.H.; Hantson, S.; Homgren, M.; Dekker, S.C.; Pueyo, S.; Xu, C.; Scheffer, M. Reslience of tropical tree cover: The roles of climate, fire, and herbivory. Glob. Change Biol. 2018, 24, 5096–5109. [Google Scholar] [CrossRef]
- Wellington, A.B.; Noble, I.R. Seed dynamics and factors limiting recruitment of the mallee Eucalyptus incrassata Labill. in semi-arid, south-eastern Australia. J. Ecol. 1985, 73, 657–666. [Google Scholar] [CrossRef]
- Setterfield, S.A. Seedling establishment in an Australian tropical savanna: effects of seed supply, soil disturbance and fire. J. Appl. Ecol. 2002, 39, 949–959. [Google Scholar] [CrossRef] [Green Version]
- Scholes, R.J.; Archer, S.R. Tree-grass interactions in savannas. Ann. Rev. Ecol. Syst. 1997, 28, 517–544. [Google Scholar] [CrossRef]
- Van Langevelde, F.; van de Vijver, C.A.D.M.; Kumar, L.; van de Koppel, J.; de Ridder, N.; van Andel, J.; Skidmore, A.K.; Hearne, J.W.; Stroosnijder, L.; Bond, W.J.; et al. Effects of fire and herbivory on the stability of savanna ecosystems. Ecology 2003, 84, 337–350. [Google Scholar] [CrossRef]
- Werner, P.A.; Franklin, D.C. Resprouting and mortality of juvenile eucalypts in an Australian savanna: Impacts of fire season and annual sorghum. Austr. J. Bot. 2010, 58, 619–628. [Google Scholar] [CrossRef]
- Beckage, B.; Gross, L.J.; Platt, W.J. Grass feedbacks on fire stabilize savannas. Ecol. Modell. 2011, 222, 2227–2233. [Google Scholar] [CrossRef]
- Morrison, T.A.; Holdo, R.M.; Rugemalila, D.M.; Nzunda, M.; Anderson, T.M. Grass competition overwhelms effects of herbivores and precipitation on early tree establishment in Serengeti. J. Ecol. 2019, 107, 216–228. [Google Scholar] [CrossRef]
- Higgins, S.I.; Bond, W.J.; Trollope, W.S.W. Fire, resprouting and variability: a recipe for grass-tree coexistence in savanna. J. Ecol. 2000, 88, 213–229. [Google Scholar] [CrossRef] [Green Version]
- Wiegand, K.; Saitz, D.; Ward, D. A patch-dynamics approach to savanna dynamics and woody plant encroachment - insights from an arid savanna. Perspect. Plant Ecol. Evol. Syst. 2006, 7, 229–242. [Google Scholar] [CrossRef]
- Menaut, J.C.; Gignoux, J.; Prado, C.; Clobert, J. Tree community dynamics in a humid savanna of the Cote-d’Ivoire: Modelling the effects of fire and competition with grass and neighbours. J. Biogeogr. 1990, 17, 471–481. [Google Scholar] [CrossRef]
- Jeltsch, F.; Milton, S.J.; Dean, W.R.J.; van Rooyen, N. Tree spacing and coexistence in semiarid savannas. J. Ecol. 1996, 84, 583–595. [Google Scholar] [CrossRef]
- Russell-Smith, J.; Whitehead, P.J.; Cook, G.D.; Hoare, J.L. Response of Eucalyptus-dominated savanna to frequent fires: lessons from Munmarlary, 1973–1996. Ecol. Monogr. 2003, 73, 349–375. [Google Scholar] [CrossRef]
- Beckage, B.; Platt, W.J.; Gross, L.J. Vegetation, fire, and feedbacks: a disturbance-mediated model of savannas. Am. Nat. 2009, 174. [Google Scholar]
- Midgley, J.J.; Lawes, M.J.; Chamaille-Jammes, S. Savanna woody plant dynamics: the role of fire and herbivory, separately and synergistically. Austr. J. Bot. 2010, 58, 1–11. [Google Scholar] [CrossRef]
- Clarke, P.J.; Lawes, M.J.; Midgley, J.J.; Lamont, B.B.; Ojeda, F.; Burrows, G.E.; Enright, N.J.; Knox, K.J.E. Resprouting as a key functional trait: how buds, protection and resource drive persistence after fire. New Phytol. 2012, 197, 19–35. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.T.; Murphy, B.P.; Baker, P.J. The existence of a fire-mediated tree-recruitment bottleneck in an Asian savanna. J. Biogeogr. 2019, 46, 745–756. [Google Scholar] [CrossRef]
- Grubb, P.J. The maintenance of species-richness in plant communities: the importance of the regeneration niche. Biol. Rev. 1977, 52, 107–145. [Google Scholar] [CrossRef]
- Moustakas, A.; Wiegand, K.; Meyer, K.M.; Ward, D.; Sankaran, M. Learning new tricks from old tress: Revisiting the savanna question. Front. Biogeogr. 2010, 2, 47–53. [Google Scholar]
- Vetaas, O.R. Micro-site effects of trees and shrubs in dry savannas. J. Veg. Sci. 1992, 3, 337–344. [Google Scholar] [CrossRef]
- Hoffmann, W.A. The effects of fire and cover on seedling establishment in a neotropical savanna. J. Ecol. 1996, 84, 383–393. [Google Scholar] [CrossRef]
- McGuire, J.P.; Mitchell, R.J.; Moser, E.B.; Pecot, S.D.; Gjerstad, D.H.; Hedman, C.W. Gaps in a gappy forest: Plant resources, longleaf pine regeneration, and understory response to tree removal in longleaf pine savannas. Can. J. For. Res. 2001, 31, 765–778. [Google Scholar] [CrossRef]
- Jeltsch, F.; Weber, G.E.; Grimm, V. Ecological buffering mechanisms in savannas: A unifying theory of long-term tree-grass coexistence. Plant Ecol. 2000, 161, 161–171. [Google Scholar] [CrossRef]
- Mutch, R.W. Wildland fires and ecosystems--A hypothesis. Ecology 1970, 51, 1046–1051. [Google Scholar] [CrossRef]
- Bond, W.E.; Midgley, J.J. Kill thy neighbor: An individualistic argument for the evolution of flammability. Oikos 1995, 73, 79–85. [Google Scholar] [CrossRef]
- Scarff, F.R.; Westoby, M. Leaf litter flammability in some semi-arid Australian woodlands. Funct. Ecol. 2006, 20, 745–752. [Google Scholar] [CrossRef] [Green Version]
- Platt, W.J.; Evans, G.W.; Rathbun, S.L. The population dynamics of a long-lived conifer (Pinus palustris). Am. Nat. 1988, 131, 491–5225. [Google Scholar] [CrossRef]
- Grace, S.L.; Platt, W.J. Effects of adult tree density and fire on the demography of pregrass stage juvenile longleaf pine (Pinus palustris Mill.). J. Ecol. 1995, 83, 75–86. [Google Scholar] [CrossRef]
- O’Brien, J.J.; Loudermilk, E.L.; Hiers, J.K.; Pokswinski, S.; Hornsby, B.; Hudak, A.; Strother, D.; Rowell, E.; Bright, B.C. Canopy derived fuels drive patterns of in-fire energy release and understory plant mortality in a longleaf pine (Pinus palustris) sandhill in northwest Florida, USA. Can. J. Remote Sens. 2016, 42, 489–500. [Google Scholar] [CrossRef]
- Platt, W.J.; Ellair, D.P.; Huffman, J.M.; Potts, S.E.; Beckage, B. Pyrogenic fuels produced by savanna trees can engineer humid savannas. Ecol. Monogr. 2016, 86, 352–372. [Google Scholar] [CrossRef]
- Hoffmann, W.A.; Jaconis, S.Y.; McKinley, K.L.; Geiger, E.L.; Cotsch, S.G.; Franco, A.C. Fuels or microclimate? Understanding the drivers of fire feedbacks at savanna-forest boundaries. Austral Ecol. 2012, 37, 634–643. [Google Scholar] [CrossRef]
- Mugnani, M.P.; Robertson, K.M.; Miller, D.L.; Platt, W.J. Longleaf pine patch dynamics influence ground-layer vegetation in old-growth pine savanna. Forests 2019, in press. [Google Scholar]
- Gilliam, F.S.; Platt, W.J. Effects of long-term fire exclusion on tree species composition and stand structure in an old-growth Pinus palustris (Longleaf pine) forest. Plant Ecol. 1999, 140, 15–26. [Google Scholar] [CrossRef]
- Gignoux, J.; Lahoreau, G.; Julliard, R.; Barot, S. Establishment and early persistence of tree seedlings in an annually burned savanna. J. Ecology 2009, 97, 484–495. [Google Scholar] [CrossRef] [Green Version]
- Bond, W.J.; Parr, C.L. Beyond the forest edge: Ecology, diversity and conservation of the grassy biomes. Biol. Conserv. 2010. [Google Scholar] [CrossRef]
- Brockway, D.G.; Outcalt, K.W. Gap-phase regeneration in longleaf pine wiregrass ecosystems. For. Ecol. Manag. 1998, 106, 125–139. [Google Scholar] [CrossRef]
- Wahlenberg, W.G. Longleaf Pine: Its Use, Ecology, Regeneration, Protection, and Management; Charles Lathrop Pack Forestry Foundation; U.S. Department of Agriculture Forest Service: Washington, DC, USA, 1946; p. 429.
- Prior, L.D.; Williams, R.J.; Bowman, D.M.J.S. Experimental evidence that fire causes a tree recruitment bottleneck in an Australian tropical savanna. J. Trop. Ecol. 2010, 26, 595–603. [Google Scholar] [CrossRef] [Green Version]
- Bond, W.J. What limits trees in C4 grasslands and savannas? Ann. Rev. Ecol. Syst. 2008, 39, 641–659. [Google Scholar] [CrossRef]
- Archibald, S.; Lehmann, C.E.R.; Belcher, C.M.; Bond, W.J.; Bradstock, R.A.; Daniau, A.L.; Dexter, K.G.; Forresetel, E.J.; Greve, M.; He, T.; et al. Biological and geophysical feedbacks with fire in the Earth system. Environ. Res. Lett. 2018, 13, 033003. [Google Scholar] [CrossRef] [Green Version]
- Glitzenstein, J.S.; Streng, D.R.; Wade, D.D. Fire frequency effects on longleaf pine (Pinus palustris P. Miller) vegetation in South Carolina and Northeast Florida, USA. Research Article. Nat. Areas J. 2003, 23, 22–37. [Google Scholar]
- Cox, J.A.; Jones, C.D. Influence of prescribed fire on winter abundance of Bachman’s sparrow. Wilson J. Ornithol. 2009, 121, 359–365. [Google Scholar] [CrossRef]
- Robertson, K.M.; Hmielowski, T.L. Effects of fire frequency and season on resprouting of woody plants in southeastern US pine-grassland communities. Oecologia 2014, 174, 765–776. [Google Scholar] [CrossRef]
- Huffman, J.M. Historical Fire Regimes In Southeastern Pine Savannas. Ph.D. Thesis, Louisiana State University and Agricultural and Mechanical College, Baton Rouge, LA, USA, 2006. [Google Scholar]
- Bale, A.M. Fire effects and litter accumulation dynamics in a montane longleaf pine system. Master’s Thesis, University of Missouri-Columbia, Columbia, MO, USA, May 2009. [Google Scholar]
- Stambaugh, M.C.; Guyette, R.P.; Marschall, J.M. Longleaf pine (Pinus palustris Mill.) fire scars reveal new details of a frequent fire regime. J. veg. Sci. 2011, 22, 1094–1104. [Google Scholar] [CrossRef]
- Freeman, M.E.; Murphy, B.P.; Richards, A.E.; Vesk, P.A.; Cook, G.D. Facultative and obligate trees in a mesic savanna: Fire effects on savanna structure imply contrasting strategies of eco-taxonomic groups. Front. Plant Sci. 2018, 9, 644. [Google Scholar] [CrossRef] [PubMed]
- Rebertus, A.J.; Williamson, G.B.; Moser, E.B. Longleaf pine pyrogenicity and turkey oak mortality in Florida xeric sandhills. Ecology 1989, 70, 60–70. [Google Scholar] [CrossRef]
- Robertson, K.M.; Ostertag, T.E. Effects of land use on fuel characteristics and fire behavior in pinelands of Southwest Georgia. Tall Timbers Fire Ecol. Conf. Proc. 2007, 23, 181–191. [Google Scholar]
- Williamson, G.B.; Black, E.M. High temperature of forest fires under pines as a selective advantage over oaks. Nature 1981, 293, 643–644. [Google Scholar] [CrossRef]
- Glitzenstein, J.S.; Platt, W.J.; Streng, D.R. Effects of fire regime and habitat on tree dynamics in North Florida longleaf pine savannas. Ecol. Monogr. 1995, 65, 141–476. [Google Scholar] [CrossRef]
- Gagnon, J.L.; Jokela, E.J.; Moser, W.K.; Huber, D.A. Characteristics of gaps and natural regeneration in mature longleaf pine flatwoods ecosystems. For. Ecol. Manag. 2004, 187, 373–380. [Google Scholar] [CrossRef]
- Hammond, D.H.; Varner, J.M.; Fan, Z.; Kush, J.S. Long-term stand dynamics of old-growth mountain longleaf pine (Pinus palustris) woodlands. For. Ecol. Manag. 2016, 364, 154–164. [Google Scholar] [CrossRef]
- Noel, J.M.; Platt, W.J.; Moser, E.B. Structural characteristics of old- and second-growth stands of longleaf pine (Pinus palustris) in the Gulf Coastal Region of the U.S.A. Conserv. Biol. 1998, 12, 533–548. [Google Scholar] [CrossRef]
- Platt, W.J.; Rathbun, S.L. Dynamics of an old-growth longleaf pine population. Tall Timbers Fire Ecol. Conf. Proc. 1993, 18, 275–297. [Google Scholar]
- Brockway, D.G.; Outcalt, K.W.; Boyer, W.D. Longleaf pine regeneration ecology and methods. In Springer Series on Environmental Management; Jose, S., Jokela, J., Miller, D.L., Eds.; Springer: New York, NY, USA, 2006; pp. 95–133. [Google Scholar]
- Chen, X.W.; Brockway, D.G.; Guo, Q.F. Characterizing the dynamics of cone production for longleaf pine forests in the southeastern United States. For. Ecol. Manag. 2018, 429, 1–6. [Google Scholar] [CrossRef]
- Reid, A.M.; Robertson, K.M. Energy content of common fuels in upland pine savannas of the south-eastern US and their application to fire behaviour modelling. Int. J. Wildland Fire 2012, 21, 591–595. [Google Scholar] [CrossRef]
- Chapman, H.H. Is the longleaf type a climax? Ecology 1932, 13, 328–334. [Google Scholar] [CrossRef]
- Pessin, L.J. Stimulating the early height growth of longleaf pine seedlings. J. For. 1944, 42, 95–98. [Google Scholar]
- Smith, L.F. Development of longleaf pine seedlings near large trees. J. For. 1955, 53, 289–290. [Google Scholar]
- Palik, B.J.; Mitchell, R.J.; Houseal, G.; Pederson, N. Effects of canopy structure on resource availability and seedling responses in a longleaf pine ecosystem. Can. J. For. Res. 1997, 27, 1458–1464. [Google Scholar] [CrossRef]
- Grace, S.L.; Platt, W.J. Neighborhood effects on juveniles in an old-growth stand of longleaf pine, Pinus palustris. Oikos 1995, 72, 99–105. [Google Scholar] [CrossRef]
- Ellair, D.P.; Platt, W.J. Fuel composition influences fire characteristics and understorey hardwoods in pine savanna. J. Ecol. 2013, 101, 192–201. [Google Scholar] [CrossRef]
- Lawton, D.E.; Marsalis, W.E.; Moye, F.J.; Murray, J.B.; O’Connor, B.J.; Penley, H.M.; Sandrock, G.S.; Friddell, M.S.; Hetrick, J.H.; Huddlestun, P.F.; et al. Geologic Map of Georgia; Georgia Department of Natural Resources and Georgia Geological Survey: Atlanta, GA, USA, 1976.
- Sanders, T.E. Soil survey of Leon County, Florida; U.S. Department of Agriculture Soil Conservation Service; University of Florida: Tallahassee, FL, USA, 1981; p. 151.
- Staff, S.S. Web Soil Survey. Available online: https://websoilsurvey.nrcs.usda.gov/ (accessed on 15 January 2019).
- Gonzalez-Benecke, C.A.; Gezan, S.A.; Samuelson, L.J.; Cropper, W.P.J.; Leduc, D.J.; Martin, T.A. Estimating Pinus palustris tree diameter and stem volume from tree height, crown area and stand-level parameters. J. For. Res. 2014, 25, 43–52. [Google Scholar] [CrossRef]
- Gagnon, J.L.; Jokela, E.J.; Moser, W.K.; Huber, D.A. Dynamics of artificial regeneration in gaps within a longleaf pine flatwoods ecosystem. For. Ecol. Manag. 2003, 172, 133–144. [Google Scholar] [CrossRef]
- O’Brien, J.J.; Hiers, J.K.; Callaham, M.A.J.; Mitchell, R.J.; Jack, S.B. Interactions among overstory structure, seedling life-history traits, and fire in frequently burned neotropical pine forests. Ambio 2008, 37, 542–547. [Google Scholar] [CrossRef] [PubMed]
- Koenig, W.D. Spatial autocorrelation of ecological phenomena. Trends Ecol. Evol. 1999, 14, 22–26. [Google Scholar] [CrossRef]
- Grace, S.L.; Hamrick, J.L.; Platt, W.J. Estimation of seed dispersal in an old-growth population of longleaf pine (Pinus Palustris) using maternity exclusion analysis. Castanea 2004, 69, 207–215. [Google Scholar] [CrossRef]
- Pessin, L.J. Density of stocking and character of ground cover as factors in longleaf pine reproduction. J. For. 1939, 37, 255–258. [Google Scholar]
- Boyer, W.D. Development of Longleaf Pine Seedlings Under Parent Trees. U.S. Forest Research Paper SO-4; Southern Forest Experiment Station, Forest Service, U.S. Department of Agriculture: Asheville, NC, USA, 1963.
- Harrington, T.B.; Dagley, C.M.; Edwards, M.B. Above- and belowground competition from longleaf pine plantations limits performance of reintroduced herbaceous species. For. Sci. 2003, 49, 681–695. [Google Scholar]
- Pessin, L.J.; Chapman, R.A. The effect of living grass on the growth of longleaf pine seedlings in pots. Ecology 1944, 25, 85–90. [Google Scholar] [CrossRef]
- Loudermilk, E.L.; O’Brien, J.J.; Mitchell, R.J.; Cropper, W.P.; Hiers, J.K.; Grunwald, S.; Grego, J.; Fernandez-Diaz, J.C. Linking complex forest fuel structure and fire behaviour at fine scales. Int. J. Wildland Fire 2012, 21, 882–893. [Google Scholar] [CrossRef]
- Dell, J.E.; Richards, L.A.; O’Brien, J.J.; Loudermilk, E.L.; Hudak, A.T.; Pokswinski, S.M.; Bright, B.C.; Hiers, J.K.; Williams, B.W.; Dyer, L.A. Overstory-derived fuels mediate plant species diversity in frequently burned longleaf forests. Ecosphere 2017, 8, e01964. [Google Scholar] [CrossRef]
- Hermann, S.M. Small-scale disturbances in longleaf pine forests. Tall Timbers Fire Ecol. Conf. Proc. 1993, 18, 265–274. [Google Scholar]
- Beckage, B.; Gross, L.J.; Platt, W.J. Modelling responses of pine savannas to climate change and large-scale disturbance. Appl. Veg. Sci. 2006, 9, 75–82. [Google Scholar] [CrossRef]
- Moss, P.T.; Greenwood, D.R.; Archibald, S.B. Regional and local vegetation community dynamics of the Eocene Okanagan Highlands (British Columbia – Washington State) from palynology. Can. J. Earth Sci. 2005, 42, 187–204. [Google Scholar] [CrossRef]
- Boyer, W.D. Impact of Prescribed Fires on Mortality of Released and Unreleased Longleaf Pine Seedlings Under Parent Trees; U.S. Forest Service Southern Forest Experiment Station Research Note SO-182; U.S. Department of Agriculture: New Orleans, LA, USA, 1974; p. 24.
- Ford, C.R.; Minor, E.S.; Fox, G.A. Long-term effects of fire and fire-return interval on population structure and growth of longleaf pine (Pinus palustris). Can. J. For. 2010, 40, 1410–1420. [Google Scholar] [CrossRef]
- Touboul, J.D.; Staver, A.C.; Levin, S.A. On the complex dynamics of savanna landscapes. Proc. Natl. Acad. Sci. USA 2018, 115, E1336–E1345. [Google Scholar] [CrossRef] [PubMed]
- Cox, J.; Jones, C. Bachman’s Sparrow the Order of the Pheonix. Birding 2008, May/June, 38–45. [Google Scholar]
- Brudvig, L.A.; Grman, E.; Habeck, C.W.; Orrock, J.L.; Ledvina, J.A. Strong legacy of agricultural land use on soils and undestory plant communities in longleaf pine woodlands. For. Ecol. Manag. 2013, 310, 944–955. [Google Scholar] [CrossRef]
- Martin, R.E.; Sapsis, D.B. Fires as agents of biodiversity: Pyrodiversity promotes biodiversity. In Proceedings on the Symposium of Biodiversity in Northwestern California, 1991 pgs 150–151; Kerner, H.M., Ed.; Wildland Resources Center, University of California, Berkeley: Berkeley, CA, USA, 1992; pp. 150–157. [Google Scholar]
- Brockett, B.C.; Biggs, H.C.; van Wilgen, V.W. A patch mosaic burning system for conservation areas in southern African savannas. Int. J. Wildland Fire 2001, 10, 169–183. [Google Scholar] [CrossRef]
- Parr, C.L.; Anderson, A.N. Patch mosaic burning for biodiversity conservation: A critique of the pyrodiversity paradigm. Conserv. Biol. 2006, 20, 1610–1619. [Google Scholar] [CrossRef] [PubMed]
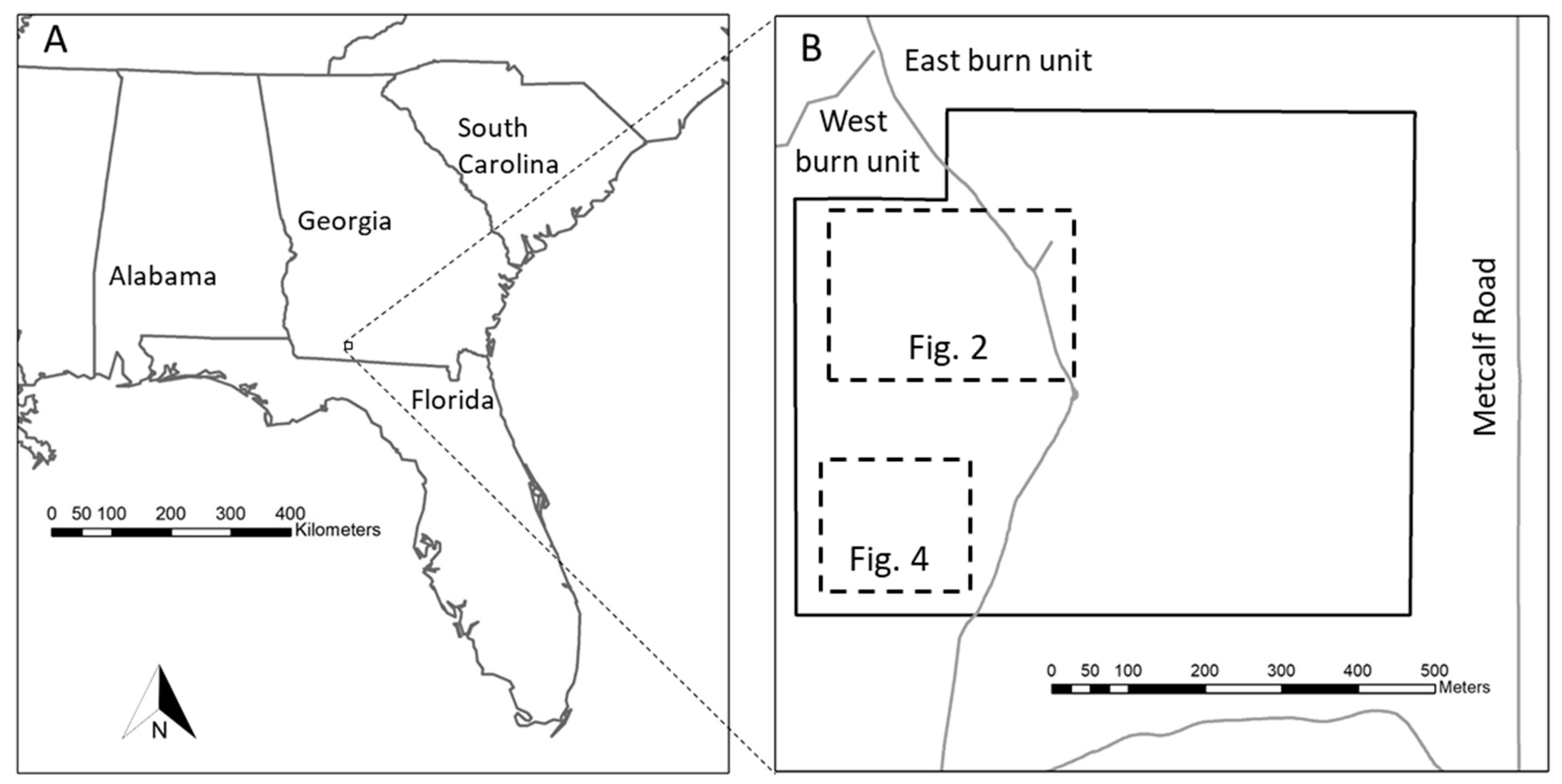
Figure 1.
(A) Location of the study area within the southeastern U.S. (B) Study area (bold solid line) and locations of areas shown in Figure 2 and Figure 4 (dashed lines). Roads are shown as grey lines.
Figure 1.
(A) Location of the study area within the southeastern U.S. (B) Study area (bold solid line) and locations of areas shown in Figure 2 and Figure 4 (dashed lines). Roads are shown as grey lines.

Figure 2.
Portion of the longleaf pine juvenile census area (see Figure 1) showing tendency for juveniles to be located in gaps within patches that were unburned during one or more of 11 fires from 2005–2018 (in contrast to areas unburned in zero fires, i.e., burned in each fire) and within areas re-opened by the death of canopy pine trees.
Figure 2.
Portion of the longleaf pine juvenile census area (see Figure 1) showing tendency for juveniles to be located in gaps within patches that were unburned during one or more of 11 fires from 2005–2018 (in contrast to areas unburned in zero fires, i.e., burned in each fire) and within areas re-opened by the death of canopy pine trees.

Figure 3.
Percentage of total study area and percentage of longleaf pine juveniles in each category of number of prescribed fires in which the area was unburned out of 11 in total from 2005 to 2018.
Figure 3.
Percentage of total study area and percentage of longleaf pine juveniles in each category of number of prescribed fires in which the area was unburned out of 11 in total from 2005 to 2018.

Figure 4.
A portion of the longleaf pine juvenile census (see Figure 1) showing locations of longleaf pine juveniles with respect to the number of times areas were unburned during 11 fires from 2005–2018.
Figure 4.
A portion of the longleaf pine juvenile census (see Figure 1) showing locations of longleaf pine juveniles with respect to the number of times areas were unburned during 11 fires from 2005–2018.

Table 1.
Results from logistic regression analysis testing the effects of Gap, Inner gap, and Re-opened area on areas unburned in ≥1 prescribed burn from 2005–2018. Analyses were based on 500 random points within and outside of unburned areas. S.E. = standard error of the estimate. Additional metrics for analyses are provided in Appendix A Table A1.
Table 1.
Results from logistic regression analysis testing the effects of Gap, Inner gap, and Re-opened area on areas unburned in ≥1 prescribed burn from 2005–2018. Analyses were based on 500 random points within and outside of unburned areas. S.E. = standard error of the estimate. Additional metrics for analyses are provided in Appendix A Table A1.
| Parameter | Estimate | S.E. | Z Score | p | Odds Ratio |
|---|---|---|---|---|---|
| Constant | −0.779 | 0.121 | −6.453 | <0.001 | |
| Gap | 1.132 | 0.144 | 7.881 | <0.001 | 3.103 |
| Constant | −0.260 | 0.071 | −3.638 | <0.001 | |
| Inner gap | 1.399 | 0.179 | 7.812 | <0.001 | 4.049 |
| Constant | −0.061 | 0.070 | −0.870 | 0.384 | |
| Re-opened area | 0.362 | 0.168 | 2.148 | 0.032 | 1.436 |
Table 2.
Percentage of surface categories in row titles that overlap with surface categories in column titles within the study area, excluding wetlands. UB = unburned.
Table 2.
Percentage of surface categories in row titles that overlap with surface categories in column titles within the study area, excluding wetlands. UB = unburned.
| UB ≥ 1 Fire | Gap | Inner Gap | Re-Opened | |
|---|---|---|---|---|
| UB ≥ 1 fire | 100 | 81.6 | 28.9 | 17.0 |
| Gap | 26.9 | 100 | 19.7 | 17.5 |
| Inner gap | 48.1 | 100 | 100 | 22.5 |
| Re-opened | 32.2 | 100 | 20.1 | 100 |
Table 3.
Results from logistic regression analysis with presence or absence of longleaf pine juvenile patches as the binary response variable and unburned area in ≥1 prescribed burn from 2005–2018, gap, inner gap, and re-opened areas as the binary independent variables. Analyses are based on 132 locations within and outside of longleaf pine juvenile patches.
Table 3.
Results from logistic regression analysis with presence or absence of longleaf pine juvenile patches as the binary response variable and unburned area in ≥1 prescribed burn from 2005–2018, gap, inner gap, and re-opened areas as the binary independent variables. Analyses are based on 132 locations within and outside of longleaf pine juvenile patches.
| Parameter | Estimate | S.E. | Z Score | p | Odds Ratio | |
|---|---|---|---|---|---|---|
| Constant | −1.562 | 0.319 | −4.892 | <0.001 | ||
| UB area | 0.640 | 0.292 | 2.192 | 0.028 | 1.897 | |
| Gap | 1.270 | 0.332 | 3.825 | <0.001 | 3.561 | |
| Inner gap | 0.718 | 0.357 | 2.012 | 0.044 | 2.051 | |
| Re-opened area | 0.936 | 0.329 | 2.846 | 0.004 | 2.550 | |
| Overall metrics | ||||||
| Chi-square | 46.7, 4 d.f., p <0.0001 | Juveniles present % correct | 63.6 | |||
| Cox & Snell R-sq | 0.162 | Juveniles absent % correct | 70.5 | |||
| Nagelkerke’s R-sq | 0.216 | Total % correct | 67.0 | |||
| Hosmer-Lemeshow | 3.535, p = 0.739, 6 d.f. | Log likelihood constant model | −182.9 | |||
| ROC Area Under Curve | 0.730 | Log likelihood full model | −159.6 | |||
Table 4.
Descriptive statistics for longleaf pine juveniles in relation to selected land cover categories, including number of fires out of 11 in which areas were unburned (# UB), total hectares covered by each cover category, and their percent of the total area. Gaps include inner gaps and re-opened areas. Juveniles within each land cover category are described by their total number, percentage of the total number, overall density (individuals per hectare), and percentage of juveniles occurring in clusters as defined in the text. Areas excluded wetlands in which longleaf pine juveniles did not occur.
Table 4.
Descriptive statistics for longleaf pine juveniles in relation to selected land cover categories, including number of fires out of 11 in which areas were unburned (# UB), total hectares covered by each cover category, and their percent of the total area. Gaps include inner gaps and re-opened areas. Juveniles within each land cover category are described by their total number, percentage of the total number, overall density (individuals per hectare), and percentage of juveniles occurring in clusters as defined in the text. Areas excluded wetlands in which longleaf pine juveniles did not occur.
| Cover Category | # UB | ha | % Area | Juveniles | % Total | Density | % in Patches |
|---|---|---|---|---|---|---|---|
| All | 0–11 | 49.4 | 100.0 | 2565 | 100.0 | 51.9 | 80.7 |
| ≥1 | 9.6 | 19.4 | 1553 | 58.7 | 161.8 | 92.5 | |
| 0 | 39.8 | 80.6 | 1012 | 41.3 | 25.4 | 63.1 | |
| Canopy | ≥1 | 2.3 | 4.7 | 143 | 5.6 | 62.2 | 79.7 |
| 0 | 17.3 | 35.1 | 190 | 7.4 | 11.0 | 51.1 | |
| Gap | ≥1 | 7.3 | 14.8 | 1410 | 55.0 | 193.2 | 90.6 |
| 0 | 22.5 | 45.4 | 822 | 32.0 | 36.5 | 69.5 | |
| Inner gap (IG) | ≥1 | 2.5 | 5.1 | 638 | 25.7 | 255.2 | 92.9 |
| 0 | 3.1 | 6.4 | 138 | 5.4 | 44.5 | 71.7 | |
| Re-opened (RO) | ≥1 | 2.1 | 4.1 | 660 | 25.7 | 314.3 | 93.8 |
| 0 | 5.6 | 11.3 | 311 | 12.1 | 55.5 | 82.6 | |
| IG/RO | ≥1 | 0.6 | 1.2 | 268 | 10.4 | 446.7 | 96.6 |
| 0 | 0.8 | 1.6 | 57 | 2.2 | 71.3 | 84.2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Robertson, K.M.; Platt, W.J.; Faires, C.E. Patchy Fires Promote Regeneration of Longleaf Pine (Pinus palustris Mill.) in Pine Savannas. Forests 2019, 10, 367. https://doi.org/10.3390/f10050367
AMA Style
Robertson KM, Platt WJ, Faires CE. Patchy Fires Promote Regeneration of Longleaf Pine (Pinus palustris Mill.) in Pine Savannas. Forests. 2019; 10(5):367. https://doi.org/10.3390/f10050367
Chicago/Turabian StyleRobertson, Kevin M., William J. Platt, and Charles E. Faires. 2019. "Patchy Fires Promote Regeneration of Longleaf Pine (Pinus palustris Mill.) in Pine Savannas" Forests 10, no. 5: 367. https://doi.org/10.3390/f10050367
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.