

No Tail No Fail: Life Cycles of the Zoogonidae (Digenea)
by
 , ,
, ,
Georgii Kremnev
1,*,†,
Anna Gonchar
1,2,
Alexandra Uryadova
1,
Vladimir Krapivin
1,
Olga Skobkina
1,
Arseniy Gubler
1 and
Darya Krupenko
1,† 1
Department of Invertebrate Zoology, Saint Petersburg University, 199034 Saint Petersburg, Russia
2
Laboratory of Parasitic Worms and Protists, Zoological Institute, Russian Academy of Sciences, 199034 Saint Petersburg, Russia
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Diversity 2023, 15(1), 121; https://doi.org/10.3390/d15010121
Submission received: 18 November 2022
/
Revised: 4 January 2023
/
Accepted: 8 January 2023
/
Published: 16 January 2023
(This article belongs to the Special Issue Diversity of Macroparasites in Marine Fishes)
Abstract
:The Zoogonidae is the only digenean family where known cercariae lack the tail but actively search for the second intermediate host. However, the data on the zoogonid life cycles are scarce. In the present study, we elucidated and verified life cycles of the Zoogonidae from the White Sea. Using rDNA data, we showed that Pseudozoogonoides subaequiporus utilizes gastropods from the family Buccinidae as the first intermediate host and protobranch bivalves as the second one. This life cycle can be facultatively truncated: some cercariae of P. subaequiporus encyst within the daughter sporocysts. Molecular data also confirmed previous hypotheses on Zoogonoides viviapus life cycle with buccinid gastropods acting as the first intermediate hosts, and annelids and bivalves as the second intermediate hosts. We demonstrated the presence of short tail primordium in the developing cercariae of both species. Based on the reviewed and our own data, we hypothesize that the emergence of tailless cercariae in the evolution of the Zoogonidae is linked to the switch to non-arthropod second intermediate hosts, and that it possibly happened only in the subfamily Zoogoninae. Basally branching zoogonids have retained the ancestral second intermediate host and might have also retained the tail.
1. Introduction
Life cycles of the Digenea are complex, typically with two or three hosts, alteration of reproductive modes and two free-dwelling larval stages, miracidium and cercariae. Locomotion of miracidia is provided by ciliated epithelial cells. Cercariae lack cilia and move using their tail—a specialized muscular hind part of the body. The tailed cercaria is most likely a plesiomorphic condition for the Digenea [1,2,3].
Cercarial tails are diverse in structure and function, depending on the specific transmission pattern. The tail gets modified to facilitate swimming (in the Heterophyidae and Lepocreadiidae [4,5,6]) and floating (in the Diplostomidae and Cyathocotylidae [7,8]), to attract the next host (in the Bivesiculidae and Azygiidae [9]) and even to penetrate the second intermediate host (in the Hemiuroidea [10,11]). Alternatively, in a number of the digenean taxa there is a clear evolutionary trend towards tail reduction. In the Opecoelidae and Lepidapedidae, tails are very short and used for attachment, not locomotion [12,13]. Tails are also reduced in the cercariae of the digeneans with secondary dixenous life cycles. Those cercariae do not leave the intermediate host and transform into infective metacercariae therein [14,15,16].
A special case is the family Zoogonidae, parasites of marine and freshwater fish [17]. Known zoogonid cercariae lack the tail, but they leave the first intermediate host and actively crawl over the substrate in a leech-like manner to find and infect the second intermediate host which is typically a slow-moving benthic invertebrate [18,19]. However, our general knowledge on the zoogonid life cycles stays rather poor since they are completely reconstructed only for five species [19,20,21,22,23]. The question is: how do the Zoogonidae, having such a bizarre cercariae strategy, thrive as a cosmopolitan parasite group, abundant in various communities, from the intertidal to the abyssal zone [24,25]?
The main goal of this study was to elucidate and verify the life cycles of the Zoogonidae from the White Sea: Pseudozoogonoides subaequiporus (Odhner, 1911) and Zoogonoides viviparus (Olsson, 1868). The second objective was to study the development of the zoogonid cercariae to check upon the tail primordium. Finally, we aimed to review published data on the Zoogonidae life cycles and speculate on the possible premises of the tailless cercariae emergence.
2. Materials and Methods
2.1. Sampling
Sampling was conducted at the Keret Archipelago, White Sea, in 2018–2021. We examined four fish species to find maritae of the Zoogonidae: Atlantic wolffish Anarhichas lupus Linnaeus, 1758, common dab Limanda limanda (Linnaeus, 1758), Arctic flounder Liopsetta glacialis (Pallas, 1776) and European flounder Platichthys flesus (Linnaeus, 1758). To detect sporocysts, cercariae and metacercariae of zoogonids we dissected a variety of subtidal invertebrates listed in Table 1.
2.2. Morphological Analysis
For the whole mounts, zoogonid maritae and metacercariae extracted from the cyst were heat-killed and then fixed in 96% ethanol. We used a pair of insect pins to rupture the cyst wall and extract metacercariae. Sporocysts and cercariae were fixed in Shaudin’s solution, or mercury (II) chloride with acetic acid, or Zenker’s solution for whole mounts and histological study. To identify mucoid substances, we used metachromatic staining with toluidine blue. Acetic carmine staining was used to visualize general structure. Histological sections were stained with Azur II-eosin. All procedures were the same as previously described [26]. Whole mounts were observed using a compound microscope Leica DM 2500 (Leica Microsystems, Wetzlar, Germany), and photos were taken with a Nikon DS Fi3 camera (Nikon, Tokyo, Japan). Measurements were conducted in Fiji software [27]. All measurements are in micrometers, the range of values is followed by the mean in parentheses. Visualization of flame cells arrangement, digestive and muscular systems in cercariae was performed by means of confocal laser scanning microscopy (CLSM). Specimens were fixed in 4% paraformaldehyde in 0.01 M phosphate buffered saline (PBS) for 6–24 h, then washed in PBS and stored in PBS with 0.1% sodium azide (NaN3) at 4 °C. Specimens were stained with TRITC-labeled phalloidin (Sigma Aldrich, Darmstadt, Germany, P1951) in PBS (500 ng/mL), incubated for 60 min and washed in PBS for 20 min. Finally, specimens were mounted in Fluoroshield™ medium with DAPI (Sigma Aldrich, Darmstadt, Germany, F6057) and examined under the CLSM Leica TCS SP5 (Leica Microsystems, Wetzlar, Germany).
2.3. Molecular Analysis
For molecular analysis, we utilized ethanol preserved samples. Eighteen isolates of putative and identified zoogonids were used (Table 2). To extract DNA, samples were taken from 96% ethanol and dried completely, incubated in 200 μL of 5% Chelex® 100 resin (Bio-Rad, Hercules, California, USA) solution with 0.2 mg/mL of proteinase K at 56 °C for 4 h, then kept for 8 min at 90 °C and centrifuged at 16,000× g for 10 min. The supernatant containing DNA was then transferred to a new tube and stored at −20 °C. Several primers were used to amplify fragments of the ribosomal operon (Table 3). Amplifications were performed in 20 μL reaction mixture containing 4 μL of ScreenMix-HS (Evrogen, Moscow, Russia), 0.5 μL of forward and reverse primers (10 pmol/µL), 2 μL of the DNA and 13 μL of PCR grade water (Evrogen, Moscow, Russia). PCR products were stained with ethidium bromide and visualized through electrophoresis on a 1% agarose gel. Sequencing was carried out with PCR primers on an ABI Prism 3500xl genetic analyzer (Applied Biosystems, Waltham, Massachusetts, USA). To analyze and edit chromatograms and to build alignments, we used Geneious Prime 2022.0.1 (https://www.geneious.com (accessed on 10 November 2022)). We used three datasets: the first one to link the life cycle stages (28S and 5.8S+ITS2 rDNA), the second and the third ones to resolve the phylogenetic position of Pseudozoogonoides subaequiporus (28S rDNA). All the information on GenBank sequences used in the second and the third datasets is listed in Supplementary Table S1. The maximum likelihood (ML) analysis was conducted for all datasets at the CIPRES Science Gateway (https://www.phylo.org (accessed on 10 November 2022)) using RAxML [28] with 1000 bootstrap iterations. The stability of clades was assessed using non-parametric bootstrapping with 1000 pseudoreplicates. The Bayesian analysis was conducted for the second and the third datasets using MrBayes v.3.2.6 [29] with 1,000,000 generations. The model of nucleotide substitution was estimated as GTR+G (28S) and K2+G (5.8S+ITS2) for the first dataset (Figure 1), as GTR+G+I for the second dataset (Supplementary Figure S1) and as GTR+G for the third dataset (Figure 2), using JModelTest [30] in MEGA 11 [31] under the maximum likelihood model.
2.4. Visualization
We used CorelDraw® 2017 software to create the schemes of parasite life cycle stages as well as intermediate and definitive hosts.
3. Results
3.1. General Observations
In the studied White Sea fish, we found zoogonid maritae of three species. Zoogonoides viviparus was obtained from the three examined flatfish species and from Atlantic wolffish. Pseudozoogonoides subaequiporus and Lepidophyllum steenstrupi Odhner, 1902 were found only in Atlantic wolffish. Sporocyst and cercariae similar to the ones described by Lebour [36] and Køie [19] as Z. viviparus were obtained from the common whelk Buccinum undatum Linnaeus, 1758. Zoogonid sporocysts with morphologically different cercariae and also encysted metacercariae were found in B. undatum and Neptunea despecta (Linnaeus, 1758). These larvae resembled maritae of P. subaequiporus. In one more whelk species, B. scalariforme Møller, 119, we found zoogonid sporocysts at the earlier stage of development, containing only the germ balls. Other dissected gastropods were free from zoogonid sporocysts (Table 1). Metacercariae similar to Z. viviparus maritae were found in several bivalve species and in the annelid Nephtys sp. We also recovered another type of zoogonid metacercariae, morphologically similar to maritae of P. subaequiporus, from protobranch bivalve Nuculana pernula (O. F. Müller, 1779). All the other examined benthic invertebrates were not infected with zoogonid metacercariae (Table 1).
3.2. Molecular Data and Life Cycles Elucidation
To link life cycle stages from intermediate hosts and maritae from definitive hosts we obtained 1244–1263 b.p. sequences of the 28S rDNA fragment (domains D1–D3) for all 18 isolates (Table 2). We also included 28S rDNA sequences of Z. viviparus (AY222271) and L. steenstrupi (AY157175) maritae into the analysis. Steganoderma cf. eamiqtrema Blend and Racz, 2020 (MW264135) was added as an outgroup. After trimming to match the shortest sequence, the alignment was 1269 b.p. long, including gaps. Three distinct groups were evident in the maximum likelihood (ML) tree (Figure 1). The first one comprised Z. viviparus from GenBank (AY222271), newly obtained putative and identified isolates of Z. viviparus and the zoogonid sporocyst from B. scalariforme (Figure 1). The second group incorporated putative and identified isolates of P. subaequiporus (Figure 1). Both groups had no intraspecific variability in 28S fragments and differed from each other in 86 positions. The third group comprised sequences of L. steenstrupi, the one from GenBank (AY157175) and the one newly obtained from the White Sea isolate (Figure 1). These sequences differed by a single nucleotide substitution (C/A) in the position 293.
For all the 18 isolates, we also sequenced a 431–545 b.p. long fragment containing partial 5.8S, ITS2 and the beginning of the 28S rDNA (Table 2). After trimming to match the shortest sequence, the alignment was 456 b.p. long, including gaps. Sequence of S. cf. eamiqtrema (MW264129) was added as an outgroup. In the ML tree, our sequences are grouped the same way as in the 28S rDNA analysis (Figure 1). The clades including Z. viviparus and P. subaequiporus isolates had no intraspecific variability and differed from each other in 97 positions. Sequence of L. steenstrupi differed from these groups in 133 and 91 positions, respectively (Figure 1).
We also generated a 1378 b.p. long sequence of 18S rDNA fragment for one isolate of P. subaequiporus marita and deposited it in the GenBank under accession number OP956057.
3.3. Phylogenetic Position of Pseudozoogonoides subaequiporus
As we obtained the first molecular data for P. subaequiporus, they were used to resolve its phylogenetic position. Analysis was run with two datasets: the first one comprised 51 sequences and the second one just 26 sequences. The resulting phylogenetic trees showed the same relationship between families and within the Zoogonidae. The tree based on the larger dataset is shown in Supplementary Figure S1. Below we describe results of the analysis inferred from the smaller dataset.
The alignment comprised 26 sequences of 28S rDNA fragments and yielded 1231 characters, including gaps. Phylogenetic trees inferred with ML and Bayesian approaches revealed almost the same topology. The Bayesian tree is presented in Figure 2; ML bootstrap support values are additionally mapped onto it next to the posterior probability (PP) values for all the ML-supported nodes.
Similarly to previous studies [37,38,39,40], the Zoogonidae was resolved within the Microphalloidea as a well-supported clade, though containing some members of the family Faustulidae (Figure 2). The latter is known to be polyphyletic as several faustulid species fall into the superfamily Gymnophalloidea [41,42,43], thus we place this family name into the quotation marks. The clade Renicolidae + Eucotylidae was inferred as a sister group to the Zoogonidae with a low BI and ML support. Psedudozoogonoides subaequiporus grouped together with Z. viviparus in a well-supported clade within the Zoogoninae (Figure 2). Monophyly of the latter was poorly supported. The former Lepidophyllinae was resolved as highly paraphyletic, and the Zoogoninae was inferred as the closest relative of the ”faustulids”, similarly to previous studies [37,38,39,40,41,42,43].
3.4. Infection Data and Morphological Descriptions
3.4.1. Maritae of Zoogonoides viviparus
Locality: White Sea, Keret Archipelago.
Hosts and infection rates: Anarhichas lupus (prevalence 25%, N = 28; mean intensity = 5.7); Limanda limanda (91%, N = 116; MI = 21.1); Liopsetta glacialis (37%, N = 27; MI = 7.6); Platichthys flesus (52%, N = 79; MI = 19.3).
Other reported hosts: 20 fish species (summarized in [44]).
Sites: Intestine, occasionally urinary bladder
Vouchers: Paragenophores (four slides) deposited in the collection of the Department of Invertebrate Zoology of Saint Petersburg University.
Description: Measurements in Table 4.
Remarks: Ovigerous specimens located predominantly in rectum.
3.4.2. Metacercariae of Zoogonoides viviparus
Locality: White Sea, Keret Archipelago.
Hosts and infection rates: Bivalvia: Nuculana pernula (2.1%, N = 394; MI = 1.3); Yoldia hyperborea (Gould, 1841) (1.3%, N = 78; MI = 1); Serripes groenlandicus (Mohr, 1786) (10.6%, N = 66; MI = 1.1); Ciliatocardium ciliatum (Fabricius, 1780) (5.1%, N = 39, MI = 1.5). Annelida: Nephtys sp. (25%, N = 8; MI = 2.5).
Other reported hosts: Various benthic invertebrates (summarized in [44]).
Sites: Bivalvia: Mantle, siphons, gills, oral lobes, visceral mass, adductor muscles, digestive gland. Annelida: Somatic musculature, gut wall.
Vouchers: Syngenophores (seven slides) deposited in the collection of the Department of Invertebrate Zoology of Saint Petersburg University.
Description: [Measurements based on 7 fixed specimens from bivalves. See also Table 4].
Body of metacercaria extracted from cyst 385 (341–446) × 115 (101–137), oval, with narrowed anterior and posterior ends. Tegument spinose. Stylet present (five of six metacercariae). Oral sucker subterminal, oval, 68 (59–81) × 73 (60–99). Ventral sucker oval, 99 (82–114) × 94 (82–132). Sucker ratio 1:1.47 (1.33–1.68). Prepharynx very short, pharynx oval, 23 (18–36) × 30 (24–34), esophagus long. Ceca reaching posterior third of ventral sucker; lumen present. Excretory vesicle in hindbody, small, oval. Two testes primordia behind ventral sucker, 39 (23–57) × 31 (18–47) and 37 (25–57) × 31 (21–44). Cirrus-sac primordium 95 (81–115) × 23 (19–27). Ovary primordium 31 (21–41) × 28 (18–39), between testes, dorsally. Other primordia of female reproductive system (oviduct, Laurer’s canal, vitellarium, uterus and metraterm) present.
3.4.3. Sporocysts, Developing and Infective Cercariae of Zoogonoides viviparus (Figure 3A–C and Figure 4)
Locality: White Sea, Keret Archipelago.
Hosts and infection rates: Buccinum undatum (0.9%, N = 2174); B. scalariforme (5.2%, N = 19).
Sites: Digestive and reproductive glands. In heavily infected whelks, daughter sporocysts are also located in the kidney and body wall near esophagus and stomach. Infective cercariae commonly accumulate in the kidney.
Vouchers: Isogenophores (two slides) deposited in the collection of the Department of Invertebrate Zoology of Saint Petersburg University.
Description:
Daughter sporocysts (Figure 4A)
[Measurements based on 10 fixed specimens from B. undatum].
Sporocysts oval, 904 (476–1178) × 343 (252–443). Brood cavity occupying almost all inner space, containing germ balls, developing and infective cercariae. Birth pore terminal at anterior body end. Germinal mass subterminal at posterior body end, embedded in parenchyma.
[Measurements based on 11 fixed specimens. See also Table 4].
Tailless distome xiphidiocercariae. Body 388 (337–451) × 139 (125–158), oval, with narrowed anterior and posterior ends. Tegument spinose. Oral sucker subterminal, oval, 73 (63–79) × 73 (65–83). Ventral sucker oval, 103 (93–119) × 87 (71–98), closer to posterior end. Sucker ratio 1:1.40 (1.19–1.63). Prepharynx very short, pharynx oval, 23 (17–36) × 29 (26–32), esophagus long, 61 (46–76). Ceca reaching posterior third of ventral sucker; lumen present. Cerebral ganglion dorsal to prepharynx. Stylet in pocket within oral sucker, dorsally to mouth opening, 16 (12–20) × 5 (4–6), single-pointed, two-layered; external refractive layer with two small shoulders; posterior end of internal layer naked. One group of 17 large unicellular penetration glands in forebody. Five glands without ducts. Twelve ducts of penetration glands arranged according to formula (2 + 3 + 1) + (1 + 3 + 2). Each duct with individual pore near anterior edge of oral sucker. Excretory vesicle small, oval. Excretory system of ‘‘Mesostoma” type. Main collecting ducts dividing into anterior and posterior parts at posterior third of ventral sucker. Excretory formula 2[(2 + 2) + (2 + 2)] = 16. Two testes primordia 48 (42–57) × 36 (33–40) and 45 (40–50) × 33 (27–37), in hindbody, symmetrical, anterior testis slightly larger. Cirrus-sac primordium anterior to ventral sucker. Vasa efferentia fusing at cirrus-sac primordium. Ovary primordium between testes, dorsally, 28 (24–30) × 23 (20–27). Vitellarium primordium smaller, ventral to ovary. Forming Laurer’s canal posterior to ovary. Uterus primordium running anteriorly, its dilated distal part close to cirrus-sac primordium.
In underdeveloped cercariae, a very short knob-like tail primordium was evident. It disappears during further development; therefore, infective cercariae are tailless. In underdeveloped cercariae, mucoid substances were found in the mucoid gland cells which lie ventrally, three pairs in the forebody and two in the hindbody. During the final stages of cercarial development, the mucoid substances are transferred into the tegument which becomes swollen.
Remarks: Morphology of studied cercariae agrees with the previous descriptions of this species [19,36,46]. Remarkable features revealed in our study are the number of the penetration glands and their ducts, stylet structure, structure of the excretory and genital systems, number of the mucoid glands and presence of the tail primordium in developing cercariae.
Figure 3.
Zoogonoides viviparus (A–C) and Pseudozoogonoides subaequiporus (D–F) cercariae. (A,D) Infective cercariae, general structure, acetic carmine staining and differential interference contrast (DIC). (B,E) Infective cercariae, flame cells, digestive and excretory systems, confocal laser scanning microscopy and TRITC-phalloidin staining. (C,F) Developing cercariae, mucoid gland cells and the tail primordium, toluidine blue staining and DIC. Abbreviations: c—cecum; cg—cerebral ganglion; gp—genital system primordium; os—oral sucker; ph—pharynx; t—testis primordium; vs—ventral sucker. Arrowheads indicate flame cells (B,E) and mucoid gland cells (C,F). Arrows indicate the tail primordium. Asterisk marks magnified fragment in square of (C).
Figure 3.
Zoogonoides viviparus (A–C) and Pseudozoogonoides subaequiporus (D–F) cercariae. (A,D) Infective cercariae, general structure, acetic carmine staining and differential interference contrast (DIC). (B,E) Infective cercariae, flame cells, digestive and excretory systems, confocal laser scanning microscopy and TRITC-phalloidin staining. (C,F) Developing cercariae, mucoid gland cells and the tail primordium, toluidine blue staining and DIC. Abbreviations: c—cecum; cg—cerebral ganglion; gp—genital system primordium; os—oral sucker; ph—pharynx; t—testis primordium; vs—ventral sucker. Arrowheads indicate flame cells (B,E) and mucoid gland cells (C,F). Arrows indicate the tail primordium. Asterisk marks magnified fragment in square of (C).

3.4.4. Maritae of Pseudozoogonoides subaequiporus
Locality: White Sea, Keret Archipelago.
Host and infection rates: Anarhichas lupus (93%, N = 28; MI = 38.1).
Site: Intestine.
Vouchers: Paragenophores (three slides) deposited in the collection of the Department of Invertebrate Zoology of Saint Petersburg University.
Description: Measurements in Table 4.
Remarks: Ovigerous specimens located predominantly in rectum.
3.4.5. Metacercariae of Pseudozoogonoides subaequiporus (Figure 5)
Locality: White Sea, Keret Archipelago.
Hosts and infection rates: Neptunea despecta (1 of 103); Buccinum undatum (1 of 2174); Nuculana pernula (1%, N = 394; MI = 1).
Sites:
Bivalvia: Mantle, siphons.
Vouchers: Hologenophore (single slide) deposited in the collection of the Department of Invertebrate Zoology of Saint Petersburg University.
Description:
Metacercariae enclosed in oval cyst, 153 (123–187) × 137 (116–157) (measurements based on 22 fixed specimens from N. despecta and B. undatum). Following description based on single hologenophore from N. pernula. Body of metacercaria extracted from cyst 275 × 119, oval, with slightly narrowed posterior end. Tegument spinose. Oral sucker subterminal, oval, 53 × 59. Ventral sucker roundish, 49 × 50. Sucker ratio 1:0.92. Prepharynx very short, pharynx oval, esophagus long. Ceca ending behind ventral sucker; lumen present. Excretory vesicle in hindbody, small, oval. Two testes primordia behind ventral sucker, 40 × 30 and 40 × 31. Cirrus-sac primordium 63 × 23. Ovary primordium 23 × 23, between testes, dorsally. Other primordia of female reproductive system (oviduct, ootype, Laurer’s canal, vitellarium, uterus and metraterm) present.
Remarks: Two of four metacercariae recovered from N. pernula were dead. Zhukov [47] claimed he found metacercariae of P. subaequiporus in mesenteries of small flounders from the Sea of Japan, but since he provided neither dimensions nor figure, we are unable to compare his finding with our results.
Figure 4.
Drawings of life cycle stages of Zoogonoides viviparus: daughter sporocyst (A), infective cercaria body structure (B), stylet, ventral view (C), mucoid gland cells and the tail primordium in underdeveloped cercaria (D).
Figure 4.
Drawings of life cycle stages of Zoogonoides viviparus: daughter sporocyst (A), infective cercaria body structure (B), stylet, ventral view (C), mucoid gland cells and the tail primordium in underdeveloped cercaria (D).

3.4.6. Sporocysts, Developing and Infective Cercariae of Pseudozoogonoides subaequiporus (Figure 3D–F and Figure 6)
Locality: White Sea, Keret Archipelago.
Hosts and infection rates: Neptunea despecta (1 of 103); Buccinum undatum (1 of 2174).
Sites: Digestive and reproductive glands.
Vouchers: Isogenophores (eight slides) deposited in the collection of the Department of Invertebrate Zoology of Saint Petersburg University.
Description:
Daughter sporocysts (Figure 6A)
[Measurements based on 20 fixed specimens, 10 from N. despecta and 10 from B. undatum].
Sporocysts elongated, 955 (528–1879) × 269 (171–350). Broad brood cavity containing germ balls, developing and infective cercariae and encysted metacercariae. Birth pore terminal at anterior body end. Germinal mass subterminal at posterior body end, embedded in parenchyma.
[Measurements based on 11 fixed specimens. See also Table 4].
Tailless distome xiphidiocercariae. Body 301 (248–385) × 118 (98–136), oval, with slightly narrowed posterior end. Tegument spinose. Oral sucker subterminal, roundish, 58 (50–66) × 57 (50–66). Ventral sucker roundish, 60 (54–65) × 60 (55–66), closer to posterior end. Sucker ratio 1:1.05 (0.92–1.24). Prepharynx very short, pharynx oval, 19 (15–21) × 23 (21–25), esophagus long, 43 (26–69). Ceca ending behind ventral sucker; lumen present. Cerebral ganglion dorsal to prepharynx. Stylet in pocket within oral sucker, dorsally to mouth opening, 13 (10–16) × 4 (3–5), single-pointed, two-layered; external refractive layer with two small shoulders; posterior end of internal layer naked. One group of 12 large unicellular penetration glands in forebody. Two glands without ducts. Ten ducts of penetration glands arranged according to formula (3 + 2) + (2 + 3). Each duct with individual pore near anterior edge of oral sucker. Excretory vesicle elongated, oval. Excretory formula 2[(2 + 2) + (2 + 2)] = 16. Two testes primordia 25 (16–30) × 18 (12–23) and 25 (18–30) × 19 (14–23), in hindbody, symmetrical. Cirrus-sac primordium anterior to ventral sucker. Vasa efferentia fuse at cirrus-sac primordium. Ovary primordium between testes, dorsally, 18 (17–20) × 15 (14–17). Vitellarium primordium small, bipartite, ventral to ovary. Ootype primordium between testes, ventrally. Forming Laurer’s canal posterior to ovary. Uterus primordium running anteriorly, its dilated distal part close to cirrus-sac primordium.
In underdeveloped cercariae, a very short knob-like tail primordium was evident. It disappears during further development; therefore, infective cercariae are tailless. In underdeveloped cercariae, mucoid substances were found in mucoid gland cells which lie ventrally, three pairs in the forebody and two in the hindbody. During the final stages of cercarial development, the mucoid substances are transferred into the tegument which becomes swollen.
Remarks: Encysted metacercariae were more abundant in the infected specimen of B. undatum (collected at the end of June) than in the infected specimen of N. despecta (collected at the end of July).
Figure 5.
Histological section through infected whelk Neptunea despecta showing encysted metacercariae of Pseudozoogonoides subaequiporus within daughter sporocyst, Azur II-eosin staining. Abbreviations: ev—excretory vesicle; os—oral sucker; vs—ventral sucker. Arrows indicate cyst wall.
Figure 5.
Histological section through infected whelk Neptunea despecta showing encysted metacercariae of Pseudozoogonoides subaequiporus within daughter sporocyst, Azur II-eosin staining. Abbreviations: ev—excretory vesicle; os—oral sucker; vs—ventral sucker. Arrows indicate cyst wall.

Figure 6.
Drawings of life cycle stages of Pseudozoogonoides subaequiporus: sporocyst (A), infective cercaria body structure (B), stylet, ventral view (C), mucoid gland cells and the tail primordium in underdeveloped cercaria (D).
Figure 6.
Drawings of life cycle stages of Pseudozoogonoides subaequiporus: sporocyst (A), infective cercaria body structure (B), stylet, ventral view (C), mucoid gland cells and the tail primordium in underdeveloped cercaria (D).

4. Discussion
The first hypothesis on the life cycle of Zoogonoides viviparus was proposed by Lebour [36]. She found sporocysts and tailless distome xiphidiocercariae in a common whelk, Buccinum undatum, and suggested these were conspecific with maritae of Z. viviparus. Later, based on both natural and experimental evidence, Køie [19] showed that the first intermediate host of Z. viviparus was indeed B. undatum, and the second ones were mollusks, annelids and brittle stars. By molecular data, we confirmed that buccinid gastropods B. undatum and B. scalariforme are the first intermediate hosts, and bivalves and annelids are the second intermediate hosts of this species (Figure 7A).
Køie [19] proposed brittle stars, and particularly Ophiura albida Forbes, 1839, to be the most important second intermediate hosts of Z. viviparus in Danish waters, considering high infection and survival rates of metacercariae. Studies of Scott in Canada indicated that brittle stars could serve as the second intermediate host of Pseudozoogonoides subaequiporus as well, judging from higher infection rates with maritae of this species in bigger flatfishes, which consume more echinoderms [48,49]. However, O. albida is absent in the White Sea [50], and we did not find any zoogonid metacercariae in the abundant local brittle stars, O. robusta (Ayres, 1852) and Ophiopholis aculeata (Linnaeus, 1767). We found metacercariae of Z. viviparus only in the annelids Nephtys sp. and several bivalve species from the families Cardiidae, Nuculanidae and Yoldiidae. Such infection routes match feeding preferences of the White Sea definitive hosts of Z. viviparus: local flatfishes feed predominantly on annelids and bivalves, and Atlantic wolffish consume mainly hard-shelled invertebrates ([51,52]; our unpublished observations), including bivalves.
We elucidated the three-host life cycle of P. subaequiporus using molecular data and demonstrated that it utilizes buccinid gastropods Neptunea despecta and B. undatum as the first intermediate hosts, and bivalves as the second intermediate hosts (Figure 7B). We found metacercariae of P. subaequiporus in a single bivalve species, N. pernula. Other benthic invertebrates also should be considered as potential second intermediate hosts of this species. However, the main transmission route of P. subaequiporus to the Atlantic wolffish in the White Sea is likely through direct consumption of the infected first intermediate hosts, buccinid gastropods, containing encysted metacercariae (Figure 7B). Shulman-Albova [53], therefore, correctly predicted that P. subaequiporus has a facultatively dixenous life cycle based on high infection rates of the Atlantic wolffish.
According to the results presented here and published earlier, the range of the first intermediate hosts of the Zoogonidae comprises gastropods from the Trochoidea (Vetigastropoda), Buccinoidea (Neogastropoda) and Naticidae (Littorinimorpha) (reviewed by Barnett et al. [54]). However, we disagree with the idea that naticid gastropods harbor zoogonid sporocysts and cercariae proposed by Palombi [20]. He did not find zoogonid infection in the Naticidae himself, but hypothesized Cercariae crispata Pelseneer, 1906 from Euspira pulchella (Risso, 1826), belongs to the species Diphterostomum brusinae (Stossich, 1888). However, from Pelseneer [55] drawings it is evident that the structure of digestive system, shape of the excretory bladder, and sucker position do not match zoogonid cercariae or metacercariae. Instead, these larvae strongly resemble metacercariae of the Microphallidae. The host, naticid gastropod, and the absence of cyst makes us suspect that Cercariae crispata should be identified as Microphallus pseudopygmaeus Galaktionov, 1980. This species has a secondary dixenous life cycle and utilizes a great variety of gastropods as the first intermediate hosts [56].
Sharing the same first (common whelk) and second (bivalve N. pernula) intermediate hosts by Z. viviparus and P. subaequiporus may lead to misidentification of their cercarial and metacercarial stages. The most reliable discriminative feature is the sucker ratio: ventral sucker in Z. viviparus is relatively larger in all the ontogenetic stages. The body size can also help to tell Z. viviparus and P. subaequiporus apart: cercariae and metacercariae of Z. viviparus are larger (Table 4).
Studied material allowed us to specify two important morphological traits of the zoogonid cercariae. The first one is the presence of five pairs of mucoid glands at the ventral body side of developing cercariae (Figure 5D and Figure 6D). Such a pattern is unique among Xiphidiata since cercariae of the other Microphalloidea and Plagiorchioidea have just four pairs of the mucoid glands [57,58,59]. Previously, only Køie [46] noted the presence of mucoid glands in developing cercariae of Z. viviparus, though she did not identify their number. The second significant trait is a short knob-like tail primordium in the developing cercariae (Figure 5D and Figure 6D). This primordium disappears during further development and the infective cercariae become tailless. Previously, only Shimura and Ito [60] noted the tail primordium in immature larvae of Cercaria brachycaeca Shimura & Ito, 1980, putative Zoogonidae. The presence of tail primordium proves that tailless zoogonid cercariae evolved from an ancestor with a tail.
What are the benefits of tail loss? Apparently, it is increased longevity of cercariae as tail-based locomotion requires a lot of energy. The lifespan of actively swimming cercariae is quite short and commonly does not exceed 24 h [61]. However, in short-tailed cercariae the longevity is higher [62,63,64,65,66]. Tailless zoogonid cercariae have a lifespan between 24 and 120 h [19,46,60,67,68,69,70], and low temperature prolongs it up to 8–12 days [67].
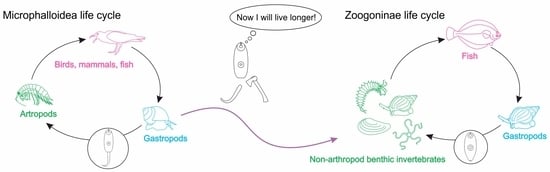
Together with the longevity benefit, tail loss obviously makes cercariae less motile, and that must affect the range of second intermediate hosts. Judging from the known life cycles, an arthropod second intermediate host is a common plesiomorphic feature for the Microphalloidea and Plagiorhioidea in general, and the Zoogonidae in particular (Figure 2). The only zoogonid group with documented tailless cercariae is the subfamily Zoogoninae, and their range of second intermediate hosts is wide and includes a variety of benthic invertebrates, such as polychaetes, gastropods, bivalves, sea lilies, brittle stars and sea urchins, but never arthropods ([19,21,22,23,24,45,67,70,71,72,73,74,75]; Figure 2). Conversely, in the basally branching Zoogonidae (Limnoderetrema minutum (Manter, 1954) and Steganoderma spp.), the second intermediate host range is restricted to arthropods: shrimps, crabs, hermit crabs and mysids ([38,76,77,78,79]; Figure 2).
Although from some studies one may conclude that all zoogonid cercariae are tailless [80], this inference seems to be premature since we have no data on the cercariae of the basally branching zoogonids. These species have the ancestral life cycle pattern, and their cercariae could have retained the tail. It may turn out arthropods are too active to be chased by tailless cercariae.
Additionally, metacercariae of the Zoogoninae and the basally branching zoogonids differ in their growth capacity. Metacercariae of Steganoderma spp. are large, up to five millimeters, and both Steganoderma spp. and L. minutum sometimes start egg production within the second intermediate host [38,76,77,78,79]. All the metacercariae of the Zoogoninae are much smaller (their length does not exceed 600 μm), and they never reach sexual maturity in the second intermediate host [19,22,23,24,45,67,71,72,73,74,75]. Thus, growth capacity seems to be the cost of the host switch to diverse non-arthropod invertebrates.
Based on the reviewed and retrieved data, we hypothesize that two major events in the evolution of the Zoogonidae, tail loss and switch to non-arthropod second intermediate host, might have co-occurred in the Zoogoninae lineage. This led to lower host specificity and growth capacity of the metacercariae in this group.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/d15010121/s1, Table S1: List of species and 28S rDNA sequences used in this study (the second and the third datasets); Figure S1: Analysis of the second dataset. Phylogenetic position of Pseudozoogonoides subaequiporus (indicated bold) based on 28S rDNA sequence data analysis inferred with Bayesian inference (BI) and maximum likelihood (ML). Nodal support: ML/BI. Scale bar shows the substitution rate. Model of nucleotide substitution: GTR+G+I. Refs. [81,82,83,84,85,86,87,88,89,90,91,92,93,94,95,96,97,98,99,100,101] are in the supplementary materials.
Author Contributions
D.K. conceived and designed the study. G.K., A.U., V.K., O.S., A.G. (Arseniy Gubler) and D.K. conducted sampling. G.K., D.K., A.G. (Anna Gonchar) and A.G. (Arseniy Gubler) performed molecular analyses. G.K., O.S. and D.K. performed morphological analyses. G.K., D.K. and A.G. (Anna Gonchar) wrote the article. D.K. received funding. All authors have read and agreed to the published version of the manuscript.
Funding
The study was financially supported by the Russian Science Foundation (Russia), grant no. 19-74-10029. Work at the White Sea Biological Station ‘‘Kartesh” of the Zoological Institute of the Russian Academy of Sciences (ZIN RAS) was supported by a research program of ZIN RAS (project no. 122031100260-0).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The fieldwork was carried out at the Educational and Research Station ‘‘Belomorskaia” of Saint Petersburg University (SPbU), Russia, and the White Sea Biological Station ‘‘Kartesh” of the Zoological Institute of the Russian Academy of Sciences (ZIN RAS), Russia. Our sincere thanks for the help with the sampling are due to Sergei Bagrov and George Slyusarev. The authors are particularly grateful to Anna Romanovich and Aleksey Masharsky for excellent sequencing support. We are also thankful to Leonie Barnett who provided us with relevant literature on the zoogonid life cycles. Finally, we thank three anonymous reviewers whose comments were a great help in improving this manuscript. During research we utilized the equipment of the resource centers “Molecular and Cell Technologies” of the Research Park of SPbU and “Taxon” Research Resource Center (http://www.ckp-rf.ru/ckp/3038/ (accessed on 10 November 2022)) of ZIN RAS.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Pearson, J.C. A phylogeny of life-cycle patterns of the Digenea. Adv. Parasitol. 1972, 10, 153–189. [Google Scholar] [PubMed]
- Cribb, T.H.; Bray, R.A.; Kudlai, P.D.; Littlewood, D.T.J. Life cycle evolution in the Digenea: A new perspective from phylogeny. Adv. Parasitol. 2003, 54, 197–254. [Google Scholar] [PubMed] [Green Version]
- Galaktionov, K.V.; Dovrovolskij, A.A. The Biology and Evolution of Trematodes: An Essay on the Biology, Morphology, Life Cycles, Transmissions, and Evolution of Digenetic Trematodes, 1st ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2003; 592p. [Google Scholar]
- Stunkard, H.W. The life history of Cryptocotyle lingua (Creplin), with notes on the physiology of the metacercariae. J. Morphol. 1930, 50, 143–191. [Google Scholar] [CrossRef]
- Cable, R.M. A new marine cercaria from the Woods Hole region and its bearing on the interpretation of larval types in the Fellodistomatidae (Trematoda: Digenea). Biol. Bull. 1954, 106, 15–20. [Google Scholar] [CrossRef]
- Ito, J.; Shimura, S. On a new lepocreadiid cercaria, Cercaria isoninae n. sp. (Trematoda) from a littoral gastropod, Japeuthria ferrea from Kanagawa and Chiba Prefectures, Japan. Jpn. J. Parasitol. 1980, 29, 181–187. [Google Scholar]
- Cable, R.M. Marine cercariae of Puerto Rico. Sci. Surv. Porto Rico Virgin Isl. 1956, 16, 491–577. [Google Scholar]
- Galaktionov, K.V.; Olenev, A.V.; Dobrovolskij, A.A. Two species of cyathocothylate cercariae from the freshwater snail Melanopsis praemorsa. Parazitologiia 1980, 14, 299–307. (In Russian) [Google Scholar]
- Cable, R.M.; Nahhas, F.M. Bivesicula caribbensis sp. n. (Trematoda: Digenea) and its life history. J. Parasitol. 1962, 48, 536–538. [Google Scholar] [CrossRef]
- Matthews, B.F. Cercaria vaullegeardi Pelseneer, 1906 (Digenea: Hemiuridae); the infection mechanism. Parasitology 1981, 83, 587–593. [Google Scholar] [CrossRef]
- Zelmer, D.A.; Esch, G.W. The infection mechanism of the cystophorous cercariae of Halipegus occidualis (Digenea: Hemiuridae). Invertebr. Biol. 1998, 117, 281–287. [Google Scholar] [CrossRef]
- Hunninen, A.V.; Cable, R.M. The life history of Podocotyle atomon (Rudolphi) (Trematoda: Opecoelidae). Trans. Am. Microsc. Soc. 1943, 62, 57–68. [Google Scholar] [CrossRef]
- Køie, M. On the morphology and life-history of Lepidapedon elongatum (Lebour, 1908) Nicoll, 1910 (Trematoda, Lepocreadiidae). Ophelia 1985, 24, 135–153. [Google Scholar] [CrossRef]
- Bartoli, P.; Jousson, O.; Russell-Pinto, F. The life cycle of Monorchis parvus (Digenea: Monorchiidae) demonstrated by developmental and molecular data. J. Parasitol. 2000, 86, 479–489. [Google Scholar] [CrossRef] [PubMed]
- Galaktionov, K.V.; Blasco-Costa, I. Microphallus ochotensis sp. nov. (Digenea, Microphallidae) and relative merits of two-host microphallid life cycles. Parasitol. Res. 2018, 117, 1051–1068. [Google Scholar] [CrossRef] [PubMed]
- Kremnev, G.; Gonchar, A.; Krapivin, V.; Uryadova, A.; Miroliubov, A.; Krupenko, D. Life cycle truncation in Digenea, a case study of Neophasis spp. (Acanthocolpidae). Int. J. Parasitol. Parasites Wildl. 2021, 15, 158–172. [Google Scholar] [CrossRef]
- Bray, R.A.; Justine, J.L. A review of the Zoogonidae (Digenea: Microphalloidea) from fishes of the waters around New Caledonia, with the description of Overstreetia cribbi n. sp. PeerJ 2014, 2, e292. [Google Scholar] [CrossRef] [Green Version]
- Miller Jr, H.M.; Northup, F.E. The seasonal infestation of Nassa obsoleta (Say) with larval trematodes. Biol. Bull. 1926, 50, 490–508. [Google Scholar] [CrossRef]
- Køie, M. On the morphology and life-history of Zoogonoides viviparus (Olsson, 1868) Odhner, 1902 (Trematoda, Zoogonidae). Ophelia 1976, 15, 1–14. [Google Scholar]
- Palombi, A. Il ciclo biologico di Diphterostomum brusinae Stossich (Trematode digenetico: Fam. Zoogonidae Odhner). Considerazioni sui cicli evolutivi dell especie affini e dei trematodi in generale. Pubbl. Stn. Zool. Napoli 1930, 10, 111–149. (In Italian) [Google Scholar]
- Stunkard, H.W. Distomum lasium Leidy, 1891 (syn. Cercariaeum lintoni Miller and Northup, 1926), the larval stage of Zoogonus rubellus (Olsson, 1868) (syn. Z. mirus Looss, 1901). Biol. Bull. 1938, 75, 308–334. [Google Scholar] [CrossRef]
- Stunkard, H.W. The morphology and life history of the digenetic trematode, Zoogonoides laevis Linton, 1940. Biol. Bull. 1943, 85, 227–237. [Google Scholar] [CrossRef]
- Gilardoni, C.; Etchegoin, J.; Cribb, T.; Pina, S.; Rodrigues, P.; Diez, M.E.; Cremonte, F. Cryptic speciation of the zoogonid digenean Diphterostomum flavum n. sp. demonstrated by morphological and molecular data. Parasite 2020, 27, PMC7301638. [Google Scholar] [CrossRef]
- Pina, S.; Tajdari, J.; Russell-Pinto, F.; Rodrigues, P. Morphological and molecular studies on life cycle stages of Diphtherostomum brusinae (Digenea: Zoogonidae) from northern Portugal. J. Helminthol. 2009, 83, 321–331. [Google Scholar] [CrossRef]
- Bray, R.A. Digenean parasites of deep-sea teleosts: A progress report. Int. J. Parasitol. Parasites Wildl. 2020, 12, 251–264. [Google Scholar] [CrossRef] [PubMed]
- Kremnev, G.; Gonchar, A.; Krapivin, V.; Knyazeva, O.; Krupenko, D. First elucidation of the life cycle in the family Brachycladiidae (Digenea), parasites of marine mammals. Int. J. Parasitol. 2020, 50, 997–1009. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, S.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Hernández-Mena, D.I.; García-Varela, M.; de León, G.P.P. Filling the gaps in the classification of the Digenea Carus, 1863: Systematic position of the Proterodiplostomidae Dubois, 1936 within the superfamily Diplostomoidea Poirier, 1886, inferred from nuclear and mitochondrial DNA sequences. Syst. Parasitol. 2017, 94, 833–848. [Google Scholar] [CrossRef] [PubMed]
- Tkach, V.; Grabda-Kazubska, B.; Pawlowski, J.; Swiderski, Z. Molecular and morphological evidence for close phylogenetic affinities of the genera Macrodera, Leptophallus, Metaleptophallus and Paralepoderma [Digenea, Plagiorchiata]. Acta Parasitol. 1999, 44, 170–179. [Google Scholar]
- Olson, P.D.; Cribb, T.H.; Tkach, V.V.; Bray, R.A.; Littlewood, D.T.J. Phylogeny and classification of the Digenea (Platyhelminthes: Trematoda). Int. J. Parasitol. 2003, 33, 733–755. [Google Scholar] [CrossRef]
- Morgan, J.A.T.; Blair, D. Nuclear rDNA ITS sequence variation in the trematode genus Echinostoma: An aid to establishing relationships within the 37-collar-spine group. Parasitology 1995, 111, 609–615. [Google Scholar] [CrossRef] [PubMed]
- Lebour, M.V. A trematode larva from Buccinum undatum and notes on trematodes from post-larval fish. J. Mar. Biol. Assoc. UK 1918, 11, 514–518. [Google Scholar] [CrossRef] [Green Version]
- Cutmore, S.C.; Miller, T.L.; Bray, R.A.; Cribb, T.H. A new species of Plectognathotrema Layman, 1930 (Trematoda: Zoogonidae) from an Australian monacanthid, with a molecular assessment of the phylogenetic position of the genus. Syst. Parasitol. 2014, 89, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Sokolov, S.; Gordeev, I.; Lebedeva, D. Redescription of Proctophantastes gillissi (Overstreet et Pritchard, 1977) (Trematoda: Zoogonidae) with discussion on the systematic position of the genus Proctophantastes Odhner, 1911. Acta Parasitol. 2016, 61, 529–536. [Google Scholar] [CrossRef]
- Sokolov, S.; Shchenkov, S.; Gordeev, I.; Ryazanova, T. Description of a metacercaria of a zoogonid trematode Steganoderma cf. eamiqtrema Blend and Racz, 2020 (Microphalloidea: Zoogonidae), with notes on the phylogenetic position of the genus Steganoderma Stafford, 1904, and resurrection of the subfamily Lecithostaphylinae Odhner, 1911. Parasitol. Res. 2021, 120, 1669–1676. [Google Scholar]
- Atopkin, D.M.; Besprozvannykh, V.V.; Ha, N.D.; Nguyen, H.V.; Nguyen, T.V. New trematode species Lecithostaphylus halongi n. sp. (Zoogonidae, Microphalloidea) and Gymnotergestia strongyluri n. sp. (Fellodistomidae, Gymnophalloidea) from beloniform fishes in Vietnam. J. Helminthol. 2022, 96, 529–536. [Google Scholar] [CrossRef]
- Cutmore, S.C.; Bray, R.A.; Cribb, T.H. Two new species of Bacciger Nicoll, 1914 (Trematoda: Faustulidae) in species of Herklotsichthys Whitley (Clupeidae) from Queensland waters. Syst. Parasitol. 2018, 95, 645–654. [Google Scholar] [CrossRef]
- Sokolov, S.G.; Shchenkov, S.V.; Gordeev, I.I. A phylogenetic assessment of Pronoprymna spp. (Digenea: Faustulidae) and Pacific and Antarctic representatives of the genus Steringophorus Odhner, 1905 (Digenea: Fellodistomidae), with description of a new species. J. Nat. Hist. 2021, 55, 867–887. [Google Scholar] [CrossRef]
- Curran, S.S.; Warren, M.B.; Bullard, S.A. Description of a New Species of Bacciger (Digenea: Gymnophalloidea) Infecting the American Gizzard Shad, Dorosoma cepedianum (Lesueur, 1818), and Molecular Characterization of Cercaria rangiae Wardle, 1983, with Molecular Phylogeny of Related Digenea. Comp. Parasitol. 2022, 89, 9–29. [Google Scholar] [CrossRef]
- Bray, R.A.; Gibson, D.I. The Zoogonidae (Digenea) of fishes from the northeast Atlantic. Bull. Br. Mus. Nat. Hist. Zool. 1986, 51, 127–206. [Google Scholar]
- Orrhage, L. Description of the Metacercaria of Zoogonoides viviparus (Olsson, 1868) Odhner, 1902 with Some Remarks on Life Cycles in the Genus Zoogonoides (Trematoda, Digenea, Zoogonidae). Zool. Scr. 1974, 2, 179–182. [Google Scholar] [CrossRef]
- Køie, M. On the histochemistry and ultrastructure of the tegument and associated structures of the cercaria of Zoogonoides viviparus in the first intermediate host. Ophelia 1971, 9, 165–206. [Google Scholar] [CrossRef]
- Zhukov, E.V. Endoparasitic worms of the fishes in the Sea of Japan and South-Kuril shallow-waters. Tr. Zool. Inst. 1960, 28, 3–146. (In Russian) [Google Scholar]
- Scott, J.S. Incidence of trematode parasites of American plaice (Hippoglossoides platessoides) of the Scotian Shelf and Gulf of St. Lawrence in relation to fish length and food. Can. J. Fish Aquat. Sci. 1975, 32, 479–483. [Google Scholar] [CrossRef]
- Scott, J.S. Digenean parasite communities in flatfishes of the Scotian Shelf and southern Gulf of St. Lawrence. Can. J. Zool. 1982, 60, 2804–2811. [Google Scholar] [CrossRef]
- Sirenko, B.I. List of Species of Free-Living Invertebrates of Eurasian Arctic Seas and Adjacent Deep Waters; Russian Academy of Science, Zoological Institute: Saint-Petersburg, Russia, 2001; 134p. [Google Scholar]
- Bray, R.A. A study of the helminth parasites of Anarhichas lupus (Perciformes: Anarhichadidae) in the North Atlantic. J. Fish Biol. 1987, 31, 237–264. [Google Scholar] [CrossRef]
- Falk-Petersen, I.B.; Kanapathippilai, P.; Primicerio, R.; Hansen, T.K. Size, locality and seasonally related feeding preferences of common wolffish (Anarhichas lupus L.) from north-Norwegian waters. Mar. Biol. Res. 2010, 6, 201–212. [Google Scholar] [CrossRef]
- Shulman-Albova, R.E. Fish parasites of the White Sea around the village of Gridina. Part I. Monogenetic and digenetic trematodes. Uchenye Zap. Karelo-Fin. Gos. Univ. 1952, 4, 78–97. (In Russian) [Google Scholar]
- Barnett, L.J.; Miller, T.L.; Cribb, T.H. A review of the currently recognised zoogonid cercariae, including the identification and emergence ecology of Cercaria capricornia XI (Digenea: Zoogonidae) from Nassarius olivaceus (Gastropoda: Nassariidae) in Central Queensland, Australia. Folia Parasitol. 2014, 61, 322–330. [Google Scholar] [CrossRef]
- Pelseneer, P. Trematodes parasites de mollusques marins. Bull. Biol. Fr. Belg. 1906, 40, 161–186. (In French) [Google Scholar]
- Galaktionov, K.V.; Blasco-Costa, I.; Olson, P.D. Life cycles, molecular phylogeny and historical biogeography of the ‘pygmaeus’ microphallids (Digenea: Microphallidae): Widespread parasites of marine and coastal birds in the Holarctic. Parasitology 2012, 139, 1346–1360. [Google Scholar] [CrossRef] [Green Version]
- Galaktionov, K.V.; Malkova, I.I. The glands of trematode cercariae of the family Microphallidae Travassos, 1920. Int. J. Parasitol. 1994, 24, 595–604. [Google Scholar] [CrossRef]
- Shchenkov, S.V.; Denisova, S.A.; Smirnova, A.D.; Shunatova, N.N. Mucoid glands of cercariae. Invert Zool. 2019, 16, 377–392. (In Russian) [Google Scholar] [CrossRef]
- Krupenko, D.; Gonchar, A.; Kremnev, G.; Efeykin, B.; Krapivin, V. New type of xiphidiocercariae (Digenea: Microphalloidea) from South Vietnam. Folia Parasitol. 2020, 67, 1–6. [Google Scholar] [CrossRef]
- Shimura, S.; Ito, J. Two new cercariae, Cercaria brachycaeca n. sp. and Cercaria misakiana n. sp. from top shells, Batillus cornutus and Marmarostoma stenogyrum, with notes of their effects on the hosts. Jpn. J. Parasitol. 1980, 29, 69–76. [Google Scholar]
- Selbach, C.; Poulin, R. Some like it hotter: Trematode transmission under changing temperature conditions. Oecologia 2020, 194, 745–755. [Google Scholar] [CrossRef]
- Ulmer, M.J. Postharmostomum helicis (Leidy, 1847) Robinson 1949, (Trematoda), Its Life History and a Revision of the Subfamily Brachylaeminae: Part I. Trans. Am. Microsc. Soc. 1951, 70, 189–238. [Google Scholar] [CrossRef]
- González-Moreno, O.; Gracenea, M. Life cycle and description of a new species of brachylaimid (Trematoda: Digenea) in Spain. J. Parasitol. 2006, 92, 1305–1312. [Google Scholar] [CrossRef]
- Chernogorenko, M.I.; Komarova, T.I.; Kurandina, D.P. The life cycle of the trematode Plagioporus skrjabini Kgwal, 1951 (Allocreadiata, Opecoelidae). Parazitologiia 1978, 12, 479–486. (In Russian) [Google Scholar]
- Køie, M. On the morphology and life-history of Podocotyle reflexa (Creplin, 1825) Odhner, 1905, and a comparison of its developmental stages with those of P. atomon (Rudolphi, 1802) Odhner, 1905 (Trematoda, Opecoelidae). Ophelia 1981, 20, 17–43. [Google Scholar] [CrossRef]
- Prokofiev, V.V. Influence of temperature and salinity on a life span of cercariae of marine litoral trematodes Cryptocotyle sp. (Heterophyidae), Levinseniella brachysoma and Maritrema subdolum (Microphallidae). Parazitologiia 1999, 33, 520–526. [Google Scholar]
- Shaw, C.R. Observations on Cercariaeum lintoni Miller and Northup and its metacercarial development. Biol. Bull. 1933, 64, 262–275. [Google Scholar] [CrossRef]
- Madhavi, R.; Shameem, U. Cercaria chilkaensis II, a new zoogonid cercaria from the snail, Nassarius orissaensis, from Chilka Lake, India. Proc. Helminthol. Soc. Wash. 1991, 58, 31–34. [Google Scholar]
- Gilardoni, C.; Etchegoin, J.; Diaz, J.I.; Ituarte, C.; Cremonte, F. A survey of larval digeneans in the commonest intertidal snails from Northern Patagonian coast, Argentina. Acta Parasitol. 2011, 56, 163–179. [Google Scholar] [CrossRef] [Green Version]
- Køie, M. On the endoparasites of Buccinum undatum L. with special reference to the trematodes. Ophelia 1969, 6, 251–279. [Google Scholar] [CrossRef]
- Lebour, M.V. A review of the British marine cercariae. Parasitology 1911, 4, 416–456. [Google Scholar] [CrossRef]
- Stunkard, H.W. Specificity and host-relations in the trematode genus Zoogonus. Biol. Bull. 1941, 81, 205–214. [Google Scholar] [CrossRef]
- Prévot, G. Sur deux trématodes larvaires d’Antedon mediterranea Lmk. (Echinoderme): Metacercaria sp. (Monorchiidae Odhner, 1911), et métacercaire de Diphterostomum brusinae Stoss., 1904 (Zoogonidae Odhner, 1911). Ann. Parasitol. Hum. Comp. 1966, 41, 233–242. (In French) [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francisco, C.J.; Almeida, A.; Castro, A.M.; Pina, S.; Russell-Pinto, F.; Rodrigues, P.; Santos, M.J. Morphological and molecular analysis of metacercariae of Diphtherostomum brusinae (Stossich, 1888) Stossich, 1903 from a new bivalve host Mytilus galloprovincialis. J. Helminthol. 2011, 85, 179–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martorelli, S.R.; Montes, M.M.; Marcotegui, P.S.; Alda, M.D.P. Primer registro de Diphterostomum brusinae (Digenea, Zoogonidae) parasitando a la corvina Micropogonias furnieri con datos sobre su ciclo biológico. Rev. Argent. Parasitol. 2014, 2, 22–27. (In Spanish) [Google Scholar]
- Uspenskaya, A.V. Parasitic Fauna of Benthic Crustaceans from the Barents Sea; Academia Nauk SSSR, Kolskij Filial: Moskva, Russia, 1963; 128p. (In Russian)
- Kagei, N.; Kon, T. Metacercariae of genus Steganoderma (Trematoda) in a crab, Chionoecetes opilio (O. Fabricius), from Northern-Sea. Jpn. J. Parasitol. 1978, 27, 27–30. [Google Scholar]
- Holton, A.L. Observations on the life history of Deretrema minutum Manter, 1954 (Digenea: Zoogonidae) in freshwater crustacean and fish hosts from Canterbury, New Zealand. N. Z. J. Mar. Freshw. Res. 1983, 17, 373–376. [Google Scholar] [CrossRef] [Green Version]
- Marcogliese, D.J. Evidence for specificity of Steganoderma formosum for its second intermediate host in the Northwest Atlantic. J. Helminthol. 1996, 70, 215–218. [Google Scholar] [CrossRef]
- Abdul-Salam, J.; Sreelatha, B.S. Studies on Cercariae from Kuwait Bay. IX. Description and surface topography of Cercaria kuwaitae IX sp. n. (Digenea: Zoogonidae). J. Helminthol. Soc. Wash. 1998, 65, 141–146. [Google Scholar]
- Briscoe, A.G.; Bray, R.A.; Brabec, J.; Littlewood, D.T.J. The mitochondrial genome and ribosomal operon of Brachycladium goliath (Digenea: Brachycladiidae) recovered from a stranded minke whale. Parasitol. Int. 2016, 65, 271–275. [Google Scholar] [CrossRef]
- Tkach, V.V.; Snyder, S.D. Choanocotyle platti sp. nov. from the northern long-necked turtle, Chelodina rugosa (Pleurodira, Chelidae) in Australia. Acta Parasitol. 2007, 52, 318–324. [Google Scholar] [CrossRef] [Green Version]
- Heneberg, P.; Literák, I. Molecular phylogenetic characterization of Collyriclum faba with reference to its three host-specific ecotypes. Parasitol. Int. 2013, 62, 262–267. [Google Scholar] [CrossRef]
- Unwin, S.; Chantrey, J.; Chatterton, J.; Aldhoun, J.A.; Littlewood, D.T.J. Renal trematode infection due to Paratanaisia bragai in zoo housed Columbiformes and a red bird-of-paradise (Paradisaea rubra). Int. J. Parasitol. Parasites Wildl. 2013, 2, 32–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shchenkov, S.V.; Denisova, S.A.; Kremnev, G.A.; Dobrovolskij, A.A. Five new morphological types of virgulate and microcotylous xiphidiocercariae based on morphological and molecular phylogenetic analyses. J. Helminthol. 2020, 94, E94. [Google Scholar] [CrossRef]
- Cribb, T.H.; Bray, R.A.; Hall, K.A.; Cutmore, S.C. A review of the genus Antorchis Linton, 1911 (Trematoda: Faustulidae) from Indo-Pacific fishes with the description of a new species. Syst. Parasitol. 2015, 92, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Tkach, V.; Pawlowski, J.; Mariaux, J.; Świderski, Z. Molecular phylogeny of the suborder Plagiorchiata and its position in the system of Digenea. In Interrelationships of the Platyhelminthes; Littlewood, D.T.J., Bray, R.A., Eds.; Taylor and Francis: London, UK, 2001; pp. 186–193. [Google Scholar]
- Bell, J.A.; González-Acuña, D.; Tkach, V.V. First record of Gyrabascus (Digenea, Pleurogenidae) from Dromiciops bozinovici D’Elia et al., 2016 (Marsupialia: Microbiotheriidae) in Chile and its phylogenetic relationships. Comp. Parasitol. 2018, 85, 58–65. [Google Scholar] [CrossRef]
- Kasl, E.L.; Font, W.F.; Criscione, C.D. Resolving evolutionary changes in parasite life cycle complexity: Molecular phylogeny of the trematode genus Alloglossidium indicates more than one origin of precociousness. Mol. Phylogenet. Evol. 2018, 126, 371–381. [Google Scholar] [CrossRef]
- Kudlai, O.; Cribb, T.H.; Cutmore, S.C. A new species of microphallid (Trematoda: Digenea) infecting a novel host family, the Muraenidae, on the northern Great Barrier Reef, Australia. Syst. Parasitol. 2016, 93, 863–876. [Google Scholar] [CrossRef]
- Tkach, V.; Pawlowski, J.; Mariaux, J. Phylogenetic analysis of the suborder Plagiorchiata (Platyhelminthes, Digenea) based on partial lsrDNA sequences. Int. J. Parasitol. 2000, 30, 83–93. [Google Scholar] [CrossRef]
- Tkach, V.V.; Littlewood, D.T.J.; Olson, P.D.; Kinsella, J.M.; Swiderski, Z. Molecular phylogenetic analysis of the Microphalloidea Ward, 1901 (Trematoda: Digenea). Syst. Parasitol. 2003, 56, 1–15. [Google Scholar] [CrossRef]
- Tkach, V.; Grabda-Kazubska, B.; Swiderski, Z. Systematic position and phylogenetic relationships of the family Omphalometridae (Digenea, Plagiorchiida) inferred from partial lsrDNA sequences. Int. J. Parasitol. 2001, 31, 81–85. [Google Scholar] [CrossRef]
- Razo-Mendivil, U.J.; León-Règagnon, V.; de Leon, G.P.P. Monophyly and systematic position of Glypthelmins (Digenea), based on partial lsrDNA sequences and morphological evidence. Org. Divers. Evol. 2006, 6, 308–320. [Google Scholar] [CrossRef] [Green Version]
- Zikmundová, J.; Georgieva, S.; Faltýnková, A.; Soldánová, M.; Kostadinova, A. Species diversity of Plagiorchis Lühe, 1899 (Digenea: Plagiorchiidae) in lymnaeid snails from freshwater ecosystems in central Europe revealed by molecules and morphology. Syst. Parasitol. 2014, 88, 37–54. [Google Scholar] [CrossRef]
- Kanarek, G.; Zaleśny, G.; Czujkowska, A.; Sitko, J.; Harris, P.D. On the systematic position of Collyricloides massanae Vaucher, 1969 (Platyhelminthes: Digenea) with notes on distribution of this trematode species. Parasitol. Res. 2015, 114, 1495–1501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pulis, E.E.; Tkach, V.V.; Newman, R.A. Helminth parasites of the wood frog, Lithobates sylvaticus, in prairie pothole wetlands of the Northern Great Plains. Wetlands 2011, 31, 675–685. [Google Scholar] [CrossRef]
- O’Dwyer, K.; Blasco-Costa, I.; Poulin, R.; Faltýnková, A. Four marine digenean parasites of Austrolittorina spp. (Gastropoda: Littorinidae) in New Zealand: Morphological and molecular data. Syst. Parasitol. 2014, 89, 133–152. [Google Scholar] [CrossRef] [PubMed]
- Cabañas-Granillo, J.; Solórzano-García, B.; Mendoza-Garfias, B.; Pérez-Ponce de León, G. A new species of Lecithostaphylus Odhner, 1911 (Trematoda: Zoogonidae) from the Pacific needlefish, Tylosurus pacificus, off the Pacific coast of Mexico, with a molecular assessment of the phylogenetic position of this genus within the family. Mar. Biodivers. 2020, 50, 83. [Google Scholar] [CrossRef]
- Lockyer, A.E.; Olson, P.D.; Littlewood, D.T.J. Utility of complete large and small subunit rRNA genes in resolving the phylogeny of the Neodermata (Platyhelminthes): Implications and a review of the cercomer theory. Biol. J. Linn. Soc. Lond. 2003, 78, 155–171. [Google Scholar] [CrossRef] [Green Version]
- Tkach, V.V.; Greiman, S.E.; Pulis, E.E.; Brooks, D.R.; Bonilla, C.C. Phylogenetic relationships and systematic position of the enigmatic Urotrema Braun, 1900 (Platyhelminthes: Digenea). Parasitol. Int. 2019, 70, 118–122. [Google Scholar] [CrossRef]
Figure 1.
Maximum likelihood phylogenetic trees based on the 28S and 5.8S+ITS2 rDNA sequence data. Bootstrap values are printed in nodes. GenBank accession numbers of newly generated sequences are indicated in bold. Blue and yellow squares mark life cycle stages of Zoogonoides viviparus and Pseudozoogonoides subaequiporus, respectively. Scale bar shows the substitution rate. Abbreviations: AL—Anarhichas lupus; BU—Buccinum undatum; BS—Buccinum scalariforme; CC—Ciliatocardium ciliatum; CL—Callionymus lyra; LL—Limanda limanda; ND—Neptunea despecta; NE—Nephtys sp.; NP—Nuculana pernula.
Figure 1.
Maximum likelihood phylogenetic trees based on the 28S and 5.8S+ITS2 rDNA sequence data. Bootstrap values are printed in nodes. GenBank accession numbers of newly generated sequences are indicated in bold. Blue and yellow squares mark life cycle stages of Zoogonoides viviparus and Pseudozoogonoides subaequiporus, respectively. Scale bar shows the substitution rate. Abbreviations: AL—Anarhichas lupus; BU—Buccinum undatum; BS—Buccinum scalariforme; CC—Ciliatocardium ciliatum; CL—Callionymus lyra; LL—Limanda limanda; ND—Neptunea despecta; NE—Nephtys sp.; NP—Nuculana pernula.

Figure 2.
Phylogenetic position of Pseudozoogonoides subaequiporus (indicated in bold) based on 28S rDNA sequence data analysis inferred with Bayesian inference (BI) and maximum likelihood (ML). Nodal support: ML/BI. Scale bar shows the substitution rate.
Figure 2.
Phylogenetic position of Pseudozoogonoides subaequiporus (indicated in bold) based on 28S rDNA sequence data analysis inferred with Bayesian inference (BI) and maximum likelihood (ML). Nodal support: ML/BI. Scale bar shows the substitution rate.

Figure 7.
Life cycle schemes of Zoogonoides viviparus (A) and Pseudozoogonoides subaequiporus (B) based on the data retrieved at the White Sea.
Figure 7.
Life cycle schemes of Zoogonoides viviparus (A) and Pseudozoogonoides subaequiporus (B) based on the data retrieved at the White Sea.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
List of the dissected White Sea benthic invertebrates and their infection with zoogonid life cycle stages.
Table 1.
List of the dissected White Sea benthic invertebrates and their infection with zoogonid life cycle stages.
| Host | Number of Dissected Individuals | Zoogonid Infection | ||
|---|---|---|---|---|
| Gastropoda | Buccinidae | Buccinum undatum Linnaeus, 1758 | 2174 | + |
| B. scalariforme Møller, 119 | 19 | + | ||
| Neptunea despecta (Linnaeus, 1758) | 103 | + | ||
| Naticidae | Cryptonatica affinis (Gmelin, 1791) | 270 | - | |
| Euspira pallida (Broderip & GB Sowerby I, 1829) | 24 | - | ||
| Amauropsis islandica (Gmelin, 1791) | 3 | - | ||
| Capulidae | Ariadnaria borealis (Broderip & G. B. Sowerby I, 1829) | 28 | - | |
| Margaritidae | Margarites helicinus (Phipps, 1774) | 17 | - | |
| Muricidae | Boreotrophon clavatus (G. O. Sars, 1878) | 8 | - | |
| Annelida | Nereididae | Nereis pelagica Linnaeus, 1758 | 11 | - |
| Pectinariidae | Cistenides hyperborea Malmgren, 1866 | 9 | - | |
| Nephtyidae | Nephtys sp. | 8 | + | |
| Polynoidae | Lepidonotus squamatus (Linnaeus, 1758) | 5 | - | |
| Orbiniidae | Scoloplos armiger (Müller, 1776) | 4 | - | |
| Trichobranchidae | Terebellides stroemii Sars, 1835 | 1 | - | |
| Lumbrineridae | Scoletoma fragilis (O.F. Müller, 1776) | 1 | - | |
| Bivalvia | Mytilidae | Musculus discors (Linnaeus, 1767) | 31 | - |
| Cardiidae | Serripes groenlandicus (Mohr, 1786) | 66 | + | |
| Ciliatocardium ciliatum (Fabricius, 1780) | 39 | + | ||
| Nuculanidae | Nuculana pernula (O. F. Müller, 1779) | 394 | + | |
| Yoldiidae | Yoldia hyperborea (Gould, 1841) | 78 | + | |
| Portlandia arctica (Gray, 1824) | 6 | - | ||
| Nuculidae | Ennucula tenuis (Montagu, 1808) | 5 | - | |
| Decapoda | Paguridae | Pagurus pubescens Krøyer, 1838 | 12 | - |
| Pandalidae | Pandalus montagui Leach, 1814 | 8 | - | |
| Thoridae | Eualus gaimardii (H. Milne Edwards, 1837) | 4 | - | |
| Crangonidae | Crangon crangon (Linnaeus, 1758) | 2 | - | |
| Sabinea septemcarinata (Sabine, 1824) | 1 | - | ||
| Ophiuroidea | Ophiuridae | Ophiura robusta (Ayres, 1852) | 67 | - |
| Ophiopholidae | Ophiopholis aculeata (Linnaeus, 1767) | 40 | - | |
Table 2.
Isolates of putative and identified Zoogonidae from the White Sea, their origin and GenBank accession numbers.
Table 2.
Isolates of putative and identified Zoogonidae from the White Sea, their origin and GenBank accession numbers.
| ID | Species | Stage | Host Species | GenBank Accession Numbers | |
|---|---|---|---|---|---|
| 28S rDNA | 5.8S rDNA+ITS2 | ||||
| G7.3 | Zoogonoides viviparus | Marita | Limanda limanda | OP956058 | OP956013 |
| G7.2 | Z. viviparus | Marita | Anarhichas lupus | OP956059 | OP956014 |
| A5.36 | Z. viviparus | Marita | L. limanda | OP956060 | OP956015 |
| G7.8 | Putative Z. viviparus | Metacercaria | Nephthys sp. | OP956061 | OP956016 |
| G7.5 | Putative Z. viviparus | Metacercaria | Nuculana pernula | OP956062 | OP956017 |
| G7.4 | Putative Z. viviparus | Metacercaria | Ciliatocardium ciliatum | OP956063 | OP956018 |
| G7.9 | Putative Z. viviparus | Sporocyst with cercariae | Buccinum undatum | OP956064 | OP956019 |
| G7.10 | Putative Z. viviparus | Sporocyst with cercariae | B. undatum | OP956065 | OP956020 |
| A5.37 | Putative Z. viviparus | Sporocyst with cercariae | B. undatum | OP956066 | OP956021 |
| A5.38 | Putative Z. viviparus | Sporocyst with cercariae | B. undatum | OP956067 | OP956022 |
| G7.6 | Pseudozoogonoides subaequiporus | Marita | A. lupus | OP956068 | OP956023 |
| G7.7 | P. subaequiporus | Marita | A. lupus | OP956069 | OP956024 |
| D9.6 | Putative P. subaequiporus | Metacercaria | N. pernula | OP956070 | OP956025 |
| D9.7 | Putative P. subaequiporus | Metacercaria | N. pernula | OP956071 | OP956026 |
| D9.4 | Putative P. subaequiporus | Sporocyst with cercariae and metacercariae | Neptunea despecta | OP956072 | OP956027 |
| D11.3 | Putative P. subaequiporus | Sporocyst with cercariae and metacercariae | B. undatum | OP956073 | OP956028 |
| G7.1 | Lepidophyllum steenstrupi | Marita | A. lupus | OP956074 | OP956029 |
| AG4.12 | Zoogonidae gen. sp. | Sporocyst with germ balls | B. scalariforme | OP956075 | OP956030 |
Table 3.
PCR primers and thermocycling conditions; in all reactions, initial denaturation was at 95 °C for 5 min and final extension was at 72 °C for 10 min.
Table 3.
PCR primers and thermocycling conditions; in all reactions, initial denaturation was at 95 °C for 5 min and final extension was at 72 °C for 10 min.
| Product | Primer | Sequence (5′-3′), Forward (F) and Reverse (R) | Thermocycling Profile | Reference |
|---|---|---|---|---|
| 18S rDNA | 18S1A | F, GGCGATCGAAAAGATTAAGCCATGCA | 94 °C—1 m 52 °C—1 m 72 °C—1 m ×35 | [32] primers and conditions |
| 32 | R, CGAAGTCCTATTCCATTATTC | |||
| 652 | F, GCAGCCGCGGTAATTCCAGCTC | |||
| 28 | R, AGCGACGGGCGGTGTGT | |||
| 28S rDNA | digl2 | F, AAGCATATCACTAAGCGG | 95 °C—30 s 54 °C—30 s 72 °C—2 m ×40 | [33] primers and conditions |
| 1500R | R, GCTATCCTGAGGGAAACTTCG | [34] primers and conditions | ||
| 5.8S+ITS2 | 3S | F, GTACCGGTGGATCACGTGGCTAGTG | 94 °C—30 s 55 °C—30 s 72 °C—1 m ×30 | [35] primers |
| ITS2.2 | R, CCTGGTTAGTTTCTTTTCCTCCGC |
Table 4.
Dimensions of the White Sea Zoogonidae life cycle stages.
| Zoogonoides viviparus (Maritae) n = 10 | Z. viviparus (Metacercariae) n = 6 | Z. viviparus (Cercariae) n = 11 | Pseudozoogonoides subaequiporus (Maritae) n = 12 | P. subaequiporus (Cercariae) n = 11 | |
|---|---|---|---|---|---|
| Length | 766 (665–831) | 385 (341–446) | 388 (337–451) | 747 (622–929) | 301 (248–385) |
| Width | 246 (206–281) | 115 (101–137) | 139 (125–158) | 295 (237–341) | 118 (98–136) |
| Forebody, length | 273 (227–294) | 172 (151–196) | 179 (153–242) | 255 (219–304) | 128 (96–180) |
| Length:forebody ratio | 1:0.36 (0.31–0.39) | 1:0.45 (0.40–0.50) | 1:0.46 (0.43–0.54) | 1:0.34 (0.29–0.39) | 1:0.42 (0.35–0.48) |
| Oral sucker | 114 (88–137) × 127 (102–144) | 68 (59–81) × 73 (60–99), n = 7 | 73 (63–79) × 73 (65–83) | 129 (107–152) × 130 (113–146) | 58 (50–66) × 57 (50–66) |
| Ventral sucker | 187 (168–222) × 182 (164–202) | 99 (82–114) × 94 (82–132), n = 7 | 103 (93–119) × 87 (71–98) | 138 (127–156) × 137 (121–182) | 60 (54–65) × 60 (55–66) |
| Sucker ratio | 1:1.66 (1.43–1.99) | 1:1.47 (1.33–1.68) n = 7 | 1:1.40 (1.19–1.63) | 1:1.08 (0.97–1.27) | 1:1.05 (0.92–1.24) |
| Stylet | - | - | 16 (12–20) × 5 (4–6) n = 9 | - | 13 (10–16) × 4 (3–5) n = 8 |
| Pharynx | 37 (30–55) × 47 (43–50) | 23 (18–36) × 30 (24–34), n = 7 | 23 (17–36) × 29 (26–32) | 40 (28–50) × 45 (36–53) | 19 (15–21) × 23 (21–25) |
| Esophagus | 84 (71–95) | 52 (36–77) | 61 (46–76) | 78 (44–97) | 43 (26–69) |
| Cirrus-sac | 261 (210–296) × 52 (44–65) | 95 (81–115) × 23 (19–27), n = 4 | 87 (62–102) × 19 (17–23) | 278 (203–325) × 74 (64–89) | 67 (56–85) × 14 (12–15) |
| Ovary | 96 (77–121) × 72 (60–89) | 31 (21–41) × 28 (18–39), n = 5 | 28 (24–30) × 23 (20–27) | 102 (81–150) × 93 (75–144) | 18 (17–20) × 15 (14–17) |
| Testis, anterior | 87 (65–123) × 72 (53–94) | 39 (23–57) × 31 (18–47), n = 5 | 48 (42–57) × 36 (33–40) | 112 (78–144) × 83 (64–103) | 25 (16–30) × 18 (12–23) |
| Testis, posterior | 83 (67–113) × 75 (63–101) | 37 (25–57) × 31 (21–44), n = 5 | 45 (40–50) × 33 (27–37) | 99 (78–138) × 80 (67–112) | 25 (18–30) × 19 (14–23) |
| Eggs | 64 (56–76) × 28 (23–31) | - | - | 70 (61–79) × 30 (27–33), n = 10 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kremnev, G.; Gonchar, A.; Uryadova, A.; Krapivin, V.; Skobkina, O.; Gubler, A.; Krupenko, D. No Tail No Fail: Life Cycles of the Zoogonidae (Digenea). Diversity 2023, 15, 121. https://doi.org/10.3390/d15010121
AMA Style
Kremnev G, Gonchar A, Uryadova A, Krapivin V, Skobkina O, Gubler A, Krupenko D. No Tail No Fail: Life Cycles of the Zoogonidae (Digenea). Diversity. 2023; 15(1):121. https://doi.org/10.3390/d15010121
Chicago/Turabian StyleKremnev, Georgii, Anna Gonchar, Alexandra Uryadova, Vladimir Krapivin, Olga Skobkina, Arseniy Gubler, and Darya Krupenko. 2023. "No Tail No Fail: Life Cycles of the Zoogonidae (Digenea)" Diversity 15, no. 1: 121. https://doi.org/10.3390/d15010121
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.