1. Introduction
The recent global report on cancer introduced lung cancer as the leading cause of cancer-related morbidity and mortality among men, whereas, in women, it ranks third for incidence, after breast and colorectal cancer, and second for mortality, after breast cancer [
1]. By 2020, there were 2,206,771 new cases and 1,796,144 deaths because of lung cancer, accounting for 11.4% of new cases and 18% of new deaths from 36 cancers of all sites [
2]. Lung cancer is directly related to tobacco smoking, and it has been estimated that regarding the widespread use of tobacco smoking in both genders, we are facing an increased risk of lung cancer for the next decades [
1]. The 5-year survival of lung cancer is reported to be up to 30 percent in different countries, mainly depending on their management strategies and early detection programs [
2,
3,
4]. It has been demonstrated that the early detection of lung cancer improves clinical outcomes and longer survival [
2,
3,
4].
Recent studies mainly focused on identifying novel biomarkers for lung cancer, as the screening programs using computed tomography and lung biopsy are invasive and not cost-effective [
5,
6]. Among these biomarkers, extracellular vesicles are of great interest. These small particles circulating in the human bloodstream travel to every part of the body. Each extracellular vesicle contains specific proteins or nucleic acid cargos based on the originating tissue. Therefore, these small vesicles may act as messengers or indicators of specific events in their originating tissue [
7,
8]. Cancerous cells produce extracellular vesicles similar to normal tissue; however, the content of these vesicles is different from the tissue and has diagnostic potentials. Moreover, it has been suggested that tumor-derived extracellular vesicles have the potential to promote tumor progression and metastasis as well as promoting chemoresistance [
9,
10]. Long non-coding ribonucleic acids (lncRNAs) are among the extracellular vesicles’ cargos. It has been reported that every cancerous tissue has specific lncRNAs, and the extracellular vesicles originating from these tissues may contain different lncRNAs [
11,
12,
13,
14]. The extracellular vesicles containing such lncRNAs have the potential to re-program the target cells and build-up a favorable tumor environment facilitating tumor growth and metastasis [
10,
11,
15,
16]. Although the role of most of these extracellular vesicles including lncRNAs is addressed in the literature, a different expression of miRNAs between the tumor tissue, extracellular vesicles, and adjacent non-cancerous tissue (ANCT) is not widely studied in non-small cell lung cancer (NSCLC) [
17]. Among various lncRNAs introduced to be involved in the development of lung cancer, GHSROS, HMlincRNA717, HNF1A-AS1, HOTAIR, and LNCRNA-p21 are reported to have oncogenic roles [
18]. GHSROS overexpression has been linked to increased cancer cell migration in lung cancer patients. Similarly, HMlincRNA717 is reported to play its oncogenic role in NSCLC by mediating metastasis. HNF1A-AS1 and HOTAIR are regulators of the tumor cell cycle and progression in lung cancer [
18,
19]. Regarding the role of these lncRNAs in the development and progression of lung cancer, and due to a lack of experimental evidence about the effect of these lncRNAs as cargos of extracellular vesicles, the present study aimed to evaluate the differential expression of 5 lncRNAs including GHSROS, HMlincRNA717, HNF1A-AS1, HOTAIR, and LINCRNA-p21 in the tumors, tumor-derived exosomes, and ANCT in NSCLC patients.
4. Discussion
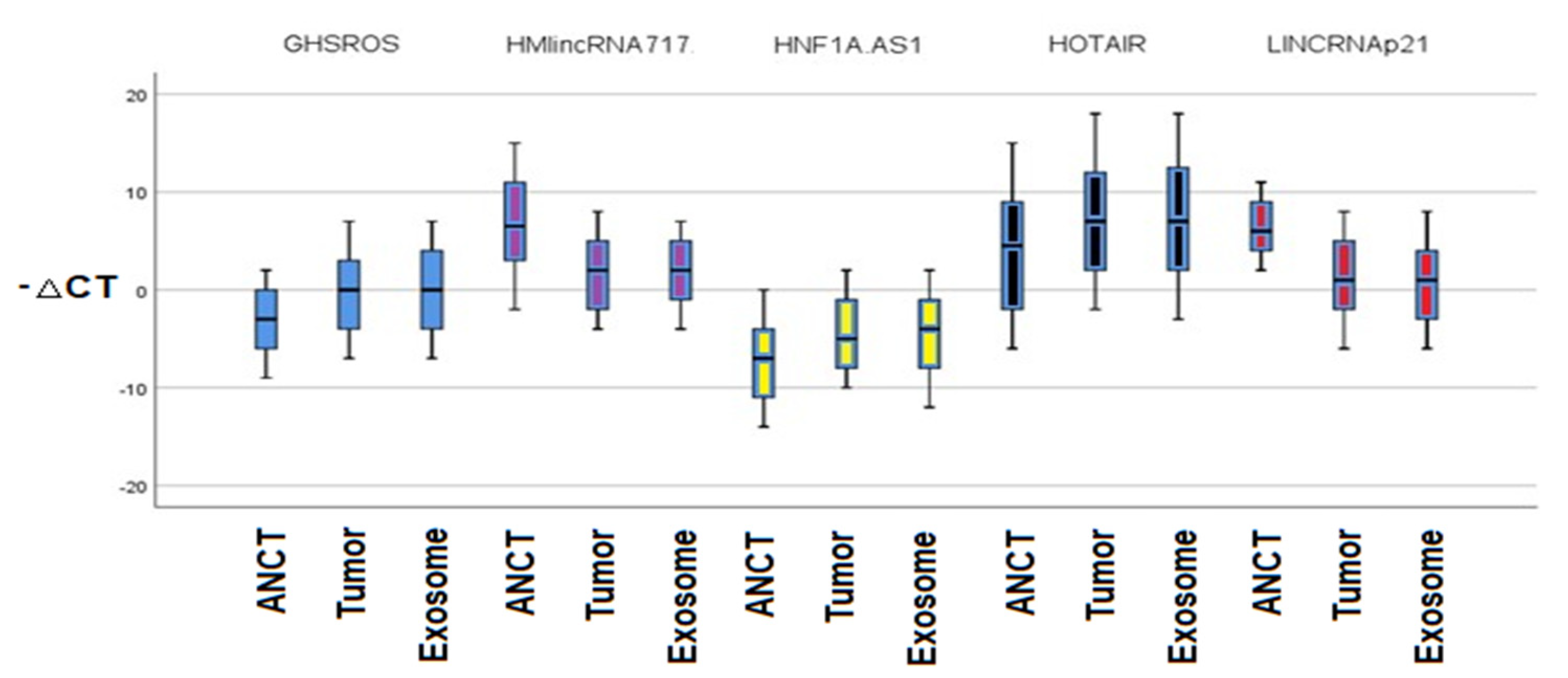
According to our study results, the relative expressions of 5 different lncRNAs including GHSROS, HMlincRNA717, HNF1A-AS1, HOTAIR, and P21 are different among the NSCLC tissue, ANCT, and plasma exosomes. The lncRNA-GHSROS, lncRNA-HNF1A-AS1, and lncRNA-HOTAIR were upregulated and the lncRNA-P21 and lncRNA-HMlincRNA717 were downregulated in tumor tissue in comparison to normal tissue. The relative expression between lncRNAs in exosome and normal tissue was significantly upregulated for lncRNA-GHSROS, lncRNA-HNF1A-AS1, and lncRNA-HOTAIR and downregulated for exosomal lncRNA-HMlincRNA717 and lncRNA-P21 in contrast to ANCT. The lncRNAs’ relative expression between tumor tissue and exosomes was not different among the studied lncRNAs.
Lung cancer is a leading cause of mortality and morbidity, and early diagnosis has been linked to better overall survival [
20,
22,
23]. Among various screening methods, serum biomarkers are a growing field in the early detection of various cancers including NSCLC. The role of lncRNAs as a regulator of many cellular processes is evident, and therefore the aberrant expression of these RNA molecules has been linked to the development of cancers. Similar to other RNA molecules, lncRNAs are secreted from various cells and also can be found in body fluids [
20,
24]. The secreted lncRNAs act as a messenger and can be packed in extracellular vesicles including exosomes. Recently, the exosomal lncRNAs in patients with cancers had provided both diagnostic and prognostic clues for the management of their malignancies [
20,
25]. We demonstrated different expression levels of 5 lncRNAs, which have been thought to have oncogenic or tumor suppressor roles in various cancers in the exosomes, ANCT, and tumor tissues of patients with NSCLC.
The lncRNA-GHSROS is an oncogenic lncRNA upregulated in many cancers. The gene coding lncRNA-GHSROS is antisense located on the ghrelin receptor encoding a 1.1 kb transcript [
26]. It has been reported that lncRNA-GHSROS is expressed in prostate and breast cancers, and its overexpression is correlated with advanced tumor stages [
25,
27]. Overexpression of this lncRNA results in increased cellular proliferation and vascularization of tumor tissues [
27]. It has been demonstrated that increased expression of this lncRNA has been linked to the migration of A549 lung cancer cell lines [
26]. Our study showed that lncRNA-GHSROS is upregulated in tumor tissue compared to the ANCT, which is in line with the previous research by Whiteside et al. Moreover, we revealed that this lncRNA is significantly overexpressed in plasma exosomes in contrast to the ANCT in NSCLC patients. However, the expression of this lncRNA is not significantly different among tumor tissues and plasma exosomes. Moreover, the expression is not related to the clinical stage or subtype of lung cancer.
The LncRNA-HMlincRNA717 has been widely studied in gastric cancer [
28]. The LncRNA-HMlincRNA717 or “gastric cancer-associated transcript-2” is among the 135 lncRNAs with aberrant expression in gastric cancer downregulated in cancer cell lines in contrast to the normal tissue [
28]. The expression level of LncRNA-HMlincRNA717 is related to venous invasion and distant metastasis [
28]. Like gastric cancer, a decreased level of this lncRNA is associated with the poor overall survival of patients with pancreatic cancer [
29]. Similarly, downregulation of LncRNA-HMlincRNA717 has been related to the poor prognosis of patients with lung cancer [
30]. A study by Xie et al. reported that the expression of LncRNA-HMlincRNA717 in NSCLC is related to metastasis and invasion, and this lncRNA can be used as a prognostic marker for lung tumors [
30]. Similarly, we demonstrated that this LncRNA-HMlincRNA717 is downregulated in lung cancer tissue in contrast to the ANCT. Furthermore, we demonstrated that this lncRNA expression level has been downregulated in plasma exosomes compared to the ANCT. However, there is no significant relationship between the expression level of this lncRNA and tumor stage or subtype.
HOTAIR is a 2.1 kb nucleotide non-coding RNA and a transcript from HOXC locus [
31]. This lncRNA interacts with various genes including HOXA5 and MMP. The lncRNA-HOTAIR promotes exosome secretion from hepatocellular carcinoma and is a novel biomarker introduced for various types of cancer. The level of this circulating lncRNA has been considered as a diagnostic marker in breast cancer patients. LncRNA-HOTAIR enhances tumor proliferation and migration in many cancers including gastric cancer, hepatocellular carcinoma, colorectal cancer, and prostate cancer [
32]. To date, only a few studies evaluated the role of exosomal lncRNA-HOTAIR in cancer. The overexpression of exosomal lncRNA-HOTAIR has been described as an indicator of poor prognosis, and also as a salivary biomarker for the early diagnosis of pancreatic cancer [
33]. Wang et al. reported that exosomal HOTAIR is positively correlated with the HER2/neu status of breast cancer [
34]. Zhang et al. study showed that exosomal HOTAIR promotes the invasion and proliferation of lung cancer [
35]. Similarly, according to our results, the expression level of lncRNA-HOTAIR is greater in tumoral tissue in contrast to the ANCT, and plasma exosomes in comparison to the ANCT.
lncRNA-P21 is a tumor suppressor which is mainly linked to p53 activity. lncRNA-p21 has been reduced in many cancers including hepatocellular carcinoma, prostate cancer, and gastric cancer. In NSCLC, the lncRNA-p21 is also downregulated, and it has been demonstrated that tumor cells with overexpression of lncRNA-p21 have a higher microvascular density and decreased expression of E-cadherin [
36]. Ao et al. demonstrated that lncRNA-p21 can inhibit NCSLC proliferation and migration [
37]. Castellano et al. demonstrated that the expression of lncRNA-p21 is related to the prognosis in lung cancer and regulation of angiogenesis [
36]. Moreover, this lncRNA can be found in body fluids, including blood, urine and plasma, and exosomes [
8]. A study by Işın et al. demonstrated lncRNA-p21 is present in urine exosomes of prostate cancer patients [
38]. Moreover, the exosomal level of lncRNA-p21 is a useful marker in distinguishing prostate cancer from benign prostate disease [
38]. Our results demonstrated that lncRNA-p21 has decreased tumor tissue and serum exosome expression in contrast to the ANCT in NSCLC patients. A study by Gezer et al. demonstrated that HOTAIR and lincRNA-p21, which show relatively low expression in Hela and MCF-7 cell lines, are enriched in exosomes [
39]. After inducing DNA damage, the lncRNA-p21 is a reliable predictor of exposure to bleomycin in contrast to the other lncRNAs [
39].
We demonstrated that the lncRNA-HNF1A-AS1 has increased relative expression in tumoral tissue and plasma exosome in contrast to ANCT. The Liu et al. study showed that lncRNA-HNF1A-AS1 upregulates in lung cancer tissue and cell lines [
40]. They demonstrated that the overexpression of lncRNA-HNF1A-AS1 is linked to NSCLC proliferation and invasion [
40]. Ma et al. reported that overexpression of this lncRNA is related to poor prognosis in NSCLC patients [
41]. However, the exosomal lncRNA-HNF1A-AS1 has not been widely studied in the literature. Luo et al. reported that exosomal lncRNA-HNF1A-AS1 secreted by the cisplatin-resistant cells could inhibit apoptosis of cervical tumors [
42,
43]. Moreover, the inhibition of exosomal lncRNA-HNF1A-AS1 could inhibit tumor formation in nude mice [
42].
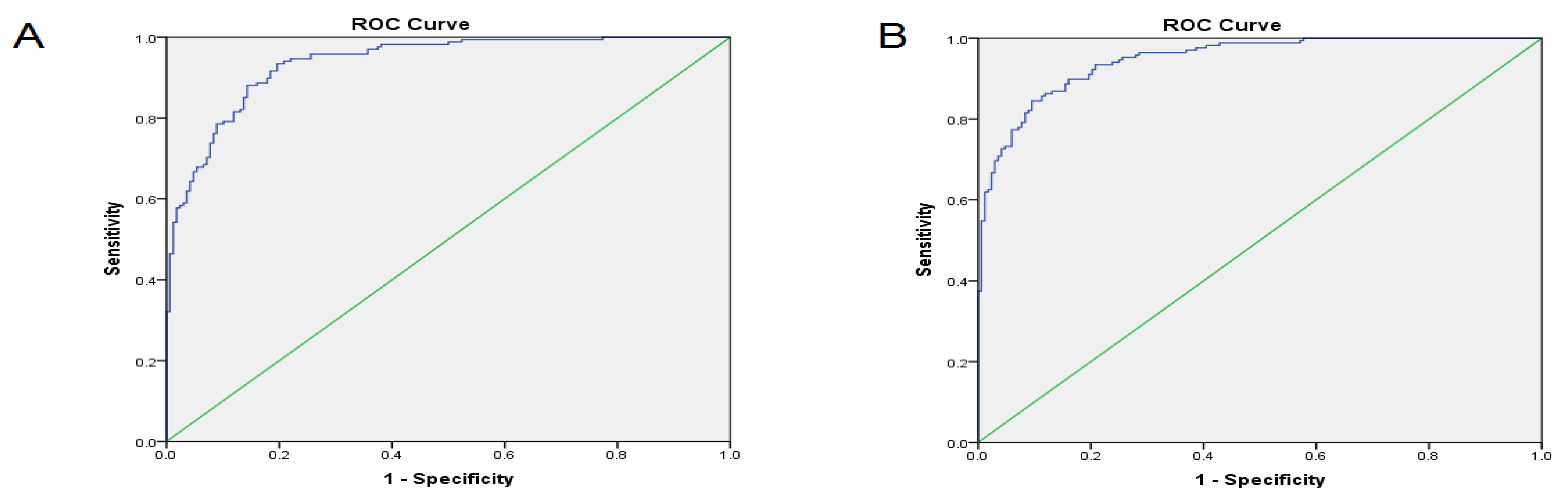
Considering a panel of these lncRNAs may be helpful in the determination of other malignancies, and further studies may consider more lung cancer-specific lncRNAs in combination with the five lncRNAs to consider a higher diagnostic accuracy for the detection of NSCLC. Furthermore, the stage of NSCLC may affect the helpfulness of considering these five lncRNAs as a diagnostic marker while we consider NSCLC from every stage. Further studies may consider specific NSCLC stages to achieve results that are more reliable.
One of the major limitations of our study was the limited sample size and absence of a control group. We used ANCT as a normal tissue, but further studies may consider normal tissues besides ANCT. Moreover, the five lncRNAs including GHSROS, HNF1A-AS1, HOTAIR, HMlincRNA717, and LINCRNA-p21 are not specific for NSCLC.







{kind=link}
{kind=link}
{kind=link}
{kind=link}