
Muscle Oxygenation Level Might Trigger the Regulation of Capillary Venous Blood Filling during Fatiguing Isometric Muscle Actions



Abstract
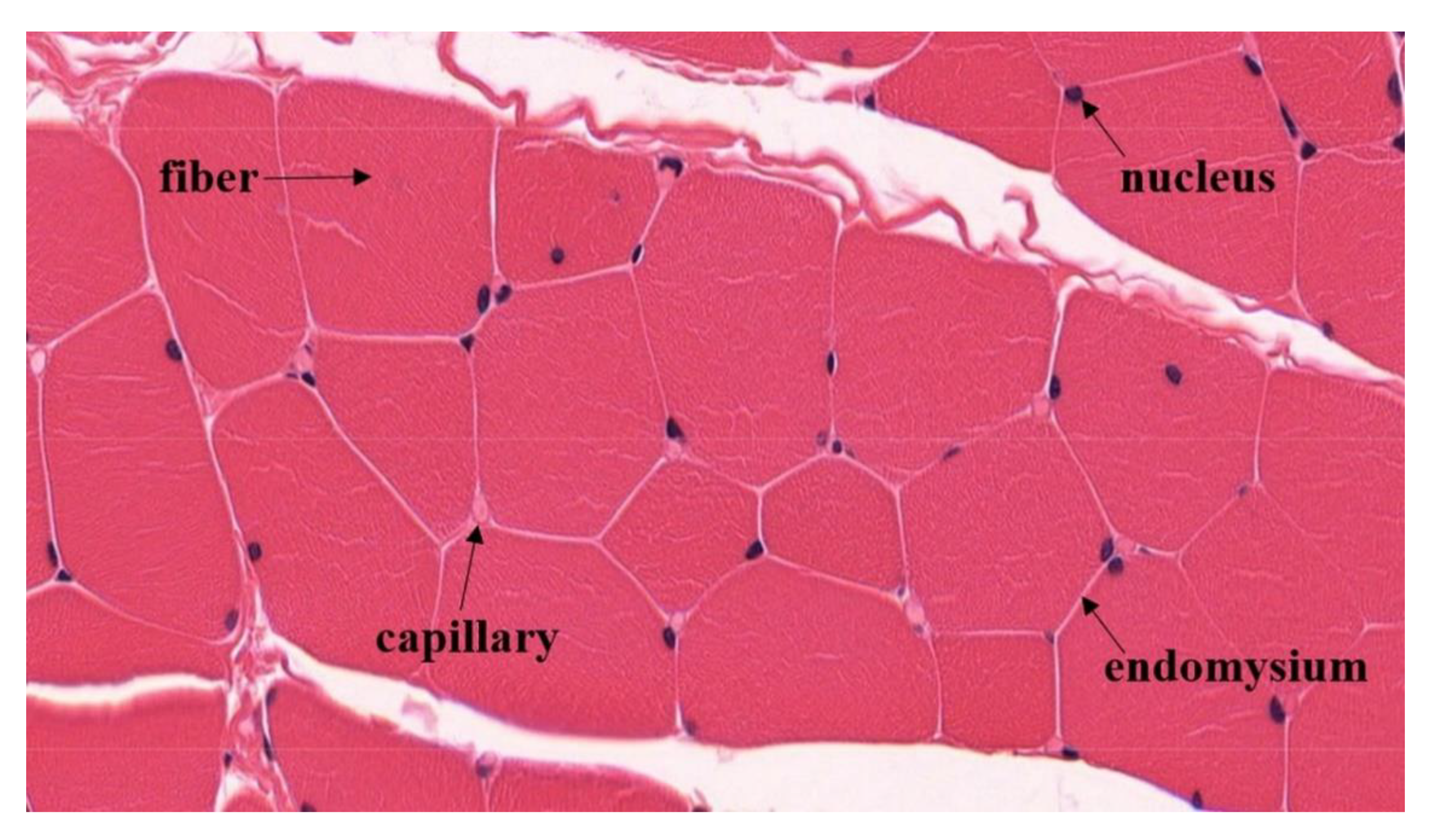
:1. Introduction
2. Materials and Methods
2.1. Subjects
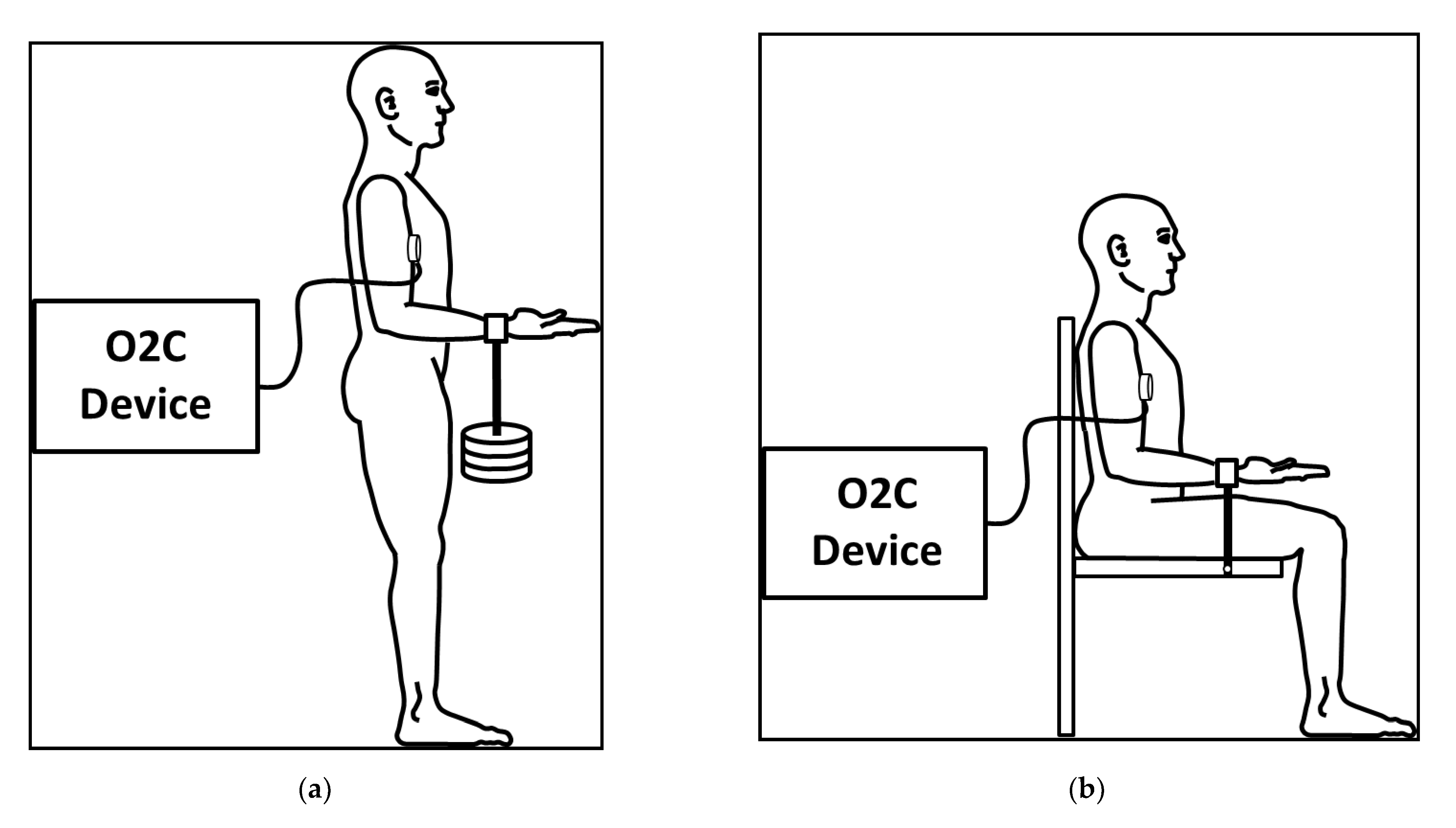
2.2. Measuring Technique
2.3. Setting and Procedure
2.4. Data Processing and Statistical Analysis
- (1)
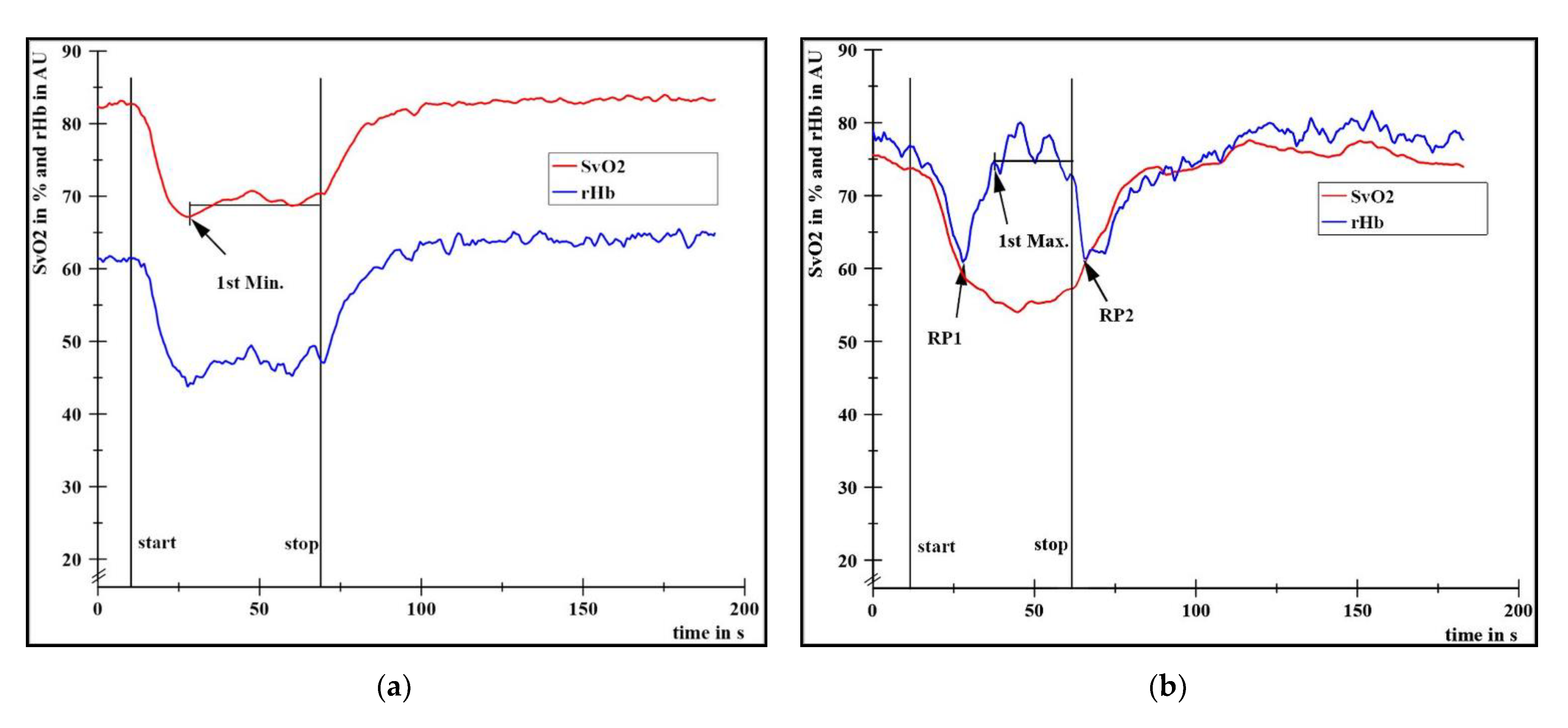
- Extent of SvO2 decrease (deoxygenation in type I and type II), calculated by the difference of the arithmetic mean (M) of the baseline values (initial 400 data points ≙ first 10 s) to the M of the plateau between the first local minimum (1st Min., Figure 2a) and stop of the task. Values of deoxygenation are presented in percent points (pp) and additionally in % related to the respective baseline value.
- (2)
- Time to task failure (TTF), defined as the time period in s from start to end of loading.
- (3)
- SvO2 levels at the moment of RP1 and RP2 (only in type II, Figure 2b), presented as Ms and 1.96-fold standard deviations (1.96SDs) in %.
- (4)
- Time period in s from start until the minimum of rHb before start of its plateau (steady state). This corresponds to RP1 in type II.
3. Results
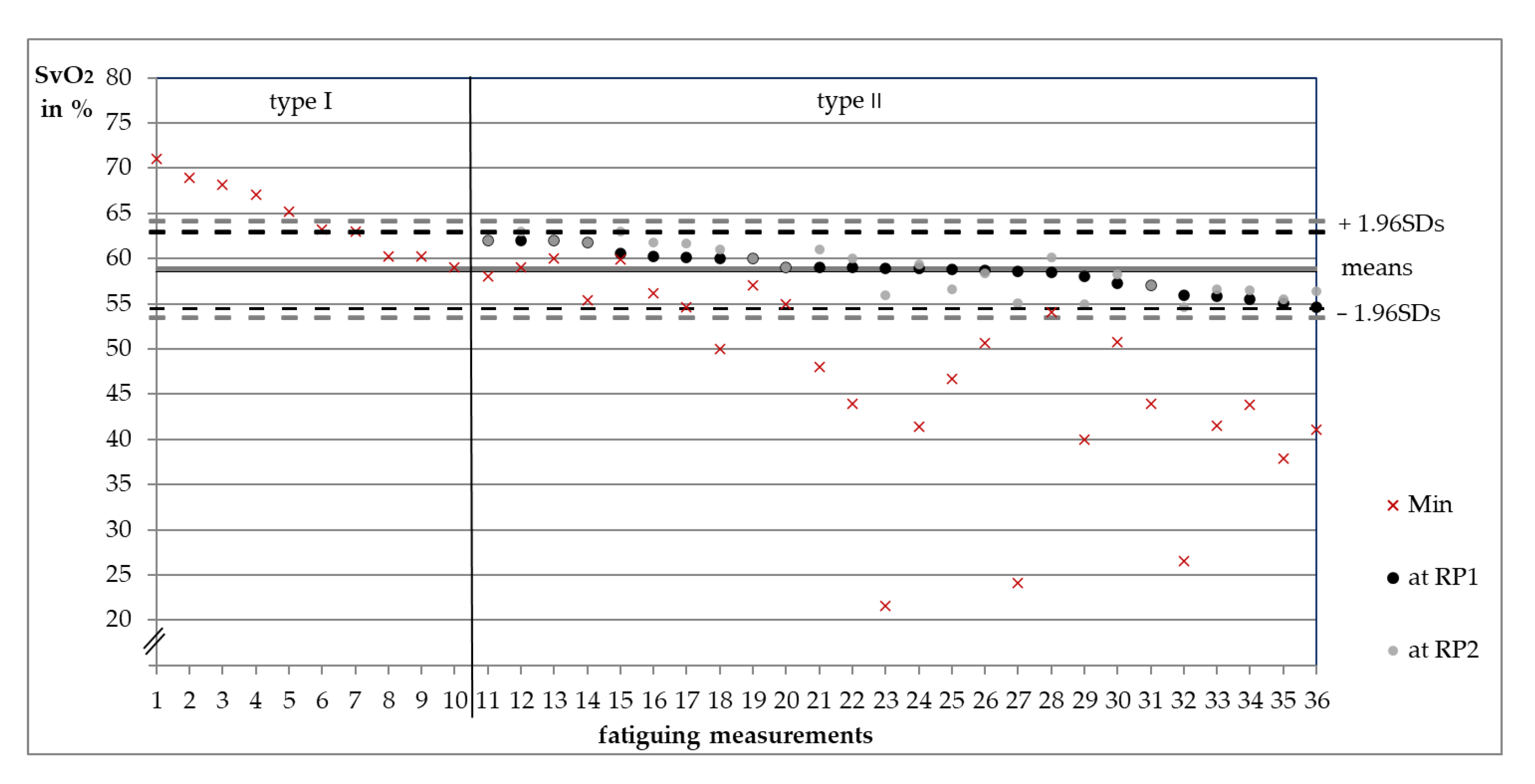
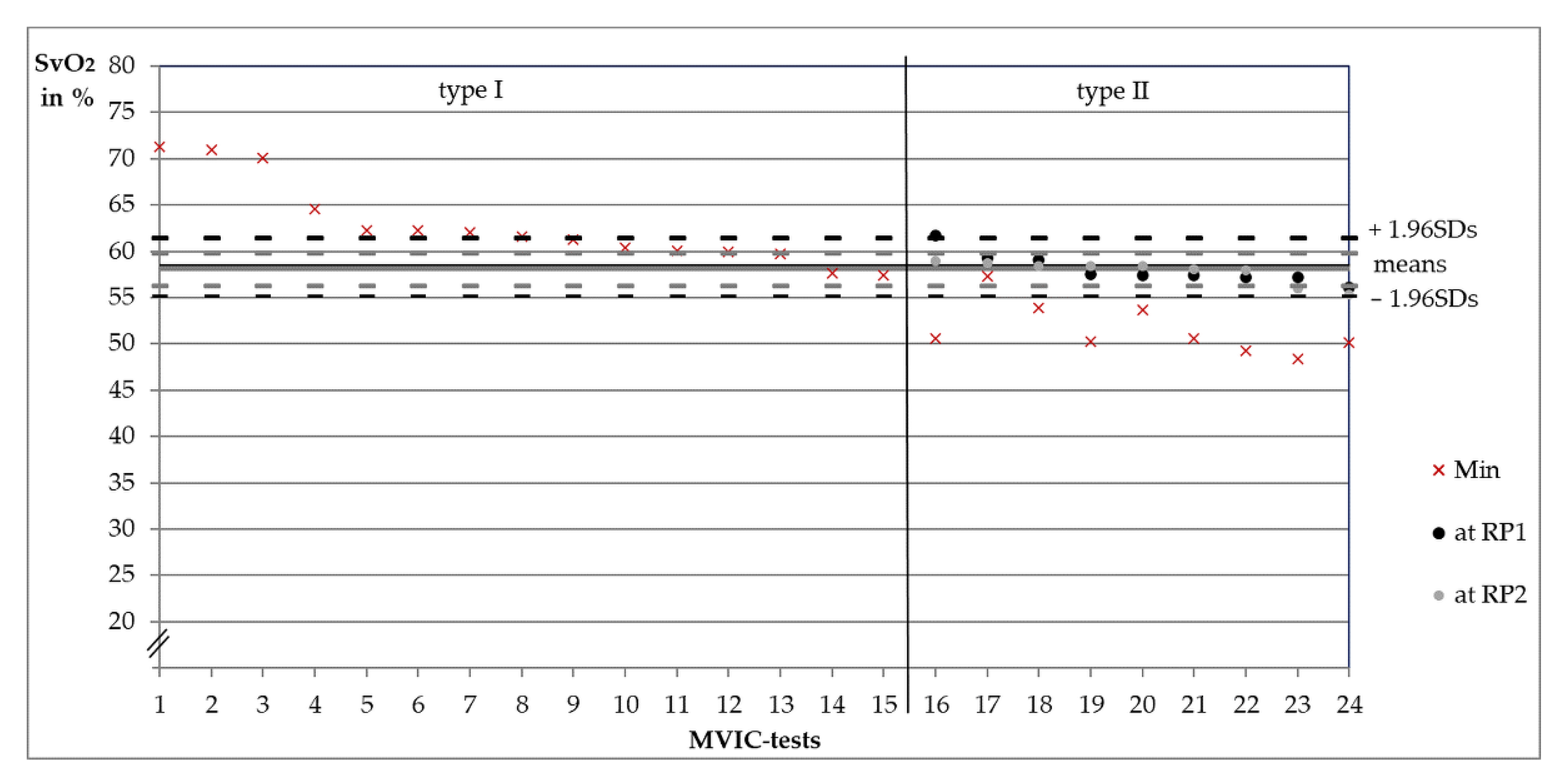
3.1. Categorization of Measurements into Type I or II
3.2. Comparisons between Behavioral Types
3.3. Oxygenation Level at Reversal Points
4. Discussion
4.1. Triggered Regulation of the Capillary Venous Blood Filling
4.2. Possible Physiological Explanation of the Regulative Response in Type II
4.3. Increased Capillary Perfusion during Isometric Muscle Actions
4.4. Study Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 1st Max. | first local maximum |
| 1st Min. | first local minimum |
| 1.96SD | 1.96-fold standard deviation |
| AU | arbitrary units |
| BMI | body mass index |
| BV | blood volume |
| HIMA | holding isometric muscle action |
| IMA | isometric muscle action |
| M | arithmetic mean |
| MMG | mechanomyography |
| MVIC | maximal voluntary isometric contraction |
| NAD+/NADH | nicotine adenine dinucleotide (oxidized/reduced, H for hydrogen) |
| NAD(P)H | reduced form of nicotinamide adenine dinucleotide phosphate |
| Nox | NAD(P)H-oxidase |
| NI DIAdemTM | National Instruments DIAdemTM |
| NIRS | near infrared spectroscopy technique |
| NO | nitric oxide |
| O2− | superoxide anion |
| O2C | Oxygen To See (device, LEA Medizintechnik GmbH) |
| PIMA | pushing or pulling isometric muscle action |
| rHb | relative hemoglobin amount |
| RP | reversal point |
| SD | standard deviation |
| SvO2 | capillary venous oxygen saturation of hemoglobin |
| tHb | total hemoglobin |
| THI | total hemoglobin index |
| TTF | time to task failure |
References
- Sadamoto, T.; Bonde-Petersen, F.; Suzuki, Y. Skeletal Muscle Tension, Flow, Pressure, and EMG during Sustained Isometric Contractions in Humans. Eur. J. Appl. Physiol. Occup. Physiol. 1983, 51, 395–408. [Google Scholar] [CrossRef] [PubMed]
- Sejersted, O.M.; Hargens, A.R.; Kardel, K.R.; Blom, P.; Jensen, O.; Hermansen, L. Intramuscular Fluid Pressure during Isometric Contraction of Human Skeletal Muscle. J. App. Physiol. 1984, 56, 287–295. [Google Scholar] [CrossRef]
- Sjøgaard, G.; Savard, G.; Juel, C. Muscle Blood Flow during Isometric Activity and Its Relation to Muscle Fatigue. Eur. J. Appl. Physiol. Occup. Physiol. 1988, 57, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Jensen, B.R.; Jørgensen, K.; Hargens, A.R.; Nielsen, P.K.; Nicolaisen, T. Physiological Response to Submaximal Isometric Contractions of the Paravertebral Muscles. Spine 1999, 24, 2332. [Google Scholar] [CrossRef]
- Järvholm, U.; Styf, J.; Suurkula, M.; Herberts, P. Intramuscular Pressure and Muscle Blood Flow in Supraspinatus. Eur. J. Appl. Physiol. 1988, 58, 219–224. [Google Scholar] [CrossRef]
- Dech, S.; Bittmann, F.; Schaefer, L. Behavior of Oxygen Saturation and Blood Filling in the Venous Capillary System of the Biceps Brachii Muscle during a Fatiguing Isometric Action. Eur. J. Transl. Myol. 2020, 30, 79–87. [Google Scholar] [CrossRef] [Green Version]
- Moalla, W.; Merzouk, A.; Costes, F.; Tabka, Z.; Ahmaidi, S. Muscle Oxygenation and EMG Activity during Isometric Exercise in Children. J. Sports Sci. 2006, 24, 1195–1201. [Google Scholar] [CrossRef]
- Akima, H.; Ando, R. Oxygenation and Neuromuscular Activation of the Quadriceps Femoris Including the Vastus Intermedius during a Fatiguing Contraction. Clin. Physiol. Funct. Imaging 2017, 37, 750–758. [Google Scholar] [CrossRef]
- Katayama, K.; Yoshitake, Y.; Watanabe, K.; Akima, H.; Ishida, K. Muscle Deoxygenation during Sustained and Intermittent Isometric Exercise in Hypoxia. Med. Sci. Sports Exerc. 2010, 42, 1269–1278. [Google Scholar] [CrossRef] [PubMed]
- Booghs, C.; Baudry, S.; Enoka, R.; Duchateau, J. Influence of Neural Adjustments and Muscle Oxygenation on Task Failure during Sustained Isometric Contractions with Elbow Flexor Muscles. Exp. Physiol. 2012, 97, 918–929. [Google Scholar] [CrossRef]
- Taelman, J.; Vanderhaegen, J.; Robijns, M.; Naulaers, G.; Spaepen, A.; Van Huffel, S. Estimation of Muscle Fatigue Using Surface Electromyography and Near-Infrared Spectroscopy. In Oxygen Transport to Tissue XXXII; LaManna, J.C., Puchowicz, M.A., Xu, K., Harrison, D.K., Bruley, D.F., Eds.; Springer: Boston, MA, USA, 2011; pp. 353–359. [Google Scholar]
- Kell, R.T.; Bhambhani, Y. Relationship between Erector Spinae Muscle Oxygenation via in Vivo near Infrared Spectroscopy and Static Endurance Time in Healthy Males. Eur. J. Appl. Physiol. 2008, 102, 243–250. [Google Scholar] [CrossRef]
- Fryer, S.; Stoner, L.; Scarrott, C.; Lucero, A.; Witter, T.; Love, R.; Dickson, T.; Draper, N. Forearm Oxygenation and Blood Flow Kinetics during a Sustained Contraction in Multiple Ability Groups of Rock Climbers. J. Sports Sci. 2015, 33, 518–526. [Google Scholar] [CrossRef] [PubMed]
- Muthalib, M.; Kerr, G.; Nosaka, K.; Perrey, S. Local Muscle Metabolic Demand Induced by Neuromuscular Electrical Stimulation and Voluntary Contractions at Different Force Levels: A NIRS Study. Eur. J. Transl. Myol. 2016, 26. [Google Scholar] [CrossRef] [Green Version]
- Pereira, M.I.R.; Gomes, P.S.C.; Bhambhani, Y.N. Acute Effects of Sustained Isometric Knee Extension on Cerebral and Muscle Oxygenation Responses. Clin. Physiol. Funct. Imaging 2009, 29, 300–308. [Google Scholar] [CrossRef] [PubMed]
- Felici, F.; Quaresima, V.; Fattorini, L.; Sbriccoli, P.; Filligoi, G.C.; Ferrari, M. Biceps Brachii Myoelectric and Oxygenation Changes during Static and Sinusoidal Isometric Exercises. J. Electromyogr. Kinesiol. 2009, 19, e1–e11. [Google Scholar] [CrossRef] [PubMed]
- Muthalib, M.; Lee, H.; Millet, G.Y.; Ferrari, M.; Nosaka, K. The Repeated-Bout Effect: Influence on Biceps Brachii Oxygenation and Myoelectrical Activity. J. Appl. Physiol. 2011, 110, 1390–1399. [Google Scholar] [CrossRef] [Green Version]
- Jones, B.; Dat, M.; Cooper, C.E. Underwater Near-Infrared Spectroscopy Measurements of Muscle Oxygenation: Laboratory Validation and Preliminary Observations in Swimmers and Triathletes. J. Biomed. Opt. 2014, 19, 127002. [Google Scholar] [CrossRef] [Green Version]
- Yoshitake, Y.; Ue, H.; Miyazaki, M.; Moritani, T. Assessment of Lower-Back Muscle Fatigue Using Electromyography, Mechanomyography, and near-Infrared Spectroscopy. Eur. J. Appl. Physiol. 2001, 84, 174–179. [Google Scholar] [CrossRef]
- Maikala, R.V.; Bhambhani, Y.N. Microvascularity of the Lumbar Erector Spinae Muscle During Sustained Prone Trunk Extension Test. In Oxygen Transport to Tissue XXX; Liss, P., Hansell, P., Bruley, D.F., Harrison, D.K., Eds.; Springer: Boston, MA, USA, 2009; pp. 67–73. [Google Scholar]
- McNeil, C.J.; Allen, M.D.; Olympico, E.; Shoemaker, J.K.; Rice, C.L. Blood Flow and Muscle Oxygenation during Low, Moderate, and Maximal Sustained Isometric Contractions. Am. J. Physiol. 2015, 309, R475–R481. [Google Scholar] [CrossRef] [Green Version]
- Frank, K.H.; Kessler, M.; Appelbaum, K.; Dummler, W. The Erlangen Micro-Lightguide Spectrophotometer EMPHO I. Phys. Med. Biol. 1989, 34, 1883–1900. [Google Scholar] [CrossRef]
- Knobloch, K.; Kraemer, R.; Lichtenberg, A.; Jagodzinski, M.; Gosling, T.; Richter, M.; Krettek, C. Microcirculation of the Ankle after Cryo/Cuff Application in Healthy Volunteers. Int. J. Sports Med. 2005, 27, 250–255. [Google Scholar] [CrossRef] [PubMed]
- Beckert, S.; Witte, M.B.; Königsrainer, A.; Coerper, S. The Impact of the Micro-Lightguide O2C for the Quantification of Tissue Ischemia in Diabetic Foot Ulcers. Diabetes Care 2004, 27, 2863–2867. [Google Scholar] [CrossRef] [Green Version]
- Jørgensen, L.P.; Schroeder, T.V. Micro-Lightguide Spectrophotometry for Tissue Perfusion in Ischemic Limbs. J. Vasc. Surg. 2012, 56, 746–752. [Google Scholar] [CrossRef] [Green Version]
- Abel, G.; Allen, J.; Drinnan, M. A Pilot Study of a New Spectrophotometry Device to Measure Tissue Oxygen Saturation. Physiol. Meas. 2014, 35, 1769–1780. [Google Scholar] [CrossRef] [PubMed]
- Forst, T.; Hohberg, C.; Tarakci, E.; Forst, S.; Kann, P.; Pfützner, A. Reliability of Lightguide Spectrophotometry (O2C®) for the Investigation of Skin Tissue Microvascular Blood Flow and Tissue Oxygen Supply in Diabetic and Nondiabetic Subjects. J. Diabetes Sci. Technol. 2008, 2, 1151–1156. [Google Scholar] [CrossRef]
- Joshi, D.; Shiwalkar, A.; Cross, M.R.; Sharma, S.K.; Vachhani, A.; Dutt, C. Continuous, Non-Invasive Measurement of the Haemodynamic Response to Submaximal Exercise in Patients with Diabetes Mellitus: Evidence of Impaired Cardiac Reserve and Peripheral Vascular Response. Heart 2010, 96, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, L.V.; Bittmann, F.N. Are There Two Forms of Isometric Muscle Action? Results of the Experimental Study Support a Distinction between a Holding and a Pushing Isometric Muscle Function. BMC Sports Sci. Med. Rehabilit. 2017, 9, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rudroff, T.; Barry, B.K.; Stone, A.L.; Barry, C.J.; Enoka, R.M. Accessory Muscle Activity Contributes to the Variation in Time to Task Failure for Different Arm Postures and Loads. J. Appl. Physiol. 2007, 102, 1000–1006. [Google Scholar] [CrossRef]
- Rudroff, T.; Justice, J.N.; Holmes, M.R.; Matthews, S.D.; Enoka, R.M. Muscle Activity and Time to Task Failure Differ with Load Compliance and Target Force for Elbow Flexor Muscles. J. Appl. Physiol. 2010, 110, 125–136. [Google Scholar] [CrossRef] [Green Version]
- Rudroff, T.; Kalliokoski, K.K.; Block, D.E.; Gould, J.R.; Klingensmith, W.C.; Enoka, R.M. PET/CT Imaging of Age- and Task-Associated Differences in Muscle Activity during Fatiguing Contractions. J. Appl. Physiol. 2013, 114, 1211–1219. [Google Scholar] [CrossRef] [Green Version]
- Garner, J.C.; Blackburn, T.; Weimar, W.; Campbell, B. Comparison of Electromyographic Activity during Eccentrically versus Concentrically Loaded Isometric Contractions. J. Electromyogr. Kinesiol. 2008, 18, 466–471. [Google Scholar] [CrossRef] [PubMed]
- Hunter, S.K.; Ryan, D.L.; Ortega, J.D.; Enoka, R.M. Task Differences With the Same Load Torque Alter the Endurance Time of Submaximal Fatiguing Contractions in Humans. J. Neurophysiol. 2002, 88, 3087–3096. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, L.V.; Bittmann, F.N. Paired Personal Interaction Reveals Objective Differences between Pushing and Holding Isometric Muscle Action. PLoS ONE 2021, 16, e0238331. [Google Scholar] [CrossRef]
- Dech, S.; Bittmann, F.N.; Schaefer, L.V. Assessment of the Adaptive Force of Elbow Extensors in Healthy Subjects Quantified by a Novel Pneumatically Driven Measurement System with Considerations of Its Quality Criteria. Diagnostics 2021, 11, 923. [Google Scholar] [CrossRef]
- Semmler, J.G.; Kornatz, K.W.; Dinenno, D.V.; Zhou, S.; Enoka, R.M. Motor Unit Synchronisation Is Enhanced during Slow Lengthening Contractions of a Hand Muscle. J. Physiol. 2002, 545, 681–695. [Google Scholar] [CrossRef]
- Kahn, J.F.; Kapitaniak, B.; Monod, H. Comparison of Two Modalities When Exerting Isometric Contractions. Europ. J. Appl. Physiol. 1985, 54, 331–335. [Google Scholar] [CrossRef]
- Schoenfeld, B.J.; Grgic, J.; Van Every, D.W.; Plotkin, D.L. Loading Recommendations for Muscle Strength, Hypertrophy, and Local Endurance: A Re-Examination of the Repetition Continuum. Sports 2021, 9, 32. [Google Scholar] [CrossRef]
- Delcanho, R.E.; Kim, Y.J.; Clark, G.T. Haemodynamic Changes Induced by Submaximal Isometric Contraction in Painful and Non-Painful Human Masseter Using near-Infra-Red Spectroscopy. Arch. Oral Biol. 1996, 41, 585–596. [Google Scholar] [CrossRef]
- Usaj, A. Differences in the Oxygenation of the Forearm Muscle during Isometric Contraction in Trained and Untrained Subjects. Cell. Mol. Biol. Lett. 2002, 7, 375–377. [Google Scholar]
- McArdle, W.D.; Katch, F.I.; Katch, V.L. Exercise Physiology: Nutrition, Energy, and Human Performance, 7th ed.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2010; ISBN 978-0-7817-9781-8. [Google Scholar]
- Golub, A.S.; Pittman, R.N. Bang-Bang Model for Regulation of Local Blood Flow. Microcirculation 2013, 20, 455–483. [Google Scholar] [CrossRef] [Green Version]
- Aizawa, S.; Tsukiyama, Y.; Koyano, K.; Clark, G.T. Reperfusion Response Changes Induced by Repeated, Sustained Contractions in Normal Human Masseter Muscle. Arch. Oral Biol. 2002, 47, 537–543. [Google Scholar] [CrossRef]
- Allen, B.W.; Stamler, J.S.; Piantadosi, C.A. Hemoglobin, Nitric Oxide and Molecular Mechanisms of Hypoxic Vasodilation. Trends Mol. Med. 2009, 15, 452–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poole, D.C.; Copp, S.W.; Hirai, D.M.; Musch, T.I. Dynamics of Muscle Microcirculatory and Blood–Myocyte O2 Flux during Contractions. Acta Physiol. 2011, 202, 293–310. [Google Scholar] [CrossRef] [PubMed]
- Poole, D.C.; Copp, S.W.; Ferguson, S.K.; Musch, T.I. Skeletal Muscle Capillary Function: Contemporary Observations and Novel Hypotheses. Exp. Physiol. 2013, 98, 1645–1658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poole, D.C.; Pittman, R.N.; Musch, T.I.; Østergaard, L. August Krogh’s Theory of Muscle Microvascular Control and Oxygen Delivery: A Paradigm Shift Based on New Data. J. Physiol. 2020, 598, 4473–4507. [Google Scholar] [CrossRef] [PubMed]
- Cayot, T.E.; Lauver, J.D.; Silette, C.R.; Scheuermann, B.W. Effects of Blood Flow Restriction Duration on Muscle Activation and Microvascular Oxygenation during Low-Volume Isometric Exercise. Clin. Physiol. Funct. Imaging 2016, 36, 298–305. [Google Scholar] [CrossRef]
- Angleys, H.; Østergaard, L. Krogh’s Capillary Recruitment Hypothesis, 100 Years on: Is the Opening of Previously Closed Capillaries Necessary to Ensure Muscle Oxygenation during Exercise? Am. J. Physiol. Heart Circ. 2019, 318, H425–H447. [Google Scholar] [CrossRef]
- Krogh, A. The Number and Distribution of Capillaries in Muscles with Calculations of the Oxygen Pressure Head Necessary for Supplying the Tissue. J. Physiol. 1919, 52, 409–415. [Google Scholar] [CrossRef]
- Krogh, A. The Supply of Oxygen to the Tissues and the Regulation of the Capillary Circulation. J. App. Physiol. 1919, 52, 457–474. [Google Scholar] [CrossRef]
- Sakai, T.; Hosoyamada, Y. Are the Precapillary Sphincters and Metarterioles Universal Components of the Microcirculation? An Historical Review. J. Physiol. Sci. 2013, 63, 319–331. [Google Scholar] [CrossRef] [Green Version]
- Segal, S.S. Regulation of Blood Flow in the Microcirculation. Microcirculation 2005, 12, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Kulandavelu, S.; Balkan, W.; Hare, J.M. Regulation of Oxygen Delivery to the Body via Hypoxic Vasodilation. Proc. Natl. Acad. Sci. USA 2015, 112, 6254–6255. [Google Scholar] [CrossRef] [Green Version]
- Dinenno, F.A. Skeletal Muscle Vasodilation during Systemic Hypoxia in Humans. J. Appl. Physiol. 2016, 120, 216–225. [Google Scholar] [CrossRef] [Green Version]
- Casey, D.P.; Joyner, M.J. Local Control of Skeletal Muscle Blood Flow during Exercise: Influence of Available Oxygen. J. Appl. Physiol. 2011, 111, 1527–1538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, A.; Rossi, E.; Scicchitano, B.M.; Coletti, D.; Moresi, V.; Adamo, S. Neurohypophyseal Hormones: Novel Actors of Striated Muscle Development and Homeostasis. Eur. J. Transl. Myol. 2014, 24, 3790. [Google Scholar] [CrossRef]
- Delong, C.; Sharma, S. Physiology, Peripheral Vascular Resistance. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Demura, S.; Nakada, M. Relationships between Force and Muscle Oxygenation Kinetics during Sustained Static Gripping Using a Progressive Workload. J. Physiol. Anthropol. 2009, 28, 109–114. [Google Scholar] [CrossRef] [Green Version]
- Haas, T.L.; Nwadozi, E. Regulation of Skeletal Muscle Capillary Growth in Exercise and Disease. Appl. Physiol. Nutr. Metab. 2015, 40, 1221–1232. [Google Scholar] [CrossRef]
- Plyley, M.; Groom, A. Geometrical Distribution of Capillaries in Mammalian Striated Muscle. Am. J. Physiol. 1975, 228, 1376–1383. [Google Scholar] [CrossRef] [PubMed]
- Al-Shammari, A.A.; Gaffney, E.A.; Egginton, S. Modelling Capillary Oxygen Supply Capacity in Mixed Muscles: Capillary Domains Revisited. J. Theor. Biol. 2014, 356, 47–61. [Google Scholar] [CrossRef] [Green Version]
- Gartner, L.P. Color Atlas and Text of Histology, 7th ed.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2017; ISBN 978-1-4963-4673-5. [Google Scholar]
- Brelje, T.C.; Sorenson, R.L. HistologyGuide.Com, University of Minnesota, MN, USA. Available online: http://histologyguide.com/slideview/MHS-262-skeletal-muscle/04-slide-1.html (accessed on 30 July 2020).
- Schaefer, L.V.; Torick, A.H.; Matuschek, H.; Holschneider, M.; Bittmann, F.N. Synchronization of Muscular Oscillations Between Two Subjects During Isometric Interaction. Eur. J. Transl. Myol. 2014, 24, 2237. [Google Scholar] [CrossRef]
- Schaefer, L.V.; Bittmann, F.N. Coherent Behavior of Neuromuscular Oscillations between Isometrically Interacting Subjects: Experimental Study Utilizing Wavelet Coherence Analysis of Mechanomyographic and Mechanotendographic Signals. Sci. Rep. 2018, 8, 15456. [Google Scholar] [CrossRef] [PubMed]
- McAuley, J.H.; Marsden, C.D. Physiological and Pathological Tremors and Rhythmic Central Motor Control. Brain 2000, 123, 1545–1567. [Google Scholar] [CrossRef]
- Schaefer, L.V.; Löffler, N.; Klein, J.; Bittmann, F.N. Mechanomyography and Acceleration Show Interlimb Asymmetries in Parkinson Patients without Tremor Compared to Controls during a Unilateral Motor Task. Sci. Rep. 2021, 11, 2631. [Google Scholar] [CrossRef]
- Schaefer, L.; Bittmann, F. Mechanotendography in Achillodynia Shows Reduced Oscillation Variability of Pre-Loaded Achilles Tendon: A Pilot Study. Eur. J. Transl. Myol. 2020, 30, 247–257. [Google Scholar] [CrossRef]
- Schaefer, L.V.; Bittmann, F.N. Parkinson Patients without Tremor Show Changed Patterns of Mechanical Muscle Oscillations during a Specific Bilateral Motor Task Compared to Controls. Sci. Rep. 2020, 10, 1168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibitoye, M.O.; Hamzaid, N.A.; Zuniga, J.M.; Abdul Wahab, A.K. Mechanomyography and Muscle Function Assessment: A Review of Current State and Prospects. Clin. Biomech. 2014, 29, 691–704. [Google Scholar] [CrossRef] [PubMed]
- Beck, T.W.; Housh, T.J.; Fry, A.C.; Cramer, J.T.; Weir, J.P.; Schilling, B.K.; Falvo, M.J.; Moore, C.A. The Influence of Muscle Fiber Type Composition on the Patterns of Responses for Electromyographic and Mechanomyographic Amplitude and Mean Power Frequency during a Fatiguing Submaximal Isometric Muscle Action. Electromyogr. Clin. Neurophysiol. 2007, 47, 221–232. [Google Scholar]
- Shinohara, M.; Søgaard, K. Mechanomyography for Studying Force Fluctuations and Muscle Fatigue. Exerc. Sport Sci. Rev. 2006, 34, 59–64. [Google Scholar] [CrossRef]
- Feldmann, A.M.; Erlacher, D.; Pfister, S.; Lehmann, R. Muscle Oxygen Dynamics in Elite Climbers during Finger-Hang Tests at Varying Intensities. Sci. Rep. 2020, 10, 3040. [Google Scholar] [CrossRef] [Green Version]
- McCully, K.K.; Hamaoka, T. Near-Infrared Spectroscopy: What Can It Tell Us about Oxygen Saturation in Skeletal Muscle? Exerc. Sport Sci. Rev. 2000, 28, 123–127. [Google Scholar]
- Ferrari, M.; Mottola, L.; Quaresima, V. Principles, Techniques, and Limitations of Near Infrared Spectroscopy. Can. J. Appl. Physiol. 2011, 29, 463–487. [Google Scholar] [CrossRef] [Green Version]
- Pereira, M.I.R.; Gomes, P.S.C.; Bhambhani, Y.N. A Brief Review of the Use of Near Infrared Spectroscopy with Particular Interest in Resistance Exercise. Sports Med. 2007, 37, 615–624. [Google Scholar] [CrossRef]
- Binzoni, T.; Cooper, C.E.; Wittekind, A.L.; Beneke, R.; Elwell, C.E.; Ville, D.V.D.; Leung, T.S. A New Method to Measure Local Oxygen Consumption in Human Skeletal Muscle during Dynamic Exercise Using Near-Infrared Spectroscopy. Physiol. Meas. 2010, 31, 1257–1269. [Google Scholar] [CrossRef] [Green Version]
- Kirkebø, A.; Wisnes, A. Regional Tissue Fluid Pressure in Rat Calf Muscle during Sustained Contraction or Stretch. Acta Physiol. 1982, 114, 551–556. [Google Scholar] [CrossRef]
- Zanato, R.; Martino, L.; Carraro, U.; Kern, H.; Rossato, E.; Masiero, S.; Stramare, R. Functional Echomyography: Thickness, Ecogenicity, Contraction and Perfusion of the LMN Denervated Human Muscle before and during h-BFES. Eur. J. Transl. Myol. 2010, 20, 33–40. [Google Scholar] [CrossRef]
- Recenti, M.; Ricciardi, C.; Edmunds, K.; Gislason, M.K.; Gargiulo, P. Machine Learning Predictive System Based upon Radiodensitometric Distributions from Mid-Thigh CT Images. Eur. J. Transl. Myol. 2020, 30, 121–124. [Google Scholar] [CrossRef] [Green Version]
- Kern, H.; Carraro, U. Home-Based Functional Electrical Stimulation of Human Permanent Denervated Muscles: A Narrative Review on Diagnostics, Managements, Results and Byproducts Revisited 2020. Diagnostics 2020, 10, 529. [Google Scholar] [CrossRef]
- Van Beekvelt, M.C.P.; Borghuis, M.S.; Van Engelen, B.G.M.; Wevers, R.A.; Colier, W.N.J.M. Adipose Tissue Thickness Affects in Vivo Quantitative Near-IR Spectroscopy in Human Skeletal Muscle. Clin. Sci. 2001, 101, 21–28. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HIMA | PIMA | |||||
|---|---|---|---|---|---|---|
| Task | Fatiguing | Fatiguing | MVIC-Test | |||
| arm | left | right | left | right | left | right |
| type I (n = 25) | 5 | 2 | 3 | 0 | 8 | 7 |
| 7 | 3 | 15 | ||||
| type II (n = 35) | 7 | 10 | 3 | 6 | 4 | 5 |
| 17 | 9 | 9 | ||||
| total (n = 60) | 12 | 12 | 6 | 6 | 12 | 12 |
| 24 | 12 | 24 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dech, S.; Bittmann, F.N.; Schaefer, L.V. Muscle Oxygenation Level Might Trigger the Regulation of Capillary Venous Blood Filling during Fatiguing Isometric Muscle Actions. Diagnostics 2021, 11, 1973. https://doi.org/10.3390/diagnostics11111973
Dech S, Bittmann FN, Schaefer LV. Muscle Oxygenation Level Might Trigger the Regulation of Capillary Venous Blood Filling during Fatiguing Isometric Muscle Actions. Diagnostics. 2021; 11(11):1973. https://doi.org/10.3390/diagnostics11111973
Chicago/Turabian StyleDech, Silas, Frank N. Bittmann, and Laura V. Schaefer. 2021. "Muscle Oxygenation Level Might Trigger the Regulation of Capillary Venous Blood Filling during Fatiguing Isometric Muscle Actions" Diagnostics 11, no. 11: 1973. https://doi.org/10.3390/diagnostics11111973