Beyond Uterine Natural Killer Cell Numbers in Unexplained Recurrent Pregnancy Loss: Combined Analysis of CD45, CD56, CD16, CD57, and CD138

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Human Endometrial Samples
2.2. Immunohistochemical Staining of Immune Cell Markers
2.3. Cell Counting and Statistical Analysis
3. Results
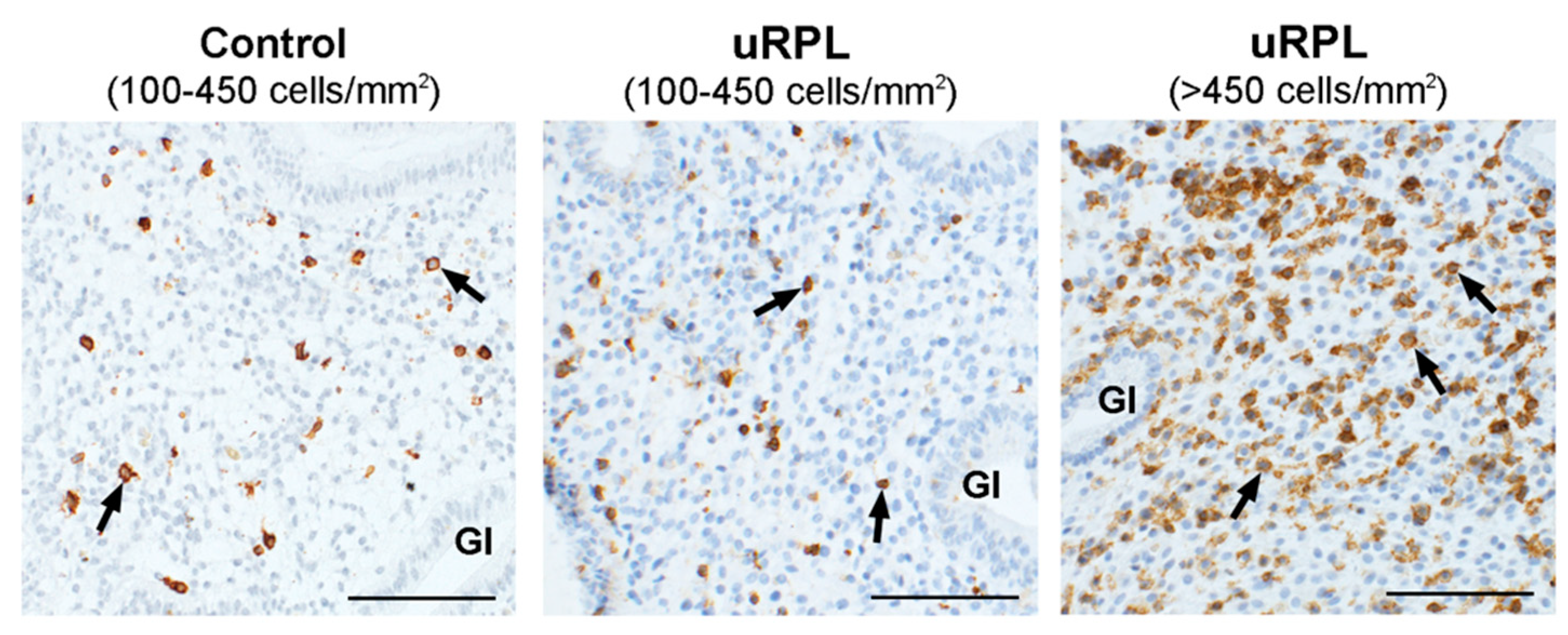
3.1. CD45
3.2. CD56
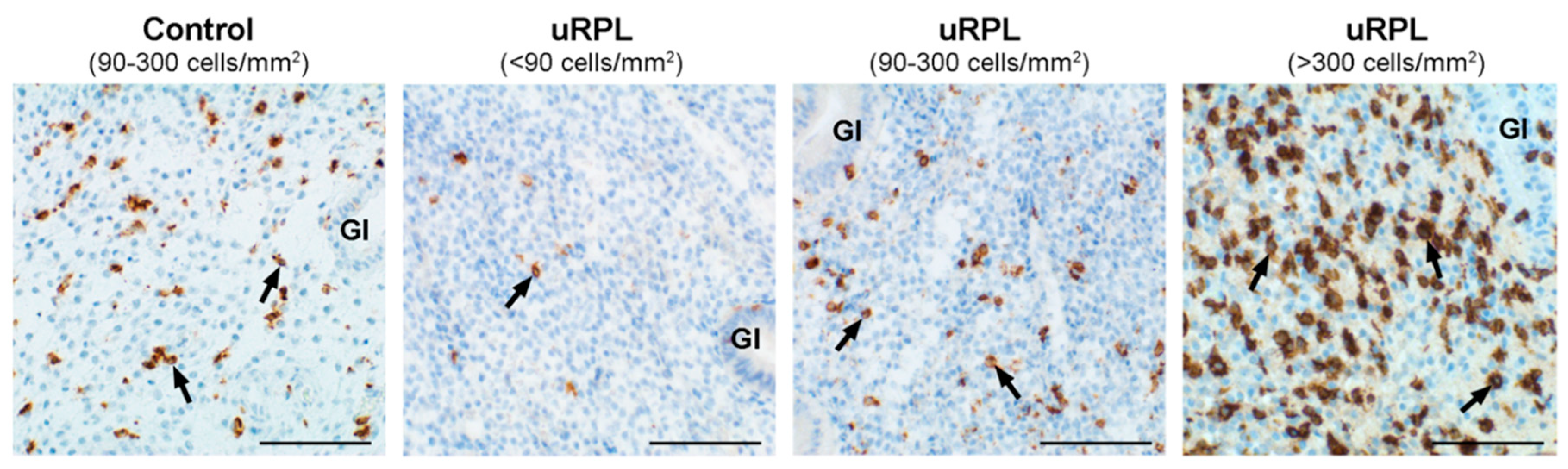
3.3. CD16
3.4. CD57
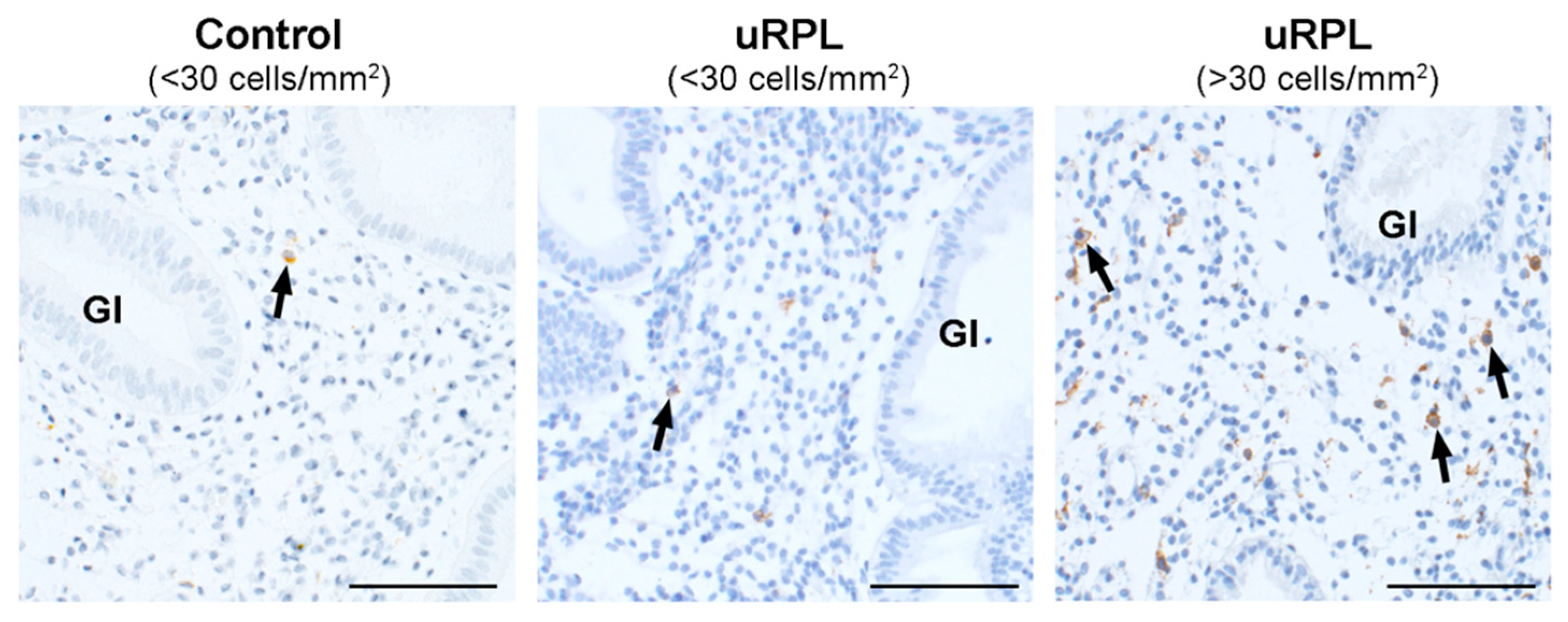
3.5. CD56/CD45 Correlation and Ratio
3.6. CD16/CD56 Correlation and Ratio
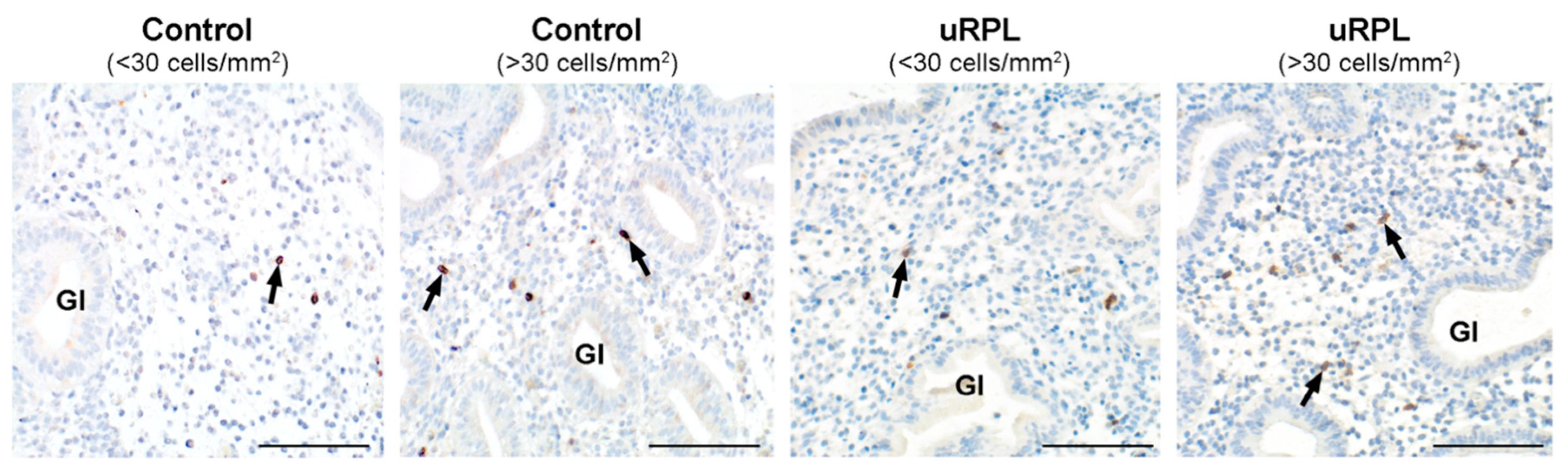
3.7. CD57/CD56 Correlation and Ratio
3.8. CD16/CD57 Correlation and Ratio
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CMV | cytomegalovirus |
| DAB | 3,3′-diaminobenzidine |
| HHV6 | human herpes virus 6 |
| IVig | intravenous immunoglobulin |
| KIR | killer cell Ig-like receptors |
| NCR | natural cytotoxicity receptors |
| MIP-1β | macrophage inflammatory protein 1β |
| NK | natural killer cells |
| uNK | uterine natural killer cells |
| NCAM | neural cell adhesion molecule |
| RIF | recurrent implantation failure |
| RPL | recurrent pregnancy loss |
| RT | room temperature |
| TBST | Tris-buffered saline-Tween20 |
| uRPL | unexplained recurrent pregnancy loss |
| uRPL-CD56high | uRPL subgroup of high uNK cell counts |
| uRPL-CD56low | uRPL subgroup of low uNK cell counts |
| uRPL-CD56normal | uRPL subgroup of normal uNK cell counts |
References
- Practice Committee of the American Society for Reproductive Medicine. Evaluation and treatment of recurrent pregnancy loss: A committee opinion. Fertil. Steril. 2012, 98, 1103–1111. [Google Scholar] [CrossRef] [PubMed]
- El Hachem, H.; Crepaux, V.; May-Panloup, P.; Descamps, P.; Legendre, G.; Bouet, P.-E. Recurrent pregnancy loss: Current perspectives. Int. J. Womens Health 2017, 9, 331–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ESHRE Guideline Group on RPL; Atik, R.B.; Christiansen, O.B.; Elson, J.; Kolte, A.M.; Lewis, S.; Middeldorp, S.; Nelen, W.; Peramo, B.; Quenby, S.; et al. ESHRE guideline: Recurrent pregnancy loss. Hum. Reprod. Open 2018, 2018. [Google Scholar] [CrossRef]
- Kolte, A.M.; Olsen, L.R.; Mikkelsen, E.M.; Christiansen, O.B.; Nielsen, H.S. Depression and emotional stress is highly prevalent among women with recurrent pregnancy loss. Hum. Reprod. 2015, 30, 777–782. [Google Scholar] [CrossRef] [PubMed]
- Carrington, B.; Sacks, G.; Regan, L. Recurrent miscarriage: Pathophysiology and outcome. Curr. Opin. Obstet. Gynecol. 2005, 17, 591–597. [Google Scholar] [CrossRef] [PubMed]
- Ford, H.B.; Schust, D.J. Recurrent pregnancy loss: Etiology, diagnosis, and therapy. Rev. Obstet. Gynecol. 2009, 2, 76–83. [Google Scholar] [PubMed]
- Wang, N.F.; Kolte, A.M.; Larsen, E.C.; Nielsen, H.S.; Christiansen, O.B. Immunologic Abnormalities, Treatments, and Recurrent Pregnancy Loss: What Is Real and What Is Not? Clin. Obstet. Gynecol. 2016, 59, 509–523. [Google Scholar] [CrossRef] [PubMed]
- Saito, S.; Nakashima, A.; Shima, T. Future directions of studies for recurrent miscarriage associated with immune etiologies. J. Reprod. Immunol. 2011, 90, 91–95. [Google Scholar] [CrossRef]
- Clifford, K.; Flanagan, A.M.; Regan, L. Endometrial CD56+ natural killer cells in women with recurrent miscarriage: A histomorphometric study. Hum. Reprod. 1999, 14, 2727–2730. [Google Scholar] [CrossRef] [Green Version]
- Quenby, S.; Bates, M.; Doig, T.; Brewster, J.; Lewis-Jones, D.I.; Johnson, P.M.; Vince, G. Pre-implantation endometrial leukocytes in women with recurrent miscarriage. Hum. Reprod. 1999, 14, 2386–2391. [Google Scholar] [CrossRef]
- Russell, P.; Sacks, G.; Tremellen, K.; Gee, A. The distribution of immune cells and macrophages in the endometrium of women with recurrent reproductive failure. III: Further observations and reference ranges. Pathology 2013, 45, 393–401. [Google Scholar] [CrossRef] [PubMed]
- Kuon, R.-J.; Weber, M.; Heger, J.; Santillán, I.; Vomstein, K.; Bär, C.; Strowitzki, T.; Markert, U.R.; Toth, B. Uterine natural killer cells in patients with idiopathic recurrent miscarriage. Am. J. Reprod. Immunol. 2017, 78. [Google Scholar] [CrossRef] [PubMed]
- Erlebacher, A. Immunology of the maternal-fetal interface. Annu. Rev. Immunol. 2013, 31, 387–411. [Google Scholar] [CrossRef] [PubMed]
- Arck, P.C.; Hecher, K. Fetomaternal immune cross-talk and its consequences for maternal and offspring’s health. Nat. Med. 2013, 19, 548–556. [Google Scholar] [CrossRef]
- Mor, G.; Aldo, P.; Alvero, A.B. The unique immunological and microbial aspects of pregnancy. Nat. Rev. Immunol. 2017, 17, 469–482. [Google Scholar] [CrossRef] [PubMed]
- Bulmer, J.N.; Morrison, L.; Longfellow, M.; Ritson, A.; Pace, D. Granulated lymphocytes in human endometrium: Histochemical and immunohistochemical studies. Hum. Reprod. 1991, 6, 791–798. [Google Scholar] [CrossRef]
- Flynn, L.; Byrne, B.; Carton, J.; Kelehan, P.; O’Herlihy, C.; O’Farrelly, C. Menstrual cycle dependent fluctuations in NK and T-lymphocyte subsets from non-pregnant human endometrium. Am. J. Reprod. Immunol. 2000, 43, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Williams, P.J.; Searle, R.F.; Robson, S.C.; Innes, B.A.; Bulmer, J.N. Decidual leucocyte populations in early to late gestation normal human pregnancy. J. Reprod. Immunol. 2009, 82, 24–31. [Google Scholar] [CrossRef]
- King, A.; Balendran, N.; Wooding, P.; Carter, N.P.; Loke, Y.W. CD3- leukocytes present in the human uterus during early placentation: Phenotypic and morphologic characterization of the CD56++ population. Dev. Immunol. 1991, 1, 169–190. [Google Scholar] [CrossRef] [Green Version]
- Koopman, L.A.; Kopcow, H.D.; Rybalov, B.; Boyson, J.E.; Orange, J.S.; Schatz, F.; Masch, R.; Lockwood, C.J.; Schachter, A.D.; Park, P.J.; et al. Human decidual natural killer cells are a unique NK cell subset with immunomodulatory potential. J. Exp. Med. 2003, 198, 1201–1212. [Google Scholar] [CrossRef] [Green Version]
- Moffett, A.; Colucci, F. Uterine NK cells: Active regulators at the maternal-fetal interface. J. Clin. Investig. 2014, 124, 1872–1879. [Google Scholar] [CrossRef] [PubMed]
- Sojka, D.K.; Yang, L.; Yokoyama, W.M. Uterine natural killer cells: To protect and to nurture. Birth Defects Res. 2018, 110, 1531–1538. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, A.P.; Gerber, S.A.; Croy, B.A. Uterine natural killer cells pace early development of mouse decidua basalis. Mol. Hum. Reprod. 2014, 20, 66–76. [Google Scholar] [CrossRef] [PubMed]
- Brighton, P.J.; Maruyama, Y.; Fishwick, K.; Vrljicak, P.; Tewary, S.; Fujihara, R.; Muter, J.; Lucas, E.S.; Yamada, T.; Woods, L.; et al. Clearance of senescent decidual cells by uterine natural killer cells in cycling human endometrium. eLife 2017, 6, e31274. [Google Scholar] [CrossRef] [PubMed]
- Hanna, J.; Goldman-Wohl, D.; Hamani, Y.; Avraham, I.; Greenfield, C.; Natanson-Yaron, S.; Prus, D.; Cohen-Daniel, L.; Arnon, T.I.; Manaster, I.; et al. Decidual NK cells regulate key developmental processes at the human fetal-maternal interface. Nat. Med. 2006, 12, 1065–1074. [Google Scholar] [CrossRef]
- Hiby, S.E.; Apps, R.; Sharkey, A.M.; Farrell, L.E.; Gardner, L.; Mulder, A.; Claas, F.H.; Walker, J.J.; Redman, C.C.; Morgan, L.; et al. Maternal activating KIRs protect against human reproductive failure mediated by fetal HLA-C2. J. Clin. Investig. 2010, 120, 4102–4110. [Google Scholar] [CrossRef]
- Robson, A.; Harris, L.K.; Innes, B.A.; Lash, G.E.; Aljunaidy, M.M.; Aplin, J.D.; Baker, P.N.; Robson, S.C.; Bulmer, J.N. Uterine natural killer cells initiate spiral artery remodeling in human pregnancy. FASEB J. 2012, 26, 4876–4885. [Google Scholar] [CrossRef]
- Lash, G.E.; Otun, H.A.; Innes, B.A.; Percival, K.; Searle, R.F.; Robson, S.C.; Bulmer, J.N. Regulation of extravillous trophoblast invasion by uterine natural killer cells is dependent on gestational age. Hum. Reprod. 2010, 25, 1137–1145. [Google Scholar] [CrossRef] [Green Version]
- Croy, B.A.; Chen, Z.; Hofmann, A.P.; Lord, E.M.; Sedlacek, A.L.; Gerber, S.A. Imaging of vascular development in early mouse decidua and its association with leukocytes and trophoblasts. Biol. Reprod. 2012, 87, 125. [Google Scholar] [CrossRef]
- Tilburgs, T.; Evans, J.H.; Crespo, Â.C.; Strominger, J.L. The HLA-G cycle provides for both NK tolerance and immunity at the maternal-fetal interface. Proc. Natl. Acad. Sci. USA 2015, 112, 13312–13317. [Google Scholar] [CrossRef] [Green Version]
- Vacca, P.; Pietra, G.; Falco, M.; Romeo, E.; Bottino, C.; Bellora, F.; Prefumo, F.; Fulcheri, E.; Venturini, P.L.; Costa, M.; et al. Analysis of natural killer cells isolated from human decidua: Evidence that 2B4 (CD244) functions as an inhibitory receptor and blocks NK-cell function. Blood 2006, 108, 4078–4085. [Google Scholar] [CrossRef] [PubMed]
- Gaynor, L.M.; Colucci, F. Uterine Natural Killer Cells: Functional Distinctions and Influence on Pregnancy in Humans and Mice. Front. Immunol. 2017, 8, 467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Mariee, N.; Jiang, L.; Liu, Y.; Wang, C.C.; Li, T.C.; Laird, S. Measurement of uterine natural killer cell percentage in the periimplantation endometrium from fertile women and women with recurrent reproductive failure: Establishment of a reference range. Am. J. Obstet. Gynecol. 2017, 217, 680.e1–680.e6. [Google Scholar] [CrossRef] [PubMed]
- Collins, P.L.; Cella, M.; Porter, S.I.; Li, S.; Gurewitz, G.L.; Hong, H.S.; Johnson, R.P.; Oltz, E.M.; Colonna, M. Gene Regulatory Programs Conferring Phenotypic Identities to Human NK Cells. Cell 2019, 176, 348–360. [Google Scholar] [CrossRef] [Green Version]
- Di Vito, C.; Mikulak, J.; Mavilio, D. On the Way to Become a Natural Killer Cell. Front. Immunol. 2019, 10. [Google Scholar] [CrossRef]
- Vassiliadou, N.; Bulmer, J.N. Immunohistochemical evidence for increased numbers of ‘classic’ CD57+ natural killer cells in the endometrium of women suffering spontaneous early pregnancy loss. Hum. Reprod. 1996, 11, 1569–1574. [Google Scholar] [CrossRef] [Green Version]
- Junovich, G.; Azpiroz, A.; Incera, E.; Ferrer, C.; Pasqualini, A.; Gutierrez, G. Endometrial CD16(+) and CD16(-) NK cell count in fertility and unexplained infertility. Am. J. Reprod. Immunol. 2013, 70, 182–189. [Google Scholar] [CrossRef]
- Giuliani, E.; Parkin, K.L.; Lessey, B.A.; Young, S.L.; Fazleabas, A.T. Characterization of uterine NK cells in women with infertility or recurrent pregnancy loss and associated endometriosis. Am. J. Reprod. Immunol. 2014, 72, 262–269. [Google Scholar] [CrossRef] [Green Version]
- Vacca, P.; Cantoni, C.; Prato, C.; Fulcheri, E.; Moretta, A.; Moretta, L.; Mingari, M.C. Regulatory role of NKp44, NKp46, DNAM-1 and NKG2D receptors in the interaction between NK cells and trophoblast cells. Evidence for divergent functional profiles of decidual versus peripheral NK cells. Int. Immunol. 2008, 20, 1395–1405. [Google Scholar] [CrossRef] [Green Version]
- Kwak-Kim, J.; Bao, S.; Lee, S.K.; Kim, J.W.; Gilman-Sachs, A. Immunological modes of pregnancy loss: Inflammation, immune effectors, and stress. Am. J. Reprod. Immunol. 2014, 72, 129–140. [Google Scholar] [CrossRef]
- Bayer-Garner, I.B.; Nickell, J.A.; Korourian, S. Routine syndecan-1 immunohistochemistry aids in the diagnosis of chronic endometritis. Arch. Pathol. Lab. Med. 2004, 128, 1000–1003. [Google Scholar] [CrossRef] [PubMed]
- McQueen, D.B.; Perfetto, C.O.; Hazard, F.K.; Lathi, R.B. Pregnancy outcomes in women with chronic endometritis and recurrent pregnancy loss. Fertil. Steril. 2015, 104, 927–931. [Google Scholar] [CrossRef] [PubMed]
- Romero, R.; Espinoza, J.; Mazor, M. Can endometrial infection/inflammation explain implantation failure, spontaneous abortion, and preterm birth after in vitro fertilization? Fertil. Steril. 2004, 82, 799–804. [Google Scholar] [CrossRef] [PubMed]
- Kitaya, K.; Takeuchi, T.; Mizuta, S.; Matsubayashi, H.; Ishikawa, T. Endometritis: New time, new concepts. Fertil. Steril. 2018, 110, 344–350. [Google Scholar] [CrossRef]
- Matteo, M.; Cicinelli, E.; Greco, P.; Massenzio, F.; Baldini, D.; Falagario, T.; Rosenberg, P.; Castellana, L.; Specchia, G.; Liso, A. Abnormal pattern of lymphocyte subpopulations in the endometrium of infertile women with chronic endometritis. Am. J. Reprod. Immunol. 2009, 61, 322–329. [Google Scholar] [CrossRef]
- Ticconi, C.; Pietropolli, A.; Di Simone, N.; Piccione, E.; Fazleabas, A. Endometrial Immune Dysfunction in Recurrent Pregnancy Loss. Int. J. Mol. Sci. 2019, 20, 5332. [Google Scholar] [CrossRef] [Green Version]
- Kitaya, K.; Matsubayashi, H.; Yamaguchi, K.; Nishiyama, R.; Takaya, Y.; Ishikawa, T.; Yasuo, T.; Yamada, H. Chronic Endometritis: Potential Cause of Infertility and Obstetric and Neonatal Complications. Am. J. Reprod. Immunol. 2016, 75, 13–22. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, X.; Huang, J.; Wang, C.C.; Yu, M.Y.; Laird, S.; Li, T.C. Comparison of the prevalence of chronic endometritis as determined by means of different diagnostic methods in women with and without reproductive failure. Fertil. Steril. 2018, 109, 832–839. [Google Scholar] [CrossRef] [Green Version]
- Sacks, G. Enough! Stop the arguments and get on with the science of natural killer cell testing. Hum. Reprod. 2015, 30, 1526–1531. [Google Scholar] [CrossRef] [Green Version]
- Tuckerman, E.; Laird, S.M.; Prakash, A.; Li, T.C. Prognostic value of the measurement of uterine natural killer cells in the endometrium of women with recurrent miscarriage. Hum. Reprod. 2007, 22, 2208–2213. [Google Scholar] [CrossRef]
- Moffett, A.; Shreeve, N. First do no harm: Uterine natural killer (NK) cells in assisted reproduction. Hum. Reprod. 2015, 30, 1519–1525. [Google Scholar] [CrossRef] [PubMed]
- Donoghue, J.F.; Paiva, P.; Teh, W.T.; Cann, L.M.; Nowell, C.; Rees, H.; Bittinger, S.; Obers, V.; Bulmer, J.N.; Stern, C.; et al. Endometrial uNK cell counts do not predict successful implantation in an IVF population. Hum. Reprod. 2019, 34, 2456–2466. [Google Scholar] [CrossRef] [PubMed]
- Tsujimoto, H.; Uchida, T.; Efron, P.A.; Scumpia, P.O.; Verma, A.; Matsumoto, T.; Tschoeke, S.K.; Ungaro, R.F.; Ono, S.; Seki, S.; et al. Flagellin enhances NK cell proliferation and activation directly and through dendritic cell-NK cell interactions. J. Leukoc. Biol. 2005, 78, 888–897. [Google Scholar] [CrossRef] [PubMed]
- Yamin, R.; Berhani, O.; Peleg, H.; Aamar, S.; Stein, N.; Gamliel, M.; Hindi, I.; Scheiman-Elazary, A.; Gur, C. High percentages and activity of synovial fluid NK cells present in patients with advanced stage active Rheumatoid Arthritis. Sci. Rep. 2019, 9, 1351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mariee, N.; Li, T.C.; Laird, S.M. Expression of leukaemia inhibitory factor and interleukin 15 in endometrium of women with recurrent implantation failure after IVF; correlation with the number of endometrial natural killer cells. Hum. Reprod. 2012, 27, 1946–1954. [Google Scholar] [CrossRef] [Green Version]
- Comba, C.; Bastu, E.; Dural, O.; Yasa, C.; Keskin, G.; Ozsurmeli, M.; Buyru, F.; Serdaroglu, H. Role of inflammatory mediators in patients with recurrent pregnancy loss. Fertil. Steril. 2015, 104, 1467–1474. [Google Scholar] [CrossRef]
- D’Ippolito, S.; Tersigni, C.; Marana, R.; Di Nicuolo, F.; Gaglione, R.; Rossi, E.D.; Castellani, R.; Scambia, G.; Di Simone, N. Inflammosome in the human endometrium: Further step in the evaluation of the “maternal side”. Fertil. Steril. 2016, 105, 111–118. [Google Scholar] [CrossRef] [Green Version]
- Kitaya, K.; Yamaguchi, T.; Yasuo, T.; Okubo, T.; Honjo, H. Post-ovulatory rise of endometrial CD16(−) natural killer cells: In situ proliferation of residual cells or selective recruitment from circulating peripheral blood? J. Reprod. Immunol. 2007, 76, 45–53. [Google Scholar] [CrossRef]
- Wu, Y.; Tian, Z.; Wei, H. Developmental and Functional Control of Natural Killer Cells by Cytokines. Front. Immunol. 2017, 8. [Google Scholar] [CrossRef]
- Berbic, M.; Fraser, I.S. Immunology of normal and abnormal menstruation. Womens Health 2013, 9, 387–395. [Google Scholar] [CrossRef] [Green Version]
- Lash, G.E.; Bulmer, J.N.; Li, T.C.; Innes, B.A.; Mariee, N.; Patel, G.; Sanderson, J.; Quenby, S.; Laird, S.M. Standardisation of uterine natural killer (uNK) cell measurements in the endometrium of women with recurrent reproductive failure. J. Reprod. Immunol. 2016, 116, 50–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marci, R.; Gentili, V.; Bortolotti, D.; Lo Monte, G.; Caselli, E.; Bolzani, S.; Rotola, A.; Di Luca, D.; Rizzo, R. Presence of HHV-6A in Endometrial Epithelial Cells from Women with Primary Unexplained Infertility. PLoS ONE 2016, 11, e0158304. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, V.D.; Falconer, K.; Björkström, N.K.; Blom, K.G.; Weiland, O.; Ljunggren, H.-G.; Alaeus, A.; Sandberg, J.K. Expansion of functionally skewed CD56-negative NK cells in chronic hepatitis C virus infection: Correlation with outcome of pegylated IFN-alpha and ribavirin treatment. J. Immunol. 2009, 183, 6612–6618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiesa, M.D.; Falco, M.; Podestà, M.; Locatelli, F.; Moretta, L.; Frassoni, F.; Moretta, A. Phenotypic and functional heterogeneity of human NK cells developing after umbilical cord blood transplantation: A role for human cytomegalovirus? Blood 2012, 119, 399–410. [Google Scholar] [CrossRef]
- Müller-Durovic, B.; Grählert, J.; Devine, O.P.; Akbar, A.N.; Hess, C. CD56-negative NK cells with impaired effector function expand in CMV and EBV co-infected healthy donors with age. Aging (Albany NY) 2019, 11, 724–740. [Google Scholar] [CrossRef]
- Long, E.O.; Kim, H.S.; Liu, D.; Peterson, M.E.; Rajagopalan, S. Controlling natural killer cell responses: Integration of signals for activation and inhibition. Annu. Rev. Immunol. 2013, 31, 227–258. [Google Scholar] [CrossRef] [Green Version]
- Chester, C.; Fritsch, K.; Kohrt, H.E. Natural Killer Cell Immunomodulation: Targeting Activating, Inhibitory, and Co-stimulatory Receptor Signaling for Cancer Immunotherapy. Front. Immunol. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Bansal, A.S.; Bajardeen, B.; Thum, M.Y. The basis and value of currently used immunomodulatory therapies in recurrent miscarriage. J. Reprod. Immunol. 2012, 93, 41–51. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Species | Type | Supplier | Dilution |
|---|---|---|---|---|
| CD16 | Mouse | Monoclonal—DJ130 | Santa Cruz | 1:200 |
| CD45 | Mouse | Monoclonal—2B11 + PD7/26 | Dako | 1:200 |
| CD56 | Mouse | Monoclonal—123C3 | Dako | 1:100 |
| CD57 | Mouse | Monoclonal—TB01 | Dako | 1:100 |
| CD138 | Mouse | Monoclonal—MI15 | Dako | 1:100 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiokadze, M.; Bär, C.; Pastuschek, J.; Dons’koi, B.V.; Khazhylenko, K.G.; Schleußner, E.; Markert, U.R.; Favaro, R.R. Beyond Uterine Natural Killer Cell Numbers in Unexplained Recurrent Pregnancy Loss: Combined Analysis of CD45, CD56, CD16, CD57, and CD138. Diagnostics 2020, 10, 650. https://doi.org/10.3390/diagnostics10090650
Chiokadze M, Bär C, Pastuschek J, Dons’koi BV, Khazhylenko KG, Schleußner E, Markert UR, Favaro RR. Beyond Uterine Natural Killer Cell Numbers in Unexplained Recurrent Pregnancy Loss: Combined Analysis of CD45, CD56, CD16, CD57, and CD138. Diagnostics. 2020; 10(9):650. https://doi.org/10.3390/diagnostics10090650
Chicago/Turabian StyleChiokadze, Maia, Christin Bär, Jana Pastuschek, Boris V. Dons’koi, Kseniia G. Khazhylenko, Ekkehard Schleußner, Udo R. Markert, and Rodolfo R. Favaro. 2020. "Beyond Uterine Natural Killer Cell Numbers in Unexplained Recurrent Pregnancy Loss: Combined Analysis of CD45, CD56, CD16, CD57, and CD138" Diagnostics 10, no. 9: 650. https://doi.org/10.3390/diagnostics10090650