Local and Systemic Interleukin-32 in Esophageal, Gastric, and Colorectal Cancers: Clinical and Diagnostic Significance


1
Department of Gastrointestinal and General Surgery, Wroclaw Medical University, 50-368 Wroclaw, Poland
2
Department of Nervous System Diseases, Wroclaw Medical University, 51-618 Wroclaw, Poland
3
Department of Medical Biochemistry, Wroclaw Medical University, 50-368 Wroclaw, Poland
*
Author to whom correspondence should be addressed.
†
Both authors contributed equally.
Diagnostics 2020, 10(10), 785; https://doi.org/10.3390/diagnostics10100785
Submission received: 17 September 2020
/
Revised: 1 October 2020
/
Accepted: 2 October 2020
/
Published: 4 October 2020
(This article belongs to the Section Pathology and Molecular Diagnostics)
Abstract
:Little is known on clinical and diagnostic relevance of interleukin-32 in gastrointestinal tract (GIT) cancers. We determined its mRNA (n = 52) and protein (n = 63) expression in paired (tumor-normal) samples from esophageal squamous cell carcinoma (ESCC) and gastric (GC) and colorectal cancer (CRC) patients, with reference to cancer-associated genes, and quantified circulating interleukin-32 in 70 cancer patients and 28 controls. IL32 expression was significantly upregulated solely in ESCC, reflecting T stage in non-transformed tumor-adjacent tissue. Fold-change in IL32 and IL-32 was higher in left-sided CRC, owing to high interleukin expression in non-transformed right-sided colonic mucosa. IL32 was independently and positively associated with Ki67, HIF1A, and ACTA2 and negatively with TJP1 in tumors and with IL10Ra and BCLxL in non-transformed tumor-adjacent tissue. IL-32 protein was significantly upregulated in colorectal tumors. In ESCC, advanced stage and lymph node metastasis were associated with significant IL-32 upregulation. Circulating interleukin was significantly elevated in cancer patients, more so in ESCC and GC than CRC. As biomarker, IL-32 detected gastroesophageal cancers with 99.5% accuracy. In conclusion, IL-32 is upregulated in GIT cancers at local and systemic level, reflecting hypoxia and proliferative and invasive/metastatic capacity in tumors and immunosuppressive and antiapoptotic potential in non-transformed mucosa, while being an accurate biomarker of gastroesophageal cancers.
1. Introduction
Gastrointestinal tract (GIT) cancers, encompassing adenocarcinomas of the colorectum (CRC) and stomach (GC) and esophageal squamous cell carcinoma (ESCC), are the most common and deadliest malignancies [1]. Radical surgery, alone or in combination with chemotherapy or radiation, remains the main therapeutic option. Mortality rates are tightly associated with cancer stage at presentation. However, the GIT cancers, ESCC and GC in particular, are frequently diagnosed when advanced, rendering them non-amenable for curative resection. Available therapeutic options fail to improve outcomes for patients with gross metastatic disease and cancers resistant to chemo- and radiotherapy [2,3,4,5]. There is a growing awareness that in order to improve prognosis, a better understanding of cancer-associated abnormalities at molecular level is urgently needed to facilitate biomarker discovery and develop and implement patient-tailored approach, referred to as “precision medicine” [6,7,8,9].
Chronic inflammation with accompanying oxidative stress plays a crucial role in initiation of neoplastic transformation in GC and CRC [3,10]. Infection with H. pylori is a main risk factor in non-cardia GC, accounting for up-to 90% of cases. Other GC risk factors include infection with Epstein–Barr virus or autoimmune gastritis and thus are inflammation-related as well. Global attempt for H. pylori eradication managed to significantly reduce incidence of this subtype. However, it also contributed to substantial increase in cardia GC incidence. Cardia GC may have common etiology with non-cardia subtype or be associated with obesity and gastroesophageal reflux, conditions of persistent low-grade inflammation and oxidative stress [3]. Crohn’s disease and ulcerative colitis are two main phenotypes of inflammatory bowel disease (IBD), chronic relapsing condition, currently incurable, associated with increased risk for CRC [10]. While esophageal adenocarcinoma is a typical inflammation-associated cancer [11], the contribution of chronic inflammation to ESCC is more subtle. Overuse of strong alcohol and prolonged exposure to tobacco smoke, irritants and carcinogens inducing oxidative and genotoxic stress and evoking inflammation, are dominant risk factors only in some regions. Disturbed oral microbiome, infections with human papillomavirus, and improper diet—low on antioxidants and contaminated with nitrosamines or mycotoxins—are other increasingly recognized and inflammation- and oxidative stress-associated risk factors [4]. Noteworthy, in addition to its role in cancer initiation, inflammatory milieu supports tumor growth by providing mitogens and pro-survival cues and allows cancer cells to evade immune system and disseminate [12].
Interleukin (IL)-32 is a relatively recently discovered cytokine of potent pro-inflammatory activity. It is expressed in natural killer cells, monocytes, lymphocytes T, peripheral blood mononuclear cells, epithelial and endothelial cells, and fibroblasts in a number of isoforms with varying biological activity. Its expression is triggered by IL-1β, IL-18, tumor necrosis factor (TNF)-α, and interferon (IFN)-γ. IL-32 is engaged in setting an inflammatory loop as it in turn induces the synthesis of IL-1β, TNFα, IL-6, IL-8, and macrophage inflammatory protein (MIP)-2 [13,14,15]. Consistently, the involvement of IL-32 has been documented in infectious diseases, chronic inflammatory conditions, including gastritis and inflammatory bowel disease, and in cancer [13,14,15,16,17,18]. Amassing evidence indicates that the biological activity of IL-32 is cell type- and context-depended and displays isoform-specific nuances. Consequently, it may either facilitate or hamper cancer development, gaining IL-32 a catching label of “frenemy in cancer” [15]. Our view on the interleukin shifted from a simple inflammation amplifier to a modulator of inflammatory response and cell fate. Importantly, IL-32, as well as mechanisms employed in regulating formation of its endogenous isoforms, is considered a potential target for anti-neoplastic strategy [13,14,15,16].
Limited reports regarding GIT cancers show that the interleukin aids invasiveness of gastric cancer [19] but may act as a tumor suppressor in the colon [20]. In view of existing controversies and growing interest in the cytokine as potential anti-neoplastic target, the relative scarcity of data is surprising. Therefore, we aimed at comparative analysis of its local expression patterns in ESCC, GC, and CRC and at an appraisal of diagnostic power of circulating IL-32. The interleukin in the present study was quantified at mRNA and protein level and referred to cancer pathology and the local expression of a panel of cancer-associated genes (IL4, IL4Ra, IL7, IL7Ra, IL10, IL10Ra, IL13, IL13Ra, ACTA2, BCL2, BCLxL, CCL2, CDKN1A, CLDN2, SLC2A1, HIF1A, Ki67, NOS2, ODC1, PTGS2, TJP1, and VEGFA) as well as circulating cytokines and growth factors (IL-1β, IL-4, IL-6, IL-8, IL-12p70, fibroblast growth factor (FGF)-2, granulocyte colony-stimulating factor (G-CSF), granulocyte-macrophage colony-stimulating factor (GM-CSF), monocyte chemoattractant protein (MCP)-1, MIP-1α, platelet-derived growth factor (PDGF)-BB, TNFα, and vascular endothelial growth factor (VEGF)-A).
2. Materials and Methods
2.1. Study Population
Study population consisted of 100 individuals: 28 controls and 72 cancer patients with histologically confirmed esophageal squamous cell carcinoma (n = 17), gastric adenocarcinoma (n = 14), or colorectal adenocarcinoma (n = 41). Details are given in Table 1. Cancer patients were admitted to the Department of Gastrointestinal and General Surgery (Wroclaw Medical University) for curative tumor resection. Patients with any severe systemic illness or gross metastatic disease non-amenable for curative resection or subjected to previous radio- or chemotherapy were excluded. Enrolled patients underwent standard preoperative evaluation consisting of blood work, physical examination, and imaging (ultrasonography, computed tomography and magnetic resonance). Cancers were rated pathologically using 7th edition of the Union for International Cancer Control TNM system. In all examined cases, the resection margins were cancer-free. Serum samples from apparently healthy individuals were obtained from blood donors from the Regional Center of Blood Donation and Therapeutics in Wroclaw, Poland. Information on sample availability for transcriptional analysis and determination of local and systemic IL-32 protein concentration is given in Results in respective subsections.
2.2. Ethical Considerations
The study gained the acceptance of the Medical Ethics Committee of Wroclaw Medical University (#KB 203/2016 from 21 April 2016). It was conducted in accordance with the Helsinki Declaration of 1975, as revised in 1983, and informed consent was obtained from all study participants.
2.3. Analytical Methods
2.3.1. IL32 Expression in Tissue Samples
Pairs of tissue samples from the tumor and from the macroscopically normal tissue adjacent to the tumor (taken approximately 10 cm from the tumor) were taken postoperatively and rinsed with PBS. Samples were then immersed in RNAlater purchased from Ambion Inc. (Austin, TX, USA). Solution-soaked tissue samples were then stored at −80 °C until RNA isolation.
Tissue samples of 30–40 mg were homogenized in FastPrep-24 Homogenizer from MP Biomedical (Solon, OH, USA) using lysis buffer and 2-mercaptoethanol (100:1, v/v) from Sigma-Aldrich (St. Luis, MO, USA). The RNA was isolated using phenol-chloroform extraction and then purified with PureLink™ RNA Mini Kit (Invitrogen, Carlsbad, CA, USA). Genomic DNA was removed by on-column incubation with DNase (PureLink™ DNase Set, Invitrogen). The RNA isolates were quantified using NanoDrop 2000 from Thermo-Fisher Scientific (Waltham, MA, USA). Purity of isolated RNA was evaluated using 260/280 nm and 260/230 nm absorbance ratios. Its integrity was determined using Experion RNA StdSens analysis kits and the Experion platform, employing LabChip microfluidic technology (BioRad, Hercules, CA, USA). Aliquots of RNA isolates corresponding with 1000 ng per reaction mixture (20 µL) were reversely transcribed using iScript™ cDNA Synthesis Kit (BioRad) and C1000 termocycler (BioRad). reaction conditions were set as suggested by the manufacturer. Quantitative (real-time) polymerase chain reaction (qPCR) was conducted using SsoFast EvaGreen® Supermix (BioRad) and CFX96 Real-Time PCR thermocycler (BioRad). The following cycling conditions were applied: 30 s activation at 95 °C, 5 s denaturation at 95 °C, annealing/extension for 5 s at 61 °C, 45 cycles, followed by melting step (60–95 °C with fluorescent reading every 0.5°C) to assure product specificity, further confirmed in an electrophoresis in high-resolution agarose (SeaKem LE agarose from Lonza, Basel, Switzerland) in TBE with SYBR Green (Lonza) detection. Reaction mixture consisted of cDNA (2 µL; diluted 1:5), 2×SsoFast EvaGreen® Supermix (10 µL), 10 nM forward and reverse target-specific primers (1 µL of each), and water up to 20 µL. The following primer sequences were used: 5′-TCAAAGAGGGCTACCTGGAGAC-3′ (IL32, forward); 5′-TCTGTTGCCTCGGCACCGTAAT-3′ (IL32, reverse); 5′-TAGATTATTCTCTGATTTGGTCGTATTGG-3′ (GAPDH, forward); 5′-GCTCCTGGAAGATGGTGATGG-3′ (GAPDH, reverse). Primer sequences were synthesized by Genomed (Warsaw, Poland). Technical replicates were averaged prior analysis. Geometric mean of all Cq values in a given analysis was obtained and subtracted from sample Cq (ΔCq) then linearized by 2^ΔCq conversion and normalized to GAPDH (internal control). The obtained values were referred to as a normalized relative quantity (NRQ) [21] and subjected to statistical analysis.
Data on relative expression of IL4, IL4Ra, IL7, IL7Ra, IL10, IL10Ra, IL13, IL13Ra, ACTA2, BCL2, BCLxL, CCL2, CDKN1A, CLDN2, SLC2A1, HIF1A, Ki67, NOS2, ODC1, PTGS2, TJP1, and VEGFA in tissue samples investigated were available for 45 cancer patients and were retrieved from our earlier studies [22,23] for the purpose of correlation analysis.
2.3.2. IL-32 Concentration in Tissue Homogenates
Pairs of tissue samples from the tumor and from the macroscopically normal tissue adjacent to the tumor (taken approximately 10 cm from the tumor) were taken postoperatively and rinsed with PBS. Samples were then rapidly frozen and stored at –45 °C until analysis.
Tissue fragments of 10–40 mg were placed in 10 mM Tris-HCl buffer with addition of 150 mM KCl and 1 mM EDTA (pH 7.4) in proportion 1:2 (w/v) and homogenized with ceramic spheres for 2 min at 4.0 m/s in FastPrep-24 homogenizer from MP Biomedical (Solon, OH, USA). Resulting homogenates were centrifuged (14,500× g, 10 min, 6 °C). Supernatants collected and used for IL-32 quantification using Human Interleukin 32 ELISA Kits from MyBiosource, Inc. (San Diego, CA, USA) according to manufacturer’s instructions. The assays were performed in duplicates and absorbance was measured using Microplate Reader BioTek ELx800 TS and Gen5 program (BioTek Instrument Inc., Winooski, VT, USA). Technical replicates were averaged and normalized to tissue weight and results are expressed as ng of protein per gram of analyzed tissue [ng/g].
2.3.3. IL-32 Concentration in Serum Samples
Peripheral blood was drawn into BD Vacutainer CAT tubes (Becton Dickinson, Plymouth, UK) and clotted at room temperature for 30 min. Subsequently, samples were centrifuged (1500× g for 10 min at room temperature). Resulting serum samples were aliquoted and stored at −45 °C until examination. Blood was drawn upon patient’s admission, prior to any treatment and following overnight fast. For IL-32 quantification the same immunoassays as described above were used. Results are expressed as pg of interleukin per milliliter of serum [pg/mL].
Data on serum concentration of IL-1β, IL-4, IL-6, IL-8, IL-12p70, FGF2, G-CSF, GM-CSF, MCP-1, MIP-1α, PDGF-BB, TNFα, and VEGF-A, determined using Luminex xMAP technology, were available for 43 cancer patients and were retrieved from our earlier studies [24] for the purpose of correlation analysis.
2.4. Statistical Analysis
Data were tested for homogeneity of variances and normality of distribution using Levene test and Kolmogorov-Smirnov test, respectively. Pair-wise analysis was conducted using t-test for paired samples on log-transformed data (transcriptional analysis) or Wilcoxon test (protein determination). Two-group comparisons were conducted using t-test for independent samples, with Welch correction in case of unequal variances, or Mann–Whitney U test. Multigroup comparisons were conducted using one-way ANOVA with Tukey–Kramer post-hoc test or Kruskal–Wallis H test with Conover post-hoc test. Data were presented as geometric means with 95% confidence interval or medians. Correlation analysis was conducted using Spearman’s rank correlation test (ρ) or Pearson correlation (r). Frequency analysis was conducted using χ2 test. Receiver operating characteristics (ROC) curve analysis was conducted to determine the diagnostic power of IL-32. The overall marker accuracy was expressed as area under the ROC curve (AUC). In addition, marker sensitivity and specificity at optimal cut-off value were calculated. Multiple regression (backward and forward stepwise method) was applied to identify independent predictors of IL32 expression. Their correlation with IL32 after the influence of the remaining variables is eliminated is presented as partial correlation coefficients (rp).
All calculated probabilities were two-tailed. The p values ≤0.05 were considered statistically significant. The entire analysis was conducted using MedCalc Statistical Software version 19.4.0 (MedCalc Software Ltd., Ostend, Belgium; https://www.medcalc.org; 2020).
3. Results
3.1. Local IL-32 Expression at mRNA Level
Relative IL32 expression was determined in patient-matched samples from tumor and non-transformed mucosa adjacent to tumor (52 pairs), obtained from 17 patients with ESCC, 14 with GC, and 21 with CRC using reverse-transcribed quantitative (real-time) polymerase chain reaction (RT-qPCR).
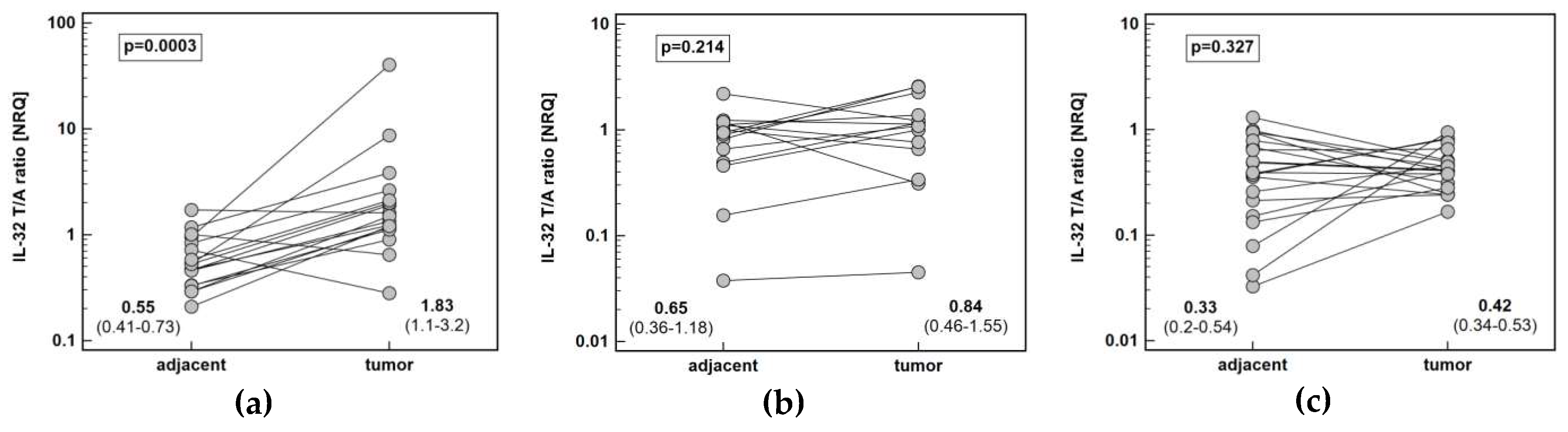
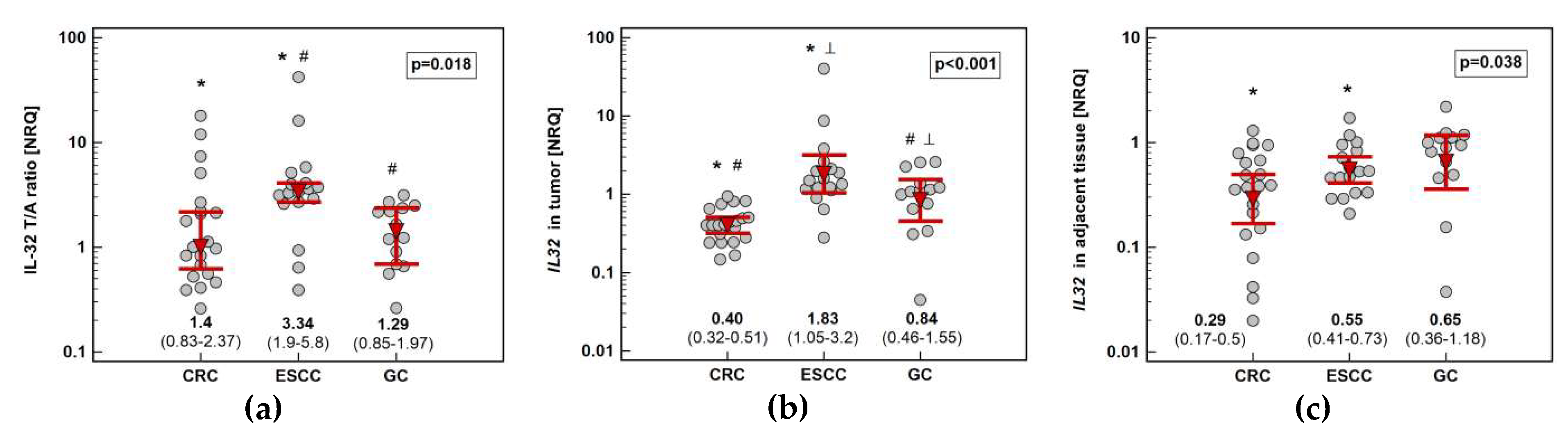
Pairwise analysis showed that IL32 expression is significantly upregulated (by 3.3-fold) solely in esophageal tumors (Figure 1). Tumor expression of IL32 was the highest in ESCC and the lowest in CRC. Esophageal tumors had higher interleukin expression by 2.2-fold as compared to gastric tumors and by 4.6-fold as compared to colorectal tumors. Gastric tumors had IL32 expression higher than colorectal tumors by 2.1-fold. In the non-cancerous tumor-adjacent tissue, IL32 expression was significantly higher in esophageal than colorectal mucosa (by 2.4-fold) and tended to be higher in gastric as compared to colorectal mucosa (by 2.3-fold). Consequently, fold-change in expression (tumor-to-adjacent) was comparable in CRC and GC and significantly lower than in ESCC by 2.4 and 2.6-fold, respectively (Figure 2).
There was no significant association between fold-change in IL32 expression and cancer overall TNM stage or its individual components. Separate analysis conducted in tumors and non-cancerous adjacent tissue showed that IL32 expression significantly increased along with growing depth of tumor invasion (T stage) in ESCC patients in adjacent tissue but not in tumors: (ρ = 0.51, p = 0.036).
Cardia subtype of GC (n = 5) tended to have higher expression of IL32 in non-cancerous adjacent tissue than non-cardia GC (n = 9) by 2.1-fold (p = 0.115), while the transcript abundance in tumors was almost identical (p = 0.885). Consequently, fold-change in IL32 expression tended to be higher in non-cardia GC by 1.9-fold (p = 0.124).
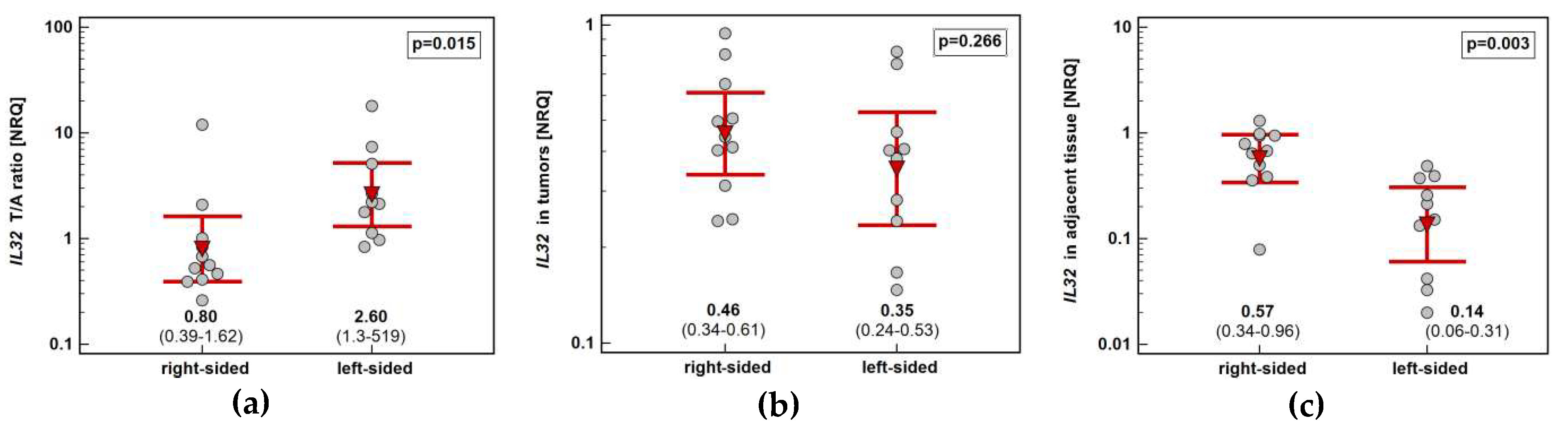
Tumor location in the left side of the colon (n = 10) was associated with higher IL32 upregulation (by 3.3-fold) than in the right side (n = 11). It resulted from significantly higher interleukin expression in non-cancerous tumor-adjacent mucosa from right than left side of the colon (by 4.1-fold) (Figure 3).
IL-32 has been implicated in modulating inflammation, immunity, proliferation, survival, angiogenesis, and epithelial-mesenchymal transition [15]. Therefore, the pattern of correlation between IL32 expression and a broad spectrum of genes encoding representative proteins implicated in cancer development, namely, IL4, IL4Ra, IL7, IL7Ra, IL10, IL10Ra, IL13, IL13Ra, ACTA2, BCL2, BCLxL, CCL2, CDKN1A, CLDN2, SLC2A1, HIF1A, Ki67, NOS2, ODC1, PTGS2, TJP1, and VEGFA, was examined in 45 cancer patients.
In tumor tissue, IL32 correlated positively with ACTA2, BCL2, BCLxL, CCL2, HIF1A, IL13Ra, IL7, Ki67, ODC1, PTGS2, SLC2A1, and TJP1. In non-cancerous tumor-adjacent tissue, IL32 correlated positively with BCL2, BCLxL, CCL2, CDKN1A, HIF1A, IL10Ra, IL13Ra, IL7Ra, Ki67, ODC1, SLC2A1, TJP1, and VEGFA. Fold-change in IL32 expression (tumor-to-adjacent) positively correlated with fold-change in expression of BCLxL, CCL2, CDKN1A, HIF1A, IL7, IL7Ra, Ki67, SLC2A1, and VEGFA (Table 2).
Multiple regression with genes significantly correlated with IL32 in univariate analysis as explanatory variables showed ACTA2 (rp = 0.33, p = 0.043), HIF1A (rp = 0.49, p = 0.002), Ki67 (rp = 0.46, p = 0.003), and TJP1 (rp = −0.48, p = 0.002) to be independent predictors of variation in IL32 expression in tumors.
In adjacent tissue, of the genes associated with IL32 expression in univariate analysis, IL10Ra (partial correlation coefficient: rp = 0.59, p < 0.0001) and SLC2A1 (rp = 0.32, p = 0.041) were independently associated with the interleukin expression. Comparably well-fit regression model could be built with BCLxL (rp = 0.64, p < 0.0001) instead of SLC2A1.
In turn, fold-change in IL32 expression was independently associated with fold-change in expression of IL7Ra (rp = 0.40, p = 0.008) and SLC2A1 (rp = 0.56, p < 0.001) or fold-change in expression of CDKN1A (rp = 0.49, p = 0.001) and IL7 (rp = 0.43, p = 0.004), depending on regression model applied.
3.2. Local IL-32 Protein Concentration
The concentration of IL-32 (protein) was determined in homogenates of patient-matched tumor and non-cancerous tumor-adjacent tissue (63 pairs) from 17 ESCC patients, 12 GC patients and 34 CRC patients using dedicated immunoassays.
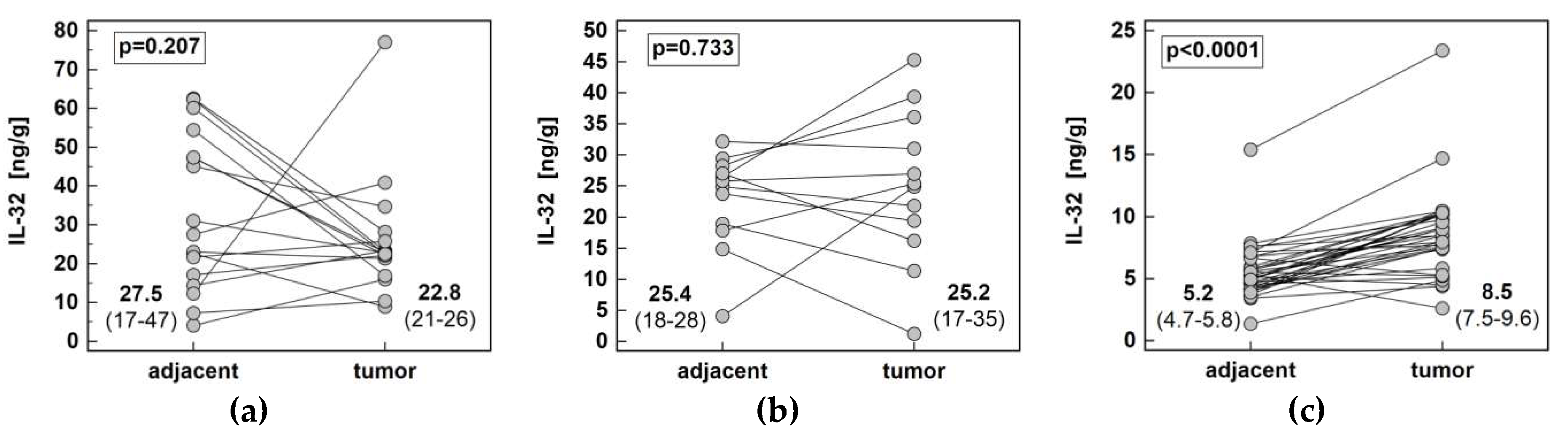
The interleukin concentration was significantly higher in tumor than adjacent tissue in CRC, showed no difference in GC and tended to be lower in ESCC (Figure 4). The IL-32 concentration in tumor (p < 0.00001) and adjacent tissue (p < 0.00001) as well as fold-change in concentration (tumor-to-adjacent) were significantly lower in CRC as compared to GC and ESCC (Figure 5).
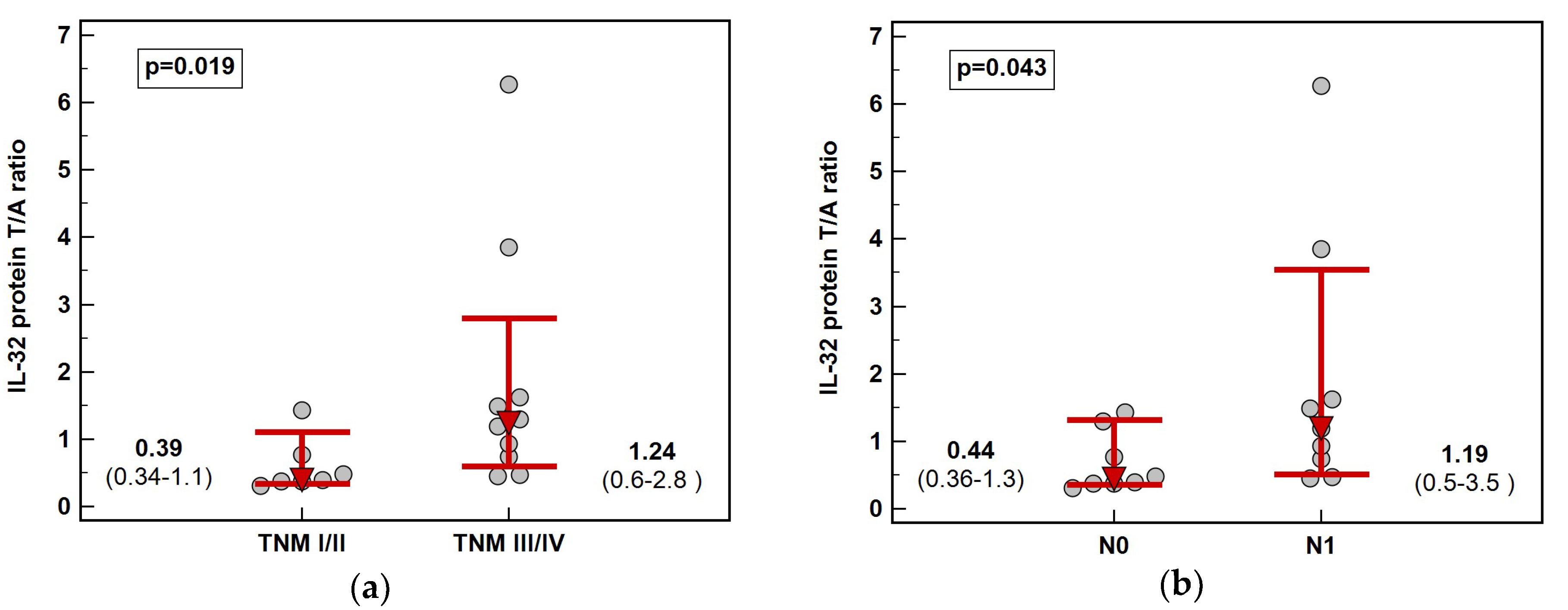
Solely in ESCC, fold-change in IL-32 protein concentration (tumor-to-adjacent) was dependent on cancer stage. The interleukin was downregulated in tumors in early (TNM I and II) and slightly upregulated in advanced (TNM III and IV) cancers and the difference in fold-change was 3.2-fold (Figure 6). Significant difference was observed between N0 and N1 cancers (Figure 2) while the difference between T1/2 and T3/4 did not reach statistical significance (respectively, 0.4 and 1.2, p = 0.131).
Tumors of gastric cardia (n = 5) tended to have higher concentration of IL-32 than non-cardia tumors (n = 7) by 2.0-fold (p = 0.169), accompanied by slightly lower abundance in non-adjacent tissue (by 1.3-fold, p = 0.565). Fold-change in IL-32 protein concentration tended to be higher in cardia GC by 2.6-fold (p = 0.099).
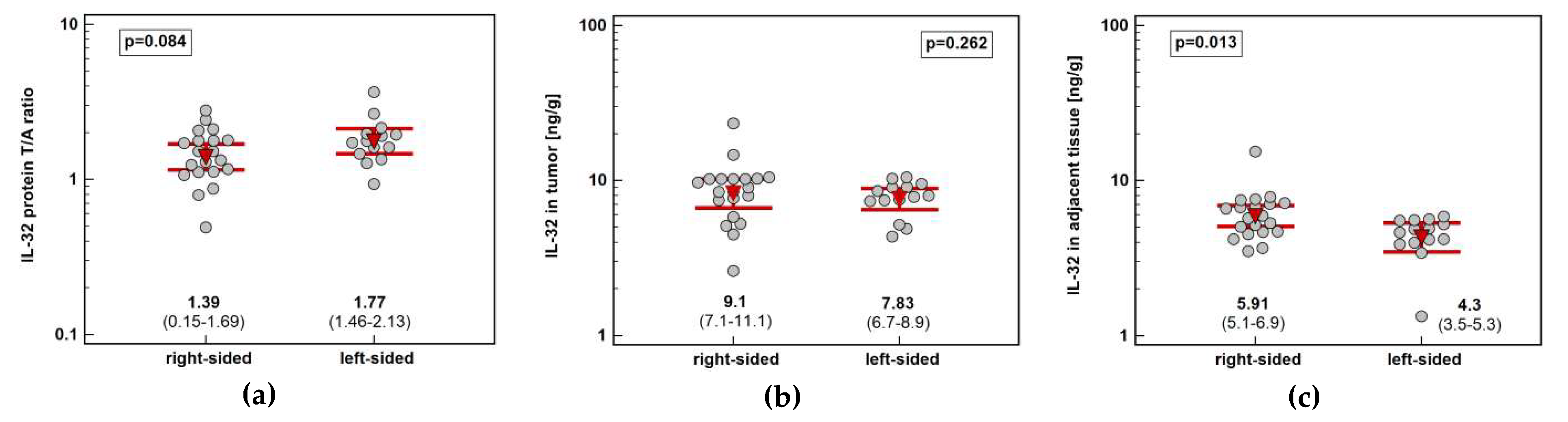
Tumor location in the left side of the colon (n = 14) tended to be associated with greater IL-32 protein concentration than in the right side (n = 20) (by 1.3-fold). It resulted from significantly higher interleukin expression in non-cancerous tumor-adjacent mucosa from right than left side of the colon (by 1.4-fold) (Figure 7).
3.3. Systemic IL-32 Protein Concentration
The concentration of IL-32 (protein) was determined in serum samples from 17 ESCC patients, 12 GC patients and 41 CRC patients using dedicated immunoassays.
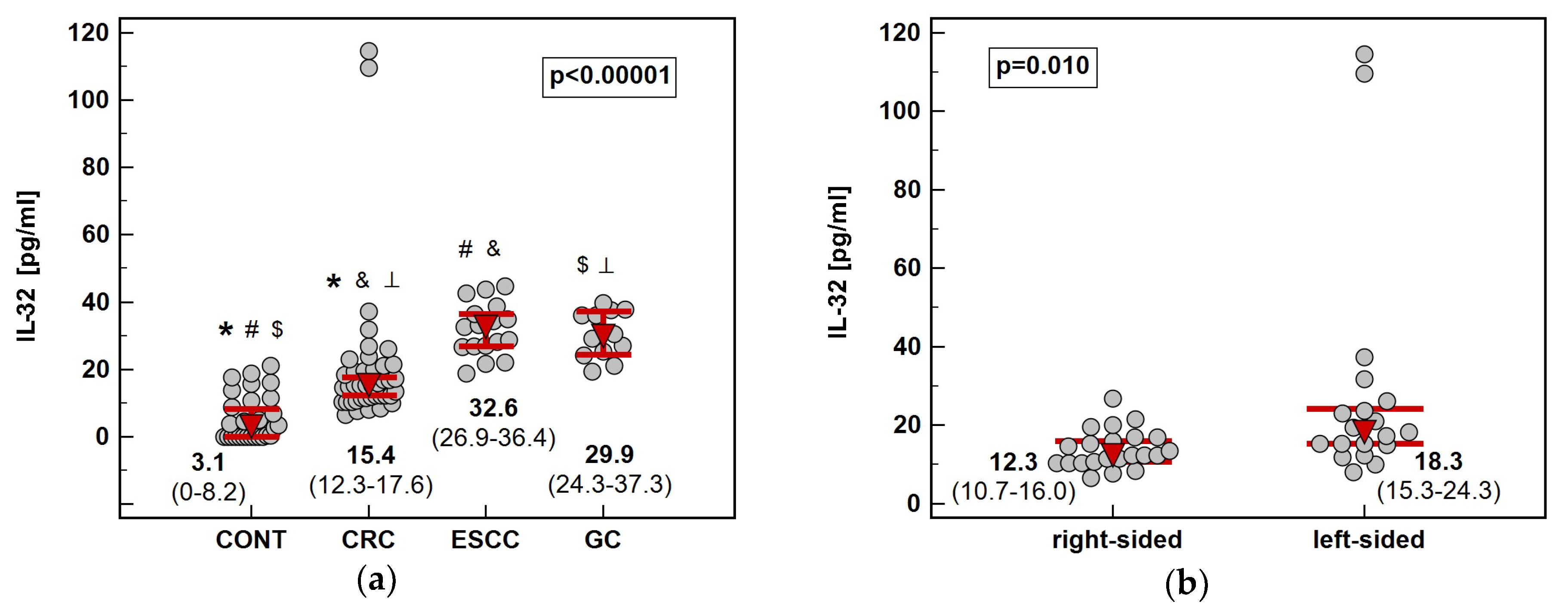
Cancer patients, regardless location, had significantly elevated circulating IL-32 as compared to healthy controls. Among cancers, CRC was accompanied by significantly lower IL-32 serum concentration than ESCC and GC. Tumor sublocation in the colon had an impact as well ‒ patients with left-sided tumors (n = 19) had significantly more elevated IL-32 than those with right-sided tumors (n = 22) (Figure 8). Regarding GC, cardia subtype (n = 5) was accompanied by insignificantly higher systemic IL-32 than non-cardia subtype (n = 7) (36 pg/mL vs. 29 pg/mL, p = 0.515).
Cancer stage had no significant effect on serum IL-32 in any cancer type.
Solely in GC, there was positive correlation between serum IL-32 and the local interleukin (protein) upregulation (r = 0.66, p = 0.020).
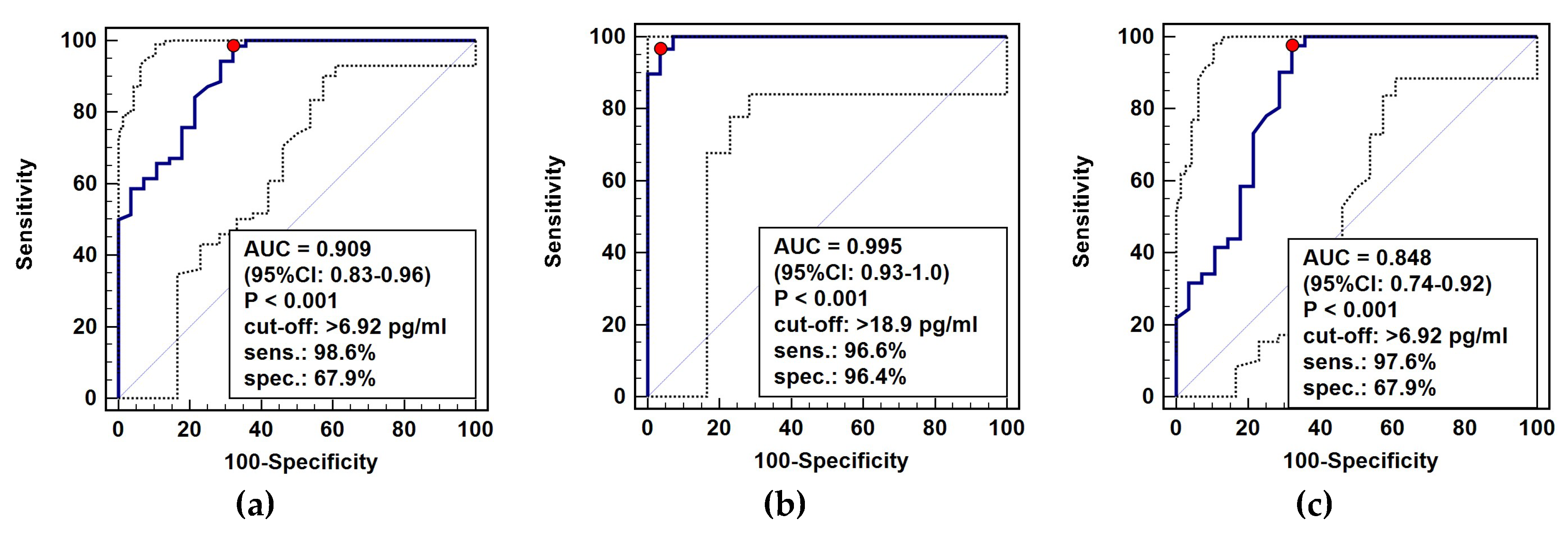
The diagnostic power of circulating IL-32 in distinguishing cancer patients from healthy individuals was tested using receiver operating characteristic (ROC) curve analysis. As general cancer marker (all cancer patients analyzed against healthy controls), IL-32 was characterized by excellent accuracy and sensitivity but only fair specificity (Figure 9a). As a marker of gastroesophageal cancers (GC and ESCC against controls), the overall accuracy was near perfect and accompanied by excellent sensitivity and specificity (Figure 9b). For CRC, IL-32 was a marker of good accuracy and excellent sensitivity accompanied by fair specificity (Figure 9c).
For the purpose of correlation analysis we have retrieved data on systemic concentrations of IL-1β, IL-4, IL-6, IL-8, IL-12p70, FGF2, G-CSF, GM-CSF, MCP-1, MIP-1α, PDGF-BB, TNFα, and VEGF-A, available for 43 of our cancer patients. Circulating IL-32 correlated positively with G-CSF, PDGF-BB, and TNFα and negatively with FGF2, MIP-1α, and VEGF-A (Table 3).
4. Discussion
Despite growing interest in IL-32, there is a scarcity of data concerning its status in the GIT cancers while its diagnostic utility has not been previously determined. In line with a pro-inflammatory character of the interleukin and inflammation-related nature of investigated cancers [15], all patients had higher serum interleukin concentration than healthy individuals. While this finding corroborates previous observations regarding the GIT cancers [25,26,27,28], here we showed it to have a diagnostic significance. The interleukin displayed an excellent accuracy in discriminating between non-cancer controls and ESCC and GC patients, which was accompanied by very good sensitivity and specificity. This observation, if confirmed on larger cohort, is of utmost clinical importance, taking into account the significance of early cancer detection for patient prognosis. Especially that the diagnosis in GC and ESCC is delayed due to uncharacteristic symptoms and, in case of ESCC, a lack of clear precancerous stage [4]. The interleukin accuracy in detecting CRC was good, but inferior to ESCC and GC. Yet, at optimal cut-off, high sensitivity was accompanied by poor specificity. Worse performance could be explained by the fact that CRC patients in our cohort had significantly lower circulating IL-32 than those with ESCC and GC. Difference between cancers regarding interleukin concentration was notable also locally, although positive correlation between systemic and local IL-32 was found only in GC patients. Here, we showed that IL-32 in esophageal and gastric mucosa was expressed more markedly than in the colorectum, both at protein and mRNA level. Markedly higher interleukin expression was not limited to esophageal and gastric tumors but was notable in non-cancerous tumor-adjacent mucosa as well. The IL32 transcripts were significantly overexpressed in tumors as compared to non-cancerous tissue only in ESCC patients. However, the observation was not corroborated at protein level, as IL-32 protein concentration was significantly upregulated solely in tumors from CRC patients and rather tended to be downregulated in ESCC. The discrepancy between the level of gene transcripts and protein is quite a frequent occurrence [29], previously observed for our patients with respect to expression of immunosuppressive IL-4 and IL-13 [22]. The phenomenon is explained by varying half-lives of mRNA and protein as well as by various posttranslational modifications which may further increase protein stability [29]. IL-32 expression has been repeatedly shown to be regulated by microRNAs [30,31], known to affect gene expression at transcriptional and/or translational level [32]. In fact, it has been suggested that transcriptomic and proteomic data should be treated as separate source of information [29]. In case of our study, the discrepancy might be associated with uncertainty of detected IL-32 isoforms. While the primer pair used in transcriptomic analysis covered all main variants, the specificity of antibody was not stated by the assay manufacturer but could be limited to some isoform.
If not for significantly higher protein content in esophageal than colonic mucosa, the discrepancy between ESCC and CRC could be attributed to differences in cancer histology. Accordingly, an analysis of IL-32 immunoreactivity in lung cancer has shown that adenocarcinomas had strong overexpression of interleukin-positive cells while the most of squamous cell carcinoma samples were lacking IL32-immunoreactivity [33]. Still, regarding the GIT cancers, IL32-positivity using the same antibody was higher in ESCC [34] than GC [35].
In an unpaired analysis, Yousif et al. [28] showed higher IL32-positivity in ESCC samples than non-cancerous mucosa obtained from patients operated for other reasons, which was also corroborated at mRNA level. We employed fully quantitative approach for IL-32 protein determination and used a paired design, analyzing patient-matched samples. While confirming Yousif’s et al. [28] results regarding mRNA, we failed to observe the IL-32 upregulation in tumors at protein level. As mentioned earlier, it may result from applied antibody being not optimal but may also indicate a cancer-associated accumulation of the interleukin in tumor-adjacent tissue. As has been repeatedly demonstrated [22,23,36,37,38], alterations in molecular profile of still non-transformed cells in tumor vicinity are common and may precede morphological and histological changes. The “tumor molecular margin” phenomenon predispose to neoplastic transformation and accounts for cancer recurrence and the synchronous tumors [39,40,41]. Supporting the notion, the abundance of IL32 transcripts in non-cancerous tumor-adjacent esophageal mucosa increased along with growing depth of tumor invasion. An association between IL-32 and the T stage of ESCC was reported also by Nabeki et al. [34], although it concerned interleukin immunoreactivity. The authors have observed that high IL32-immunopositivity coincided with greater infiltration with regulatory T-cells, indicative of a more immunosuppressive environment. Here, fold-change in protein concentration between tumor and adjacent tissue only tended to be higher in locally advanced cancers but it was significantly associated with the presence of lymph node involvement and an overall ESCC stage, linking the upregulation of IL-32 protein in tumors with gaining metastatic potential.
The role of IL-32 gastric cancer seems to be unequivocally negative. It has been shown to be overexpressed in H. pylori-infected gastric mucosa and modulate cytokine synthesis [42]. In addition, it has been upregulated in gastric tumors and predictive of poor prognosis [35,43]. The IL32-immunopositivity has been associated with greater depth of tumor invasion, lymph node involvement, and venous invasion [35], which we failed to observe in our cohort, either at protein or mRNA level. However, like in our patients, no significant association with the disease advancement or patient outcome has been noted for serum IL-32 [26]. In turn, serum IL-32 has tended to be more elevated in cardiac than distal GC [26]. Even in a small set of samples in our study, we were able to observe a similar trend but regarding local interleukin expression. Mechanistically, IL-32 has been demonstrated to promote changes in gastric cancer cell morphology, facilitating their migration, and to enhance their potential for invasiveness by upregulating the expression of matrix metalloproteinases (MMP)-2 and 9, VEGF-A, and IL-8 via HIF-1α activation [19]. It has also been suggested that IL-32 may induce immunosuppression and allow cancer cells to evade immune system facilitating metastasis [35].
In turn, there seem to be a discrepancy concerning IL-32 status and role in CRC. Experimental findings mostly indicate a tumor suppressive function. The θ isoform of IL-32 (IL-32θ) inhibits migration by hampering epithelial-mesenchymal transition (EMT) and temper the properties of cancer stem cells [20]. IL-32α and γ mediate and enhance cell death induced by TNFα [44,45] by increasing production of reactive oxygen species [44] or by upregulating pro-apoptotic Bax [45] and reducing expression of anti-apoptotic Bcl-2 [45]. In animal models, IL-32α confers protection against azoxymethane-induced carcinogenesis [44] and IL-32γ [45] and IL-32β [46,47] suppress tumor growth. Mechanistically, it is associated with induction of apoptosis, downregulation of proliferation markers, reduced expression of proinflamatory enzymes and cytokines, and increased secretion of immunosuppressive Il10 [46,47]. Contrary findings have been reported in humans. Catalan et al. [27] demonstrated IL-32α to be induced in response to hypoxia and to upregulate genes encoding mediators of inflammation and extracellular matrix remodeling. The CRC patients had elevated circulating IL-32 and their visceral adipose tissue had higher expression of the interleukin at both mRNA and protein level. Others have shown higher rates of IL32-immunopositivity in primary tumors derived from patients with lymph node metastasis as well as in metastatic as compared to corresponding primary tumors, regardless the location of distant metastases [48]. In our cohort, IL-32 has been significantly upregulated in colorectal tumors solely at protein level. Although not related to cancer pathology, its expression was dependent on the sublocation of primary tumors. Both IL32 transcripts and IL-32 protein were more abundant in non-cancerous tumor-adjacent tissue from CRC patients with right-sided tumors, while interleukin tumor expression displayed a similar tendency. Consequently, the fold-change in expression ratio (tumor-to-adjacent) was higher in left-sided CRCs, significantly so for IL32 transcripts. Circulating IL-32 was significantly more elevated in patients with tumors located in the left side of the colon. Although treated as one entity, CRC is highly heterogeneous with different set of protective and risk factors for tumors arising in right or left side of the colon and with subsite affecting disease presentation, treatment responsiveness and thus also patient prognosis. Right-sided tumors are considered to have less favorable characteristics than left-sided ones. Differences in clinical behavior stem from distinct genetic makeup and molecular patterns [49,50]. We and others have repeatedly documented dissimilarities between left- and right-sided cancers, not only locally [36,50,51,52,53] but also at systemic level [24,54,55,56] and now we showed the contribution of IL-32. This finding is of clinical relevance, since subsite heterogeneity, if not addressed, may hamper potential IL32-based therapies and reduce its diagnostic power as CRC biomarker.
To shed some light on possible role of IL-32 in GIT cancers, we analyzed the correlation patterns of the interleukin with major players in inflammation and immunity, cell proliferation and survival, angiogenesis and epithelial-mesenchymal transition, as well as metabolic reprogramming. Except for ACTA2 and PTGS2 (COX2) or IL10Ra, IL7Ra, and VEGFA, with which IL32 correlated positively only in tumors or adjacent mucosa, respectively, the correlation patterns in cancerous and non-cancerous tissue were quite similar. Fold-change in IL32 expression mirrored upregulation of proliferation marker Ki67, cell cycle regulator CDKN1A (p21CIP1/WAF1), metabolic reprogramming marker SLC2A1—encoding glucose transporter (GLUT1) angiogenic VEGFA, and IL7 and its receptor IL7Ra. Multivariate analysis, allowing for discerning associations independent from other covariates, showed a tight positive relationship between tumor IL32 expression and cell proliferation index and hypoxia, implying a tumor-promoting role for the interleukin. Mechanistically, the relationship between IL-32 and hypoxia can be bidirectional. Experimental studies in the colon have shown that cancer cell lines upregulate the interleukin expression in response to hypoxia while cell stimulation with IL-32α has no effect on HIF1A or VEGFA expression [27]. In the stomach, in turn, IL-32 have activated hypoxia-related transcriptional factor [19]. The interleukin, β and α, respectively, supported angiogenesis by upregulating VEGFA also in breast [45] and liver cancer cells [57]. Still, the proangiogenic effect of the interleukin on endothelial cells was independent from VEGF-A [58] what may explain lack of positive correlation between circulating IL-32 and VEGF-A. In fact, at the systemic level, the interleukin was inversely related with the growth factors as well as with FGF2, another potent proangiogenic factor, and a positive correlation was observed only between IL-32 and PDGF-BB. A negative relationship between IL-32 and VEGF-A has previously been reported in normal human bronchial epithelial cells [59]. An inverse relationship with FGF2 may be an echo of the inhibitory effect of the growth factor on fibroblast activation and mesenchymal transition [60]. It would be consistent with an EMT-promoting role ascribed to IL-32 overexpression, as discussed further.
Regarding proliferation, our findings oppose a role attributed to IL-32γ [46] and β [47] in the colon but are in agreement with the pro-proliferative activity of IL-32 in breast cancer [61]. When analyzed with reference to cancer type, IL32 showed strong positive correlation with Ki67 in gastric tumors (r = 0.72, p = 0.009) and only a weaker tendency in colonic tumors (r = 0.41. p = 0.072).
Tumor IL32 expression in our clinical samples was independently associated also with EMT, positively with mesenchymal marker ACTA2 (encoding α smooth muscle actin) and negatively with epithelial marker TJP1 (encoding zona occludens-1), and thus indicative of interleukin involvement in promoting invasion and metastasis. Those results are consistent with experimental findings, showing IL-32α to upregulate expression of proteins engaged in remodeling of extracellular matrix (SPP1 and MMP9) in the colon [27] or other EMT markers, namely vimentin and Slug, in the liver [57]. Noteworthy, however, the θ isoform of the interleukin has reportedly a negative impact on EMT in the colon [20]. In the non-transformed tissue surrounding tumor, IL32 expression was directly and independently associated with immunosuppressive and anti-inflammatory IL10R, consistent with the activity ascribed to IL-32β in the colon [47], or with antiapoptotic BCLxL. As others have shown that colonic cancer cells respond to IL-32 with induction of apoptosis [44], those results might indicate that IL-32 plays distinct roles at stages preceding neoplastic transformation than in already transformed cells.
In line with its role as an inflammation amplifier, the interleukin expression it is upregulated by TNFα in ESCC [62] and colon cancer [17] cell lines and, in a positive feed-back loop, activates NFκB in esophageal tumors [28] and upregulates TNFα expression and secretion in the colon [17,27]. Still, animal studies have shown IL-32γ and IL-32β to downregulate expression of inflammatory mediators [46,47]. Nonetheless, circulating interleukin is claimed to affect tumor microenvironment through positive regulation of pro-inflammatory cytokines [15] and to have modulatory effect on immune function [13,14]. Here, circulating IL-32 displayed a strong positive correlation with TNFα and G-CSF, while its local upregulation in tumors mirrored changes in the status of IL-7/IL-7Ra axis. There is no data linking IL-32 with G-CSF but GM-CSF, a colony-stimulating factor with overlapping functions in cancer, have been shown to induce expression of various isoforms of IL-32 in eosinophils [63]. The possible connection between IL-32 and IL-7/IL-7Ra axis is worth exploring in the light of increasing interest in this signaling pathway for anticancer therapy. Like IL-32, IL-7 in solid tumors is understudied and its role is poorly understood. Nonetheless, the accumulating evidence presents IL-7 as a cytokine facilitating tumor growth and metastasis and aiding drug-resistance [64].
A small number of available tissue samples, affecting the analysis in cancer subgroups, is a limitation that should be clearly acknowledged. Further studies on a larger cohort are needed to confirm the interleukin association with cancer pathology and tumor sublocation.
Author Contributions
Conceptualization, D.D. and M.K.-K.; data curation, D.D. and M.K.-K.; formal analysis, D.D. and M.K.-K.; investigation, D.D. and M.K.-.K; methodology, D.D. and M.K.-K.; resources, D.D. and M.K.-K.; visualization, D.D. and M.K.-K.; writing—original draft, D.D. and M.K.-K.; writing—review and editing, D.D. and M.K.-K. All authors have read and agreed to the published version of the manuscript
Funding
This research received no external funding.
Acknowledgments
The authors would like to thank the Foundation of Wroclaw Medical University (FAM) and its Board Chairmen for financing a lease of BioRad Experion platform and CFX96 Real-Time PCR system.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| AUC | Area under the ROC curve |
| CI | Confidence interval |
| COX | Cyclooxygenase |
| CRC | Colorectal adenocarcinoma |
| EMT | Epithelial-mesenchymal transition |
| ESCC | Esophageal squamous cell carcinoma |
| FGF | Fibroblast growth factor |
| GC | Gastric adenocarcinoma |
| G-CSF | Granulocyte colony-stimulating factor |
| GIT | Gastrointestinal tract |
| GM-CSF | Granulocyte-macrophage colony-stimulating factor |
| IBD | Inflammatory bowel disease |
| IFN | Interferon |
| IL | Interleukin |
| MCP | Monocyte chemoattractant protein |
| MIP | Macrophage inflammatory protein |
| NOS | Nitric oxide synthase |
| NRQ | Normalized relative quantities |
| PDGF | Platelet-derived growth factor |
| ROC | Receiver operating characteristics |
| RT-qPCR | Reverse-transcribed quantitative (real-time) polymerase chain reaction |
| TNF | Tumor necrosis factor |
| TNM | Tumor-node-metastasis cancer staging system |
| VEGF | Vascular endothelial growth factor |
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Tustumi, F.; Kimura, C.M.; Takeda, F.R.; Uema, R.H.; Salum, R.A.; Ribeiro-Junior, U.; Cecconello, I. Prognostic Factors and Survival Analysis in Esophageal Carcinoma. ABCD Arq. Bras. Cir. Dig. 2016, 29, 138–141. [Google Scholar] [CrossRef] [PubMed]
- Thrift, A.P.; El-Serag, H.B. Burden of Gastric Cancer. Clin. Gastroenterol. Hepatol. 2020, 18, 534–542. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.V.V.; Sagar, R.; Mathew, J. Squamous cell carcinoma: Esophagus. In Squamous Cell Carcinoma—Hallmark and Treatment Modalities; Daaboul, H.E., Ed.; IntechOpen Ltd.: London, UK, 2019. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, T.; Saikawa, Y.; Kitagawa, Y. Gastric cancer: Current status of diagnosis and treatment. Cancers 2013, 5, 48–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aftimos, P.G.; Barthelemy, P.; Awada, A. Molecular Biology in Medical Oncology: Diagnosis, Prognosis, and Precision Medicine. Discov. Med. 2014, 17, 81–91. [Google Scholar]
- Yang, J.W.; Choi, Y.L. Genomic profiling of esophageal squamous cell carcinoma (ESCC)-Basis for precision medicine. Pathol. Res. Pract. 2017, 213, 836–841. [Google Scholar] [CrossRef]
- Liu, X.; Meltzer, S.J. Gastric Cancer in the Era of Precision Medicine. Cell Mol. Gastroenterol. Hepatol. 2017, 3, 348–358. [Google Scholar] [CrossRef] [Green Version]
- Aziz, M.A. Precision medicine in colorectal cancer. Saudi J. Gastroenterol. 2019, 25, 139–140. [Google Scholar] [CrossRef]
- Kim, E.R.; Chang, D.K. Colorectal cancer in inflammatory bowel disease: The risk, pathogenesis, prevention and diagnosis. World J. Gastroenterol. 2014, 20, 9872–9881. [Google Scholar] [CrossRef]
- O’Sullivan, K.E.; Phelan, J.J.; O’Hanlon, C.; Lysaght, J.; O’Sullivan, J.N.; Reynolds, J.V. The role of inflammation in cancer of the esophagus. Expert Rev. Gastroenterol. Hepatol. 2014, 8, 749–760. [Google Scholar] [CrossRef]
- Rajput, S.; Wilber, A. Roles of inflammation in cancer initiation, progression, and metastasis. Front. Biosci. 2010, 2, 176–183. [Google Scholar] [CrossRef] [Green Version]
- Joosten, L.A.; Heinhuis, B.; Netea, M.G.; Dinarello, C.A. Novel insights into the biology of interleukin-32. Cell Mol. Life Sci. 2013, 70, 3883–3892. [Google Scholar] [CrossRef] [PubMed]
- Khawar, M.B.; Abbasi, M.H.; Sheikh, N. IL-32: A Novel Pluripotent Inflammatory Interleukin, towards Gastric Inflammation, Gastric Cancer, and Chronic Rhino Sinusitis. Mediators Inflamm. 2016, 2016, 8413768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, S.; Yang, Y. Interleukin-32: Frenemy in cancer? BMB Rep. 2019, 52, 165–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, H.; He, D.; Huang, X.; Zhang, E.; Chen, Q.; Xu, R.; Liu, X.; Zi, F.; Cai, Z. Role of interleukin-32 in cancer biology. Oncol. Lett. 2018, 16, 41–47. [Google Scholar] [CrossRef]
- Shioya, M.; Nishida, A.; Yagi, Y.; Ogawa, A.; Tsujikawa, T.; Kim-Mitsuyama, S.; Takayanagi, A.; Shimizu, N.; Fujiyama, Y.; Andoh, A. Epithelial overexpression of interleukin-32alpha in inflammatory bowel disease. Clin. Exp. Immunol. 2007, 149, 480–486. [Google Scholar] [CrossRef]
- Sloot, Y.J.E.; Smit, J.W.; Joosten, L.A.B.; Netea-Maier, R.T. Insights into the role of IL-32 in cancer. Semin. Immunol. 2018, 38, 24–32. [Google Scholar] [CrossRef]
- Tsai, C.Y.; Wang, C.S.; Tsai, M.M.; Chi, H.C.; Cheng, W.L.; Tseng, Y.H.; Chen, C.Y.; Lin, C.D.; Wu, J.I.; Wang, L.H. Interleukin-32 increases human gastric cancer cell invasion associated with tumor progression and metastasis. Clin. Cancer Res. 2014, 20, 2276–2288. [Google Scholar] [CrossRef] [Green Version]
- Bak, Y.; Kwon, T.; Bak, I.S.; Hong, J.; Yu, D.Y.; Yoon, D.Y. IL-32θ inhibits stemness and epithelial-mesenchymal transition of cancer stem cells via the STAT3 pathway in colon cancer. Oncotarget 2016, 7, 7307–7317. [Google Scholar] [CrossRef] [Green Version]
- Hellemans, J.; Vandesompele, J. qPCR data analysis—Unlocking the secret to successful results. In PCR Troubleshooting and Optimization: The Essential Guide, 1st ed.; Kennedy, S., Oswald, N., Eds.; Caister Academic Press: Poole, UK, 2011; pp. 1–13. [Google Scholar]
- Bednarz-Misa, I.; Diakowska, D.; Szczuka, I.; Fortuna, P.; Kubiak, A.; Rosińczuk, J.; Krzystek-Korpacka, M. Interleukins 4 and 13 and Their Receptors Are Differently Expressed in Gastrointestinal Tract Cancers, Depending on the Anatomical Site and Disease Advancement, and Improve Colon Cancer Cell Viability and Motility. Cancers 2020, 12, 1463. [Google Scholar] [CrossRef]
- Bednarz-Misa, I.; Fortuna, P.; Diakowska, D.; Jamrozik, N.; Krzystek-Korpacka, M. Distinct Local and Systemic Molecular Signatures in the Esophageal and Gastric Cancers: Possible Therapy Targets and Biomarkers for Gastric Cancer. Int. J. Mol. Sci. 2020, 21, 4509. [Google Scholar] [CrossRef] [PubMed]
- Krzystek-Korpacka, M.; Diakowska, D.; Kapturkiewicz, B.; Bębenek, M.; Gamian, A. Profiles of circulating inflammatory cytokines in colorectal cancer (CRC), high cancer risk conditions, and health are distinct. Possible implications for CRC screening and surveillance. Cancer Lett. 2013, 337, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Seo, E.H.; Kang, J.; Kim, K.H.; Cho, M.C.; Lee, S.; Kim, H.J.; Kim, J.H.; Kim, E.J.; Park, D.K.; Kim, S.H. Detection of expressed IL-32 in human stomach cancer using ELISA and immunostaining. J. Microbiol. Biotechnol. 2008, 18, 1606–1612. [Google Scholar] [PubMed]
- Erturk, K.; Tastekin, D.; Serilmez, M.; Bilgin, E.; Bozbey, H.U.; Vatansever, S. Clinical significance of serum interleukin-29, interleukin-32, and tumor necrosis factor alpha levels in patients with gastric cancer. Tumour Biol. 2016, 37, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Catalán, V.; Gómez-Ambrosi, J.; Rodríguez, A.; Ramírez, B.; Ortega, V.A.; Hernández-Lizoain, J.L.; Baixauli, J.; Becerril, S.; Rotellar, F.; Valentí, V. IL-32α-induced inflammation constitutes a link between obesity and colon cancer. Oncoimmunology 2017, 6, e1328338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yousif, N.G.; Al-Amran, F.G.; Hadi, N.; Lee, J.; Adrienne, J. Expression of IL32 modulates NF-κB and p38 MAP kinase pathways in human esophageal cancer. Cytokine 2013, 61, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Schlesinger-Raab, A.; Werner, J.; Friess, H.; Hölzel, D.; Engel, J. Age and outcome in gastrointestinal cancers: A population-based evaluation of oesophageal, gastric and colorectal cancer. Visc. Med. 2017, 33, 245–253. [Google Scholar] [CrossRef]
- Li, Y.; Xie, J.; Xu, X.; Liu, L.; Wan, Y.; Liu, Y.; Zhu, C.; Zhu, Y. Inducible interleukin 32 (IL-32) exerts extensive antiviral function via selective stimulation of interferon λ1 (IFN-λ1). J. Biol. Chem. 2013, 288, 20927–20941. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Ye, W.; Kou, N.; Chen, K.; Cui, B.; Zhang, X.; Hu, S.; Liu, T.; Kang, L.; Li, X. MicroRNA-29b-3p suppresses oral squamous cell carcinoma cell migration and invasion via IL32/AKT signalling pathway. J. Cell. Mol. Med. 2020, 24, 841–849. [Google Scholar] [CrossRef] [Green Version]
- Oliveto, S.; Mancino, M.; Manfrini, N.; Biffo, S. Role of microRNAs in translation regulation and cancer. World J. Biol. Chem. 2017, 8, 45–56. [Google Scholar] [CrossRef]
- Sorrentino, C.; Di Carlo, E. Expression of IL32 in human lung cancer is related to the histotype and metastatic phenotype. Am. J. Respir. Crit. Care Med. 2009, 180, 769–779. [Google Scholar] [CrossRef] [PubMed]
- Nabeki, B.; Ishigami, S.; Uchikado, Y.; Sasaki, K.; Kita, Y.; Okumura, H.; Arigami, T.; Kijima, Y.; Kurahara, H.; Maemura, K. Interleukin-32 expression and Treg infiltration in esophageal squamous cell carcinoma. Anticancer Res. 2015, 35, 2941–2947. [Google Scholar] [PubMed]
- Ishigami, S.; Arigami, T.; Uchikado, Y.; Setoyama, T.; Kita, Y.; Sasaki, K.; Okumura, H.; Kurahara, H.; Kijima, Y.; Harada, A. IL32 expression is an independent prognostic marker for gastric cancer. Med. Oncol. 2013, 30, 472. [Google Scholar] [CrossRef]
- Neubauer, K.; Bednarz-Misa, I.; Diakowska, D.; Kapturkiewicz, B.; Gamian, A.; Krzystek-Korpacka, M. Nampt/PBEF/visfatin upregulation in colorectal tumors, mirrored in normal tissue and whole blood of colorectal cancer patients, is associated with metastasis, hypoxia, IL1β, and anemia. Biomed. Res. Int. 2015, 2015, 523930. [Google Scholar] [CrossRef] [Green Version]
- Krzystek-Korpacka, M.; Gorska, S.; Diakowska, D.; Kapturkiewicz, B.; Podkowik, M.; Gamian, A.; Bednarz-Misa, I. Midkine is up-regulated in both cancerous and inflamed bowel, reflecting lymph node metastasis in colorectal cancer and clinical activity of ulcerative colitis. Cytokine 2017, 89, 68–75. [Google Scholar] [CrossRef]
- Bednarz-Misa, I.; Diakowska, D.; Krzystek-Korpacka, M. Local and systemic IL-7 concentration in gastrointestinal-tract cancers. Medicina 2019, 55, 262. [Google Scholar] [CrossRef] [Green Version]
- Mao, L.; Clark, D. Molecular margin of surgical resections—where do we go from here? Cancer 2015, 121, 1914–1916. [Google Scholar] [CrossRef] [PubMed]
- Dakubo, G.D.; Jakupciak, J.P.; Birch-Machin, M.A.; Parr, R.L. Clinical implications and utility of field cancerization. Cancer Cell Int. 2007, 7, 2. [Google Scholar] [CrossRef] [Green Version]
- Patel, A.; Tripathi, G.; Gopalakrishnan, K.; Williams, N.; Arasaradnam, R.P. Field cancerisation in colorectal cancer: A new frontier or pastures past? World J. Gastroenterol. 2015, 21, 3763–3772. [Google Scholar] [CrossRef]
- Sakitani, K.; Hirata, Y.; Hayakawa, Y.; Serizawa, T.; Nakata, W.; Takahashi, R.; Kinoshita, H.; Sakamoto, K.; Nakagawa, H.; Akanuma, M. Role of interleukin-32 in Helicobacter pylori-induced gastric inflammation. Infect. Immun. 2012, 80, 3795–3803. [Google Scholar] [CrossRef] [Green Version]
- Chang, W.J.; Du, Y.; Zhao, X.; Ma, L.Y.; Cao, G.W. Inflammation-related factors predicting prognosis of gastric cancer. World J. Gastroenterol. 2014, 20, 4586–4596. [Google Scholar] [CrossRef] [PubMed]
- Yun, H.M.; Park, K.R.; Kim, E.C.; Han, S.B.; Yoon, D.Y.; Hong, J.T. IL-32α suppresses colorectal cancer development via TNFR1-mediated death signaling. Oncotarget 2015, 6, 9061–9072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, E.S.; Yoo, J.M.; Yoo, H.S.; Yoon, D.Y.; Yun, Y.P.; Hong, J. IL-32γ enhances TNF-α-induced cell death in colon cancer. Mol. Carcinog. 2014, 53 (Suppl. 1), E23–E35. [Google Scholar] [CrossRef] [PubMed]
- Yun, H.M.; Oh, J.H.; Shim, J.H.; Ban, J.O.; Park, K.R.; Kim, J.H.; Lee, D.H.; Kang, J.W.; Park, Y.H.; Yu, D. Antitumor activity of IL-32β through the activation of lymphocytes, and the inactivation of NF-κB and STAT3 signals. Cell Death Dis. 2013, 4, e640. [Google Scholar] [CrossRef]
- Oh, J.H.; Cho, M.C.; Kim, J.H.; Lee, S.Y.; Kim, H.J.; Park, E.S.; Ban, J.O.; Kang, J.W.; Lee, D.H.; Shim, J.H. IL-32γ inhibits cancer cell growth through inactivation of NF-κB and STAT3 signals. Oncogene 2011, 30, 3345–3359. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Wang, Z.; Zhou, Y.; Wang, X.; Xiang, J.; Chen, Z. Dysregulation of over-expressed IL-32 in colorectal cancer induces metastasis. World J. Surg. Oncol. 2015, 13, 146. [Google Scholar] [CrossRef] [Green Version]
- Hemminki, K.; Santi, I.; Weires, M.; Thomsen, H.; Sundquist, J.; Bermejo, J.L. Tumor location and patient characteristics of colon and rectal adenocarcinomas in relation to survival and TNM classes. BMC Cancer 2010, 10, 688. [Google Scholar] [CrossRef] [Green Version]
- Greystoke, A.; Mullamitha, S.A. How many diseases are colorectal cancer? Gastroenterol. Res. Pract. 2012, 2012, 564741. [Google Scholar] [CrossRef]
- Yamauchi, M.; Lochhead, P.; Morikawa, T.; Huttenhower, C.; Chan, A.T.; Giovannucci, E.; Fuchs, C.; Ogino, S. Colorectal cancer: A tale of two sides or a continuum? Gut 2012, 61, 794–797. [Google Scholar] [CrossRef] [Green Version]
- Simons, C.C.; Hughes, L.A.; Smits, K.M.; Khalid-de Bakker, C.A.; de Bruïne, A.P.; Carvalho, B.; Meijer, G.A.; Schouten, L.J.; van den Brandt, P.A.; Weijenberg, M.P. A novel classification of colorectal tumors based on microsatellite instability, the CpG island methylator phenotype and chromosomal instability: Implications for prognosis. Ann. Oncol. 2013, 24, 2048–2056. [Google Scholar] [CrossRef]
- Krzystek-Korpacka, M.; Diakowska, D.; Grabowski, K.; Gamian, A. Tumor location determines midkine level and its association with the disease progression in colorectal cancer patients: A pilot study. Int. J. Colorectal. Dis. 2012, 27, 1319–1324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krzystek-Korpacka, M.; Diakowska, D.; Neubauer, K.; Gamian, A. Circulating midkine in malignant and non-malignant colorectal diseases. Cytokine 2013, 64, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Krzystek-Korpacka, M.; Zawadzki, M.; Neubauer, K.; Bednarz-Misa, I.; Górska, S.; Wiśniewski, J.; Witkiewicz, W.; Gamian, A. Elevated systemic interleukin-7 in patients with colorectal cancer and individuals at high risk of cancer: Association with lymph node involvement and tumor location in the right colon. Cancer Immunol. Immunother. 2017, 66, 171–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krzystek-Korpacka, M.; Zawadzki, M.; Kapturkiewicz, B.; Lewandowska, P.; Bednarz-Misa, I.; Gorska, S.; Witkiewicz, W.; Gamian, A. Subsite heterogeneity in the profiles of circulating cytokines in colorectal cancer. Cytokine 2018, 110, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.B.; Wang, Q.L.; Xu, Y.T.; Xu, S.F.; Qiu, Y.; Zhu, F. Overexpression of interleukin-32α promotes invasion by modulating VEGF in hepatocellular carcinoma. Oncol. Rep. 2018, 39, 1155–1162. [Google Scholar] [CrossRef] [Green Version]
- Nold-Petry, C.A.; Rudloff, I.; Baumer, Y.; Ruvo, M.; Marasco, D.; Botti, P.; Farkas, L.; Cho, S.X.; Zepp, J.A.; Azam, T. IL-32 promotes angiogenesis. J. Immunol. 2014, 192, 589–602. [Google Scholar] [CrossRef] [Green Version]
- Meyer, N.; Christoph, J.; Makrinioti, H.; Indermitte, P.; Rhyner, C.; Soyka, M.; Eiwegger, T.; Chalubinski, M.; Wanke, K.; Fujita, H. Inhibition of angiogenesis by IL-32: Possible role in asthma. J. Allergy Clin. Immunol. 2012, 129, 964–973. [Google Scholar] [CrossRef]
- Dolivo, D.M.; Larson, S.A.; Dominko, T. Fibroblast Growth Factor 2 as an Antifibrotic: Antagonism of Myofibroblast Differentiation and Suppression of Pro-Fibrotic Gene Expression. Cytokine Growth Factor Rev. 2017, 38, 49–58. [Google Scholar] [CrossRef]
- Wang, S.; Chen, F.; Tang, L. IL-32 promotes breast cancer cell growth and invasiveness. Oncol. Lett. 2015, 9, 305–307. [Google Scholar] [CrossRef] [Green Version]
- Neuzillet, J.M.; Traynor, C.; Hullmann, G.; Tyritzis, G.; Shimizu, J. TNF-α-induced IL-32 expression in esophageal cancer: Cross talk JNK/Akt signaling pathways. Am. J. BioMed. 2013, 1, 73–85. [Google Scholar] [CrossRef]
- Ribeiro-Dias, F.; Saar Gomes, R.; de Lima Silva, L.L.; Dos Santos, J.C.; Joosten, L.A. Interleukin 32: A novel player in the control of infectious diseases. J. Leukoc Biol. 2017, 101, 39–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bednarz-Misa, I.; Bromke, M.; Krzystek-Korpacka, M. Interleukin (IL)-7 signaling in the tumor microenvironment. In Tumor Microenvironment; Birbrair, A., Ed.; Springer: Berlin/Heidelberg, Germany, in press.
Figure 1.
Pair-wise analysis of IL-32 expression at mRNA level: (a) in esophageal squamous cell carcinoma; (b) in gastric adenocarcinoma; (c) in colorectal adenocarcinoma. Data presented as geometric means of normalized relative quantities (NRQ) with 95% confidence interval and analyzed using t-test for paired samples.
Figure 1.
Pair-wise analysis of IL-32 expression at mRNA level: (a) in esophageal squamous cell carcinoma; (b) in gastric adenocarcinoma; (c) in colorectal adenocarcinoma. Data presented as geometric means of normalized relative quantities (NRQ) with 95% confidence interval and analyzed using t-test for paired samples.

Figure 2.
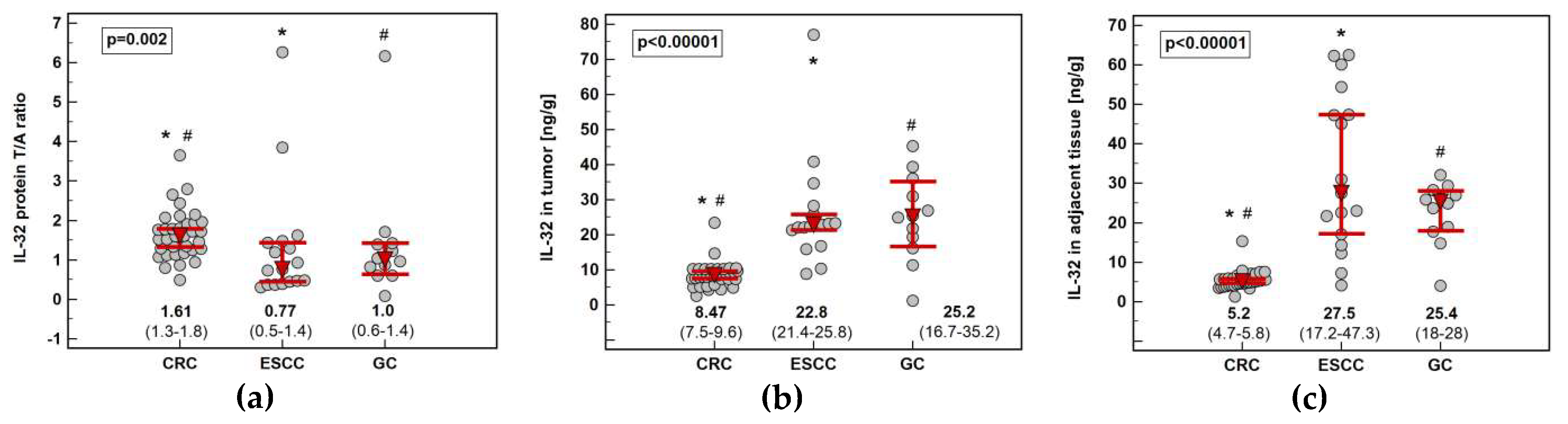
Impact of cancer type on IL32 expression: (a) fold-change in expression in tumor and non-cancerous tumor-adjacent tissue (T/A ratio); (b) in tumors; (c) in non-cancerous tumor-adjacent tissue. Data presented as geometric means of normalized relative quantities (NRQ) with 95% confidence interval (red triangles with whiskers and numeric data below the dot-plots) and analyzed using one-way analysis of variance with Student–Newman–Keuls post-hoc test. Significant between-groups differences are marked by the symbols (*, #, ⟂) of the same type. CRC, colorectal cancer; ESCC, esophageal squamous cell carcinoma; GC, gastric cancer.
Figure 2.
Impact of cancer type on IL32 expression: (a) fold-change in expression in tumor and non-cancerous tumor-adjacent tissue (T/A ratio); (b) in tumors; (c) in non-cancerous tumor-adjacent tissue. Data presented as geometric means of normalized relative quantities (NRQ) with 95% confidence interval (red triangles with whiskers and numeric data below the dot-plots) and analyzed using one-way analysis of variance with Student–Newman–Keuls post-hoc test. Significant between-groups differences are marked by the symbols (*, #, ⟂) of the same type. CRC, colorectal cancer; ESCC, esophageal squamous cell carcinoma; GC, gastric cancer.

Figure 3.
Impact of tumor location in the colon on IL32 expression: (a) fold-change in expression in tumor and non-cancerous tumor-adjacent tissue (T/A ratio); (b) in tumors; (c) in non-cancerous tumor-adjacent tissue. Data presented as geometric means of normalized relative quantities (NRQ) with 95% confidence interval (red triangles with whiskers and numeric data below the dot-plots) and analyzed using t-test for independent samples.
Figure 3.
Impact of tumor location in the colon on IL32 expression: (a) fold-change in expression in tumor and non-cancerous tumor-adjacent tissue (T/A ratio); (b) in tumors; (c) in non-cancerous tumor-adjacent tissue. Data presented as geometric means of normalized relative quantities (NRQ) with 95% confidence interval (red triangles with whiskers and numeric data below the dot-plots) and analyzed using t-test for independent samples.

Figure 4.
Pair-wise analysis of IL-32 concentration (protein) in tissue homogenates: (a) in esophageal squamous cell carcinoma; (b) in gastric adenocarcinoma; (c) in colorectal adenocarcinoma. Data presented as medians of interleukin concentration normalized to tissue weight with 95% confidence interval and analyzed using Wilcoxon test.
Figure 4.
Pair-wise analysis of IL-32 concentration (protein) in tissue homogenates: (a) in esophageal squamous cell carcinoma; (b) in gastric adenocarcinoma; (c) in colorectal adenocarcinoma. Data presented as medians of interleukin concentration normalized to tissue weight with 95% confidence interval and analyzed using Wilcoxon test.

Figure 5.
Impact of cancer type on IL-32 concentration (protein): (a) fold-change in concentration in tumor and non-cancerous tumor-adjacent tissue (T/A ratio); (b) in tumors; (c) in non-cancerous tumor-adjacent tissue. Data presented as medians with 95% confidence interval (red triangles with whiskers and numeric data below the dot-plots) and analyzed using Kruskal–Wallis H test with Conover post-hoc test. Significant between-groups differences are marked by the symbols (*, #) of the same type. CRC, colorectal cancer; ESCC, esophageal squamous cell carcinoma; GC, gastric cancer.
Figure 5.
Impact of cancer type on IL-32 concentration (protein): (a) fold-change in concentration in tumor and non-cancerous tumor-adjacent tissue (T/A ratio); (b) in tumors; (c) in non-cancerous tumor-adjacent tissue. Data presented as medians with 95% confidence interval (red triangles with whiskers and numeric data below the dot-plots) and analyzed using Kruskal–Wallis H test with Conover post-hoc test. Significant between-groups differences are marked by the symbols (*, #) of the same type. CRC, colorectal cancer; ESCC, esophageal squamous cell carcinoma; GC, gastric cancer.

Figure 6.
Impact of ESCC advancement on IL-32 protein concentration: (a) tumor-node-metastasis (TNM) stage; (b) lymph node involvement (stage N). Data presented as medians with 95% confidence interval (red triangles with whiskers and numeric data below the dot-plots). ESCC, esophageal squamous cell carcinoma.
Figure 6.
Impact of ESCC advancement on IL-32 protein concentration: (a) tumor-node-metastasis (TNM) stage; (b) lymph node involvement (stage N). Data presented as medians with 95% confidence interval (red triangles with whiskers and numeric data below the dot-plots). ESCC, esophageal squamous cell carcinoma.

Figure 7.
Impact of tumor location in the colon on IL-32 concentration: (a) fold-change in expression in tumor and non-cancerous tumor-adjacent tissue (T/A ratio); (b) in tumors; (c) in non-cancerous tumor-adjacent tissue. Data presented as geometric means with 95% confidence interval (red triangles with whiskers and numeric data below the dot-plots) and analyzed using t-test for independent samples.
Figure 7.
Impact of tumor location in the colon on IL-32 concentration: (a) fold-change in expression in tumor and non-cancerous tumor-adjacent tissue (T/A ratio); (b) in tumors; (c) in non-cancerous tumor-adjacent tissue. Data presented as geometric means with 95% confidence interval (red triangles with whiskers and numeric data below the dot-plots) and analyzed using t-test for independent samples.

Figure 8.
Circulating IL-32: (a) association with cancer type; (b) association with tumor location in the colon. Data presented as medians with 95% confidence interval (red triangles with whiskers and numeric data below the dot-plots) and analyzed using Kruskal–Wallis H test with Conover post-hoc test or Mann–Whitney U test. Significant between-group differences are marked with the same type of symbol (*, #, ⟂, $, &).
Figure 8.
Circulating IL-32: (a) association with cancer type; (b) association with tumor location in the colon. Data presented as medians with 95% confidence interval (red triangles with whiskers and numeric data below the dot-plots) and analyzed using Kruskal–Wallis H test with Conover post-hoc test or Mann–Whitney U test. Significant between-group differences are marked with the same type of symbol (*, #, ⟂, $, &).

Figure 9.
IL-32 as a cancer marker: (a) general cancer marker; (b) marker for gastroesophageal cancers (ESCC+GC vs. controls); (c) marker for colorectal cancer. Data presented as receiver operating (ROC) curves with 95% confidence interval (respectively, solid and dashed lines). AUC, area under ROC curve (indicating marker overall accuracy); CI, confidence interval; sens., sensitivity; spec., specificity accompanying optimal cut-off value (marked as a red dot); ESCC, esophageal squamous cell carcinoma; GC, gastric cancer.
Figure 9.
IL-32 as a cancer marker: (a) general cancer marker; (b) marker for gastroesophageal cancers (ESCC+GC vs. controls); (c) marker for colorectal cancer. Data presented as receiver operating (ROC) curves with 95% confidence interval (respectively, solid and dashed lines). AUC, area under ROC curve (indicating marker overall accuracy); CI, confidence interval; sens., sensitivity; spec., specificity accompanying optimal cut-off value (marked as a red dot); ESCC, esophageal squamous cell carcinoma; GC, gastric cancer.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Characteristics of study population.
| Characteristics | Controls | ESCC | GC | CRC | p Value |
|---|---|---|---|---|---|
| n | 28 | 17 | 14 | 41 | - |
| Sex (F/M), n | 12/16 | 7/10 | 4/10 | 21/20 | 0.515 1 |
| Age [yrs.], median (95%CI) | 57 (53–61.6) | 61 (58–65) | 64 (58–75.3) | 59 (54–65.2) | 0.121 2 |
| TNM stage (I/II/III/IV), n | - | 1/6/9/1 | 2/3/5/4 | 14/12/12/4 | 0.079 1 |
| Primary tumor, T (1/2/3/4), n | - | 0/6/9/2 | 1/1/8/4 | 5/11/22/3 | 0.201 1 |
| Lymph node metastasis, N (no/yes), n | - | 8/9 | 5/9 | 26/15 | 0.159 1 |
| Distant metastasis, M (no/yes), n | - | 16/1 | 10/4 | 37/4 | 0.118 1 |
1 Chi-squared test; 2 Kruskal–Wallis H test. ESCC, esophageal squamous cell carcinoma; GC, gastric cancer; CRC, colorectal cancer; n, number of observations; F/M, female-to-male ratio; yrs., years; CI, confidence interval; TNM, tumor-node-metastasis cancer staging system.
Table 2.
Correlation pattern of IL32 and cancer-promoting genes.
| Gene | Tumor IL32 | Adjacent IL32 | Fold-Change (T/A) in IL32 |
|---|---|---|---|
| ACTA2 | 0.33, p = 0.031 | ns | ns |
| BCL2 | 0.39, p = 0.019 | 0.39, p = 0.008 | ns |
| BCLxL | 0.34, p = 0.023 | 0.57, p < 0.001 | 0.37, p = 0.015 |
| CCL2 | 0.45, p = 0.002 | 0.32, p = 0.037 | 0.30, p = 0.049 |
| CDKN1A | 0.27, p = 0.079 | 0.41, p = 0.006 | 0.51, p < 0.001 |
| HIF1A | 0.57, p < 0.001 | 0.40, p = 0.007 | 0.31, p = 0.040 |
| IL10 | 0.29, p = 0.053 | ns | ns |
| IL10Ra | ns | 0.41, p = 0.005 | ns |
| IL13Ra | 0.37, p = 0.012 | 0.40, p = 0.007 | ns |
| IL7 | 0.32, p = 0.037 | 0.40, p = 0.007 | 0.41, p = 0.005 |
| IL7Ra | ns | 0.37, p = 0.012 | 0.38, p = 0.011 |
| Ki67 | 0.62, p < 0.001 | 0.49, p < 0.001 | 0.48, p = 0.001 |
| ODC1 | 0.45, p = 0.002 | 0.35, p = 0.019 | 0.29, p = 0.056 |
| PTGS2 | 0.30, p = 0.045 | ns | ns |
| SLC2A1 | 0.54, p < 0.001 | 0.53, p < 0.001 | 0.55, p < 0.001 |
| TJP1 | 0.39, p = 0.008 | 0.31, p = 0.041 | ns |
| VEGFA | ns | 0.52, p < 0.001 | 0.46, p = 0.002 |
Data analyzed following log-transformation and presented as Pearson correlation coefficient (r). T/A, tumor-adjacent ratio; ns, non-significant; ACTA2, smooth muscle actin alpha 2; BCL2, B-cell lymphoma 2; BCLxL, B-cell lymphoma-extra large; CCL2, C-C motif chemokine ligand 2; CDKN1A, p21CIP1/WAF1; HIF1A, hypoxia-inducible factor 1α; IL10, interleukin 10; IL10Ra, interleukin 10 receptor subunit α; IL13Ra, interleukin 13 receptor subunit α; IL7, interleukin 7; IL7Ra, interleukin 7 receptor subunit α; Ki67, proliferation marker Ki67; ODC1, ornithine decarboxylase 1; PTGS2, cyclooxygenase 2; SLC2A1; glucose transporter 1; TJP1; tight junction protein 1; VEGFA, vascular endothelial growth factor A.
Table 3.
Correlation pattern of serum IL-32 and circulating cytokines and growth factors.
| Cytokine/Growth Factor | Spearman Correlation Coefficient (ρ), p |
|---|---|
| FGF2 | −0.52, p < 0.001 |
| G-CSF | 0.53, p < 0.001 |
| MIP-1α | −0.35, p = 0.021 |
| PDGF-BB | 0.46, p = 0.002 |
| TNFα | 0.63, p < 0.001 |
| VEGF-A | −0.52, p < 0.001 |
FGF2, fibroblast growth factor 2, G-CSF, granulocyte colony-stimulating growth factor; MIP-1α, macrophage inflammatory protein 1α; PDGF-BB, platelet-derived growth factor BB; TNFα, tumor necrosis factor α; VEGF-A, vascular endothelial growth factor A.
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Diakowska, D.; Krzystek-Korpacka, M. Local and Systemic Interleukin-32 in Esophageal, Gastric, and Colorectal Cancers: Clinical and Diagnostic Significance. Diagnostics 2020, 10, 785. https://doi.org/10.3390/diagnostics10100785
AMA Style
Diakowska D, Krzystek-Korpacka M. Local and Systemic Interleukin-32 in Esophageal, Gastric, and Colorectal Cancers: Clinical and Diagnostic Significance. Diagnostics. 2020; 10(10):785. https://doi.org/10.3390/diagnostics10100785
Chicago/Turabian StyleDiakowska, Dorota, and Małgorzata Krzystek-Korpacka. 2020. "Local and Systemic Interleukin-32 in Esophageal, Gastric, and Colorectal Cancers: Clinical and Diagnostic Significance" Diagnostics 10, no. 10: 785. https://doi.org/10.3390/diagnostics10100785
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.