Characterization of CD147, CA9, and CD70 as Tumor-Specific Markers on Extracellular Vesicles in Clear Cell Renal Cell Carcinoma

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patient Samples
2.2. Cell Culture
2.3. Exosome Isolation from Cell Culture Supernatants
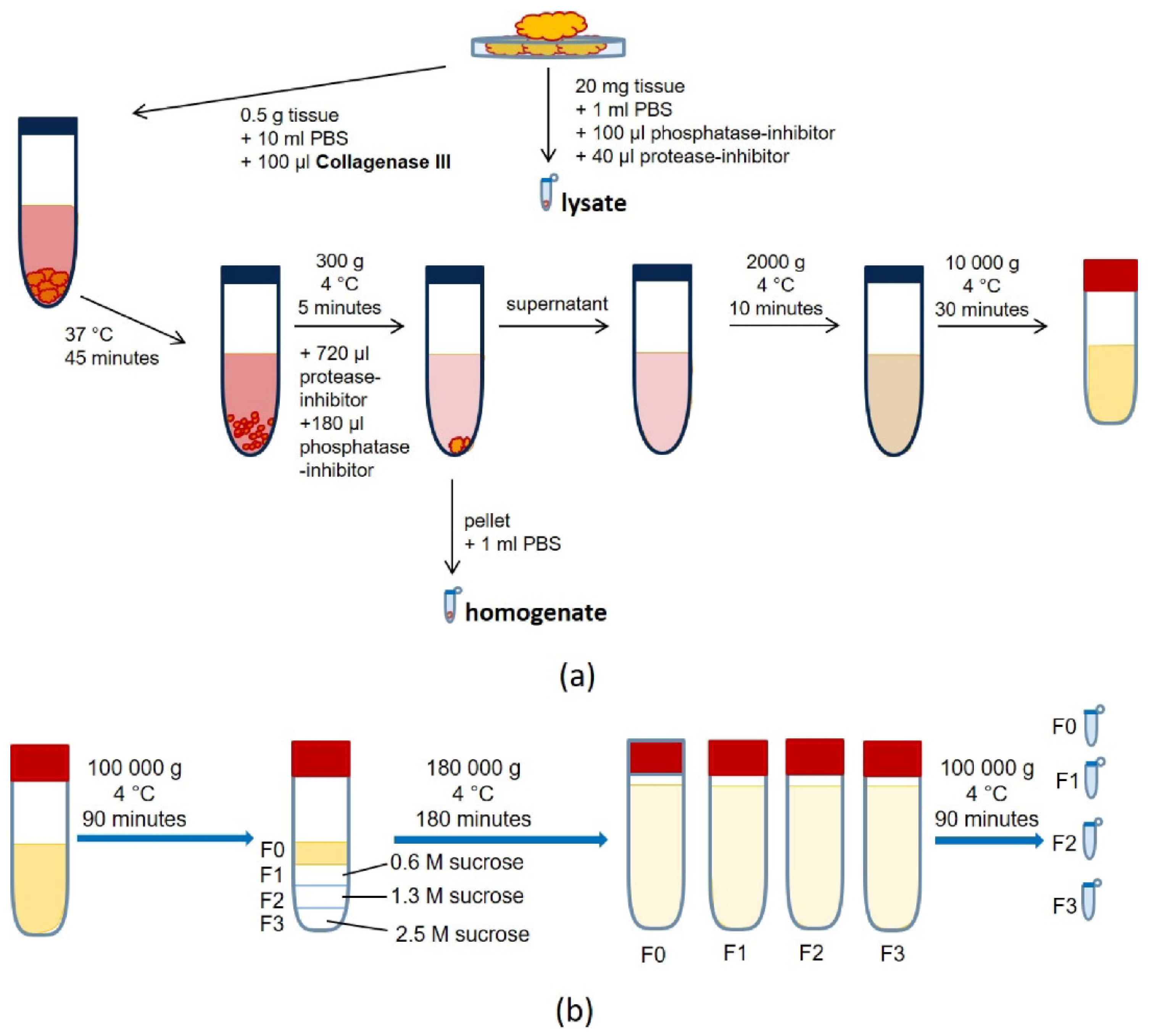
2.4. Exosome Isolation from Tissue
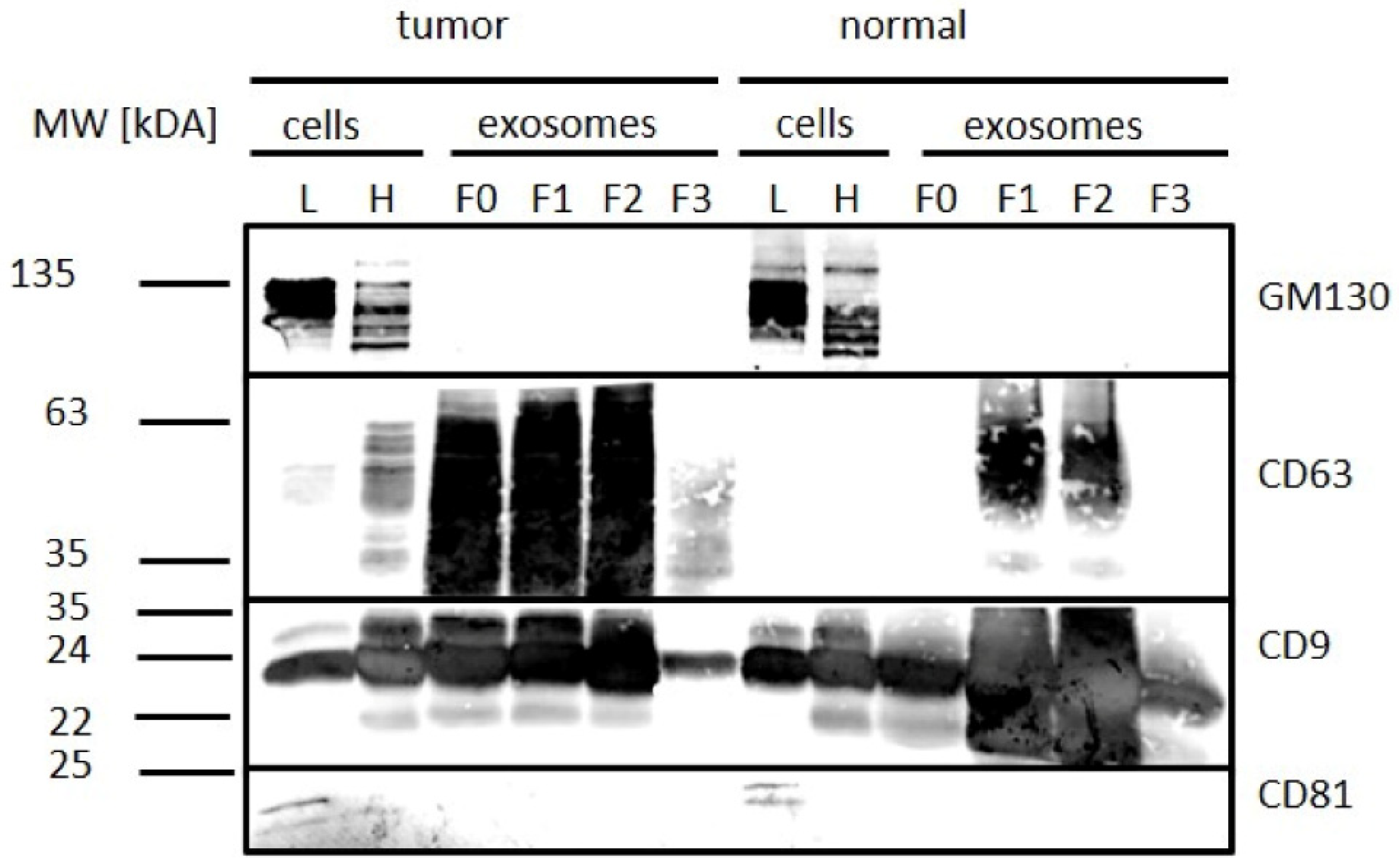
2.5. Characterization of Exosomes
2.6. Immunohistochemistry
3. Results
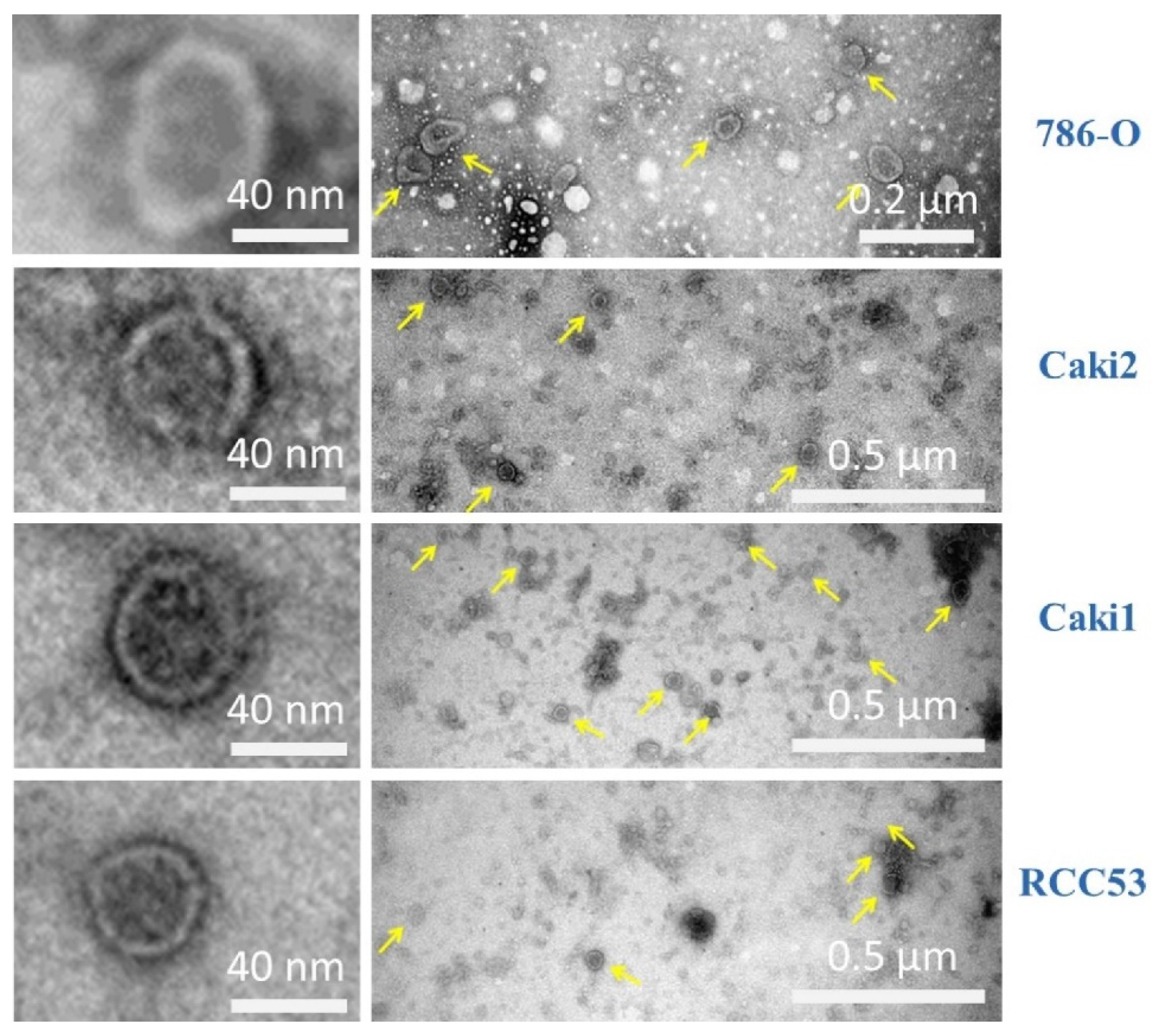
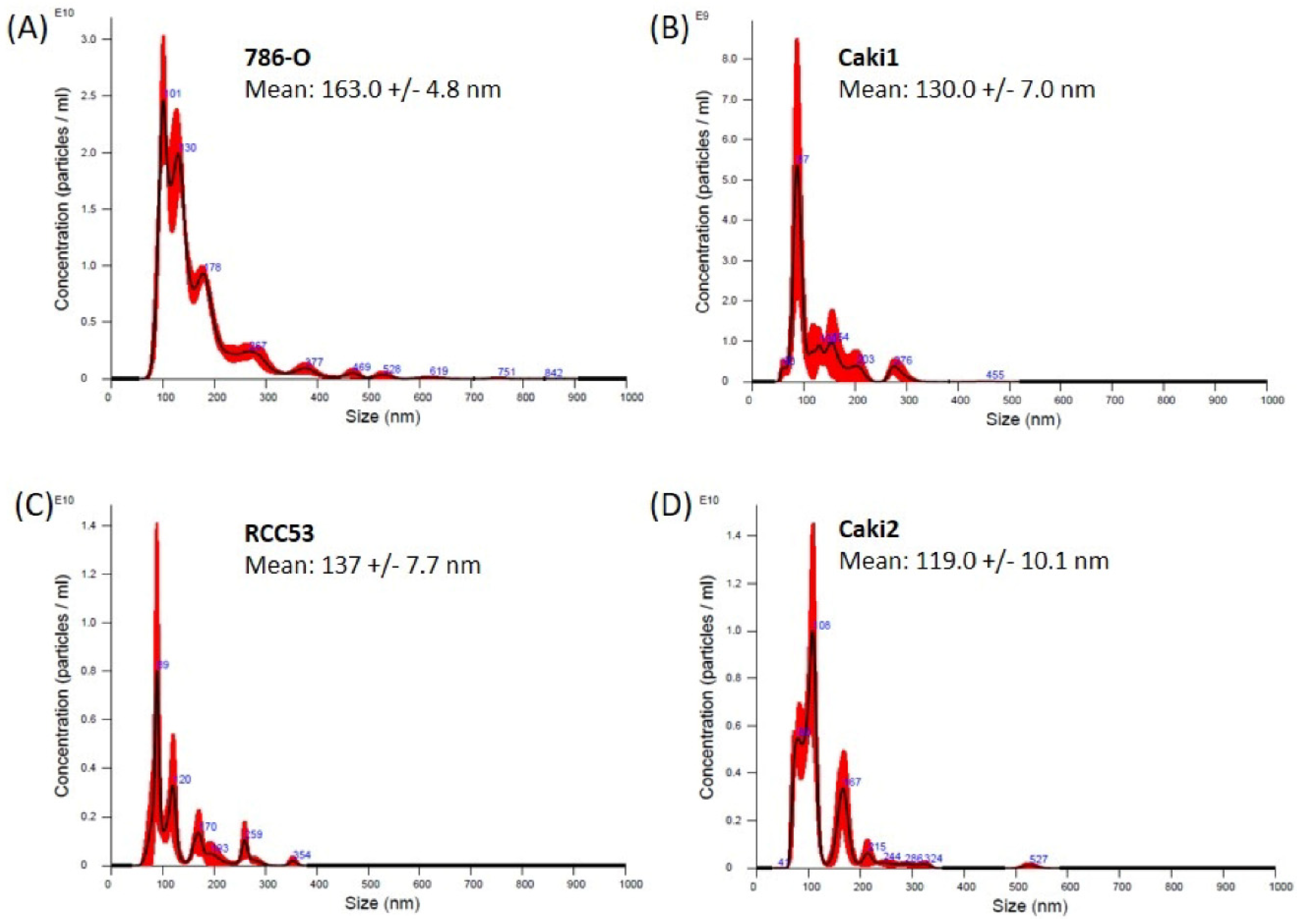
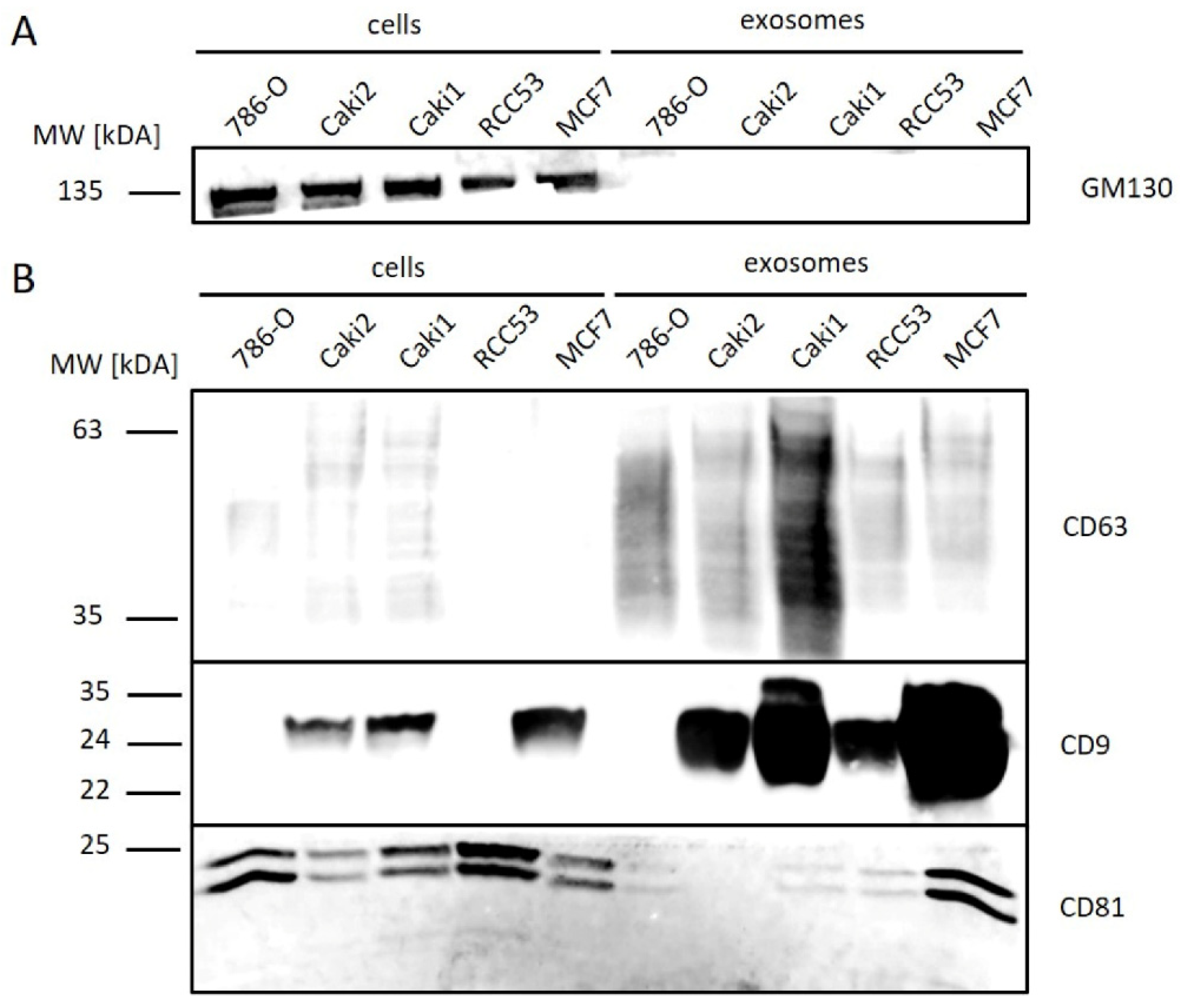
3.1. Exosome Isolation from Cell Culture
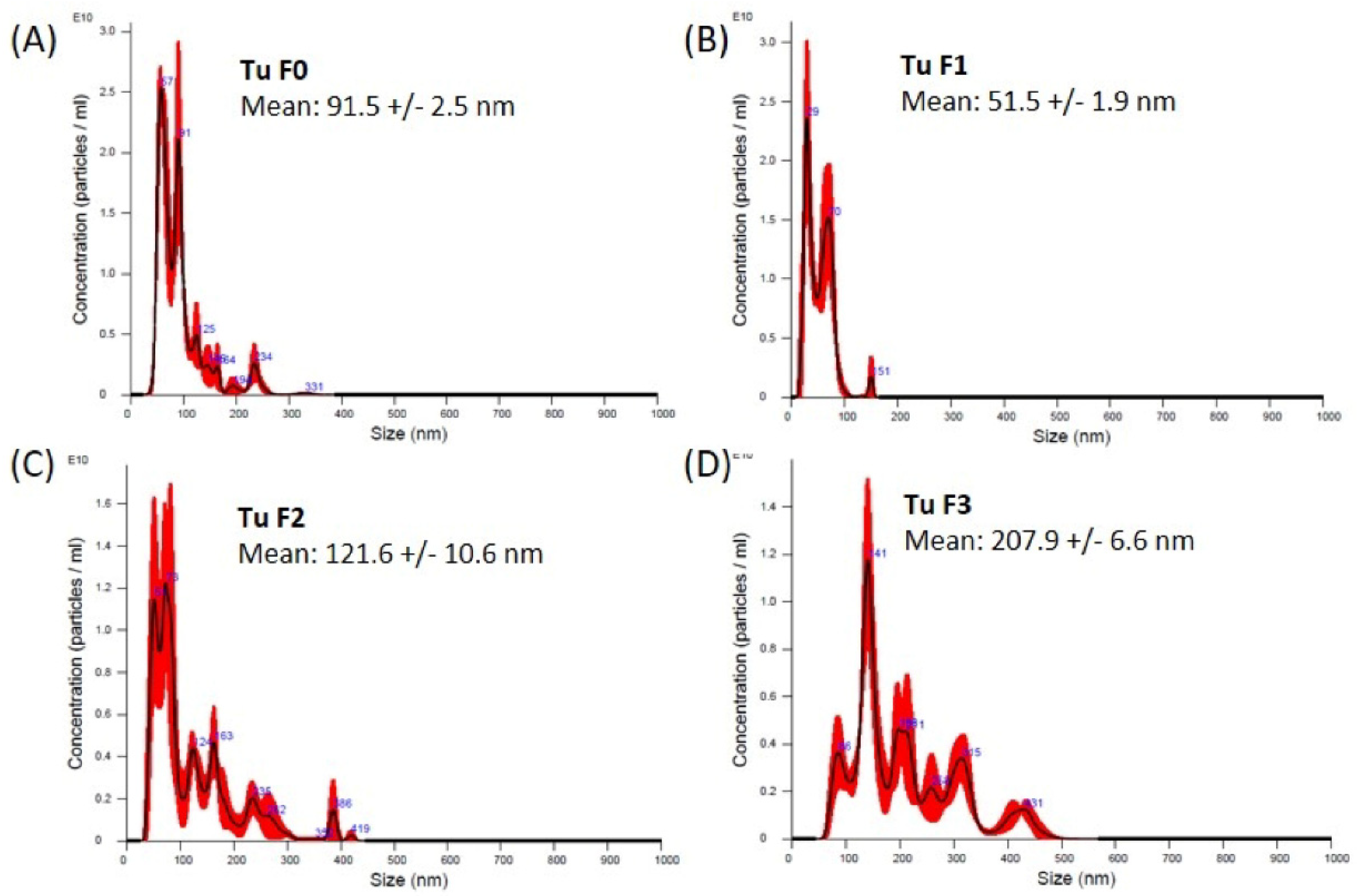
3.2. Exosome Isolation from Tissue Samples
3.3. Expression of Tumor Markers in Cells and Tissues
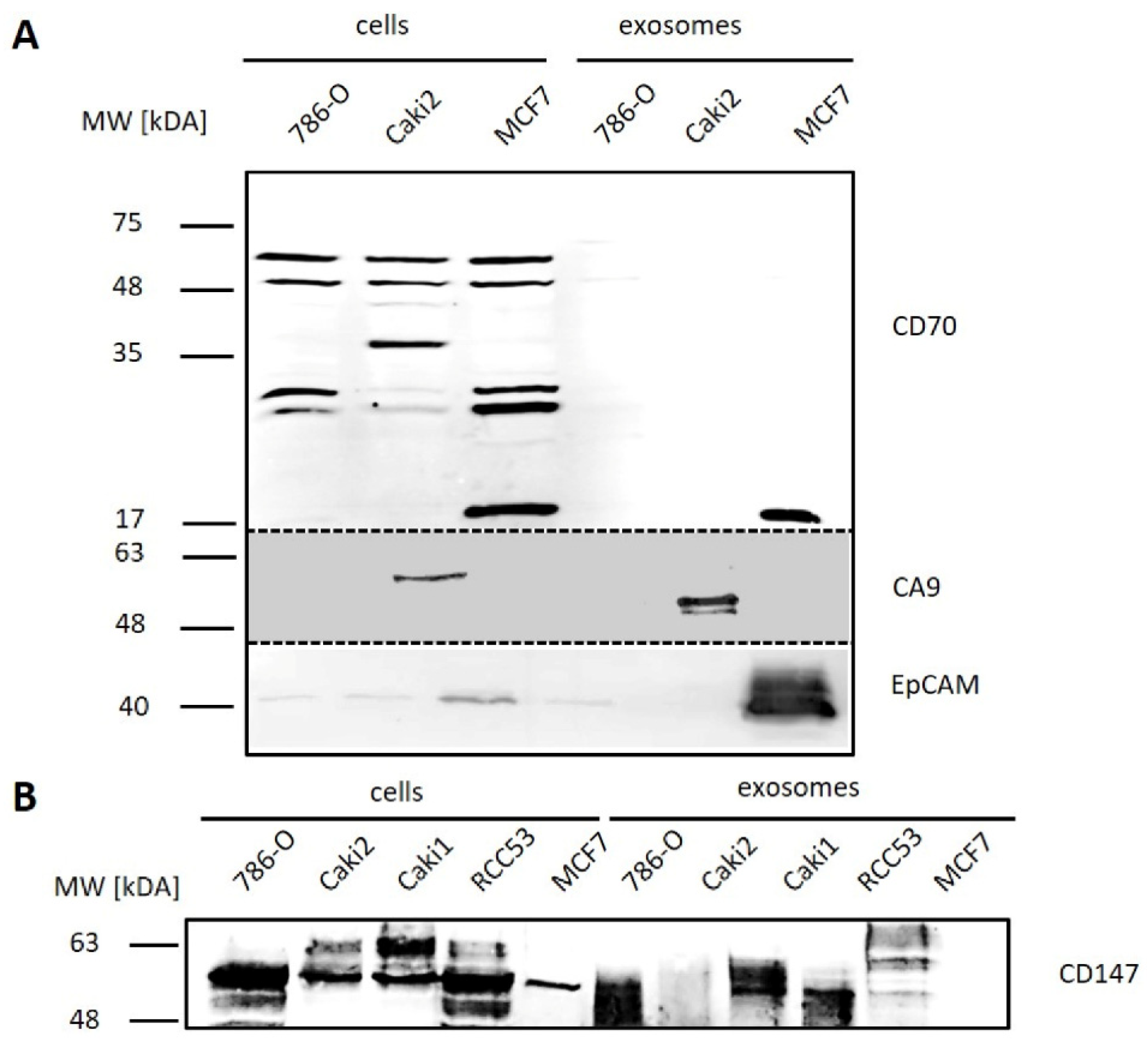
3.3.1. Cell Culture
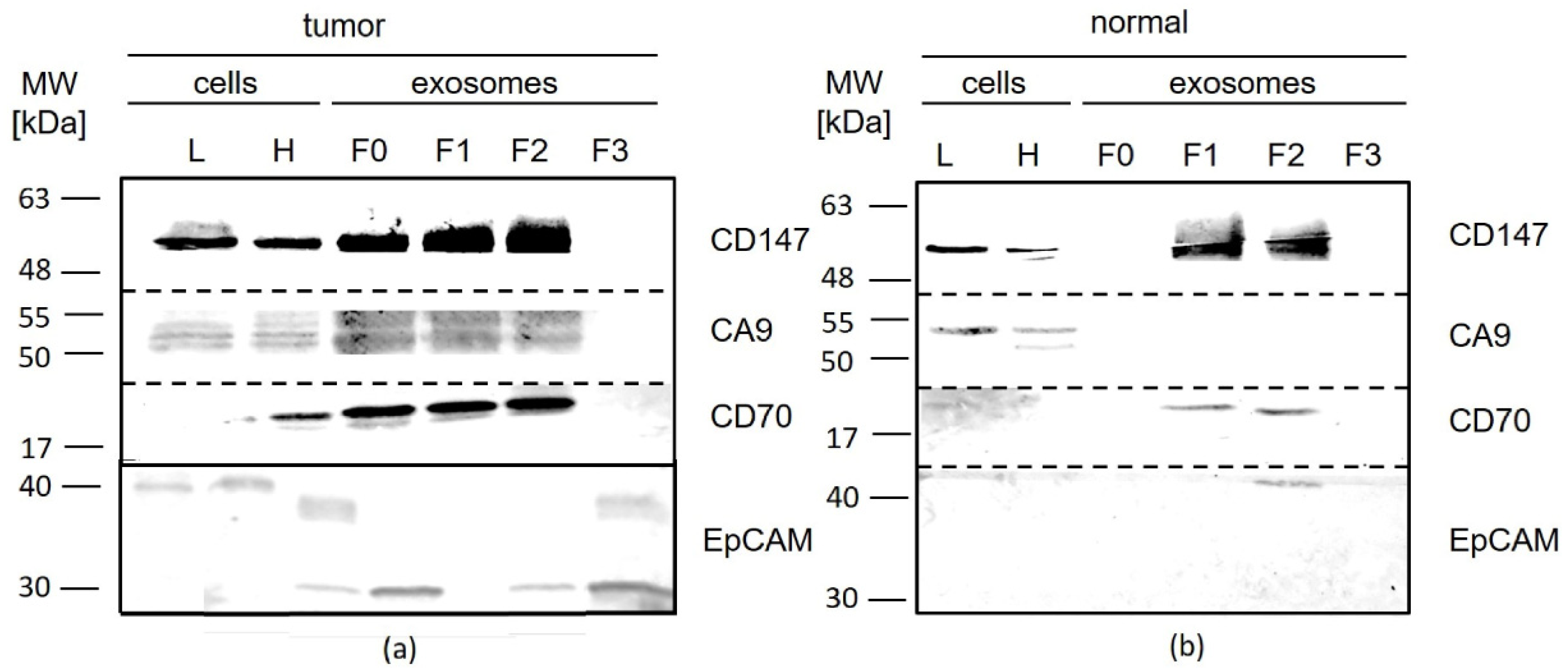
3.3.2. Primary Tumor Tissue
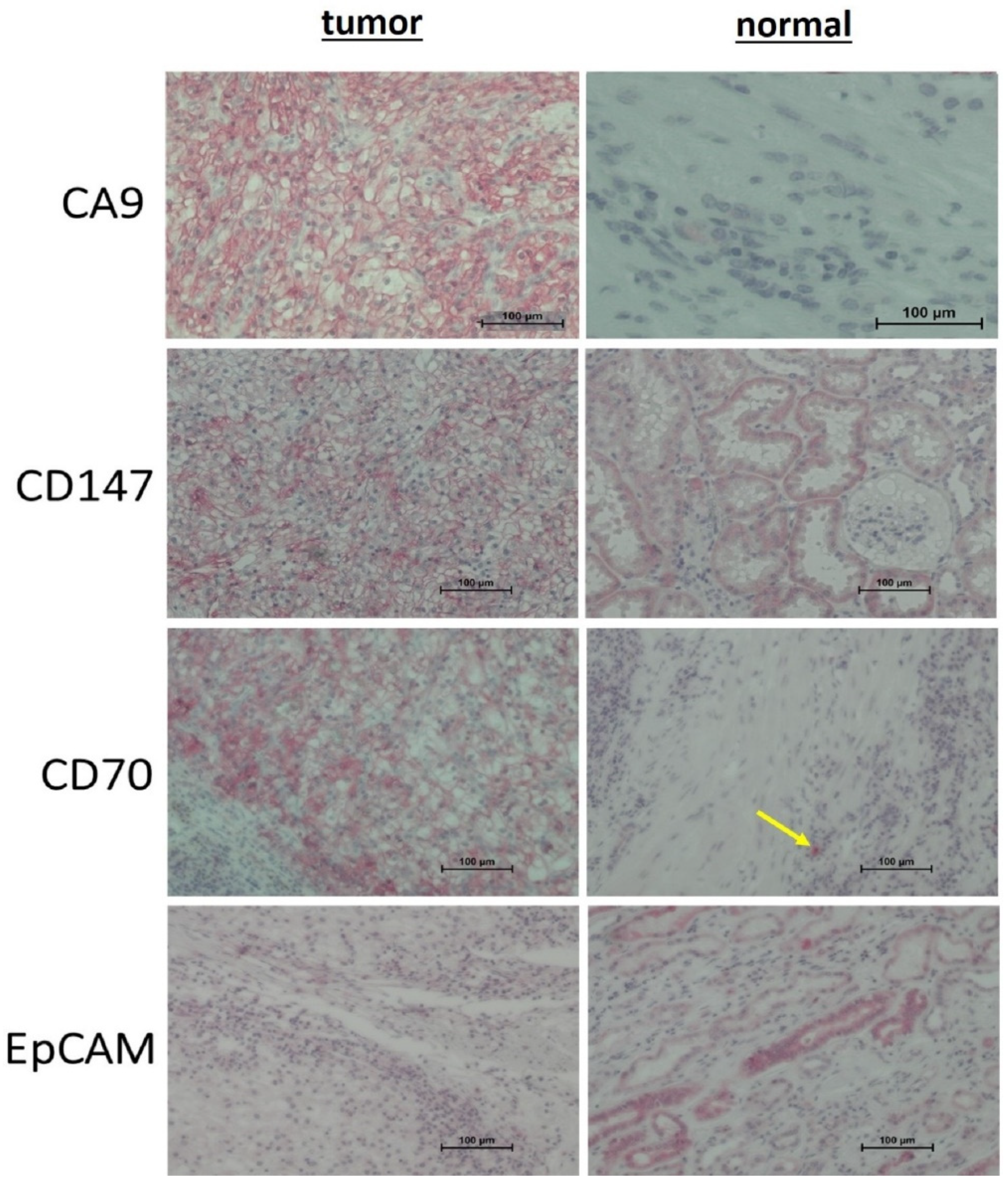
3.3.3. Immunohistochemistry
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Global Burden of Disease Cancer Collaboration; Fitzmaurice, C.; Akinyemiju, T.F.; Al Lami, F.H.; Alam, T.; Alizadeh-Navaei, R.; Allen, C.; Alsharif, U.; Alvis-Guzman, N.; Amini, E.; et al. Global, Regional, and National Cancer Incidence, Mortality, Years of Life Lost, Years Lived With Disability, and Disability-Adjusted Life-Years for 29 Cancer Groups, 1990 to 2016: A Systematic Analysis for the Global Burden of Disease Study. JAMA Oncol. 2018, 4, 1553–1568. [Google Scholar] [CrossRef]
- Barr, R.D.; Ferrari, A.; Ries, L.; Whelan, J.; Bleyer, W.A. Cancer in Adolescents and Young Adults: A Narrative Review of the Current Status and a View of the Future. JAMA Pediatr. 2016, 170, 495–501. [Google Scholar] [CrossRef]
- Glazer, A.M.; Rigel, D.S.; Winkelmann, R.R.; Farberg, A.S. Clinical Diagnosis of Skin Cancer: Enhancing Inspection and Early Recognition. Dermatol Clin. 2017, 35, 409–416. [Google Scholar] [CrossRef] [PubMed]
- Ott, J.J.; Ullrich, A.; Miller, A.B. The importance of early symptom recognition in the context of early detection and cancer survival. Eur. J. Cancer 2009, 45, 2743–2748. [Google Scholar] [CrossRef] [PubMed]
- Jones, C.E.; Maben, J.; Jack, R.H.; Davies, E.A.; Forbes, L.J.L.; Lucas, G.; Ream, E. A systematic review of barriers to early presentation and diagnosis with breast cancer among black women. BMJ Open 2014, 4, e004076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, D.; Friend, C.; Dreher, A.; Allgar, V.; Macleod, U. The diagnostic test accuracy of rectal examination for prostate cancer diagnosis in symptomatic patients: A systematic review. BMC Fam. Pract. 2018, 19, 79. [Google Scholar] [CrossRef] [Green Version]
- Armstrong, A.J.; Eisenberger, M.A.; Halabi, S.; Oudard, S.; Nanus, D.M.; Petrylak, D.P.; Sartor, A.O.; Scher, H.I. Biomarkers in the management and treatment of men with metastatic castration-resistant prostate cancer. Eur. Urol. 2012, 61, 549–559. [Google Scholar] [CrossRef] [Green Version]
- Du, M.; Van Ness, S.; Gordeuk, V.; Nouraie, S.M.; Nekhai, S.; Gladwin, M.; Steinberg, M.H.; Sebastiani, P. Biomarker signatures of sickle cell disease severity. Blood Cells Mol. Dis. 2018, 72, 1–9. [Google Scholar] [CrossRef]
- Mankarious, M.; Matthews, N.C.; Snowden, J.A.; Alfred, A. Extracorporeal Photopheresis (ECP) and the Potential of Novel Biomarkers in Optimizing Management of Acute and Chronic Graft vs. Host Disease (GvHD). Front. Immunol. 2020, 11, 81. [Google Scholar] [CrossRef]
- Hendriks, R.J.; van Oort, I.M.; Schalken, J.A. Blood-based and urinary prostate cancer biomarkers: A review and comparison of novel biomarkers for detection and treatment decisions. Prostate Cancer Prostatic Dis. 2017, 20, 12–19. [Google Scholar] [CrossRef]
- Poulet, G.; Massias, J.; Taly, V. Liquid Biopsy: General Concepts. Acta Cytol. 2019, 63, 449–455. [Google Scholar] [CrossRef] [PubMed]
- Dinges, S.S.; Hohm, A.; Vandergrift, L.A.; Nowak, J.; Habbel, P.; Kaltashov, I.A.; Cheng, L.L. Cancer metabolomic markers in urine: Evidence, techniques and recommendations. Nat. Rev. Urol. 2019, 16, 339–362. [Google Scholar] [CrossRef] [PubMed]
- Mittelbrunn, M.; Sánchez-Madrid, F. Intercellular communication: Diverse structures for exchange of genetic information. Nat. Rev. Mol. Cell Biol. 2012, 13, 328–335. [Google Scholar] [CrossRef] [PubMed]
- Elsharkawi, F.; Elsabah, M.; Shabayek, M.; Khaled, H. Urine and Serum Exosomes as Novel Biomarkers in Detection of Bladder Cancer. Asian Pac. J. Cancer Prev. 2019, 20, 2219–2224. [Google Scholar] [CrossRef] [Green Version]
- Vlassov, A.V.; Magdaleno, S.; Setterquist, R.; Conrad, R. Exosomes: Current knowledge of their composition, biological functions, and diagnostic and therapeutic potentials. Biochim. Biophys. Acta 2012, 1820, 940–948. [Google Scholar] [CrossRef]
- Li, M.; Zeringer, E.; Barta, T.; Schageman, J.; Cheng, A.; Vlassov, A.V. Analysis of the RNA content of the exosomes derived from blood serum and urine and its potential as biomarkers. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369, 20130502. [Google Scholar] [CrossRef]
- Ståhl, A.L.; Johansson, K.; Mossberg, M.; Kahn, R.; Karpman, D. Exosomes and microvesicles in normal physiology, pathophysiology, and renal diseases. Pediatr. Nephrol. 2019, 34, 11–30. [Google Scholar] [CrossRef] [Green Version]
- Qin, J.; Xu, Q. Functions and application of exosomes. Acta Pol. Pharm. 2014, 71, 537–543. [Google Scholar]
- Pisitkun, T.; Shen, R.F.; Knepper, M.A. Identification and proteomic profiling of exosomes in human urine. Proc. Natl. Acad. Sci. USA 2004, 101, 13368–13373. [Google Scholar] [CrossRef] [Green Version]
- Baumgart, S.; Hölters, S.; Ohlmann, C.H.; Bohle, R.; Stöckle, M.; Ostenfeld, M.S.; Dyrskjøt, L.; Junker, K.; Heinzelmann, J. Exosomes of invasive urothelial carcinoma cells are characterized by a specific miRNA expression signature. Oncotarget. 2017, 8, 58278–58291. [Google Scholar] [CrossRef] [Green Version]
- Zeuschner, P.; Linxweiler, J.; Junker, K. Non-coding RNAs as biomarkers in liquid biopsies with a special emphasis on extracellular vesicles in urological malignancies. Expert Rev. Mol. Diagn. 2020, 20, 151–167. [Google Scholar] [CrossRef] [PubMed]
- Junker, K.; Heinzelmann, J.; Beckham, C.; Ochiya, T.; Jenster, G. Extracellular Vesicles and Their Role in Urologic Malignancies. Eur. Urol. 2016, 70, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.H.; Ji, J.; Wang, B.C.; Chen, H.; Yang, Z.H.; Wang, K.; Luo, C.L.; Zhang, W.W.; Wang, F.-B.; Zhang, X.-L. Tumor-Derived Exosomal Long Noncoding RNAs as Promising Diagnostic Biomarkers for Prostate Cancer. Cell. Physiol. Biochem. 2018, 46, 532–545. [Google Scholar] [CrossRef] [PubMed]
- Linxweiler, J.; Junker, K. Extracellular vesicles in urological malignancies: An update. Nat. Rev. Urol. 2020, 17, 11–27. [Google Scholar] [CrossRef]
- Zhang, W.; Ni, M.; Su, Y.; Wang, H.; Zhu, S.; Zhao, A.; Li, G. MicroRNAs in Serum Exosomes as Potential Biomarkers in Clear-cell Renal Cell Carcinoma. Eur. Urol. Focus 2018, 4, 412–419. [Google Scholar] [CrossRef] [Green Version]
- Van der Gun, B.T.; Melchers, L.J.; Ruiters, M.H.; de Leij, L.F.; McLaughlin, P.M.; Rots, M.G. EpCAM in carcinogenesis: The good, the bad or the ugly. Carcinogenesis 2010, 31, 1913–1921. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Liao, G.; Li, Y.; Zhou, S.; Zou, H.; Fernando, S. Prognostic value of carbonic anhydrase IX immunohistochemical expression in renal cell carcinoma: A meta-analysis of the literature. PLoS ONE 2014, 9, e114096. [Google Scholar] [CrossRef]
- Adam, P.J.; Terrett, J.A.; Steers, G.; Stockwin, L.; Loader, J.A.; Fletcher, G.C.; Lu, L.S.; Leach, B.I.; Mason, S.; Stamps, A.C.; et al. CD70 (TNFSF7) is expressed at high prevalence in renal cell carcinomas and is rapidly internalised on antibody binding. Br. J. Cancer 2006, 95, 298–306. [Google Scholar] [CrossRef]
- Mori, Y.; Masuda, T.; Kosugi, T.; Yoshioka, T.; Hori, M.; Nagaya, H.; Maeda, K.; Sato, Y.; Kojima, H.; Kato, N.; et al. The clinical relevance of plasma CD147/basigin in biopsy-proven kidney diseases. Clin. Exp. Nephrol. 2018, 22, 815–824. [Google Scholar] [CrossRef]
- Kashii, Y.; Giorda, R.; Heberman, R.B.; Whiteside, T.L.; Vujanovic, N.L. Constitutive expression and role of the TNF family ligands in apoptotic killing of tumor cells by human NK cells. J. Immunol. 1999, 163, 5358–5366. [Google Scholar]
- Lens, S.M.; Tesselaar, K.; van Oers, M.H.J.; van Lier, R.A.W. Control oflymphocyte function through CD27-CD70 interactions. Semin. Immunol. 1998, 10, 491–499. [Google Scholar] [CrossRef] [PubMed]
- Junker, K.; Hindermann, W.; von Eggeling, F.; Diegmann, J.; Haessler, K.; Schubert, J. CD70: A new tumor specific biomarker for renal cell carcinoma. J. Urol. 2005, 173, 2150–2153. [Google Scholar] [CrossRef] [PubMed]
- Agematsu, K.; Hokibara, S.; Nagumo, H.; Komiyama, A. CD27: Amemory B-cell marker. Immunol. Today 2000, 21, 204–206. [Google Scholar] [CrossRef]
- Becker, H.M. Carbonic anhydrase IX and acid transport in cancer. Br. J. Cancer 2020, 122, 157–167. [Google Scholar] [CrossRef]
- Luong-Player, A.; Liu, H.; Wang, H.L.; Lin, F. Immunohistochemical reevaluation of carbonic anhydrase IX (CA IX) expression in tumors and normal tissues. Am. J. Clin. Pathol. 2014, 141, 219–225. [Google Scholar] [CrossRef] [Green Version]
- Ingels, A.; Hew, M.; Algaba, F.; de Boer, O.J.; van Moorselaar, R.J.; Horenblas, S.; Zondervan, P.; de la Rosette, J.J.; Pilar Laguna Pes, M. Vimentin over-expression and carbonic anhydrase IX under-expression are independent predictors of recurrence, specific and overall survival in non-metastatic clear-cell renal carcinoma: A validation study. World J. Urol. 2017, 35, 81–87. [Google Scholar] [CrossRef] [Green Version]
- Soltysova, A.; Breza, J.; Takacova, M.; Feruszova, J.; Hudecova, S.; Novotna, B.; Rozborilova, E.; Pastorekova, S.; Kadasi, L.; Krizanova, O. Deregulation of energetic metabolism in the clear cell renal cell carcinoma: A multiple pathway analysis based on microarray profiling. Int. J. Oncol. 2015, 47, 287–295. [Google Scholar] [CrossRef]
- Peng, F.; Li, H.; You, Q.; Li, H.; Wu, D.; Jiang, C.; Deng, G.; Li, Y.; Li, Y.; Wu, Y. CD147 as a Novel Prognostic Biomarker for Hepatocellular Carcinoma: A Meta-Analysis. BioMed Res. Int. 2017, 2017, 5019367. [Google Scholar] [CrossRef]
- Landras, A.; Reger de Moura, C.; Jouenne, F.; Lebbe, C.; Menashi, S.; Mourah, S. CD147 Is a Promising Target of Tumor Progression and a Prognostic Biomarker. Cancers 2019, 11, 1803. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.; Su, J.; Zhou, Y.; Xie, X.; Peng, C.; Yuan, Z.; Chen, X. Repressing CD147 is a novel therapeutic strategy for malignant melanoma. Oncotarget 2017, 8, 25806–25813. [Google Scholar] [CrossRef] [Green Version]
- Sato, M.; Nakai, Y.; Nakata, W.; Yoshida, T.; Hatano, K.; Kawashima, A.; Fujita, K.; Uemura, M.; Takayama, H.; Nonomura, N. EMMPRIN promotes angiogenesis, proliferation, invasion and resistance to sunitinib in renal cell carcinoma, and its level predicts patient outcome. PLoS ONE 2013, 8, e74313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Witt, L. Etablierung eines Anreicherungsverfahrens von tumorassoziierten Exosomen aus Nierenzellkarzinomgeweben. Master’s Thesis, Saarland University, Saarbrücken, Germany, 2018. [Google Scholar]
- Tesselaar, K.; Xiao, Y.; Arens, R.; van Schijndel, G.M.W.; Schuurhuis, D.H.; Mebius, R.; Borst, J.; van Lier, R.A.W. Expression of the murineCD27 ligand CD70in vitroandin vivo. J. Immunol. 2003, 170, 33–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, Y.; Huang, W.; Ma, L.T.; Jiang, J.L.; Chen, Z.N. Importance of N-glycosylation on CD147 for its biological functions. Int. J. Mol. Sci. 2014, 15, 6356–6377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zieren, R.C.; Dong, L.; Pierorazio, P.M.; Pienta, K.J.; de Reijke, T.M.; Amend, S.R. Extracellular vesicle isolation from human renal cancer tissue. Med. Oncol. 2020, 37, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; He, X.; Deng, Y.; Yang, C. An Update on Isolation Methods for Proteomic Studies of Extracellular Vesicles in Biofluids. Molecules 2019, 24, 3516. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Kaslan, M.; Lee, S.H.; Yao, J.; Gao, Z. Progress in Exosome Isolation Techniques. Theranostics 2017, 7, 789–804. [Google Scholar] [CrossRef]
- Tai, Y.L.; Chen, K.C.; Hsieh, J.T.; Shen, T.L. Exosomes in cancer development and clinical applications. Cancer Sci. 2018, 109, 2364–2374. [Google Scholar] [CrossRef] [Green Version]
- Spizzo, G.; Went, P.; Dirnhofer, S.; Obrist, P.; Moch, H.; Baeuerle, P.A.; Mueller-Holzner, E.; Marth, C.; Gastl, G.; Zeimet, A.G. Overexpression of epithelial cell adhesion molecule (Ep-CAM) is an independent prognostic marker for reduced survival of patients with epithelial ovarian cancer. Gynecol. Oncol. 2006, 103, 483–488. [Google Scholar] [CrossRef]
- Benko, G.; Spajić, B.; Krušlin, B.; Tomas, D. Impact of the EpCAM expression on biochemical recurrence-free survival in clinically localized prostate cancer. Urol. Oncol. 2013, 31, 468–474. [Google Scholar] [CrossRef]
- Spizzo, G.; Went, P.; Dirnhofer, S.; Obrist, P.; Simon, R.; Spichtin, H.; Maurer, R.; Metzger, U.; von Castelberg, B.; Bart, R.; et al. High Ep-CAM expression is associated with poor prognosis in node-positive breast cancer. Breast Cancer Res. Treat. 2004, 86, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Trzpis, M.; Popa, E.R.; McLaughlin, P.M.; van Goor, H.; Timmer, A.; Bosman, G.W.; de Leij, L.M.F.H.; Harmsen, M.C. Spatial and temporal expression patterns of the epithelial cell adhesion molecule (EpCAM/EGP-2) in developing and adult kidneys. Nephron Exp. Nephrol. 2007, 107, e119–e131. [Google Scholar] [CrossRef] [PubMed]
- Gires, O. EGFR-Dependent Regulated Intramembrane Proteolysis of EpCAM—Letter. Cancer Res. 2017, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.J.; Feng, X.; Lu, M.; Morimura, S.; Udey, M.C. Matriptase-mediated cleavage of EpCAM destabilizes claudins and dysregulates intestinal epithelial homeostasis. J. Clin. Investig. 2017, 127, 623–634. [Google Scholar] [CrossRef] [PubMed]
- Denzel, S.; Maetzel, D.; Mack, B.; Eggert, C.; Bärr, G.; Gires, O. Initial activation of EpCAM cleavage via cell-to-cell contact. BMC Cancer 2009, 9, 402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seligson, D.B.; Pantuck, A.J.; Liu, X.; Huang, Y.; Horvath, S.; Bui, M.H.T.; Han, K.-R.; Correa, A.J.L.; Eeva, M.; Tze, S.; et al. Epithelial cell adhesion molecule (KSA) expression: Pathobiology and its role as an independent predictor of survival in renal cell carcinoma. Clin. Cancer Res. 2004, 10, 2659–2669. [Google Scholar] [CrossRef] [Green Version]
- Iacono, K.T.; Brown, A.L.; Greene, M.I.; Saouaf, S.J. CD147 immunoglobulin superfamily receptor function and role in pathology. Exp. Mol. Pathol. 2007, 83, 283–295. [Google Scholar] [CrossRef] [Green Version]
- Tsai, W.-C.; Chao, Y.-C.; Lee, W.-H.; Chen, A.; Sheu, L.-F.; Jin, J.-S. Increasing EMMPRIN and matriptase expression in hepatocellular carcinoma: Tissue microarray analysis of immunohistochemical scores with clinicopathological parameters. Histopathology 2006, 49, 388–395. [Google Scholar] [CrossRef]
- Tsai, W.C.; Sheu, L.F.; Nieh, S.; Yu, C.-P.; Sun, G.-H.; Lin, Y.-F.; Chen, A.; Jin, J.-S. Association of EMMPRIN and fascin expression in renal cell carcinoma: Correlation with clinicopathological parameters. World J. Urol. 2007, 25, 73–80. [Google Scholar] [CrossRef]
- Du, Z.M.; Hu, C.F.; Shao, Q.; Huang, M.-Y.; Kou, C.-W.; Zhu, X.-F.; Zeng, Y.-X.; Shao, J.-Y. Upregulation of caveolin-1 and CD147 expression in nasopharyngeal carcinoma enhanced tumor cell migration and correlated with poor prognosis of the patients. Int. J. Cancer 2009, 125, 1832–1841. [Google Scholar] [CrossRef]
- Pértega-Gomes, N.; Vizcaíno, J.R.; Miranda-Gonçalves, V.; Pinheiro, C.; Silva, J.; Pereira, H.; Monteiro, P.; Henrique, R.M.; Reis, R.M.; Lopes, C.; et al. Monocarboxylate transporter 4 (MCT4) and CD147 overexpression is associated with poor prognosis in prostate cancer. BMC Cancer 2011, 11, 312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, X.; Wang, C.; Zhang, J.; Qie, G.; Zhou, J. The roles of CD147 and/or cyclophilin A in kidney diseases. Mediat. Inflamm. 2014, 2014, 728673. [Google Scholar] [CrossRef] [PubMed]
- Fisel, P.; Stühler, V.; Bedke, J.; Winter, S.; Rausch, S.; Hennenlotter, J.; Nies, A.T.; Stenzl, A.; Scharpf, M.; Fend, F.; et al. MCT4 surpasses the prognostic relevance of the ancillary protein CD147 in clear cell renal cell carcinoma. Oncotarget 2015, 6, 30615–30627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Z.D.; He, H.C.; Bi, X.C.; Qin, W.-J.; Dai, Q.-S.; Zou, J.; Ye, Y.-K.; Liang, Y.-X.; Zeng, G.-H.; Zhu, G.; et al. Expression and clinical significance of CD147 in genitourinary carcinomas. J. Surg. Res. 2010, 160, 260–267. [Google Scholar] [CrossRef] [PubMed]
- Baniak, N.; Flood, T.A.; Buchanan, M.; Dal Cin, P.; Hirsch, M.S. Carbonic Anhydrase IX (CA9) Expression in Multiple Renal Epithelial Tumor Subtypes. Histopathology 2020. [Google Scholar] [CrossRef] [PubMed]
- Purkerson, J.M.; Schwartz, G.J. The role of carbonic anhydrases in renal physiology. Kidney Int. 2007, 71, 103–115. [Google Scholar] [CrossRef] [Green Version]
- Tostain, J.; Li, G.; Gentil-Perret, A.; Gigante, M. Carbonic anhydrase 9 in clear cell renal cell carcinoma: A marker for diagnosis, prognosis and treatment. Eur. J. Cancer 2010, 46, 3141–3148. [Google Scholar] [CrossRef]
- Soyupak, B.; Erdoğan, S.; Ergin, M.; Seydaoğlu, G.; Kuzgunbay, B.; Tansuğ, Z. CA9 expression as a prognostic factor in renal clear cell carcinoma. Urol. Int. 2005, 74, 68–73. [Google Scholar] [CrossRef]
- Stillebroer, A.B.; Mulders, P.F.; Boerman, O.C.; Oyen, W.J.; Oosterwijk, E. Carbonic anhydrase IX in renal cell carcinoma: Implications for prognosis, diagnosis, and therapy. Eur. Urol. 2010, 58, 75–83. [Google Scholar] [CrossRef]
- Jilaveanu, L.B.; Sznol, J.; Aziz, S.A.; Duchen, D.; Kluger, H.M.; Camp, R.L. CD70 expression patterns in renal cell carcinoma. Hum. Pathol. 2012, 43, 1394–1399. [Google Scholar] [CrossRef] [Green Version]
- Bodmer, J.L.; Schneider, P.; Tschopp, J. The molecular architecture of the TNF superfamily. Trends Biochem. Sci. 2002, 27, 19–26. [Google Scholar] [CrossRef] [Green Version]
- Locksley, R.M.; Killeen, N.; Lenardo, M.J. The TNF and TNF receptor superfamilies: Integrating mammalian biology. Cell 2001, 104, 487–501. [Google Scholar] [CrossRef] [Green Version]
- Vella, L.J.; Scicluna, B.J.; Cheng, L.; Bawden, E.G.; Masters, C.L.; Ang, C.-S.; Willamson, N.; McLean, C.; Barnham, K.J.; Hill, A.F. A rigorous method to enrich for exosomes from brain tissue. J. Extracell. Vesicles 2017, 6, 1348885. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Origin | CA9 | CD147 | CD70 | EpCAM |
|---|---|---|---|---|
| cells | 6/6 | 8/8 | 8/8 | 5/7 |
| exosomes | 7/7 | 8/8 | 5/8 | 5/8 |
| Case No. | Cells | Exosomes | TNM | Grade | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| CA9 | CD147 | CD70 | EpCAM | CA9 | CD147 | CD70 | EpCAM | |||
| NTB1126 | n.e. | + | + | n.e. | +++ | + | - | - | pT3acN0cM0 | unknown |
| NTB1302 | +++ | ++ | + | n.e. | +++ | +++ | - | n.e. | pT3acN0cM0 | G3 |
| NTB1161 | n.e. | + | + | n.e. | n.e. | + | - | n.e. | pT1bcN0cM0 | G2 |
| NTB1208 | + | + | + | + | ++ | + | ++ | + | pT4cN0cM1 | G3 |
| NTB1175 | ++ | + | + | - | +++ | ++ | ++ | - | pT3acN0cM0 | G3 |
| NTB1288 | ++ | + | + | - | ++ | + | +++ | - | pT3acN0M0 | G3 |
| NTB1304 | n.e. | n.e. | n.e. | ++ | n.e. | n.e. | n.e. | ++ | pT3bpN0M0 | G2 |
| NTB1132 | n.e. | n.e. | n.e. | + | n.e. | n.e. | n.e. | +++ | pT1bcN0cM0 | G2 |
| NTB1197 | ++ | + | + | + | +++ | +++ | +++ | ++ | pT1bcN0cM0 | G2 |
| NTB1272 | +++ | + | + | ++ | +++ | ++ | ++ | + | pT1bcN0cM0 | G2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Himbert, D.; Zeuschner, P.; Ayoubian, H.; Heinzelmann, J.; Stöckle, M.; Junker, K. Characterization of CD147, CA9, and CD70 as Tumor-Specific Markers on Extracellular Vesicles in Clear Cell Renal Cell Carcinoma. Diagnostics 2020, 10, 1034. https://doi.org/10.3390/diagnostics10121034
Himbert D, Zeuschner P, Ayoubian H, Heinzelmann J, Stöckle M, Junker K. Characterization of CD147, CA9, and CD70 as Tumor-Specific Markers on Extracellular Vesicles in Clear Cell Renal Cell Carcinoma. Diagnostics. 2020; 10(12):1034. https://doi.org/10.3390/diagnostics10121034
Chicago/Turabian StyleHimbert, Dirk, Philip Zeuschner, Hiresh Ayoubian, Joana Heinzelmann, Michael Stöckle, and Kerstin Junker. 2020. "Characterization of CD147, CA9, and CD70 as Tumor-Specific Markers on Extracellular Vesicles in Clear Cell Renal Cell Carcinoma" Diagnostics 10, no. 12: 1034. https://doi.org/10.3390/diagnostics10121034