
It’s Time for Entropic Clocks: The Roles of Random Chain Protein Sequences in Timing Ion Channel Processes Underlying Action Potential Properties

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Thermodynamic and Timing Signatures of Entropic Clock-Based ‘Ball and Chain’ Binding Mechanism
3. The Physiological Relevance of Entropic Clocks in Kv Channel Inactivation and Clustering
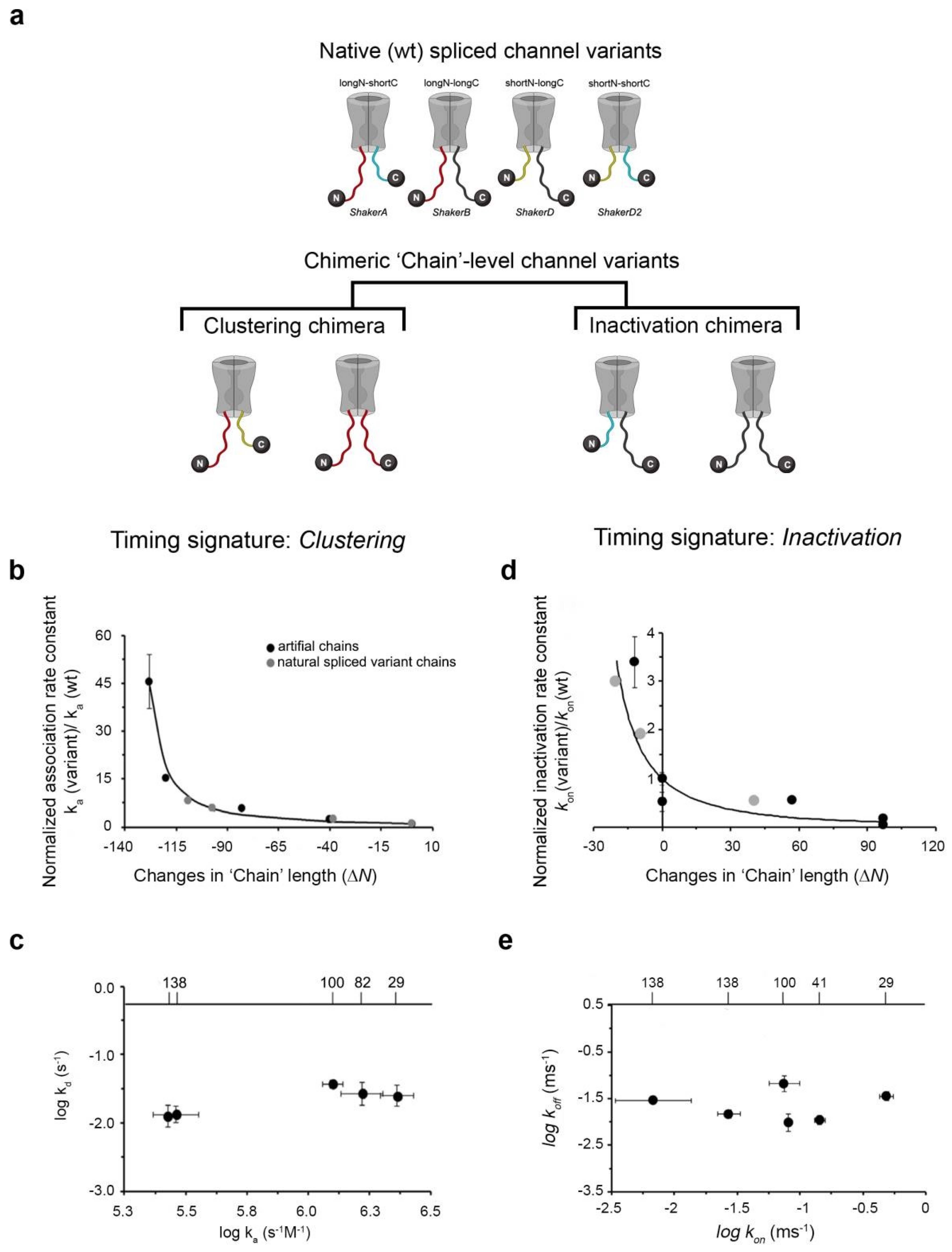
4. A Native-‘Chain’ Chimeric Channel Strategy Supports Entropic Clock-Based ‘Ball and Chain’ Mechanisms for Kv Channel Clustering and Inactivation
5. Cellular Clustering Correlates of the PSD-95-Kv Channel ‘Ball and Chain’ Binding Mechanism
6. Subunit Hereto-Oligomerization Regulates Cluster Kv Channel Density
7. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Kandel, E.R.; Schwartz, J.H.; Jessell, T.M.; Siegelbaum, S.A.; Hudspeth, A.J. Principles of Neural Science, 4th ed.; McGraw-Hill: New York, NY, USA, 2000. [Google Scholar]
- Lai, H.C.; Jan, L.Y. The Distribution and Targeting of Neuronal Voltage-Gated Ion Channels. Nat. Rev. Neurosci. 2006, 7, 548–562. [Google Scholar] [CrossRef] [PubMed]
- Trimmer, J.S. Subcellular Localization of K+ Channels in Mammalian Brain Neurons: Remarkable Precision in the Midst of Extraordinary Complexity. Neuron 2015, 85, 238–256. [Google Scholar] [CrossRef]
- Debanne, D.; Guérineau, N.C.; Gähwiler, B.H.; Thompson, S.M. Action-Potential Propagation Gated by an Axonal IA-like K+ Conductance in Hippocampus. Nature 1997, 389, 286–289. [Google Scholar] [CrossRef]
- Giese, K.P.; Storm, J.F.; Reuter, D.; Fedorov, N.B.; Shao, L.-R.; Leicher, T.; Pongs, O.; Silva, A.J. Reduced K+ Channel Inactivation, Spike Broadening, and after- Hyperpolarization in KvB1.1-Deficient Mice with Impaired Learning. Learn. Mem. 1998, 5, 257–273. [Google Scholar] [CrossRef] [PubMed]
- Johnston, D.; Hoffman, D.A.; Colbert, C.M.; Magee, J.C. Regulation of Back-Propagating Action Potentials in Hippocampal Neurons. Curr. Opin. Neurobiol. 1999, 9, 288–292. [Google Scholar] [CrossRef]
- Århem, P.; Klement, G.; Blomberg, C. Channel Density Regulation of Firing Patterns in a Cortical Neuron Model. Biophys. J. 2006, 90, 4392–4404. [Google Scholar] [CrossRef]
- Zeberg, H.; Blomberg, C.; Århem, P. Ion Channel Density Regulates Switches between Regular and Fast Spiking in Soma but Not in Axons. PLoS Comput. Biol. 2010, 6, e1000753. [Google Scholar] [CrossRef]
- Zeberg, H.; Robinson, H.P.C.; Århem, P. Density of Voltage-Gated Potassium Channels Is a Bifurcation Parameter in Pyramidal Neurons. J. Neurophysiol. 2015, 113, 537–549. [Google Scholar] [CrossRef] [PubMed]
- Zandany, N.; Lewin, L.; Nirenberg, V.; Orr, I.; Yifrach, O. Entropic Clocks in the Service of Electrical Signaling: “Ball and Chain” Mechanisms for Ion Channel Inactivation and Clustering. FEBS Lett. 2015, 589, 2441–2447. [Google Scholar] [CrossRef]
- Dunker, A.K.; Lawson, J.D.; Brown, C.J.; Williams, R.M.; Romero, P.; Oh, J.S.; Oldfield, C.J.; Campen, A.M.; Ratliff, C.M.; Hipps, K.W.; et al. Intrinsically Disordered Protein. J. Mol. Graph. Model. 2001, 19, 26–59. [Google Scholar] [CrossRef]
- Timpe, L.C.; Peller, L. A Random Flight Chain Model for the Tether of the Shaker K+ Channel Inactivation Domain. Biophys. J. 1995, 69, 2415–2418. [Google Scholar] [CrossRef] [PubMed]
- Zandany, N.; Marciano, S.; Magidovich, E.; Frimerman, T.; Yehezkel, R.; Shem-Ad, T.; Lewin, L.; Abdu, U.; Orr, I.; Yifrach, O. Alternative Splicing Modulates Kv Channel Clustering through a Molecular Ball and Chain Mechanism. Nat. Commun. 2015, 6, 6488. [Google Scholar] [CrossRef] [PubMed]
- Gomperts, S.N. Clustering Membrane Proteins: It’s All Coming Together with the PSD- 95/SAP90 Protein Family. Cell 1996, 84, 659–662. [Google Scholar] [CrossRef] [PubMed]
- Hsueh, Y.P.; Kim, E.; Sheng, M. Disulfide-Linked Head-to-Head Multimerization in the Mechanism of Ion Channel Clustering by PSD-95. Neuron 1997, 18, 803–814. [Google Scholar] [CrossRef]
- Armstrong, C.M.; Bezanilla, F. Inactivation of the Sodium Channel. II. Gating Current Experiments. J. Gen. Physiol. 1977, 70, 567–590. [Google Scholar] [CrossRef]
- Zagotta, W.N.; Hoshi, T.; Aldrich, R.W. Restoration of Inactivation in Mutants of Shaker Potassium Channels by a Peptide Derived from ShB. Science 1990, 250, 568–571. [Google Scholar] [CrossRef] [PubMed]
- Hoshi, T.; Zagotta, W.N.; Aldrich, R.W. Biophysical and Molecular Mechanisms of Shaker Potassium Channel Inactivation. Science 1990, 250, 533–538. [Google Scholar] [CrossRef] [PubMed]
- Rasmusson, R.L.; Morales, M.J.; Wang, S.; Liu, S.; Campbell, D.L.; Brahmajothi, M.V.; Strauss, H.C. Inactivation of Voltage-Gated Cardiac K+ Channels. Circ. Res. 1998, 82, 739–750. [Google Scholar] [CrossRef]
- Hille, B. Ion Channels of Excitable Membranes, 3rd ed.; Sinauer Associates: Sunderland, MA, USA, 2001. [Google Scholar]
- Aldrich, R.W. Fifty Years of Inactivation. Nature 2001, 411, 643–644. [Google Scholar] [CrossRef]
- Demo, S.D.; Yellen, G. The Inactivation Gate of the Shaker K+ Channel Behaves like an Open-Channel Blocker. Neuron 1991, 7, 743–753. [Google Scholar] [CrossRef]
- Murrell-Lagnado, R.D.; Aldrich, R.W. Interactions of Amino Terminal Domains of Shaker K Channels with a Pore Blocking Site Studied with Synthetic Peptides. J. Gen. Physiol. 1993, 102, 949–975. [Google Scholar] [CrossRef]
- Zhou, M.; Morais-Cabral, J.H.; Mann, S.; MacKinnon, R. Potassium Channel Receptor Site for the Inactivation Gate and Quaternary Amine Inhibitors. Nature 2001, 411, 657–661. [Google Scholar] [CrossRef]
- Gonzalez, C.; Lopez-Rodriguez, A.; Srikumar, D.; Rosenthal, J.J.C.; Holmgren, M. Editing of Human K(V)1.1 Channel MRNAs Disrupts Binding of the N-Terminus Tip at the Intracellular Cavity. Nat. Commun. 2011, 2, 436. [Google Scholar] [CrossRef]
- Tiffany, A.M.; Manganas, L.N.; Kim, E.; Hsueh, Y.P.; Sheng, M.; Trimmer, J.S. Psd-95 and Sap97 Exhibit Distinct Mechanisms for Regulating K+ Channel Surface Expression and Clustering. J. Cell. Biol. 2000, 148, 147–158. [Google Scholar] [CrossRef]
- Kim, E.; Niethammer, M.; Rothschild, A.; Jan, Y.N.; Sheng, M. Clustering of Shaker-Type K+ Channels by Interaction with a Family of Membrane-Associated Guanylate Kinases. Nature 1995, 378, 85–88. [Google Scholar] [CrossRef]
- Tejedor, F.J.; Bokhari, A.; Rogero, O.; Gorczyca, M.; Zhang, J.; Kim, E.; Sheng, M.; Budnik, V. Essential Role for Dlg in Synaptic Clustering of Shaker K+ Channels In Vivo. J. Neurosci. 1997, 17, 152–159. [Google Scholar] [CrossRef]
- Ruiz-Cañada, C.; Koh, Y.H.; Budnik, V.; Tejedor, F.J. DLG Differentially Localizes Shaker K+-Channels in the Central Nervous System and Retina of Drosophila. J. Neurochem. 2002, 82, 1490–1501. [Google Scholar] [CrossRef]
- Zhong, Y.; Budnik, V.; Wu, C.F. Synaptic Plasticity in Drosophila Memory and Hyperexcitable Mutants: Role of CAMP Cascade. J. Neurosci. 1992, 12, 644–651. [Google Scholar] [CrossRef]
- Budnik, V.; Zhong, Y.; Wu, C.F. Morphological Plasticity of Motor Axons in Drosophila Mutants with Altered Excitability. J. Neurosci. 1990, 10, 3754–3768. [Google Scholar] [CrossRef]
- Cooper, A. Thermodynamic Analysis of Biomolecular Interactions. Curr. Opin. Chem. Biol. 1999, 3, 557–563. [Google Scholar] [CrossRef]
- Dunker, A.K.; Brown, C.; Lawson, J.D.; Iakoucheva, L.M.; Obradović, Z. Intrinsic Disorder and Protein Function. Biochemistry 2002, 41, 6573–6582. [Google Scholar] [CrossRef]
- Lewin, L.; Nirenberg, V.; Yehezkel, R.; Naim, S.; Abdu, U.; Orr, I.; Yifrach, O. Direct Evidence for a Similar Molecular Mechanism Underlying Shaker Kv Channel Fast Inactivation and Clustering. J. Mol. Biol. 2019, 431, 542–556. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, T.L.; Tempel, B.L.; Papazian, D.M.; Jan, Y.N.; Jan, L.Y. Multiple Potassium–Channel Components Are Produced by Alternative Splicing at the Shaker Locus in Drosophila. Nature 1988, 331, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Pongs, O.; Kecskemethy, N.; Müller, R.; Krah-Jentgens, I.; Baumann, A.; Kiltz, H.H.; Canal, I.; Llamazares, S.; Ferrus, A. Shaker Encodes a Family of Putative Potassium Channel Proteins in the Nervous System of Drosophila. EMBO J. 1988, 7, 1087–1096. [Google Scholar] [CrossRef] [PubMed]
- Mottes, J.-R.; Iverson, L.-E. Tissue-Specific Alternative Splicing of Hybrid Shaker/LacZ Genes Correlates with Kinetic Differences in Shaker K+ Currents in Vivo. Neuron 1995, 14, 613–623. [Google Scholar] [CrossRef]
- Rogero, O.; Hämmerle, B.; Tejedor, F.J. Diverse Expression and Distribution of Shaker Potassium Channels during the Development of the Drosophila Nervous System. J. Neurosci. 1997, 17, 5108–5118. [Google Scholar] [CrossRef]
- Timpe, L.C.; Jan, Y.N.; Jan, L.Y. Four CDNA Clones from the Shaker Locus of Drosophila Induce Kinetically Distinct A-Type Potassium Currents in Xenopus Oocytes. Neuron 1988, 1, 659–667. [Google Scholar] [CrossRef]
- Iverson, L.E.; Tanouye, M.A.; Lester, H.A.; Davidson, N.; Rudy, B. A-Type Potassium Channels Expressed from Shaker Locus CDNA. Proc. Natl. Acad. Sci. USA 1988, 85, 5723–5727. [Google Scholar] [CrossRef] [PubMed]
- Pfeiffer, P.; Egorov, A.V.; Lorenz, F.; Schleimer, J.H.; Draguhn, A.; Schreiber, S. Clusters of Cooperative Ion Channels Enable a Membrane-Potential-Based Mechanism for Short- Term Memory. Elife 2020, 9, e49974. [Google Scholar] [CrossRef] [PubMed]
- Romero, P.R.; Zaidi, S.; Fang, Y.Y.; Uversky, V.N.; Radivojac, P.; Oldfield, C.J.; Cortese, M.S.; Sickmeier, M.; LeGall, T.; Obradovic, Z.; et al. Alternative Splicing in Concert with Protein Intrinsic Disorder Enables Increased Functional Diversity in Multicellular Organisms. Proc. Natl. Acad. Sci. USA 2006, 103, 8390–8395. [Google Scholar] [CrossRef]
- Buljan, M.; Chalancon, G.; Dunker, A.K.; Bateman, A.; Balaji, S.; Fuxreiter, M.; Babu, M.M. Alternative Splicing of Intrinsically Disordered Regions and Rewiring of Protein Interactions. Curr. Opin. Struct. Biol. 2013, 23, 443–450. [Google Scholar] [CrossRef] [PubMed]
- Topinka, J.R.; Bredt, D.S. N-Terminal Palmitoylation of PSD-95 Regulates Association with Cell Membranes and Interaction with K+ Channel Kv1.4. Neuron 1998, 20, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Rettig, J.; Heinemann, S.H.; Wunder, F.; Lorra, C.; Parcej, D.N.; Oliver Dolly, J.; Pongs, O. Inactivation Properties of Voltage-Gated K+ Channels Altered by Presence of Beta-Subunit. Nature 1994, 369, 289–294. [Google Scholar] [CrossRef]
- Lewin, L.; Nsasra, E.; Golbary, E.; Hadad, U.; Orr, I.; Yifrach, O. Molecular and Cellular Correlates in Kv Channel Clustering: Entropy-Based Regulation of Cluster Ion Channel Density. Sci. Rep. 2020, 10, e11304. [Google Scholar] [CrossRef] [PubMed]
- McCormack, K.; Lin, J.W.; Iverson, L.E.; Rudy, B. Shaker K+ Channel Subunits Form Heteromultimeric Channels with Novel Functional Properties. Biochem. Biophys. Res. Commun. 1990, 171, 1361–1371. [Google Scholar] [CrossRef] [PubMed]
- Nsasra, E.; Peretz, G.; Orr, I.; Yifrach, O. Regulating Shaker Kv Channel Clustering by Hetero-Oligomerization. Front. Mol. Biosci. 2023, 9, 1050942. [Google Scholar] [CrossRef]
- Uversky, V.N.; Gillespie, J.R.; Fink, A.L. Why Are “Natively Unfolded” Proteins Unstructured under Physiologic Conditions? Proteins 2000, 41, 415–427. [Google Scholar] [CrossRef]
- Tompa, P. Intrinsically Unstructured Proteins. Trends Biochem. Sci. 2002, 27, 527–533. [Google Scholar] [CrossRef] [PubMed]
- Wright, P.E.; Dyson, H.J. Linking Folding and Binding. Curr. Opin. Struct. Biol. 2009, 19, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Gsponer, J.; Madan Babu, M. The Rules of Disorder or Why Disorder Rules. Prog. Biophys. Mol. Biol. 2009, 99, 94–103. [Google Scholar] [CrossRef]
- Uversky, V.N.; Dunker, A.K. Understanding Protein Non-Folding. Biochim. Biophys. Acta Proteins Proteom. 2010, 1804, 1231–1264. [Google Scholar] [CrossRef] [PubMed]
- van der Lee, R.; Buljan, M.; Lang, B.; Weatheritt, R.J.; Daughdrill, G.W.; Dunker, A.K.; Fuxreiter, M.; Gough, J.; Gsponer, J.; Jones, D.T.; et al. Classification of Intrinsically Disordered Regions and Proteins. Chem. Rev. 2014, 114, 6589–6631. [Google Scholar] [CrossRef] [PubMed]
- Wright, P.E.; Dyson, H.J. Intrinsically Unstructured Proteins: Re-Assessing the Protein Structure-Function Paradigm. J. Mol. Biol. 1999, 293, 321–331. [Google Scholar] [CrossRef]
- Eunson, L.H.; Rea, R.; Zuberi, S.M.; Youroukos, S.; Panayiotopoulos, C.P.; Liguori, R.; Avoni, P.; McWilliam, R.C.; Stephenson, J.B.P.; Hanna, M.G.; et al. Clinical, Genetic, and Expression Studies of Mutations in the Potassium Channel Gene KCNA1 Reveal New Phenotypic Variability. Ann. Neurol. 2000, 48, 647–656. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nsasra, E.; Dahan, I.; Eichler, J.; Yifrach, O. It’s Time for Entropic Clocks: The Roles of Random Chain Protein Sequences in Timing Ion Channel Processes Underlying Action Potential Properties. Entropy 2023, 25, 1351. https://doi.org/10.3390/e25091351
Nsasra E, Dahan I, Eichler J, Yifrach O. It’s Time for Entropic Clocks: The Roles of Random Chain Protein Sequences in Timing Ion Channel Processes Underlying Action Potential Properties. Entropy. 2023; 25(9):1351. https://doi.org/10.3390/e25091351
Chicago/Turabian StyleNsasra, Esraa, Irit Dahan, Jerry Eichler, and Ofer Yifrach. 2023. "It’s Time for Entropic Clocks: The Roles of Random Chain Protein Sequences in Timing Ion Channel Processes Underlying Action Potential Properties" Entropy 25, no. 9: 1351. https://doi.org/10.3390/e25091351