
Statistical Assessment of Phenol Biodegradation by a Metal-Tolerant Binary Consortium of Indigenous Antarctic Bacteria


 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Culture, Maintenance and Media Preparation
2.2. Phenol Biodegradation
2.3. Design of Experiment (DoE) by Response Surface Methodology (RSM)
2.4. Heavy Metal Tolerance
3. Results and Discussion
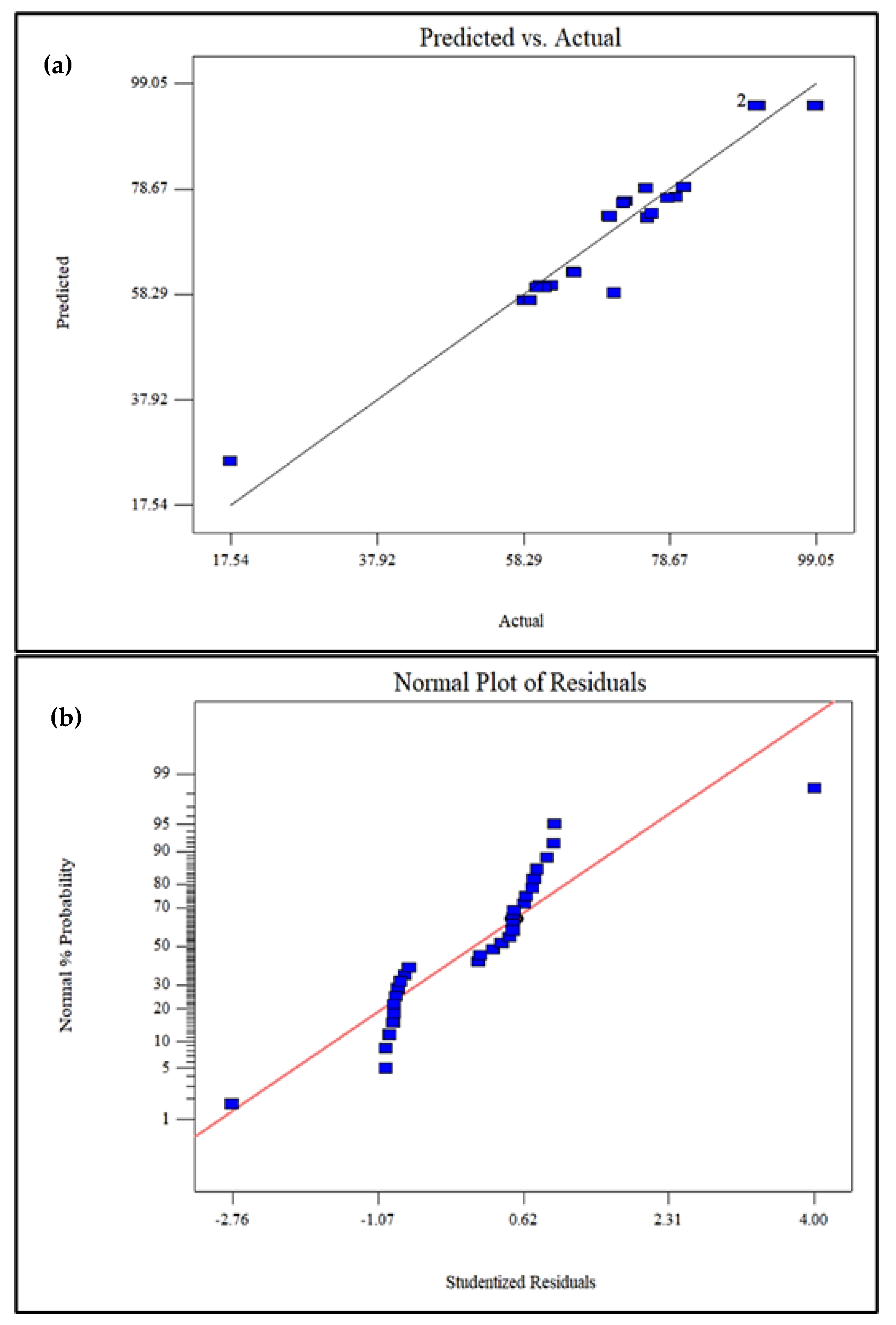
3.1. Statistical Optimization of Phenol Degradation Using RSM
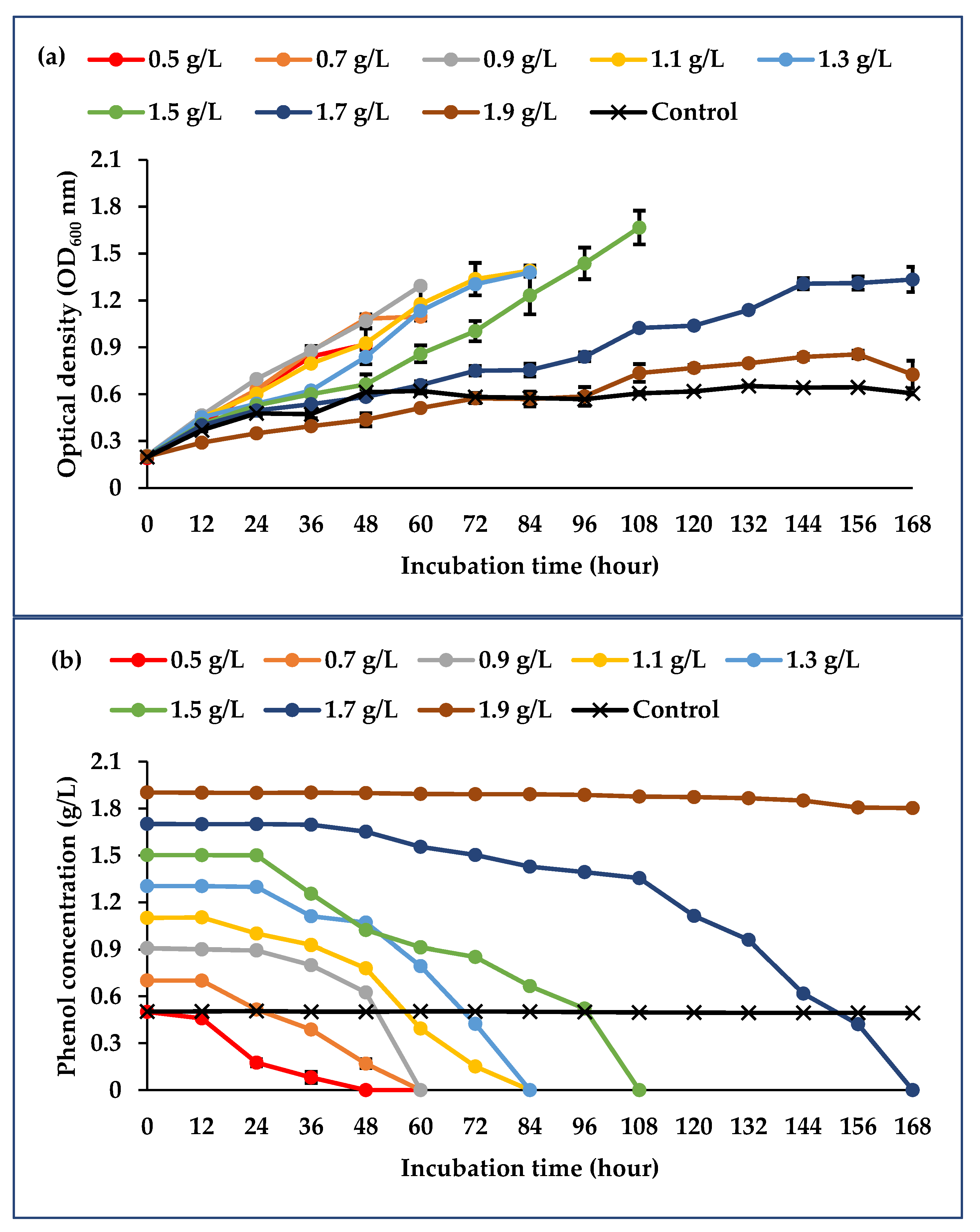
3.2. Effect of Phenol Initial Concentration on Consortium Degradation Activity
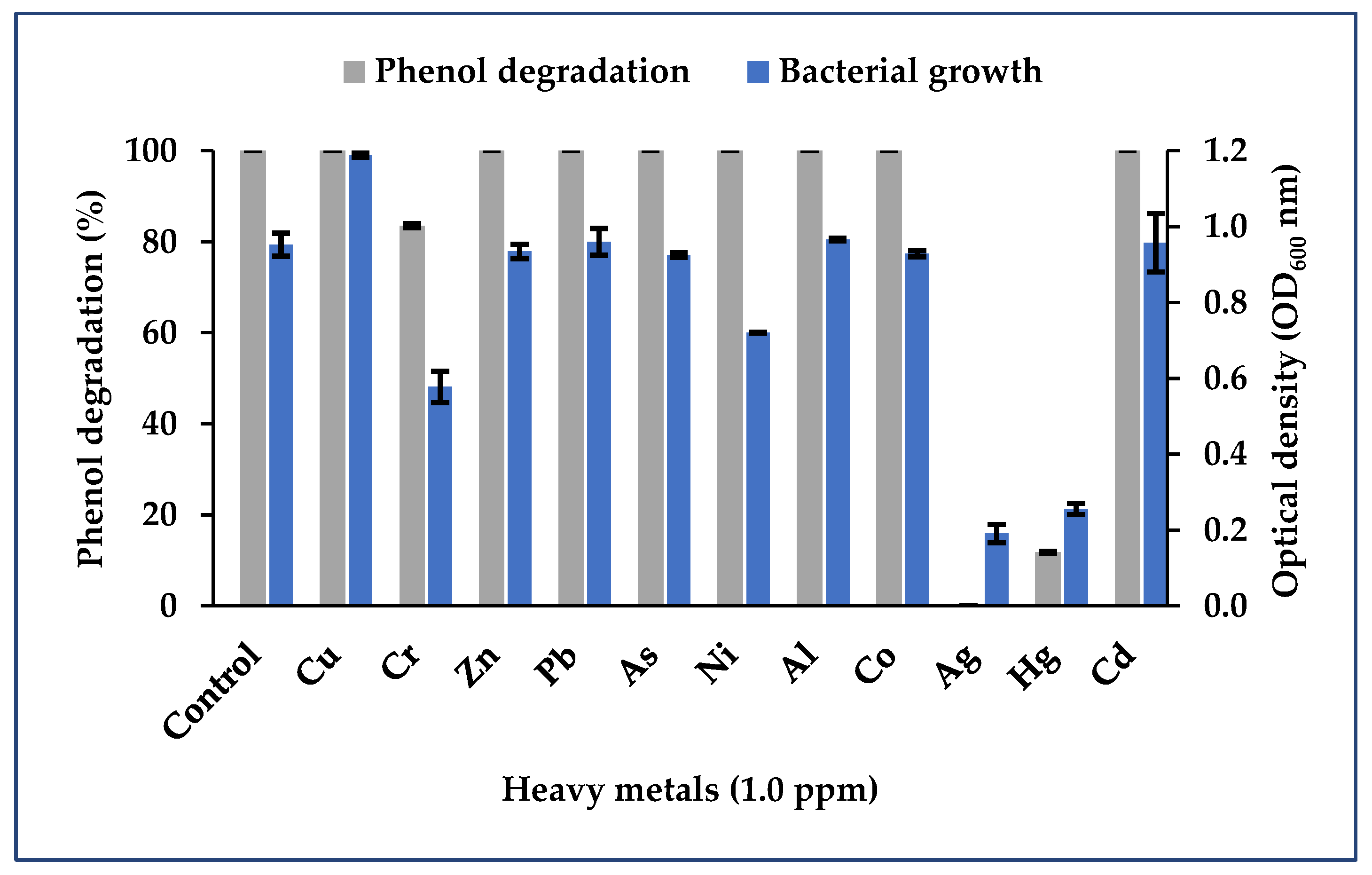
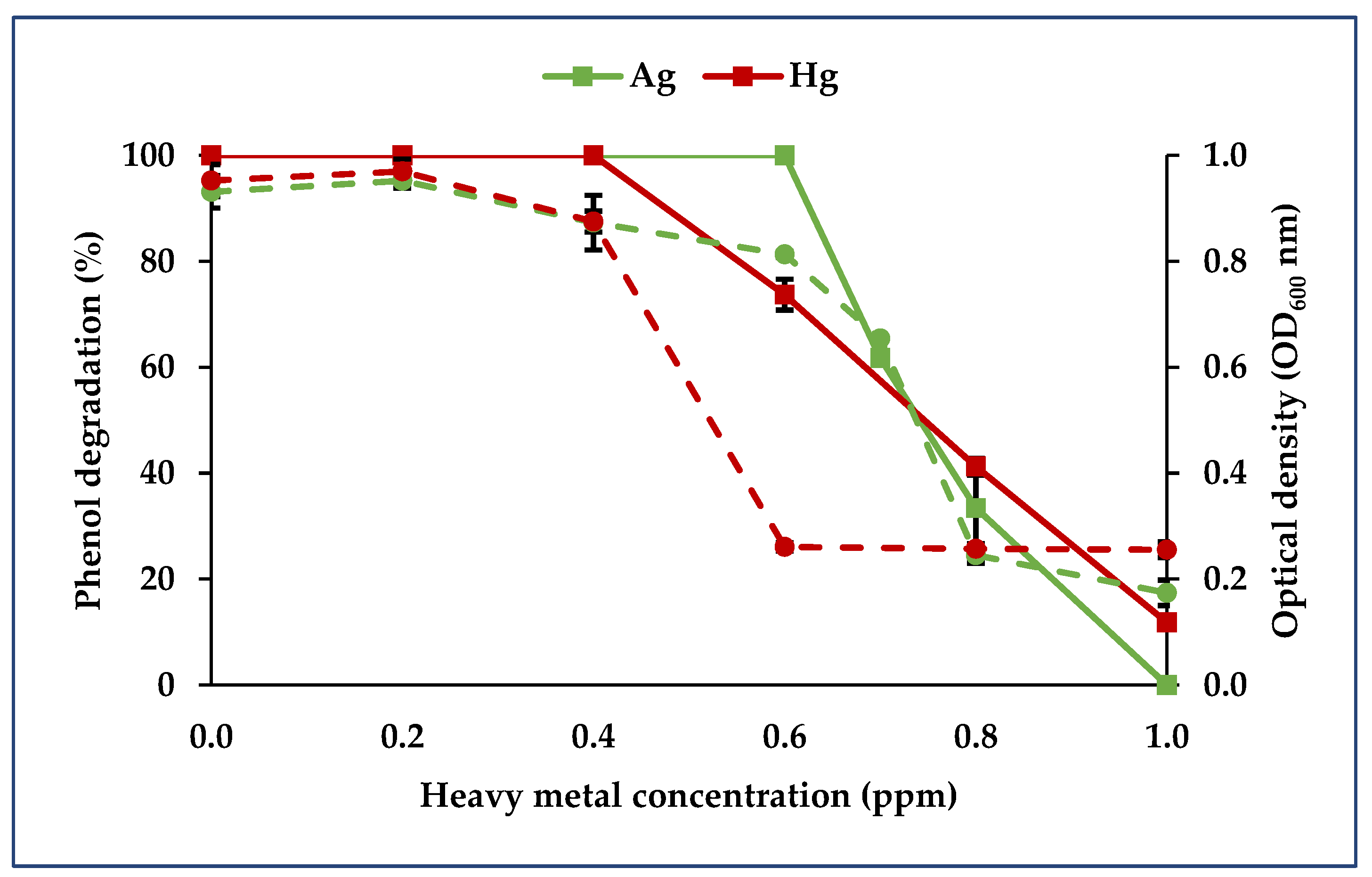
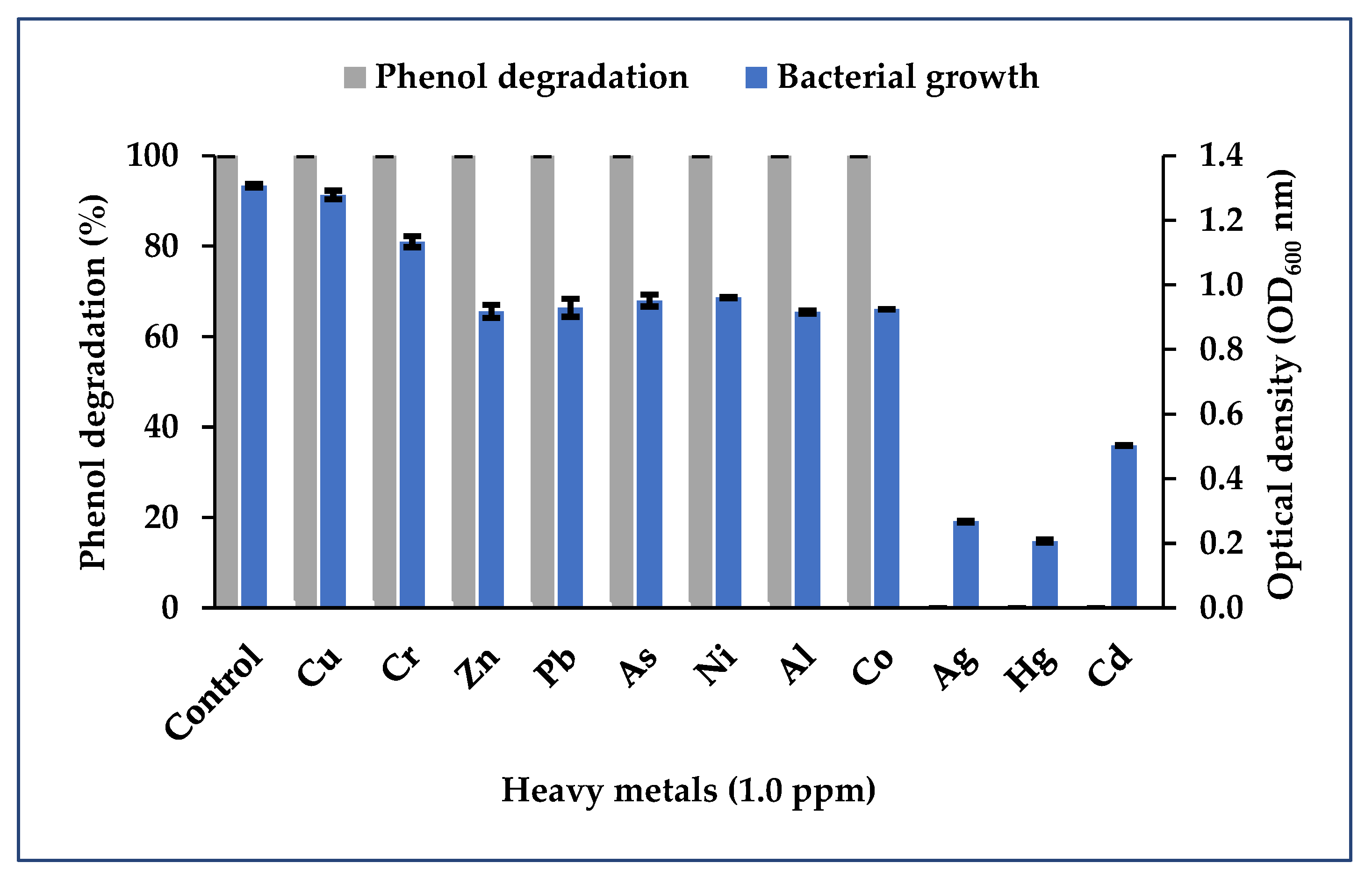
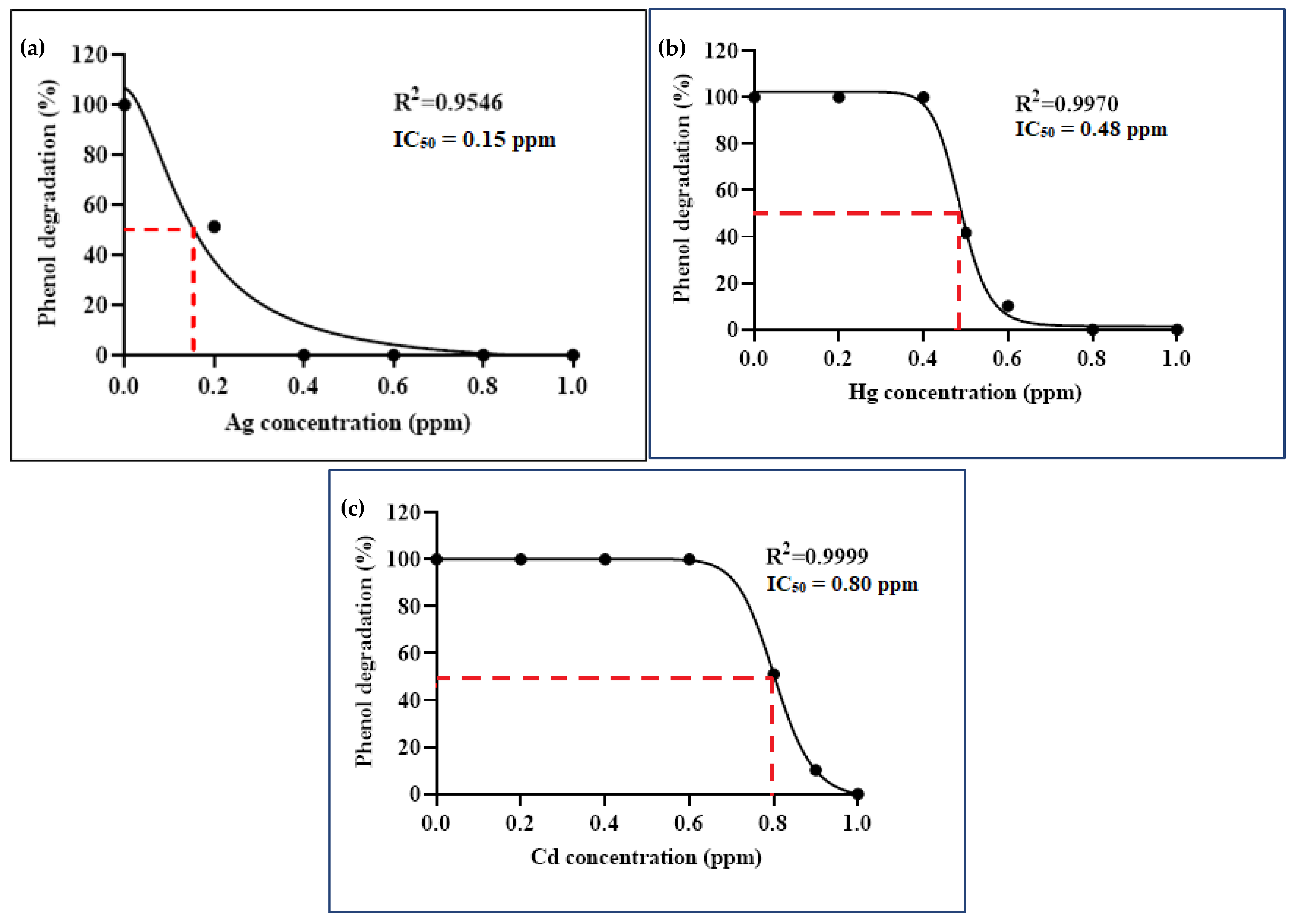
3.3. Impact of Heavy Metals on Phenol Biodegradation
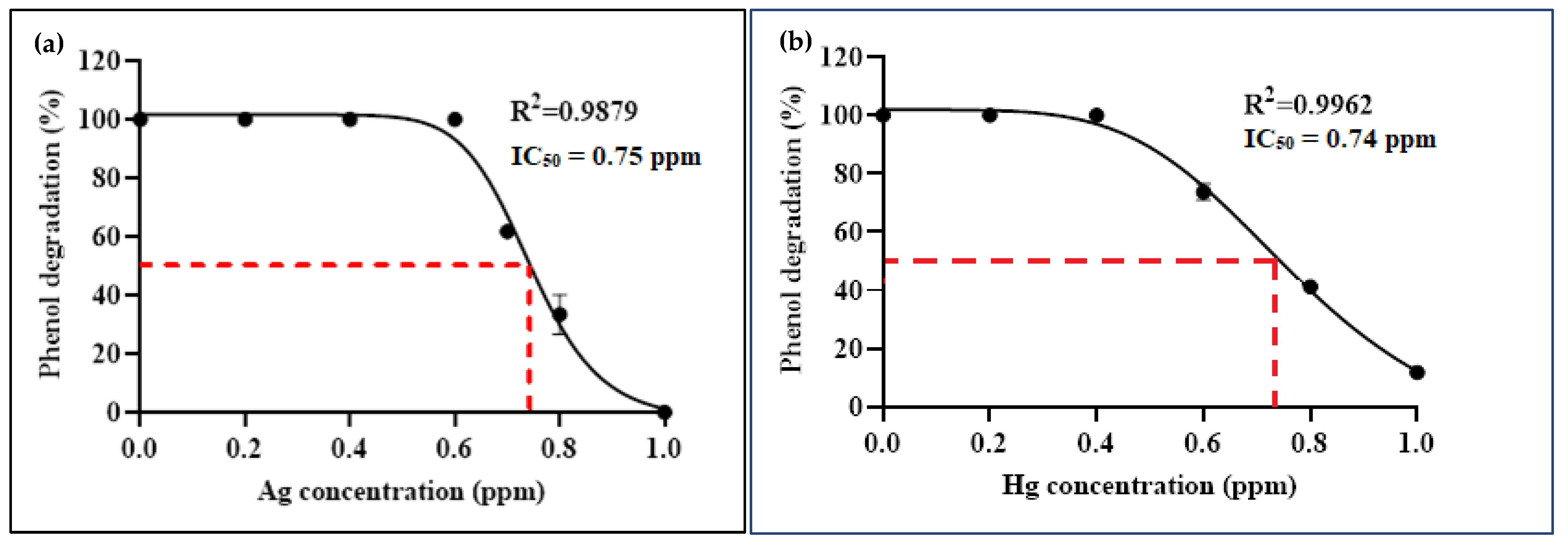
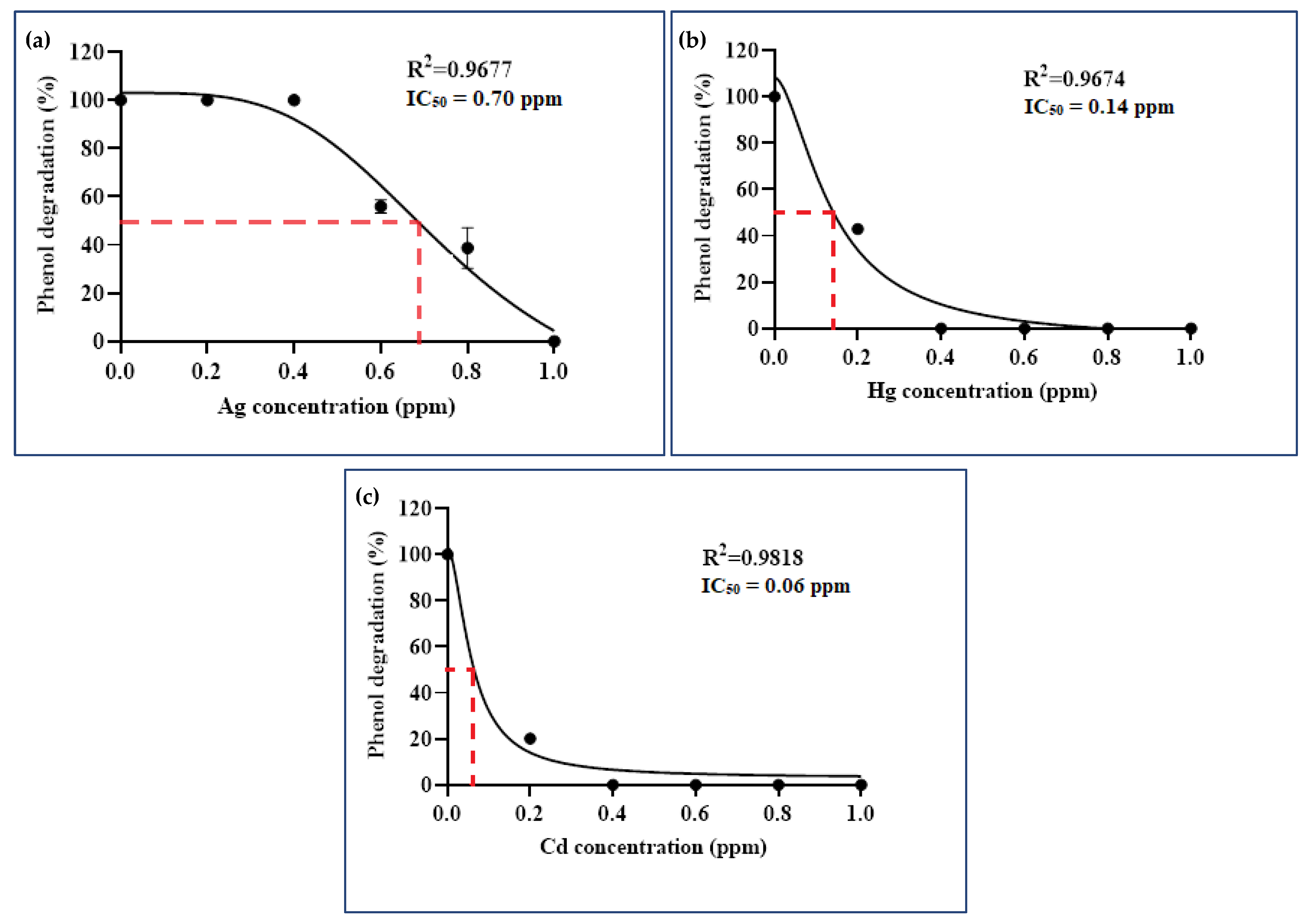
3.3.1. Arthrobacter sp. Strain AQ5-06
3.3.2. Arthrobacter sp. Strain AQ5-15
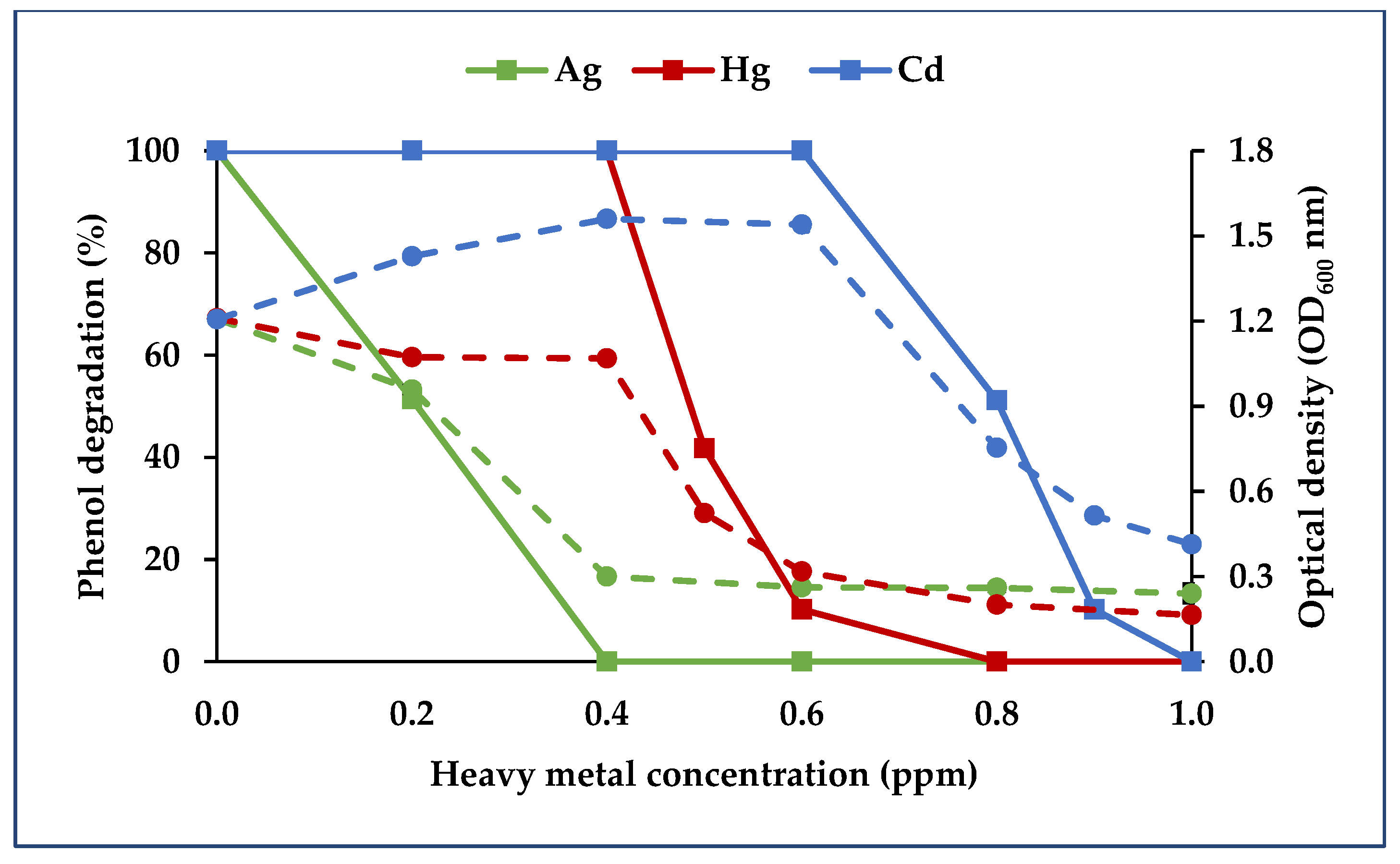
3.3.3. Binary Consortium
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Al-Chalabi, M. The final frontier? An expedition to Antarctica and implications for the future. Crit. Arts 2017, 31, 110–118. [Google Scholar] [CrossRef]
- Brown, K.E.; King, C.K.; Harrison, P.L. Lethal and behavioral impacts of diesel and fuel oil on the Antarctic amphipod Paramoera walkeri. Environ. Toxicol. Chem. 2017, 36, 2444–2455. [Google Scholar] [CrossRef]
- Bargagli, R. Antarctic Ecosystems: Environmental Contamination, Climate Change, and Human Impact; Springer Science and Business Media: Berlin, Germany, 2005; Volume 175, pp. 125–290. [Google Scholar]
- Tin, T.; Fleming, Z.L.; Hughes, K.A.; Ainley, D.G.; Convey, P.; Moreno, C.A.; Pfeiffer, S.; Scott, J.; Snape, I. Impacts of local human activities on the Antarctic environment. Antarct. Sci. 2009, 21, 3–33. [Google Scholar] [CrossRef] [Green Version]
- Hughes, K.A. How committed are we to monitoring human impacts in Antarctica? Environ. Res. Lett. 2010, 5, 041001. [Google Scholar] [CrossRef]
- Kennicutt, M.C., II; Klein, A.; Montagna, P.; Sweet, S.; Wade, T.; Palmer, T.; Sericano, J.; Denoux, G. Temporal and spatial patterns of anthropogenic disturbance at McMurdo Station, Antarctica. Environ. Res. Lett. 2010, 5, 034010. [Google Scholar] [CrossRef]
- Stark, J.S.; Kim, S.L.; Oliver, J.S. Anthropogenic disturbance and biodiversity of marine benthic communities in Antarctica: A regional comparison. PLoS ONE 2014, 9, e98802. [Google Scholar] [CrossRef]
- Stark, J.S.; Mohammad, M.; McMinn, A.; Ingels, J. The effects of hydrocarbons on meiofauna in marine sediments in Antarctica. J. Exp. Mar. Biol. Ecol. 2017, 496, 56–73. [Google Scholar] [CrossRef]
- Vázquez, S.; Monien, P.; Minetti, R.P.; Jürgens, J.; Curtosi, A.; Primitz, J.V.; Frickenhaus, S.; Abele, D.; Mac Cormack, W.; Helmke, E. Bacterial communities and chemical parameters in soils and coastal sediments in response to diesel spills at Carlini Station, Antarctica. Sci. Total Environ. 2017, 605, 26–37. [Google Scholar] [CrossRef]
- Pi, Y.; Meng, L.; Bao, M.; Sun, P.; Lu, J. Degradation of crude oil and relationship with bacteria and enzymatic activities in laboratory testing. Int. Biodeterior. Biodegrad. 2016, 106, 106–116. [Google Scholar] [CrossRef]
- McWatters, R.S.; Wilkins, D.; Spedding, T.; Hince, G.; Raymond, B.; Lagerewskij, G.; Terry, D.; Wise, L.; Snape, I. On site remediation of a fuel spill and soil reuse in Antarctica. Sci. Total Environ. 2016, 571, 963–973. [Google Scholar] [CrossRef] [PubMed]
- Hui-Peng, L.; Jian, S.; Hua, Z.; Li, Z.; Yu-Tai, Q. Effects of phenols on the stability of FCC diesel fuel. Pet. Sci. Technol. 2009, 27, 486–497. [Google Scholar] [CrossRef]
- Singh, A.; Kumar, V.; Srivastava, J.N. Assessment of bioremediation of oil and phenol contents in refinery waste water via bacterial consortium. J. Pet. Environ. Biotechnol. 2013, 4, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Allinson, M.; Kadokami, K.; Shiraishi, F.; Nakajima, D.; Zhang, J.; Knight, A.; Gray, S.R.; Scales, P.J.; Allinson, G. Wastewater recycling in Antarctica: Performance assessment of an advanced water treatment plant in removing trace organic chemicals. J. Environ. Manag. 2018, 224, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Kwon, K.D.; Jo, W.K.; Lim, H.J.; Jeong, W.S. Volatile pollutants emitted from selected liquid household products. Environ. Sci. Pollut. Res. 2008, 15, 521–526. [Google Scholar] [CrossRef]
- Vanden Eynde, J.J. How efficient is my (medicinal) chemistry? Pharmaceuticals 2016, 9, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vecchiato, M.; Argiriadis, E.; Zambon, S.; Barbante, C.; Toscano, G.; Gambaro, A.; Piazza, R. Persistent Organic Pollutants (POPs) in Antarctica: Occurrence in continental and coastal surface snow. Microchem. J. 2015, 119, 75–82. [Google Scholar] [CrossRef]
- Zangrando, R.; Barbaro, E.; Vecchiato, M.; Kehrwald, N.M.; Barbante, C.; Gambaro, A. Levoglucosan and phenols in Antarctic marine, coastal and plateau aerosols. Sci. Total Environ. 2016, 544, 606–616. [Google Scholar] [CrossRef] [PubMed]
- Zangrando, R.; Corami, F.; Barbaro, E.; Grosso, A.; Barbante, C.; Turetta, C.; Capodaglio, G.; Gambaro, A. Free phenolic compounds in waters of the Ross Sea. Sci. Total Environ. 2019, 650, 2117–2128. [Google Scholar] [CrossRef]
- Mohammadi, S.; Kargari, A.; Sanaeepur, H.; Abbassian, K.; Najafi, A.; Mofarrah, E. Phenol removal from industrial wastewaters: A short review. Desalin. Water Treat. 2014, 53, 2215–2234. [Google Scholar] [CrossRef]
- Tengku-Mazuki, T.A.; Subramanian, K.; Zakaria, N.N.; Convey, P.; Khalil, K.A.; Lee, G.L.Y.; Zulkharnain, A.; Shaharuddin, N.A.; Ahmad, S.A. Optimization of phenol degradation by Antarctic bacterium Rhodococcus sp. Antarct. Sci. 2020, 32, 486–495. [Google Scholar] [CrossRef]
- Camenzuli, D.; Freidman, B.L. On-site and in situ remediation technologies applicable to petroleum hydrocarbon contaminated sites in the Antarctic and Arctic. Polar Res. 2015, 34, 24492. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, Y.; Sawada, T. Biodegradation of phenol in the presence of heavy metals. J. Chem. Technol. Biotechnol. 2000, 75, 137–142. [Google Scholar] [CrossRef]
- Zakaria, N.N.; Ahmad, S.A.; Yin, G.L.L.; Yasid, N.A.; Motharasan, M.; Subramaniam, K.; Mazuki, T.A.T.; Nawawi, N.M.; Shukor, M.Y. Biodegradation of phenol by Antarctic bacterium Rhodococcus baikonurensis strain AQ5-001 in the presence of heavy metals. Malaysian J. Biochem. Mol. Biol. 2018, 21, 29–36. [Google Scholar]
- Tengku-Mazuki, T.A.; Zulkharnain, A.; Subramaniam, K.; Convey, P.; Gomez-Fuentes, C.; Ahmad, S.A. Effects of zinc (Zn) and chromium (Cr) on the phenol-degrading bacteria growth kinetics. Malays. J. Biochem. Mol. Biol. 2020, 23, 1–4. [Google Scholar]
- Duraisamy, P.; Sekar, J.; Arunkumar, A.D.; Ramalingam, P.V. Kinetics of phenol biodegradation by heavy metal tolerant rhizobacteria Glutamicibacter nicotianae MSSRFPD35 from distillery effluent contaminated soils. Front. Microbiol. 2020, 11, 1573. [Google Scholar] [CrossRef]
- Stark, J.S.; Bridgen, P.; Dunshea, G.; Galton-Fenzi, B.; Hunter, J.; Johnstone, G.; King, C.; Leeming, R.; Palmer, A.; Smith, J.; et al. Dispersal and dilution of wastewater from an ocean outfall at Davis Station, Antarctica, and resulting environmental contamination. Chemosphere 2016, 152, 142–157. [Google Scholar] [CrossRef] [PubMed]
- Chu, W.L.; Dang, N.L.; Kok, Y.Y.; Yap, K.S.I.; Phang, S.M.; Convey, P. Heavy metal pollution in Antarctica and its potential impacts on algae. Polar Sci. 2019, 20, 75–83. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, J.; Zhang, Y.; Hesham, A.L.; Yang, M. Molecular characterization of a bacterial consortium enriched from an oilfield that degrades phenanthrene. Biotechnol. Lett. 2006, 28, 617–621. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharya, M.; Guchhait, S.; Biswas, D.; Datta, S. Waste lubricating oil removal in a batch reactor by mixed bacterial consortium: A kinetic study. Bioprocess Biosyst. Eng. 2015, 38, 2095–2106. [Google Scholar] [CrossRef] [PubMed]
- Sivasubramanian, S.; Namasivayam, S.K.R. Phenol degradation studies using microbial consortium isolated from environmental sources. J. Environ. Chem. Eng. 2015, 3, 243–252. [Google Scholar] [CrossRef]
- Roslee, A.F.A.; Zakaria, N.N.; Convey, P.; Zulkharnain, A.; Lee, G.L.Y.; Gomez-Fuentes, C.; Ahmad, S.A. Statistical optimisation of growth conditions and diesel degradation by the Antarctic bacterium, Rhodococcus sp. strain AQ5–07. Extremophiles 2020, 24, 277–291. [Google Scholar] [CrossRef]
- Abdulrasheed, M.; Zulkharnain, A.; Zakaria, N.N.; Roslee, A.F.A.; Abdul Khalil, K.; Napis, S.; Convey, P.; Gomez-Fuentes, C.; Ahmad, S.A. Response surface methodology optimization and kinetics of diesel degradation by a cold-adapted Antarctic bacterium, Arthrobacter sp. strain AQ5-05. Sustainability 2020, 12, 6966. [Google Scholar] [CrossRef]
- Ungureanu, C.V.; Favier, L.; Bahrim, G.E. Improving Biodegradation of Clofibric Acid by Trametes pubescens through the Design of Experimental Tools. Microorganisms 2020, 8, 1243. [Google Scholar] [CrossRef]
- Lee, G.L.Y.; Ahmad, S.A.; Yasid, N.A.; Zulkharnain, A.; Convey, P.; Johari, W.L.W.; Alias, S.A.; Gonzalez-Rocha, G.; Shukor, M.Y. Biodegradation of phenol by cold-adapted bacteria from Antarctic soils. Polar Biol. 2018, 41, 553–562. [Google Scholar] [CrossRef]
- Subramaniam, K.; Mazuki, T.A.T.; Shukor, M.Y.; Ahmad, S.A. Isolation and optimisation of phenol degradation by Antarctic isolate using one factor at time. Malays. J. Biochem. Mol. Biol. 2019, 22, 79–86. [Google Scholar]
- Abdulrasheed, M.; Zakaria, N.N.; Roslee, A.F.A.; Shukor, M.Y.; Zulkharnain, A.; Napis, S.; Convey, P.; Alias, S.A.; Gonzalez-Rocha, G.; Ahmad, S.A. Biodegradation of diesel oil by cold-adapted bacterial strains of Arthrobacter spp. from Antarctica. Antarct. Sci. 2020, 32, 341–353. [Google Scholar] [CrossRef]
- APHA (American Public Health Association). Standard Method for the Examination of Water and Wastewater, 20th ed.; American Public Health Association: Washington, DC, USA, 2005; pp. 540–544. [Google Scholar]
- Suhaila, Y.N.; Ramanan, R.N.; Rosfarizan, M.; Latif, I.A.; Ariff, A.B. Optimization of parameters for improvement of phenol degradation by Rhodococcus UKMP-5M using response surface methodology. Ann. Microbiol. 2013, 63, 513–521. [Google Scholar] [CrossRef]
- Ahmad, S.A.; Asokan, G.; Yasid, N.A.; Nawawi, N.M.; Subramaniam, K.; Zakaria, N.N.; Shukor, M.Y. Effect of heavy metals on biodegradation of phenol by Antarctic bacterium: Arthrobacter bambusae strain AQ5-003. Malaysian J. Biochem. Mol. Biol. 2018, 21, 47–51. [Google Scholar]
- Carlson, H.K.; Price, M.N.; Callaghan, M.; Aaring, A.; Chakraborty, R.; Liu, H.; Kuehl, J.V.; Arkin, A.P.; Deutschbauer, A.M. The selective pressures on the microbial community in a metal-contaminated aquifer. ISME J. 2019, 13, 937–949. [Google Scholar] [CrossRef] [Green Version]
- Tecon, R.; Or, D. Cooperation in carbon source degradation shapes spatial self-organization of microbial consortia on hydrated surfaces. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Lee, G.L.Y. Biodegradation of Phenol by Cold-Adapted Bacteria Isolated from Antarctic Soils. Ph.D. Thesis, Universiti Putra Malaysia, Selangor, Malaysia, 2018. [Google Scholar]
- Subramaniam, K.; Shaharuddin, N.A.; Tengku-Mazuki, T.A.; Zulkharnain, A.; Khalil, K.A.; Convey, P.; Ahmad, S.A. Statistical optimisation for enhancement of phenol biodegradation by the Antarctic soil bacterium Arthrobacter sp. strain AQ5-15 using response surface methodology. J. Environ. Biol. 2020, 41, 1560–1569. [Google Scholar] [CrossRef]
- Clarke, K.G. Bioprocess Engineering: An Introductory Engineering and Life Science Approach. In Microbiology; Woodhead Publishing Limited: Cambridge, UK, 2013; pp. 7–24. [Google Scholar]
- Varjani, S.J.; Upasani, V.N. A new look on factors affecting microbial degradation of petroleum hydrocarbon pollutants. Int. Biodeterior. Biodegrad. 2017, 120, 71–83. [Google Scholar] [CrossRef]
- Lin, C.; Gan, L.; Chen, Z.L. Biodegradation of naphthalene by strain Bacillus fusiformis (BFN). J. Hazard. Mater. 2010, 182, 771–777. [Google Scholar] [CrossRef] [PubMed]
- Ucun, H.; Yildiz, E.; Nuhoglu, A. Phenol biodegradation in a batch jet loop bioreactor (JLB): Kinetics study and pH variation. Bioresour. Technol. 2010, 101, 2965–2971. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.; Lu, H.; King, J.; Marshall, G.J.; Phillips, T.; Bannister, D.; Colwell, S. Extreme temperatures in the Antarctic. J. Clim. 2021, 34, 2653–2668. [Google Scholar] [CrossRef]
- Tay, J.H.; Jiang, H.L.; Tay, S.T.L. High-rate biodegradation of phenol by aerobically grown microbial granules. Int. J. Environ. Eng. 2004, 130, 1415–1423. [Google Scholar] [CrossRef]
- Tay, S.T.L.; Moy, B.Y.P.; Maszenan, A.M.; Tay, J.H. Comparing activated sludge and aerobic granules as microbial inocula for phenol biodegradation. Appl. Microbiol. Biotechnol. 2005, 67, 708–713. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.; Li, C.; Song, X.; Yang, Y. Biodegradation of phenol by Rhodococcus sp. strain SKC: Characterization and kinetics study. Molecules 2020, 25, 3665. [Google Scholar] [CrossRef] [PubMed]
- Senthilvelan, T.; Kanagaraj, J.; Panda, R.C.; Mandal, A.B. Biodegradation of phenol by mixed microbial culture: An eco-friendly approach for the pollution reduction. Clean Technol. Environ. Policy 2014, 16, 113–126. [Google Scholar] [CrossRef]
- Hamitouche, A.; Amrane, A.; Bendjama, Z.; Kaouah, F. Effect of the ammonium chloride concentration on the mineral medium composition—Biodegradation of phenol by a microbial consortium. Int. J. Environ. Res. 2010, 4, 849–854. [Google Scholar]
- Bhatia, S.K.; Bhatia, R.K.; Choi, Y.K.; Kan, E.; Kim, Y.G.; Yang, Y.H. Biotechnological potential of microbial consortia and future perspectives. Crit. Rev. Biotechnol. 2018, 38, 1209–1229. [Google Scholar] [CrossRef] [PubMed]
- Deeb, B.E.; Altalhi, A.D. Degradative plasmid and heavy metal resistance plasmid naturally coexist in phenol and cyanide assimilating bacteria. Am. J. Biochem. Biotechnol. 2009, 5, 84–93. [Google Scholar] [CrossRef]
- Al-Defiery, M.E.; Reddy, G. Influence of metal ions concentration on phenol degradation by Rhodococcus pyridinivorans GM3. Mesop. Environ. J. 2014, 1, 30–38. [Google Scholar]
- Trevizani, T.H.; Petti, M.A.V.; Ribeiro, A.P.; Corbisier, T.N.; Figueira, R.C.L. Heavy metal concentrations in the benthic trophic web of martel inlet, admiralty Bay (King George Island, Antarctica). Mar. Pollut. Bull. 2018, 130, 198–205. [Google Scholar] [CrossRef] [PubMed]
- Chu, Z.; Yang, Z.; Wang, Y.; Sun, L.; Yang, W.; Yang, L.; Gao, Y. Assessment of heavy metal contamination from penguins and anthropogenic activities on Fildes peninsula and Ardley island, Antarctic. Sci. Total Environ. 2019, 646, 951–957. [Google Scholar] [CrossRef] [PubMed]
- Lischka, A.; Bustamante, P.; Braid, H.; Piatkowski, U.; Lacoue-Labarthe, T. Trophic ecology drives trace element concentrations in the Antarctic octopod community. Sci. Total Environ. 2021, 768, 144373. [Google Scholar] [CrossRef] [PubMed]
- Abdulrasheed, M.; Roslee, A.F.; Zakaria, N.N.; Zulkharnain, A.; Lee, G.L.Y.; Convey, P.; Napis, S.; Ahmad, S.A. Effects of heavy metals on diesel metabolism of psychrotolerant strains of Arthrobacter sp. from Antarctica. J. Environ. Biol. 2020, 41, 966–972. [Google Scholar] [CrossRef]
- Ibrahim, S.; Zulkharnain, A.; Zahri, K.N.M.; Lee, G.L.Y.; Convey, P.; Gomez Fuentes, C.; Sabri, S.; Khalil, K.A.; Alias, S.A.; Gonzales-Rocha, G.; et al. Effect of heavy metals and other xenobiotics on biodegradation of waste canola oil by cold-adapted Rhodococcus sp. AQ5-07. Rev. Mex. Ing. Quim. 2020, 19, 1041–1052. [Google Scholar] [CrossRef] [Green Version]
- Zahri, K.N.M.; Zulkharnain, A.; Gomez-Fuentes, C.; Sabri, S.; Ahmad, S.A. Effects of heavy metals on Antarctic bacterial cell growth kinetics and degradation of waste canola oil. J. Environ. Biol. 2020, 41, 1433–1441. [Google Scholar] [CrossRef]
- Zakaria, N.N.; Roslee, A.F.A.; Gomez-Fuentes, C.; Zulkharnain, A.; Abdulrasheed, M.; Sabri, S.; Ramirez-Moreno, N.; Calisto-Ulloa, N.; Ahmad, S.A. Kinetic studies of marine psychrotolerant microorganisms capable of degrading diesel in the presence of heavy metals. Rev. Mex. Ing. Quim. 2020, 19, 1375–1388. [Google Scholar] [CrossRef]
- Ladomersky, E.; Petris, M.J. Copper tolerance and virulence in bacteria. Metallomics 2015, 7, 957–964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Issazadeh, K.; Jahanpour, N.; Pourghorbanali, F.; Raeisi, G.; Faekhondeh, J. Heavy metals resistance by bacterial strains. Ann. Biol. Res. 2013, 4, 60–63. [Google Scholar]
- Singh, R.; Gautam, N.; Mishra, A.; Gupta, R. Heavy metals and living systems: An overview. Indian J. Pharmacol. 2011, 43, 246. [Google Scholar] [CrossRef] [Green Version]
- Yoo, J.Y.; Choi, J.; Lee, T.; Park, J.W. Organobentonite for sorption and degradation of phenol in the presence of heavy metals. Water Air Soil Pollut. 2004, 154, 225–237. [Google Scholar] [CrossRef]
- Hrynkiewicz, K.; Baum, C. Application of Microorganisms in Bioremediation of Environment from Heavy Metals. In Environmental Deterioration and Human Health; Malik, A., Grohmann, E., Akhtar, R., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 215–227. [Google Scholar]
- Bruins, M.R.; Kapil, S.; Oehme, F.W. Microbial resistance to metals in the environment. Ecotoxicol. Environ. Saf. 2000, 45, 198–207. [Google Scholar] [CrossRef] [PubMed]
- Schink, S.J.; Biselli, E.; Ammar, C.; Gerland, U. Death rate of E. coli during starvation is set by maintenance cost and biomass recycling. Cell Syst. 2019, 9, 64–73. [Google Scholar] [CrossRef]
- Al-Saleh, E.S.; Obuekwe, C. Inhibition of hydrocarbon bioremediation by lead in a crude oil-contaminated soil. Int. Biodeterior. Biodegrad. 2005, 56, 1–7. [Google Scholar] [CrossRef]
- Lu, Q.; Zou, X.; Liu, J.; Liang, Z.; Shim, H.; Qiu, R.; Wang, S. Inhibitory effects of metal ions on reductive dechlorination of polychlorinated biphenyls and perchloroethene in distinct organohalide-respiring bacteria. Environ. Int. 2020, 135, 105373. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.H.; Kwon, Y.J.; So, J.S. Bioremediation of heavy metals by using bacterial mixtures. Ecol. Eng. 2016, 89, 64–69. [Google Scholar] [CrossRef]
- Oaikhena, E.E.; Makaije, D.B.; Denwe, S.D.; Namadi, M.M.; Haroun, A.A. Bioremediation potentials of heavy metal tolerant bacteria isolated from petroleum refinery effluent. Am. J. Environ. Prot. 2016, 5, 29–34. [Google Scholar] [CrossRef] [Green Version]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Symbol | Experimental Values | ||||
|---|---|---|---|---|---|---|
| −2 | −1 | 0 | +1 | +2 | ||
| (NH4)2SO4 concentration (g/L) | A | 0.2 | 0.3 | 0.4 | 0.5 | 0.6 |
| NaCl concentration (g/L) | B | 0.08 | 0.10 | 0.13 | 0.15 | 0.17 |
| pH | C | 6.75 | 7.00 | 7.25 | 7.50 | 7.75 |
| Temperature (°C) | D | 7.0 | 10.0 | 12.5 | 15.0 | 17.50 |
| DMC | Inoculum Ratio | Growth Rate (h−1) | Incubation Time (h) |
|---|---|---|---|
| AQ5-06 + AQ5-15 | 1:1 | 0.0191 (±0.0263) | 48 |
| 1:3 | 0.0106 (±0.0017) | 84 | |
| 3:1 | 0.0121 (±0.0227) | 84 |
| Source | Sum of Squares | DF | Mean Square | F-Value | Prob > F |
|---|---|---|---|---|---|
| Model | 6879.31 | 8 | 859.91 | 37.51 | <0.0001 *** |
| A | 0.01 | 1 | 0.012 | 5.3 × 10−3 | 0.9819 |
| B | 51.92 | 1 | 51.92 | 2.26 | 0.1472 |
| C | 38.35 | 1 | 38.35 | 1.67 | 0.2099 |
| D | 1581.13 | 1 | 1581.13 | 68.97 | <0.0001 *** |
| A2 | 544.17 | 1 | 544.17 | 23.74 | <0.0001 *** |
| B2 | 598.61 | 1 | 598.61 | 26.11 | <0.0001 *** |
| C2 | 577.34 | 1 | 577.34 | 25.18 | <0.0001 *** |
| D2 | 4718.10 | 1 | 4718.10 | 205.80 | <0.0001 *** |
| Residual | 481.43 | 21 | 22.93 | ||
| Lack of Fit | 378.55 | 16 | 23.66 | 1.15 | 0.4773 |
| Pure Error | 102.88 | 5 | 20.58 | ||
| Cor Total | 7360.74 | 29 | |||
| Standard deviation | 4.79 | R2 | 0.9346 | ||
| Mean | 73.31 | Adjusted R2 | 0.9097 | ||
| Coefficient variance | 6.53 | Predicted R2 | 0.7573 | ||
| PRESS | 1786.11 | Adequate Precision | 26.1944 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Subramaniam, K.; Ahmad, S.A.; Convey, P.; Shaharuddin, N.A.; Khalil, K.A.; Tengku-Mazuki, T.A.; Gomez-Fuentes, C.; Zulkharnain, A. Statistical Assessment of Phenol Biodegradation by a Metal-Tolerant Binary Consortium of Indigenous Antarctic Bacteria. Diversity 2021, 13, 643. https://doi.org/10.3390/d13120643
Subramaniam K, Ahmad SA, Convey P, Shaharuddin NA, Khalil KA, Tengku-Mazuki TA, Gomez-Fuentes C, Zulkharnain A. Statistical Assessment of Phenol Biodegradation by a Metal-Tolerant Binary Consortium of Indigenous Antarctic Bacteria. Diversity. 2021; 13(12):643. https://doi.org/10.3390/d13120643
Chicago/Turabian StyleSubramaniam, Kavilasni, Siti Aqlima Ahmad, Peter Convey, Noor Azmi Shaharuddin, Khalilah Abdul Khalil, Tengku Athirrah Tengku-Mazuki, Claudio Gomez-Fuentes, and Azham Zulkharnain. 2021. "Statistical Assessment of Phenol Biodegradation by a Metal-Tolerant Binary Consortium of Indigenous Antarctic Bacteria" Diversity 13, no. 12: 643. https://doi.org/10.3390/d13120643