
Macrobenthos of the Tortolì Lagoon: A Peculiar Case of High Benthic Biodiversity among Mediterranean Lagoons


1
CNR-IAS, National Research Council—Institute for the Study of Anthropic Impact and Sustainability in the Marine Environment, Località Sa Mardini, Torregrande, 09170 Oristano, Italy
2
Department of Biology, University of Rome “Tor Vergata”, 00133 Rome, Italy
3
CoNISMa (Consorzio Nazionale Interuniversitario per le Scienze del Mare), 00196 Rome, Italy
*
Author to whom correspondence should be addressed.
Diversity 2023, 15(6), 783; https://doi.org/10.3390/d15060783
Submission received: 19 May 2023
/
Revised: 14 June 2023
/
Accepted: 15 June 2023
/
Published: 16 June 2023
(This article belongs to the Special Issue Investigation of the Biology, Ecology, Distribution of Marine Macrozoobenthic Communities)
Abstract
:Coastal lagoons and brackish ponds are extremely dynamic and temporary ecosystems that follow natural changes throughout their geological history. The correct management of the lagoons ensures their integrity and proper functioning. For this reason, their ecological status should be surveyed for assessing the most appropriate strategies of use. In the present study, historical datasets collected in 2003–2004 are used to investigate the spatiotemporal variation in the species composition and community structure of the macrobenthos of the Tortolì Lagoon (Sardinia, Italy) and to assess their relationship with key environmental variables. Owing to the presence of a riverine runoff at a site and confined areas at some distance from the sea inlet, we hypothesize the marked spatiotemporal changes of the macrobenthic community consistent with the high environmental variability typical of coastal lagoons. The results show a surprisingly high benthic biodiversity for a medium-sized lagoon (250 ha), with 101 species unevenly distributed across the lagoon. The environmental variables did not explain the zonation of the macrobenthic community as that typically found along a lagoonal gradient, due to a marked marine influence. The sampling sites were in fact discriminated by the species distribution according to their ecological affinity; in particular, the most distinctive characteristics of the Tortolì Lagoon emerged from the strictly marine species that represented the most abundant group, consistently with the high marinization of the lagoon. Our results show that the Tortolì Lagoon constitutes a peculiar ecosystem within Mediterranean lagoons, departing from the classic confinement theory.
1. Introduction
Transitional waters, including coastal lagoons, brackish ponds, and estuaries, are important and fragile ecosystems in the coastal landscape, and the scientific community and worldwide legislations (e.g., the US Clean Water Act, European Water Framework Directive, and Marine Strategy Framework Directive) recognize the necessities to assess their health status and ensure the proper management of their resources. Within the Mediterranean Sea, there are numerous transitional water ecosystems that are exploited for productive purposes, such as aquaculture and fisheries [1,2], and are interesting conservation sites for flora and fauna [3]. Coastal lagoons and brackish ponds are especially extremely dynamic and temporary ecosystems [4], which follow natural changes from their formation, evolution, and subsequent decline, throughout their geological history. Typically, their fate is closely connected to the presence of barriers enclosing them and cutting them off partially from the sea; on the other hand, the lagoons may be largely influenced by freshwater inflows from the continental system. Meteorological and climatic factors, e.g., winds, waves, and currents, further influence the history of the lagoons and determine their shape. This involves the great diversity of lagoonal environments, both in time and space [5,6], which is also reflected on the peculiarity of their flora and fauna [7,8,9,10,11,12,13].
Human activities are also increasingly contributing to the fate of the lagoons by reclamation works, regulating sea and freshwater inflows, and modifying the hydrographic regime, based on the intended use of the lagoons by human beings [14,15]. Their use depends on the key ecosystem services they provide for both productive and conservation purposes, such as water quality improvement, fisheries resources, habitat and food for migratory and resident animals, and recreational areas for human populations [16,17,18,19,20]. By contrast, the lagoonal ecosystems are under various pressures due to anthropogenic impacts that endanger them and may cause the reduction in or even loss of their services and functions [21,22,23,24]. Consequently, the scientific communities and stakeholders have taken care to manage lagoons in order to maintain their integrity and functioning, which contrasts with the natural fate of lagoons, in order to ensure the numerous ecosystem services that lagoons provide. In this context, it is worthy of attention to detect changes to see if they are part of natural variability or if they presage a system malfunction [25,26].
A useful tool to understand environmental changes is the analysis of historical data, when available, to help us distinguish periodical fluctuations from sudden/short-term variations [27,28,29]. The lack of knowledge about past conditions, in parallel with the gradual change in the human perception of environmental conditions, often results in a growing tolerance to environmental degradation, which leads to harmful consequences on environmental and cultural heritage sites and their conservation [30,31,32,33,34]. In this context, the composition and abundance of benthic invertebrate fauna are one of the main biological elements used to assess ecological quality status, and it is also known to be one the best effective indicators of pollution stress, as it shows predictive responses to different levels of anthropogenic impacts [35,36,37,38]. In coastal lagoons, macrobenthic assemblages show a typical zonation, with the marine species more abundant at the seaward sites and brackish and opportunistic species in the confined zone [11,12].
From these general concepts, we use historical datasets collected in the Tortolì Lagoon (Sardinia, Italy) to investigate the species composition and community structure of the macrobenthos and to assess their relationship with key environmental variables. As is typical of coastal lagoons, we hypothesize that the marked spatiotemporal changes in the macrobenthos are driven by the environmental variability typical of these systems. In particular, we aim (i) to analyze the variations in macrobenthic community structure and diversity in order to detect a possible spatial zonation; (ii) to investigate the consistency between the spatial pattern of benthos and that based on the environmental features of the lagoon; and (iii) to evaluate the possible causes influencing the species composition, diversity, and distribution of the macrobenthos within the investigated lagoon.
2. Materials and Methods
2.1. Study Area
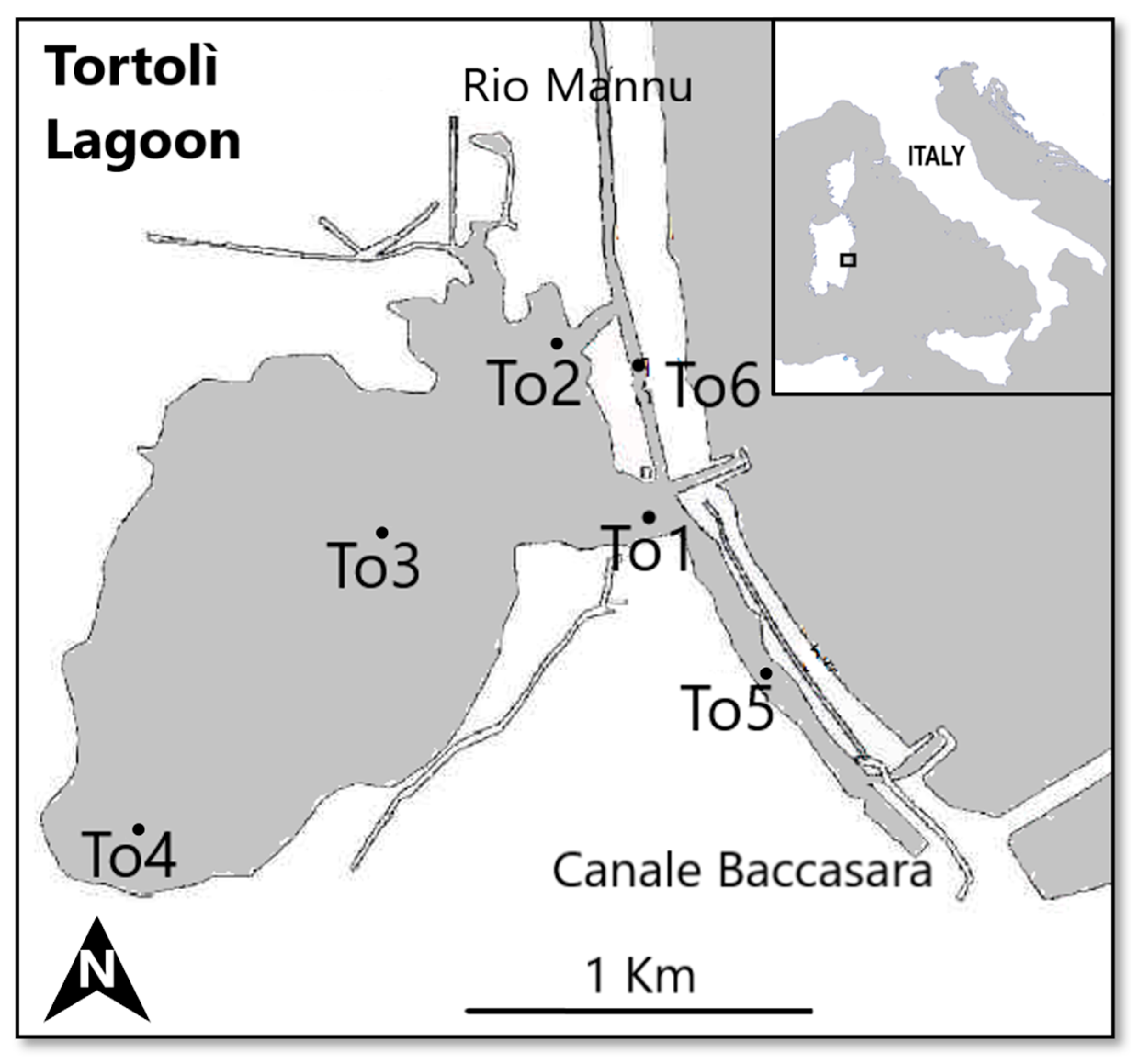
The Tortolì Lagoon, located in the central-east coast of Sardinia, has a total area of approximately 250 ha and a perimeter of 15.1 Km (Figure 1); the average depth of the water is 1.5 m. It is composed of an ovoid basin connected to the sea through two channels in its central and southern areas, while the freshwater inputs enter the lagoon from Rio Mannu through a bulkhead system in its northern area and a small canal in the southwest area. Bottoms are sandy–muddy and large areas are covered with seagrasses, mainly Zostera noltei. The lagoon hosts a flourishing fish and shellfish traditional fishery, and aquaculture activities have been practiced for more than 30 years [39].
For the present study, six sites (Figure 1) assumed to be representative of different environmental conditions according to the confinement theory (e.g., salinity, sediment grain-size, etc.) were selected. Three sites (To2, To3, To4) were located in the internal area of the lagoon and had sandy sediments; To1 and To5 were located, respectively, near the central and southern parts of the seaward channel, and both had muddy–sandy sediments; and To6 was located in Rio Mannu and had sandy–muddy sediments.
2.2. Sampling Methods and Taxonomical Analysis
The macrozoobenthos and water samples were collected at the six sites (Figure 1) in July and November 2003 and in February and June 2004; two replicates of macrozoobenthos were collected at each site to obtain a total of 48 samples. From the water samples that were collected on the same dates, the nitrates, total phosphorous, and chlorophyll-a levels were measured in the laboratory according to the method described by Parson et al. (1984) [40]. Salinity, temperature, and dissolved oxygen (mg/L) were measured monthly from July 2003 to June 2004 at each site using a multi-parametric probe (OxyGuard Handy Gamma).
The macrozoobenthos samples, collected with a Van Veen grab (sampling area: 500 cm2), were sieved using a 0.5 mm mesh-size sieve and fixed with a 10% buffered formaldehyde solution; when necessary, Rose Bengal was used to better highlight the organisms. In the laboratory, the animals were identified to the lowest possible taxonomic level, counted, and then preserved in 70% ethanol. The updated taxonomic nomenclature was checked by the World Register of Marine Species.
2.3. Data Analysis
Both abiotic and biotic data were analyzed by univariate and multivariate techniques. Principal component analysis (PCA) was performed on log-transformed and normalized abiotic data in order to highlight the variations among sites and sampling periods. Univariate diversity measures, such as average species richness (S), abundance (A), Shannon index (H′), and Evenness (J), were calculated to analyze the community structure and species composition. The non-parametric permutational analysis of variance (two-way PERMANOVA) was conducted to test the significant differences in all the abiotic data (i.e., temperature, salinity, dissolved oxygen, NO3, chlorophyll-a, and PTOT) and the biotic univariate measures (i.e., number of species, total macrofaunal abundance, Shannon diversity, and evenness). Macrozoobenthic data were analyzed by means of the multivariate non-metric multidimensional scaling (nMDS) procedure, using mean abundance values in the cumulative matrix of all the sites and months, based on the Bray–Curtis (dis)similarity measure [41]. Similarity, percentage (SIMPER) analysis [41] with a cut-off value of 95% was used to determine the contribution of each species to the (dis)similarity between sampling sites. The analysis of similarities (ANOSIM) was performed to test significant variations in the macrozoobenthos structure between sites [42]. Univariate and multivariate analyses were performed using PAST 4.05 and PRIMER v.6 software.
3. Results
3.1. Water Variables
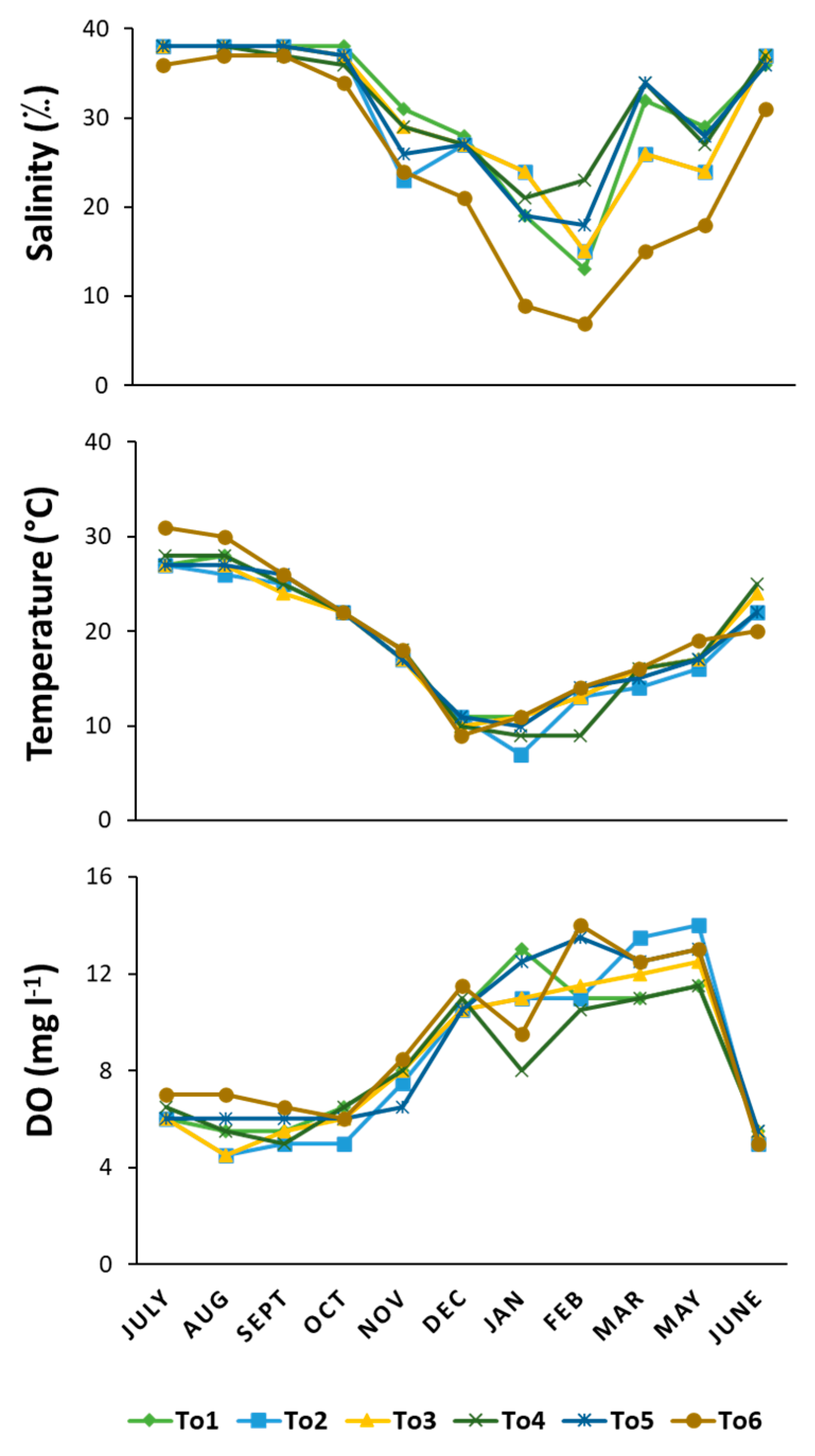
Temperature and dissolved oxygen (DO) showed similar mean values and ranges at all sites, with maximum values of temperature in July and August and minimum values in December and January, while the lowest values of DO were recorded from June to September and the highest values from February to May (Figure 2). Salinity showed relatively higher mean values at the innermost site (To4) and at those closest to the sea (To1 and To5), while the site influenced by the Rio Mannu (To6) showed the widest range of variation in salinity, from 9‰ in winter–spring to 37‰ in summer–autumn.
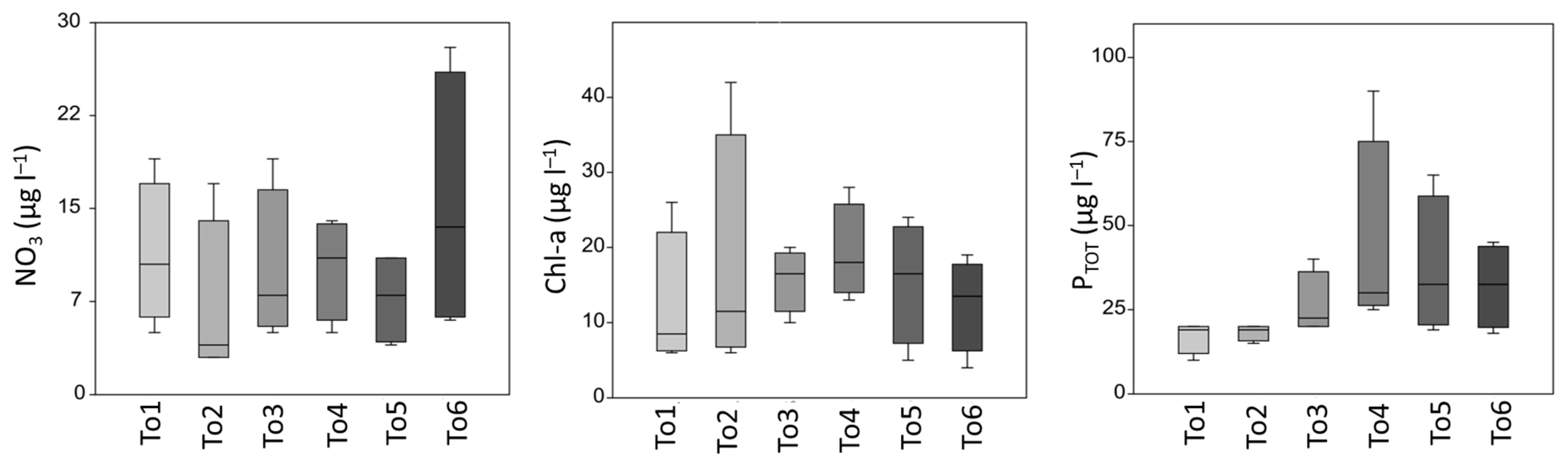
The box plots in Figure 3 show relatively limited variations in the nutrient concentrations, being most apparent for nitrates at To6 and for total phosphorous at To4; the smallest variations were found at To5 for nitrates and at To1, To2, and To3 for total phosphorous. Chlorophyll-a content varied between sites and showed the lowest mean values at site To1 close to the sea and at To2, and in the latter, the greatest range of chlorophyll-a was also recorded; the highest mean value was achieved for in the innermost site at To4.
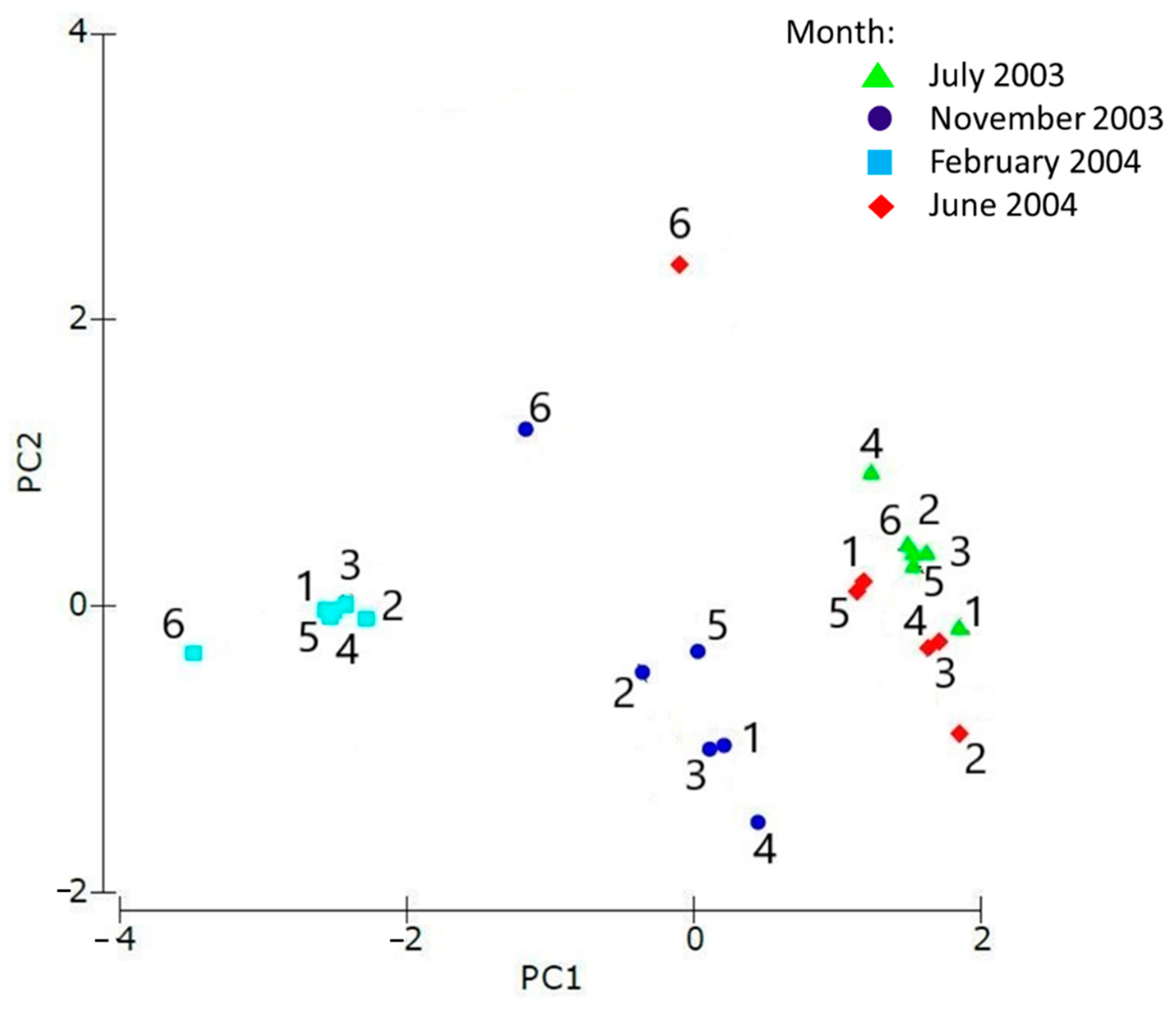
The PCA based on the environmental variables showed a clear separation between different dates (Figure 4). The sampling points for the months of June and July were grouped on the right side of the plot, as opposed to those of February, while the sampling-points of November were located in the intermediate position. At each month, no particular arrangement separated the site points that remained close together within each group, with the exception of To6 in June and November.
PERMANOVA showed no significant differences among the sampling sites for all the abiotic variables (data not shown).
3.2. Macrozoobenthic Community
The macrobenthic fauna found in the Tortolì Lagoon was composed of 101 species, for a total of 2839 individuals. Polychaetes were the richest in number of species (49.5%) and dominant in terms of number of individuals (42.7%), followed by crustaceans (28.4% of species and 34.9% of number of individuals) and mollusks (13.7%). The most abundant polychaetes were the marine species Nephtys hombergii, Myriochele heeri, and Spiochaetopterus costarum reaching the highest densities at To1 and the opportunistic species Polydora ciliata at To2 and To5 and Capitella capitata at To4. Among the crustaceans, the most abundant species were the amphipods with the brackish species Monocorophium insidiosum, which occurred at all the sites, except To1 and To5, but mostly at To4 and To6, and marine Apocorophium acutum recorded throughout the lagoon and Elasmopus pocillimanus especially at To3. The dominant mollusk was the bivalve Abra segmentum, a typical inhabitant of coastal shallow areas, which was found throughout the lagoon, and the brackish species Cerastoderma glaucum, recorded at all the sites, but only occasionally at To1 and To5.
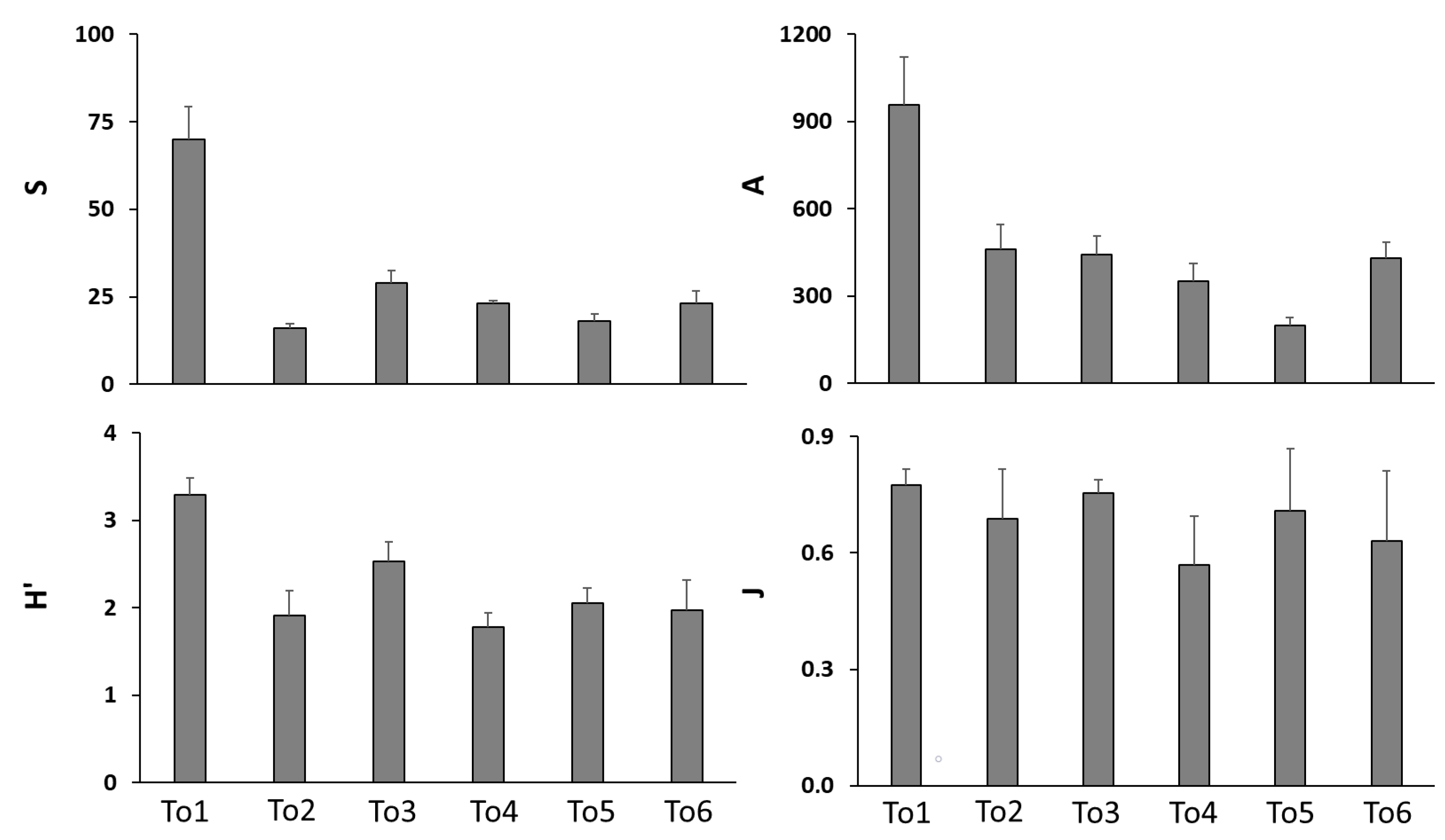
The results of the biotic synthetic variables are shown in Figure 5. The highest values for all four variables (i.e., number of species and individuals, and H′ and J indices) were found at To1; the lowest ones occurred at To2 (number of species), To5 (number of individuals), and To4 (and H′ and J indices).
The two-way PERMANOVA conducted for each biotic measure showed significant differences (p < 0.001) among sites and months in the species number, total abundance, evenness, and diversity index (Table 1).
The nMDS plot (Figure 6) shows the distinction of the sampling sites divided into three groups, based on the macrobenthic community structure. Two-way ANOSIM analysis confirmed significant differences among the sites (R = 0.93; p < 0.0001) and times (R = 0.96; p < 0.0001). The first group of sampling sites included To4 and To6 located at the opposite sides of the lagoon; the second group consisted of the sites To2 and To3 situated in the central area of the lagoon; and the third group consisted of To1 and To5, which were located at the sites closest to the sea.
Table 2 shows the list of the top discriminating species contributing to the 95% by SIMPER analysis at the investigated sites and their ecological affinity. The species were ordered according to their occurrence, and only four species (A. segmentum, A. acutum, C. glaucum, and Heteromastus filiformis) were found at all sites, most species occurring only at two or one site. The marine species group was the most abundant, followed by coastal, opportunistic, and brackish species.
Some distinction in terms of species composition emerged between the sites, the highest number of species being found at To1, the majority of which were typically marine (M. heeri, S. costarum, Aphelochaeta marioni, Microdeutopus algicola, Amphipholis squamata, Phoronis psammophila) or common inhabitants of shallow coastal waters (A. segmentum, Neanthes caudata, Microdeutopus gryllotalpa), while strictly opportunistic species were lacking, save H. filiformis and very few individuals of C. capitata, which occasionally occurred. Some of these species, both marine and common in shallow coastal waters, were shared with the benthic assemblage of To5 (i.e., A. marioni, P. psammophila, M. gryllotalpa, A. segmentum), as well as the opportunistic species H. filiformis, which occurred in To5 with the highest abundance. Many marine and shallow coastal waters species, e.g., N. hombergii, A. acutum, P. psammophila, A. segmentum, together with the opportunistic species Prionospio cirrifera, characterized the benthos at both To2 and To3. Finally, few marine species occurred at To4 and To6, which, instead, shared some brackish-water (i.e., M. insidiosum, C. glaucum) and opportunistic (i.e., C. capitata, Malacoceros fuliginosus, P. cirrifera) species between their benthic assemblages. No typical freshwater elements were recorded in the lagoon, not even at To6 located next to the Rio Mannu freshwater input.
4. Discussion
The present study highlighted a limited range of spatial variations in the environmental features of the Tortolì Lagoon, contrary to a natural gradient typically found in coastal lagoons, i.e., from marine-influenced towards confined areas [12,43]. In particular, water temperature, dissolved oxygen, and salinity showed no significant differences among the sites, even though site To6 was affected by freshwater runoff from the Rio Mannu in the cooler season. Moreover, the other measured water variables only partially contributed to arrange the sampling sites along a trophic gradient, with chlorophyll-a showing slightly higher values and smaller variations at the sites of To3, To4, To5, and To6; total phosphorus at To4, To5, and To6; and nitrate at To6. The absence of confinement and trophic gradients was reflected in the macrobenthos community structure and species composition that did not show a clear zonation. Notwithstanding, PERMANOVA highlighted significant differences in species richness, abundance, diversity, and evenness, both in space and time. These latter factors were attributable to the seasonal cycle of macrozoobenthos assemblages in lagoon ecosystems [44,45,46]. Based on the macrozoobenthos structure data, distinct differences among sites emerged from the nMDS analysis and were supported by the ANOSIM results; however, the measured environmental variables were inconsistent to explain the different groupings of communities. We therefore hypothesized that the causes of these distinctions and spatial heterogeneity lie in the biological factors. Indeed, species of different ecological affinities occurred unevenly at different sites, save the only two brackish species, M. insidiosum and C. glaucum, found all over the lagoon. Half of the opportunistic species were distributed throughout the lagoon, while the other opportunistic ones characterized only two sites, as M. fuliginosus To4 and To6, P. ciliate To1 and To5, and T. dulongii To1 and To2. A similar pattern was revealed with regard to coastal-sheltered and strictly marine species. Of the ten coastal-sheltered species, only three (A. segmentum, M. gryllotalpa, M. palmata) were distributed over the large part of the lagoon, unlike the other species that distinguished two sites or a single site. Most distinctions emerged within the group of strictly marine species, which was also the most species-rich group. Of these, only four species (M. acutum, N. hombergii, P. psammofila, I. trispinosa) were able to entirely colonize the lagoon, while half of the remaining species characterized two sites and the other half a single site. These results suggest, on the one hand, the high biodiversity in the Tortolì Lagoon and, on the other, the marked degree of marinization that characterizes it.
Such peculiarity makes the Tortolì Lagoon a peculiar ecosystem among the Mediterranean transitional water environments, particularly located those along the Sardinian coast [12]. As a result of its physiographic features that characterize it as a medium-sized lagoon (surface of 2.5 km2 and volume of 4 Mm3) and the efficient connection with the sea, the Tortolì Lagoon appeared to be strongly influenced by the sea, except for the sector of the lagoon in close proximity to the main freshwater tributary (Rio Mannu). This observation was confirmed by the composition of the macrozoobenthos assemblages, which included a large number of marine species at each site. Most of the species, typical inhabitants of strictly marine and sheltered coastal areas, mostly characterized the marine site (To1), with some of them, e.g., Oweniidae Myriochele heeri and Chaetopteridae Spiochaetopterus costarum, presenting dense populations as result of their gregarious behavior [47,48]. At the same time, the presence of several marine species, also at the other sites, verified the strong marine influence throughout the lagoon. The very high number of species (101) recorded in the Tortolì Lagoon, despite its limited size, is a unique feature among the Italian brackish-water ecosystems. As an example, in the Santa Giusta Lagoon, with a similar salinity range and shape, and in the Mistras Lagoon, with a similar surface but higher salinity, both located in the same transitional district of Oristano in the central-west coast of Sardinia [49], a total of 59 and 39 species have been recorded, respectively [3,50]. Moreover, in the two lagoons of Varano and Alimini, which are located in the Adriatic district and are similarly influenced by the adjacent sea, a total of 52 and 33 were recorded against a larger extension of the Varano Lagoon and a similar salinity range of the Alimini Lagoon compared to the Tortolì Lagoon [51]. We hypothesized that the peculiarity of the Tortolì Lagoon was attributable to its physiographic features and the marked connection with the sea, which make it suitable for the entry of a large number of marine species. In fact, these species find environmental conditions suitable for their settlement heterogeneously across the whole lagoon basin, producing a scattered distribution instead of a distribution along the typical confinement gradient. Moreover, thanks to its physiographic and hydrological peculiarities, the Tortolì Lagoon has been exploited for aquaculture purposes, particularly the experimental oyster farming that started in the early 1990s after the decline in lagoonal fish production since the 1980s [52]. During these years, salinity has increased as a result of various interventions to improve the water circulation, and the changed environmental conditions resulted in conditions suitable for oyster farming [53]. In fact, such production continued successfully for the next twenty years, until the time of the present study (2003), when the oyster production yields, added to those of fish, reached the levels found before the decline in fish production (the author’s unpublished data). These results demonstrate that the macrozoobenthos community in the Tortolì Lagoon has not been adversely affected by high-quality bivalve culture.
5. Conclusions
The present study showed the absence of zonation of the macrobenthic community driven by confinement and trophic gradients, as is typically found in Mediterranean lagoons. This was due to a marked marine influence and the dominance of strictly marine species, which contributed to an unusually high benthic biodiversity level. The peculiarity of the Tortolì Lagoon, among the Mediterranean lagoons, and the macrobenthic community analyzed here may represent a useful baseline for the future assessment and comparison of past and present environmental conditions in this and other lagoon systems.
Author Contributions
Conceptualization, J.G., A.S., P.M. and M.F.G.; metodology, J.G. and M.F.G.; formal analysis J.G. and A.S.; data curation, J.G., A.S., P.M. and M.F.G.; investigation, J.G., A.S., P.M. and M.F.G.; writing—original draft preparation, J.G., A.S., P.M. and M.F.G.; writing—review and editing, J.G., A.S., P.M. and M.F.G.; supervision, J.G., A.S., P.M. and M.F.G. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Graham, P.; Falconer, L.; Telfer, T.; Mossone, P.; Viale, I.; Carboni, S. A modelling approach to classify the suitability of shallow Mediterranean lagoons for pacific oyster, Crassostrea gigas (Thunberg, 1793) farming. Ocean Coast. Manag. 2020, 192, 105234. [Google Scholar] [CrossRef]
- Vallainc, D.; Concu, D.; Papiol, G.G.; Loi, B.; Leggieri, F.; Brundu, G.; Chindris, A.; Sanna, G.; Fois, N.; Antognarelli, F.; et al. Producing flat-head grey mullet Mugil cephalus (Linnaeus, 1758) fries in captivity from sexually mature adults collected in Sardinian lagoons. Aquac. Rep. 2021, 21, 100844. [Google Scholar] [CrossRef]
- Magni, P.; Gravina, M.F. Macrobenthos of lagoon ecosystems: A comparison in vegetated and bare sediments. Adv. Oceanogr. Limnol. 2023, 14. [Google Scholar] [CrossRef]
- Pérez-Ruzafa, A.; Marcos, C.; Pérez-Ruzafa, I.M.; Pérez-Marcos, M. Coastal lagoons: “transitional ecosystems” between transitional and coastal waters. J. Coast. Conserv. 2011, 15, 369–392. [Google Scholar] [CrossRef]
- Colombo, G. Lagoons. In The Coastline; Barnes, R.S.K., Ed.; Wiley: London, UK; New York, NY, USA, 1977; pp. 63–81. [Google Scholar]
- Barnes, R.S.K. Coastal Lagoons; University Press Cambridge: Cambridge, UK, 1980; pp. 1–106. [Google Scholar]
- Sacchi, C. Les formes biologiques animals comme indicatrices du milieu: Considerations générales et exemples pratique. Rend. Sem. Fac. Sci. Univ. Cagliari 1984, 54, 303–318. [Google Scholar]
- Cognetti, G.; Maltagliati, F. Perspectives on the ecological assessment of transitional waters. Mar. Pollut. Bull. 2008, 56, 607–608. [Google Scholar] [CrossRef]
- Magni, P.; Tagliapietra, D.; Lardicci, C.; Balthis, L.; Castelli, A.; Como, S.; Frangipane, G.; Giordani, G.; Hyland, J.; Maltagliati, F.; et al. Animal-sediment relationships: Evaluating the “Pearson–Rosenberg paradigm” in Mediterranean coastal lagoons. Mar. Pollut. Bull. 2009, 58, 478–486. [Google Scholar] [CrossRef] [Green Version]
- Magni, P.; Como, S.; Kamijo, A.; Montani, S. Effects of Zostera marina on the patterns of spatial distribution of sediments and macrozoobenthos in the boreal lagoon of Furen (Hokkaido, Japan). Mar. Environ. Res. 2017, 131, 90–102. [Google Scholar] [CrossRef]
- Tagliapietra, D.; Pessa, G.; Cornello, M.; Zitelli, A.; Magni, P. Temporal distribution of intertidal macrozoobenthic assemblages in a Nanozostera noltii dominated area (Lagoon of Venice). Mar. Environ. Res. 2016, 114, 31–39. [Google Scholar] [CrossRef]
- Gravina, M.F.; Cabiddu, S.; Como, S.; Floris, A.; Padedda, B.M.; Pusceddu, A.; Magni, P. Disentangling heterogeneity and commonalities in nanotidal Mediterranean lagoons through environmental features and macrozoobenthic assemblages. Estuar. Coast. Shelf. Sci. 2020, 237, 106688. [Google Scholar] [CrossRef]
- Brundu, G.; Magni, P. Context-dependent effect of serpulid reefs on the variability of soft-bottom macrobenthic assemblages in three Mediterranean lagoons (Sardinia, Italy). Estuarine. Coast. Shelf Sci. 2021, 262, 107589. [Google Scholar] [CrossRef]
- De Falco, G.; Magni, P.; Teräsvuori, L.M.H.; Matteucci, G. Sediment grain size and organic carbon distribution in the Cabras lagoon (Sardinia, western Mediterranean). Chem. Ecol. 2004, 20, 367–377. [Google Scholar] [CrossRef]
- Como, S.; Magni, P.; Casu, D.; Floris, A.; Giordani, G.; Natale, S.; Fenzi, G.A.; Signa, G.; De Falco, G. Sediment characteristics and macrofauna distribution along a human-modified inlet in the Gulf of Oristano (Sardinia, Italy). Mar. Pollut. Bull. 2007, 54, 733–744. [Google Scholar] [CrossRef]
- Mitsch, W.J.; Gosselink, J.G. The value of wetlands: Importance of scale and landscape setting. Ecol. Econ. 2000, 35, 25–33. [Google Scholar] [CrossRef]
- Levin, L.A.; Boesch, D.F.; Covich, A.; Dahm, C.; Erseus, C.; Ewel, K.C.; Kneib, R.T.; Moldenke, A.; Palmer, M.A.; Snelgrove, P.; et al. The function of marine critical transition zones and the importance of sediment biodiversity. Ecosystems 2001, 4, 430–451. [Google Scholar] [CrossRef]
- Pérez-Ruzafa, A.; Marcos, C.; Pérez-Ruzafa, I.M. Mediterranean coastal lagoons in an ecosystem and aquatic resources management context. Phys. Chem. Earth Parts A/B/C 2011, 36, 160–166. [Google Scholar] [CrossRef]
- Obiefuna, J.N.; Nwilo, P.C.; Atagbaza, A.O.; Okolie, C.J. Land cover dynamics associated with the spatial changes in the wetlands of Lagos/Lekki Lagoon system of Lagos, Nigeria. J. Coast. Res. 2013, 29, 671–679. [Google Scholar]
- Newton, A.; Icely, J.; Cristina, S.; Brito, A.; Cardoso, A.C.; Colijn, F.; Dalla Riva, S.; Gertz, F.; Hansen, J.W.; Holmer, M. An overview of ecological status, vulnerability and future perspectives of European large shallow, semi-enclosed coastal systems, lagoons and transitional waters. Estuar. Coast. Shelf Sci. 2014, 140, 95–122. [Google Scholar] [CrossRef]
- Crooks, S.; Turner, R.K. Integrated coastal management: Sustaining estuarine natural resources. Adv. Ecol. Res. 1999, 29, 241–289. [Google Scholar]
- Magni, P.; Micheletti, S.; Casu, D.; Floris, A.; Giordani, G.; Petrov, A.N.; De Falco, G.; Castelli, A. Relationship between chemical characteristics of sedimentsband macrofaunal communities in the Cabras lagoon (Western Mediterranean, Italy). Hydrobiologia 2005, 550, 105.e119. [Google Scholar] [CrossRef]
- Panigrahi, S.; Acharya, B.C.; Panigrahy, R.C.; Nayak, B.K.; Banarjee, K.; Sarkar, S.K. Anthropogenic impact on water quality of Chilika lagoon RAMSAR site: A statistical approach. Wetl. Ecol. Manag. 2007, 15, 113–126. [Google Scholar] [CrossRef]
- Kolios, S.; Stylios, C.D. Identification of land cover/land use changes in the greater area of the Preveza peninsula in Greece using Landsat satellite data. Appl. Geogr. 2013, 40, 150–160. [Google Scholar] [CrossRef]
- Khedhri, I.; Afli, A.; Aleya, L. Structuring factors of the spatio-temporal variability of macrozoobenthos assemblages in a southern Mediterranean lagoon: How useful for bioindication is a multi-biotic indices approach? Mar. Pollut. Bull. 2017, 114, 515–527. [Google Scholar] [CrossRef]
- Semprucci, F.; Gravina, M.F.; Magni, P. Meiofaunal Dynamics and Heterogeneity along Salinity and Trophic Gradients in a Mediterranean Transitional System. Water 2019, 11, 1488. [Google Scholar] [CrossRef] [Green Version]
- Gravina, M.F.; Bonifazi, A.; Del Pasqua, M.; Giampaoletti, J.; Lezzi, M.; Ventura, D.; Giangrande, A. Perception of changes in marine benthic habitats: The relevance of taxonomic and ecological memory. Diversity 2020, 12, 480. [Google Scholar] [CrossRef]
- Boero, F. Episodic events: Their relevance to ecology and evolution. Mar. Ecol. 1996, 17, 237–250. [Google Scholar] [CrossRef]
- Bianchi, C.N.; Boero, F.; Fonda Umani, S.; Morri, C.; Vacchi, M. Successione e cambiamento negli ecosistemi marini. Biol. Mar. Medit. 1998, 5, 117–135. [Google Scholar]
- Muxika, I.; Borja, A.; Bald, J. Using historical data, expert judgement and multivariate analysis in assessing reference conditions and benthic ecological status, according to the European Water Framework Directive. Mar. Pollut. Bull. 2007, 55, 16–29. [Google Scholar] [CrossRef] [PubMed]
- Pinnegar, J.K.; Engelhard, G.H. The “shifting baseline” phenomenon: A global perspective. Rev. Fish Biol. 2008, 18, 1–16. [Google Scholar] [CrossRef]
- Papworth, S.K.; Rist, J.; Coad, L.; Milner-Gulland, E.J. Evidence for shifting baseline syndrome in conservation. Conserv. Lett. 2009, 2, 93–100. [Google Scholar] [CrossRef]
- Balaguer, L.; Escudero, A.; Martín-Duque, J.F.; Mola, I.; Aronson, J. The historical reference in restoration ecology: Re-defining a cornerstone concept. Biol. Conserv. 2014, 176, 12–20. [Google Scholar] [CrossRef]
- Soga, M.; Gaston, K.J. Shifting baseline syndrome: Causes, consequences, and implications. Front. Ecol. Environ. 2018, 16, 222–230. [Google Scholar] [CrossRef] [Green Version]
- Magni, P. Biological benthic tools as indicators of coastal marine ecosystems health. Chem. Ecol. 2003, 19, 363–372. [Google Scholar] [CrossRef]
- Nonnis Marzano, C.; Scalera Liaci, L.; Fianchini, A.; Gravina, M.F.; Mercurio, M.; Corriero, G. Distribution, persistence and change in the macrobenthos of the lagoon of Lesina (Apulia, southern Adriatic Sea). Oceanol. Acta 2003, 26, 57–66. [Google Scholar] [CrossRef] [Green Version]
- Basset, A.; Sangiorgio, F.; Sabetta, L. (Curatori) Nuovi approcci metodologici per la classificazione dello stato di qualità degli ecosistemi acquatici di transizione—Metodologie per la determinazione della struttura dimensionale di fitoplancton e macroinvertebrati bentonici. In Manuali e Linee Guida ISPRA; ISPRA: Rome, Italy; Università del Salento: Lecce, Italy, 2009. [Google Scholar]
- Giangrande, A.; Gravina, M.F. Brackish-water polychaetes, good descriptors of environmental changes in space and time. Transit. Waters Bull. 2015, 9, 42–55. [Google Scholar]
- Salati, F.; Meloni, A.; Marongiu, E.; Kusuda, R. Microbiological study on the flora of mussel, Mytilus galloprovincialis, cultured in Tortoli Lagoon, South-East Sardinia, Italy. Fish. Sci. 1999, 65, 657–658. [Google Scholar] [CrossRef] [Green Version]
- Parson, T.; Maita, T.; Lalli, M. (Eds.) A Manual of Chemical and Biological Methods for Seawater Analysis; Pergamon Press: Oxford, UK, 1984; p. 173. [Google Scholar]
- Clarke, K.R.; Warwick, R.M. (Eds.) Changes in Marine Communities: An Approach to Statistical Analysis and Interpretation; Primer-E Ltd.: Plymouth, UK, 2001; p. 172. [Google Scholar]
- Clarke, K.R. Non-parametric multivariate analyses of change of community structure. Austral Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Tagliapietra, D.; Sigovini, M.; Magni, P. Saprobity: A unified view of benthic succession models for coastal lagoons. Hydrobiologia 2012, 686, 15–28. [Google Scholar] [CrossRef]
- Basset, A.; Sabetta, L.; Galuppo, N. Environmental heterogeneity and benthic macroinvertebrate guilds in Italian lagoons. Transit. Waters Bull. 2007, 1, 48–63. [Google Scholar]
- Basset, A.; Pinna, M.; Sabetta, L.; Barbone, E.; Galuppo, N. Hierarchical scaling of biodiversity in lagoon ecosystems. Transit. Waters Bull. 2008, 3, 75–86. [Google Scholar]
- Magni, P.; Draredja, B.; Melouah, K.; Como, S. Patterns of seasonal variation in lagoonal macrozoobenthic assemblages (Mellah lagoon, Algeria). Mar. Environ. Res. 2015, 109, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Cognetti Varriale, A.M. Su due policheti Owenidae di sabbie infralitorali del Golfo di Follonica. Atti Soc. Tosc. Sci. Nat. Mem. Ser. B. 1979, 86, 263–267. [Google Scholar]
- Bhaud, M.R. Identification of adults and larvae in Spiochaetopterus (Polychaeta, Chaetopteridae): Consequences for larval transport and recruitment. Hydrobiologia 2003, 496, 279–287. [Google Scholar] [CrossRef]
- Magni, P.; Como, S.; Cucco, A.; De Falco, G.; Domenici, P.; Ghezzo, M.; Lefrançois, C.; Simeone, S.; Perilli, A. A multidisciplinary and ecosystemic approach in the Oristano Lagoon-Gulf system (Sardinia, Italy) as a tool in management plans. Transit. Waters Bull. 2008, 2, 41–62. [Google Scholar]
- Como, S.; Floris, A.; Pais, A.; Rumolo, P.; Saba, S.; Sprovieri, M.; Magni, P. Assessing the impact of the Asian mussel Arcuatula senhousia in the recently invaded Oristano Lagoon-Gulf system (W Sardinia, Italy). Estuar. Coast. Shelf Sci. 2018, 201, 123–131. [Google Scholar] [CrossRef]
- Cardone, F.; Corriero, G.; Fianchini, A.; Gravina, M.F.; Nonnis Marzano, C. Biodiversity of transitional waters: Species composition and comparative analysis of hard bottom communities from the south-eastern Italian coast. J. Mar. Biol. Assoc. U. K. 2014, 94, 25–34. [Google Scholar] [CrossRef]
- Rossi, R.; Cannas, A. Gli stagni sardi dalla pesca tradizionale all’uso delle nuove tecnologie. Oebalia 1992, 17, 1–10. [Google Scholar]
- Cannas, A.; Trisolini, R.; Rossi, R. La pesca nello stagno di Tortolì (Sardegna). Oebalia 1992, 17, 127–128. [Google Scholar]
Figure 1.
Study area (Tortolì Lagoon, east coast of Sardinia, Italy) and sampling sites (To1, To2, To3, To4, To5, To6).
Figure 1.
Study area (Tortolì Lagoon, east coast of Sardinia, Italy) and sampling sites (To1, To2, To3, To4, To5, To6).

Figure 2.
Line graph of water column variables related to the six sampling sites for each month: salinity, temperature, dissolved oxygen (DO).
Figure 2.
Line graph of water column variables related to the six sampling sites for each month: salinity, temperature, dissolved oxygen (DO).

Figure 3.
Box plot of water column nutrients related to the six sampling sites: nitrates (NO3), chlorophyll-a (Chl-a), and total phosphorous (PTOT); maximum value, minimum value, and median line.
Figure 3.
Box plot of water column nutrients related to the six sampling sites: nitrates (NO3), chlorophyll-a (Chl-a), and total phosphorous (PTOT); maximum value, minimum value, and median line.

Figure 4.
Principal component analysis (PCA) for environmental variables; cumulative variance (95.8%), PC1 (76.3%), PC2 (15.6%); 1, 2, 3, 4, 5, 6: number of sampling sites.
Figure 4.
Principal component analysis (PCA) for environmental variables; cumulative variance (95.8%), PC1 (76.3%), PC2 (15.6%); 1, 2, 3, 4, 5, 6: number of sampling sites.

Figure 5.
Mean values for the whole period of study (n = 8, ±SE standard error) of macrofaunal community synthetic variables at the sampling sites (To1, To2, To3, To4, To5, and To6); S: number of species; A: number of individuals (ind 0.1 m−2); H′: Shannon index; J: Evenness.
Figure 5.
Mean values for the whole period of study (n = 8, ±SE standard error) of macrofaunal community synthetic variables at the sampling sites (To1, To2, To3, To4, To5, and To6); S: number of species; A: number of individuals (ind 0.1 m−2); H′: Shannon index; J: Evenness.

Figure 6.
nMDS on average values of abundance data for the whole period of study (stress = 0.04) based on Bray–Curtis (dis)similarity.
Figure 6.
nMDS on average values of abundance data for the whole period of study (stress = 0.04) based on Bray–Curtis (dis)similarity.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Results of two-way PERMANOVA testing differences among sites for the total macrofaunal abundance, number of taxa, Shannon diversity, and evenness.
Table 1.
Results of two-way PERMANOVA testing differences among sites for the total macrofaunal abundance, number of taxa, Shannon diversity, and evenness.
| Variable | Factors | Total Sum of Squares | df | Mean Square | Pseudo-F | p |
|---|---|---|---|---|---|---|
| Species | Sites | 1.31 | 5 | 0.26 | 8.96 | 0.0001 |
| Months | 0.49 | 3 | 0.16 | 5.66 | 0.0006 | |
| Sites × Months | 1.9 | 15 | 0.13 | 4.34 | 0.0001 | |
| Residual | 0.7 | 24 | 0.03 | |||
| Total | 4.4 | 47 | ||||
| Abundance | Sites | 0.78 | 5 | 0.16 | 3.86 | 0.0008 |
| Months | 3.45 | 3 | 1.15 | 28.49 | 0.0001 | |
| Sites × Months | 4.29 | 15 | 0.29 | 7.07 | 0.0001 | |
| Residual | 0.97 | 24 | 0.04 | |||
| Total | 9.49 | 47 | ||||
| Diversity | Sites | 1.1 | 5 | 0.22 | 3.57 | 0.0011 |
| Months | 0.42 | 3 | 0.14 | 2.27 | 0.0249 | |
| Sites × Months | 1.83 | 15 | 0.12 | 1.98 | 0.02 | |
| Residual | 1.48 | 24 | 0.06 | |||
| Total | 4.83 | 47 | ||||
| Evenness | Sites | 0.38 | 5 | 0.08 | 7.28 | 0.001 |
| Months | 0.53 | 3 | 0.18 | 17.15 | 0.0001 | |
| Sites × Months | 0.75 | 15 | 0.05 | 4.81 | 0.0024 | |
| Residual | 0.25 | 24 | 0.01 | |||
| Total | 1.9 | 47 |
Table 2.
List of the top discriminating taxa contributing to the 95% in SIMPER analysis, ordered by occurrence (species occur.); higher taxon: A (ascidian), B (bivalve), C (crustacean), Ch (chironomid), Cn (cnidarian), E (echinoderm), G (gastropod), P (polychaete), Ph (Phoronid); ecological affinity: Br (brackish), Cs (coastal sheltered), Mr (marine), Op (opportunistic); contrib%: percentage of contribution of each species.
Table 2.
List of the top discriminating taxa contributing to the 95% in SIMPER analysis, ordered by occurrence (species occur.); higher taxon: A (ascidian), B (bivalve), C (crustacean), Ch (chironomid), Cn (cnidarian), E (echinoderm), G (gastropod), P (polychaete), Ph (Phoronid); ecological affinity: Br (brackish), Cs (coastal sheltered), Mr (marine), Op (opportunistic); contrib%: percentage of contribution of each species.
| Species | Higher Taxon | Ecological Affinity | Species Occur. | Contrib. % | To1 | To2 | To3 | To4 | To5 | To6 |
|---|---|---|---|---|---|---|---|---|---|---|
| Abra segmentum (Récluz, 1843) | B | Cs | 6 | 5.19 |  | | | | | |
| Apocorophium acutum (Chevreux, 1908) | C | Mr | 6 | 4.22 | | | | | | |
| Cerastoderma glaucum (Bruguière, 1789) | B | Br | 6 | 2.63 | | | | | | |
| Heteromastus filiformis (Claparède, 1864) | P | Op | 6 | 2.21 | | | | | | |
| Capitella capitata (Fabricius, 1780) | P | Op | 5 | 7.69 | | | | | | |
| Microdeutopus gryllotalpa Costa, 1853 | C | Cs | 5 | 3.60 | | | | | | |
| Monocorophium insidiosum (Crawford, 1937) | C | Br | 5 | 13.27 | | | | | | |
| Nephtys hombergii Savigny in Lamarck, 1818 | P | Mr | 5 | 4.51 | | | | | | |
| Phoronis psammophila Cori, 1889 | Ph | Mr | 5 | 0.86 | | | | | | |
| Prionospio cirrifera Wirén, 1883 | P | Op | 5 | 0.63 | | | | | | |
| Iphinoe trispinosa (Goodsir, 1843) | C | Mr | 4 | 0.30 | | | | | ||
| Melita palmata (Montagu, 1804) | C | Cs | 4 | 0.57 | | | | | ||
| Chironomidae | Ch | Op | 3 | 4.59 | | | | |||
| Venerupis corrugata (Gmelin, 1791) | B | Mr | 3 | 1.68 | | | | |||
| Aphelochaeta marioni (Saint-Joseph, 1894) | P | Mr | 2 | 1.05 | | | ||||
| Chaetozone setosa Malmgren, 1867 | P | Mr | 2 | 0.26 | | | ||||
| Diopatra neapolitana Delle Chiaje, 1841 | P | Mr | 2 | 0.26 | | | ||||
| Elasmopus pocillimanus (Spence Bate, 1862) | C | Mr | 2 | 4.30 | | | ||||
| Ericthonius brasiliensis (Dana, 1853) | C | Mr | 2 | 0.25 | | | ||||
| Gammarus insensibilis Stock, 1966 | C | Cs | 2 | 0.39 | | | ||||
| Loripes orbiculatus Poli, 1795 | B | Cs | 2 | 0.45 | | | ||||
| Malacoceros fuliginosus (Claparède, 1868) | P | Op | 2 | 1.62 | | | ||||
| Microdeutopus algicola Della Valle, 1893 | C | Mr | 2 | 2.14 | | | ||||
| Neanthes acuminata (Ehlers, 1868) | P | Cs | 2 | 1.12 | | | ||||
| Oligochaeta | O | 2 | 0.41 | | | |||||
| Paranemonia cinerea (Contarini, 1844) | Cn | Cs | 2 | 0.46 | | | ||||
| Parvicardium exiguum (Gmelin, 1791) | B | Mr | 2 | 0.38 | | | ||||
| Polydora ciliata (Johnston, 1838) | P | Op | 2 | 8.64 | | | ||||
| Protodorvillea kefersteini (McIntosh, 1869) | P | Mr | 2 | 0.58 | | | ||||
| Styela plicata (Lesueur, 1823) | A | Cs | 2 | 1.53 | | | ||||
| Tanais dulongii (Audouin, 1826) | C | Op | 2 | 0.29 | | | ||||
| Amphibalanus amphitrite (Darwin, 1854) | C | Cs | 1 | 1.60 | | |||||
| Amphibalanus eburneus (Gould, 1841) | C | Br | 1 | 2.71 | | |||||
| Amphipholis squamata (Delle Chiaje, 1828) | E | Mr | 1 | 0.82 | | |||||
| Caecum clarkii P. P. Carpenter, 1859 | G | Mr | 1 | 0.47 | | |||||
| Hilbigneris gracilis (Ehlers, 1868) | P | Mr | 1 | 0.25 | | |||||
| Lagis koreni Malmgren, 1866 | P | Mr | 1 | 0.50 | | |||||
| Lumbrineris latreilli Audouin & Milne Edwards, 1833 | P | Cs | 1 | 0.50 | | |||||
| Myriochele heeri Malmgren, 1867 | P | Mr | 1 | 4.50 | | |||||
| Pseudolirius kroyeri (Haller, 1879) | C | Mr | 1 | 0.66 | | |||||
| Sphaerosyllis taylori Perkins, 1981 | P | Mr | 1 | 0.28 | | |||||
| Spiochaetopterus costarum (Claparède, 1869) | P | Mr | 1 | 4.75 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Giampaoletti, J.; Sbrana, A.; Magni, P.; Gravina, M.F. Macrobenthos of the Tortolì Lagoon: A Peculiar Case of High Benthic Biodiversity among Mediterranean Lagoons. Diversity 2023, 15, 783. https://doi.org/10.3390/d15060783
AMA Style
Giampaoletti J, Sbrana A, Magni P, Gravina MF. Macrobenthos of the Tortolì Lagoon: A Peculiar Case of High Benthic Biodiversity among Mediterranean Lagoons. Diversity. 2023; 15(6):783. https://doi.org/10.3390/d15060783
Chicago/Turabian StyleGiampaoletti, Jacopo, Alice Sbrana, Paolo Magni, and Maria Flavia Gravina. 2023. "Macrobenthos of the Tortolì Lagoon: A Peculiar Case of High Benthic Biodiversity among Mediterranean Lagoons" Diversity 15, no. 6: 783. https://doi.org/10.3390/d15060783
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.