
Developing Inside a Layer of Germs—A Potential Role for Multiciliated Surface Cells in Vertebrate Embryos


Department of Biology and Biochemistry & Molecular Biology, Gettysburg College, Gettysburg, PA 17325, USA
Diversity 2021, 13(11), 527; https://doi.org/10.3390/d13110527
Submission received: 26 July 2021
/
Revised: 18 October 2021
/
Accepted: 20 October 2021
/
Published: 23 October 2021
(This article belongs to the Special Issue Evolution, Development, and Diversification of Vertebrates)
{kind=link}
{kind=link}
{kind=link}
Abstract
:This paper reviews current research on the microbial life that surrounds vertebrate embryos. Several clades are believed to develop inside sterile—or near-sterile—embryonic microhabitats, while others thrive within a veritable zoo of microbial life. The occurrence of embryo-associated microbes in some groups, but not others, is an under-appreciated transition (possibly transitions) in vertebrate evolution. A lack of comparable studies makes it currently impossible to correlate embryo-associated microbiomes with other aspects of vertebrate evolution. However, there are embryonic features that should instruct a more targeted survey. This paper concludes with a hypothesis for the role of multiciliated surface cells in amphibian and some fish embryos, which may contribute to managing embryo-associated microbial consortia. These cells are known to exist in some species that harbor in ovo microbes or have relatively porous egg capsules, although most have not been assayed for embryo-associated microbiota. Whether the currents generated within these extraembryonic microhabitats contribute to culturing consistent microbial communities remains to be seen.
1. Introduction
Since the beginning of the 21st century, the life sciences have experienced a revolution in our understanding of microbial interactions with multicellular life. There is a growing appreciation for the influence of single-celled bacteria, fungi, protists, and viruses on many nutritional, physiological, and behavioral host phenotypes. The revelation that we are “animals in a bacterial world” [1,2] has largely been driven by new culture-free microbial identification techniques. These include rDNA metabarcoding [3] as well as the sequencing of entire microbial communities, or metagenomics [4]. However, parallel lines of research have identified organisms that do not require a (primarily gut) microbiome [5]. These apparent outliers indicate potential constraints on otherwise common host-microbiota interactions. This review proposes that comparative studies of microbial interactions with vertebrate embryos offer similar opportunities to interrogate these rules of engagement. Despite a large amount of research on the controversial human in utero microbiota, we know surprisingly little about which vertebrate embryos maintain an embryo-associated microbial communities, and less still about the developmental and physiological consequences of these consortia [6]. By reviewing the evidence at hand, this paper aims to identify gaps in our understanding in order to prioritize future efforts in vertebrate embryo-microbe research.
2. Terminology: In Ovo and In Utero
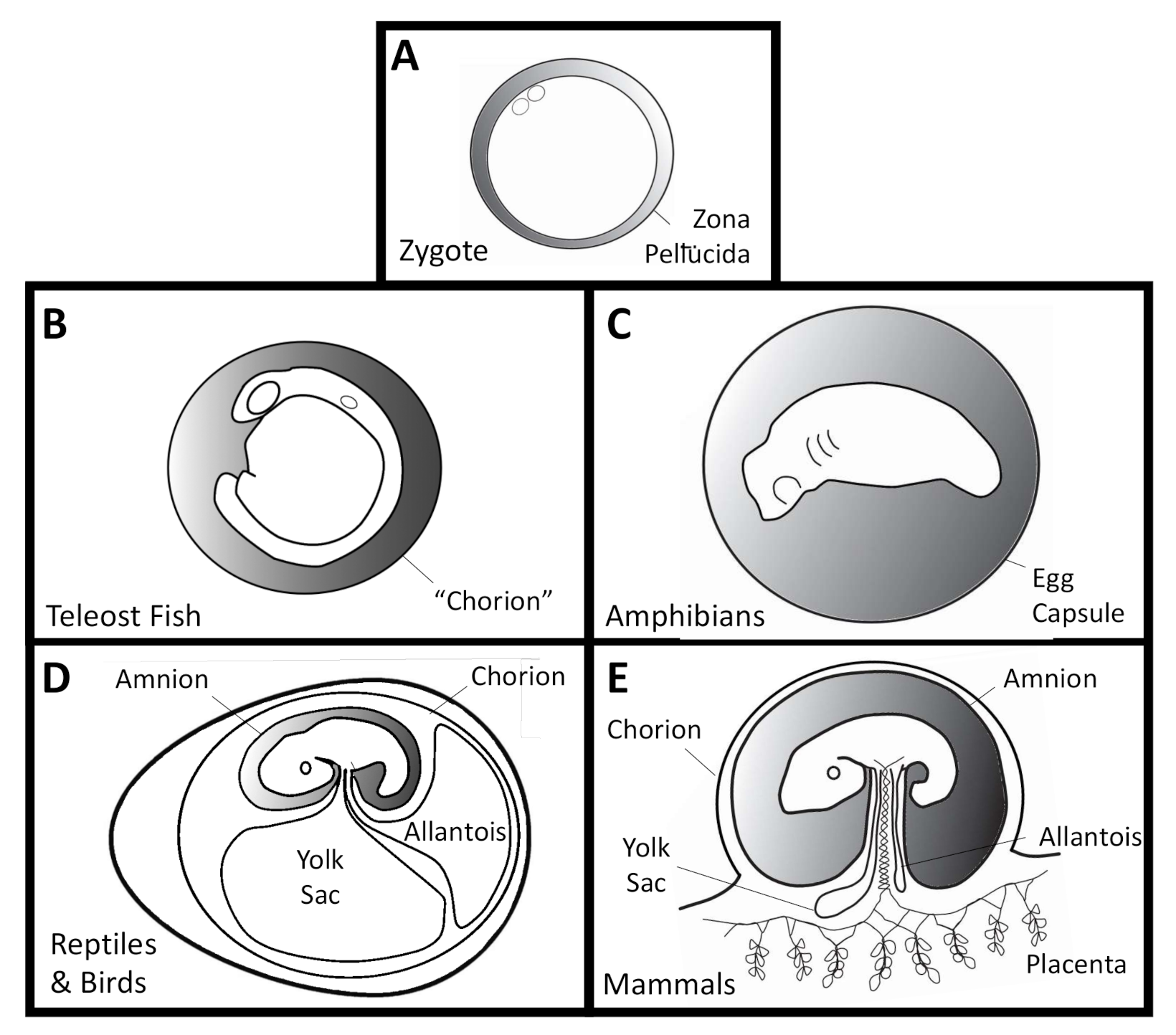
All vertebrates begin life inside a clear capsule called the oocyte coat, zona pellucida, zona radiata, vitelline membrane, vitelline envelope, or perivitelline capsule [7,8] (Figure 1A). These express zona pellucida (ZP) glycoproteins, which are involved in sperm recognition, the acrosome reaction, and associated blocks to polyspermy [9]. This innermost capsule is maternally derived and surrounds the unfertilized egg as well as the early embryo. Humans “hatch” from their zona pellucida prior to uterine implantation, and are often prematurely removed from these capsules during in vitro fertilization procedures with frozen-thawed blastocyts [10]. In fish and amphibians, the innermost vitelline/perivitelline capsule will rupture later in development, during organogenesis [11,12]. The vitelline membrane of bird eggs degrades seven days after laying [13], after which they become more susceptible to pathogens [14]. Many fish [12,15], amphibian [11], and most human embryos [7] form transient “threads” or “filaments” inside the perivitelline space. These are of largely unknown composition and function and vary in their attachments between the embryo and surrounding membrane. To date, there have been no accounts of microbial life within the perivitelline fluid. This would be a pre-requisite for transovarial transmission of microbes in vertebrates, which has also never been found.
Additional oocyte capsules of varying numbers [8] surround the innermost vitelline envelope in fish [12] and amphibians [11,16] (Figure 1B,C). The outermost capsule in fish is called the “chorion”, which is not homologous with the embryo-derived amniote chorion [17]. These in turn are surrounded by maternally-derived jelly coat(s), which can cluster eggs together. Several oviparous taxa also form post-fertilization capsules in nidamental or shell glands, such as the skate egg case or reptile and bird eggs [8].
The external aquatic development of most (but not all) fish and amphibian embryos contrasts the more terrestrial life histories enabled by the amniotic egg and its associated membranes. The most prominent of those membranes is the amnion, which is the “pond” where reptile and mammalian embryos develop. The amnion is also derived from the embryo and is not homologous with any fish or amphibian structures. Additional amniote innovations include the allantois for metabolic waste collection and chorion for gas exchange (Figure 1D,E).
The primary concern of this review is to compare the occurrence and consequences of microbes on developing vertebrate embryos. This may be either inside the embryo proper, or inside the surrounding fluid and extraembryonic membranes of oviparous and ovoviviparous species (an in ovo microbiota), or within the amniotic fluid and accompanying membranes, including the embryonic placenta, in viviparous mammals (an in utero microbiota). Variation abounds in the timing of internal “hatching” from concentric capsules, the formation and degradation of the amniote zona pellucida, alternative life history strategies in various groups, or the extent of development that occurs within these protected environments (as in differences between marsupial and placental mammals). While the “pond” of amniotic fluid is not homologous to the capsular fluid of oviparous an-amniotes, all of these embryos develop in controlled aqueous microenvironments that have the potential to harbor microbial pathogens, and potentially commensal or mutualistic microbes as well. By focusing on in ovo and in utero microbes, this review explores the evidence for embryo-associated symbionts within the extraembryonic fluids, as opposed to embryo-adjacent microbes that may grow on the outer surfaces of the rearing environment or within the mother.
3. Methods for Identifying Microbial Consortia
Multiple techniques are available to identify and characterize microbial populations. However, none are available to definitively establish the sterility of a surface or structure. As early as 1885, Thomas Escherich used culturing to claim the neonate intestine is sterile [18]. This absence of evidence has since been called into question. Traditional culture-based methods rely on mimicking the endogenous niche of target microbe(s), which may be difficult to characterize [19]. Novel developments in staged bioreactors allow the isolation and clonal culturing of a wider range of bacteria than conventional techniques [20]; however, molecular techniques have become the current standard for identifying rare environmental microbes [21].
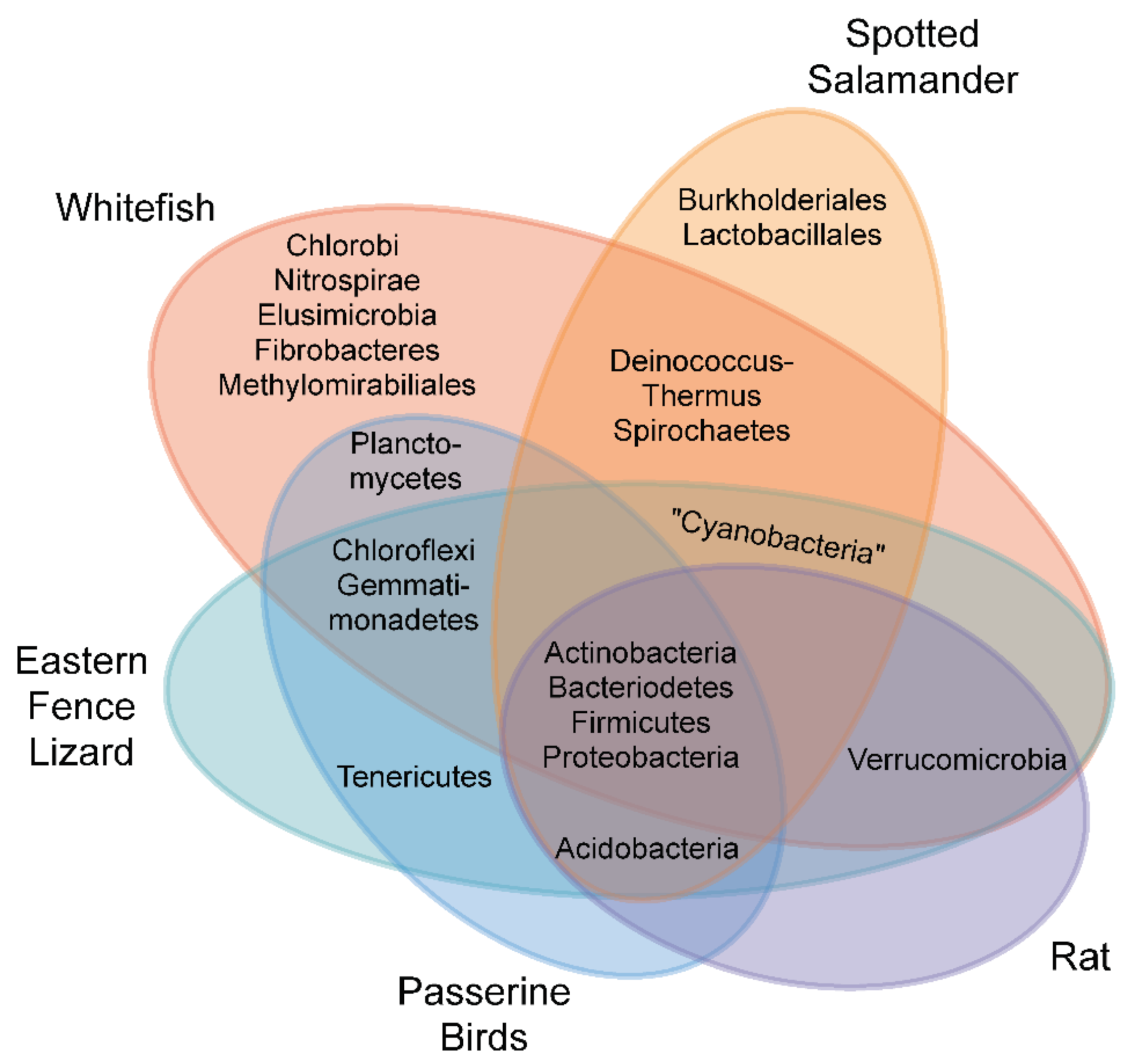
Molecular approaches typically rely on the use of conserved sequence “barcodes” for different microbial clades. These include tissue techniques such as fluorescent in situ hybridizations (FISH [22]), or newer hybridization chain reaction (HCR-FISH [23]), to detect clade-specific sequences with fluorescent probes. “Meta-barcoding” requires degenerate PCR primers or primers that target highly conserved regions of fungal (internal transcribed spacer region—ITS rDNA [24]), eukaryotic (18s rDNA [25]), bacterial (16s rDNA [26]), or green algal (rubisco [27]) genomes. The amplified PCR products are either analyzed for sequence heterogeneity through denaturing gradient gel electrophoresis [28], hybridized to a taxa-defined DNA microarray chip [29], or prepared as libraries for next generation sequencing and binned according to sequence similarity with curated reference taxonomy classifiers [30]. While these PCR amplification-dependent techniques are more sensitive than traditional culturing, they are also prone to false-positive signals from low-abundance microbial populations [31]. A summary of representative vertebrate embryo-associated bacterial communities, which have recently been reported using a range of 16s rDNA sequencing approaches, is shown in Figure 2 [32,33,34,35]. Variation in PCR primers, taxonomy classifiers, and negative control filtering makes this comparison preliminary at best. Future comparative analyses should include consistent sampling and bioinformatic approaches, including metagenomic sequencing, where all microbial genomes in a sample are sequenced, and both the identity and inferred functions of shared community members are revealed [21].
4. Evidence for In Ovo and In Utero Microbiotas in Healthy Vertebrates
4.1. Fish
With over thirty-two thousand species [39], a global distribution, and 38 reproductive guilds [40], it is difficult to generalize anything about the diversity of fish. Multiple studies treat the teleost extraembryonic fluid as sterile. However, this may not be the case for all fish. Kemp [41] describes pores in Australian lungfish chorion (Neoceratodus fosteri), which allow noticeable “particle”, and presumably microbial, entry.
The majority of fish egg-microbiota research attention has focused on reducing infections in aquaculture [42,43], and controlling the colonization of gut microbiome [44,45,46]. Studies of microbial colonization on the surface of teleost eggs have found biased bacterial populations that differ from the surrounding environment in Salmo trutta (brown trout [47,48]), Salmo salar L. (Atlantic salmon [49]), Gadus morhua and Hippoglossus hippoglossus (cod and halibut, respectively [50]). Recent studies on controlling G. morhua [36], and characterizing Danio rerio (zebrafish [51]) microbiota establishment explicitly state that the fluid inside the chorion is sterile. This may be the case for teleosts as their chorion ultrastructure has plugged canals in its outer layers, which create a semipermeable membrane that only allow small molecules to pass [52,53]. Additionally, unlike other vertebrates, sperm can only access unfertilized teleost eggs through a specialized micropyle within their oocyte capsule [12,54]. This restrictive access likely contributes to an effectively sterile in ovo environment for teleost embryos.
However, it should be noted that neither the zebrafish or aquaculture studies have assayed fish embryos in their native environments for in ovo colonization. A recent study by Wilkins et al. [32] on field-collected whitefish embryos (Corregone spp.) from Switzerland assayed whole eggs that were rinsed in sterile water before DNA extraction and 16s rDNA PCR. They describe an early rich microbial community that becomes less diverse (through alpha and beta metrics) over developmental time, potentially as a result of the maturing immune system. This study is inconclusive in revealing a microbiota within the extraembryonic fluid, since these communities could have been embedded in the outer chorion. However, the results suggest a reevaluation of the presumed-sterile teleost egg by focusing on native habitats instead of captive bred populations.
Fish eggs are immunologically active. Zebrafish embryos have an active innate immune response with macrophages and neutrophils maturing early in development [55,56], while adaptive immune responses mature after hatching [57]. Recent work using proteomics has revealed enhanced innate immune functions in perivitelline fluid in zebrafish embryos over three stages of development [58]. Peptide mass spectrometry revealed candidate lectins, glucosidases, alpha-macroglobulin protease inhibitors, and chitinase modifying enzymes inherited from the mother as well as iron co-factor chelation by zygotic transferrin-a. Multiple papers have found trans-generational immune priming in teleost fish, which is the inheritance of parental innate and adaptive immune components that can differ based on the parental infection status [59]. It should be noted that a strong early immune response does not prevent the establishment of symbiotic microbiota [60], and in many cases it is important for establishing the correct microbial communities [61].
The apparent lack of a stable microbiome inside the teleost egg does not mean that the surface microbiota of those eggs does not influence development. A stable microbiota has been found on the outer chorion of multiple wild-collected fish species, which differs from the surrounding water and/or substrate [32,62] and the composition of these chorion-associated bacteria has been found to influence hatching and survival [63,64].
Additional embryo-associated microbiomes are found in association with fish that engage in parental care. These include stable microbial communities associated with “pregnant” male pipefish (Sygnathus typhle), which is apparently attenuated by immunological priming of the father [65]. Interestingly, this study found a distinct microbial population on the surface of female gonadal eggs using 16s rDNA metabarcoding, but did not examine the inside of these eggs. In both the S. typhle brood pouch and Corregonus spp. (whitefish [32]) chorion the bacterial alpha diversity decreases over developmental time, again suggesting management by the developing immune system of the embryos. Parental care may facilitate different modes, or extent, of transgenerational microbial transmission which may be enhanced in these classically K-selected species. However, regardless of reproductive guild, most evidence to date suggests that healthy teleosts develop inside relatively sterile extraembryonic fluid. There are no studies to date on the possible sterility of cyclostome, elasmobranch, or non-teleost osteichthyan extraembryonic fluid or their potential microbiota.
4.2. Amphibians
Amphibians also have a large number of reproductive modes [66] which corresponds to a wide variety of egg structures [11]. Their developing embryos often succumb to infection from bacterial, fungal, or viral diseases [67,68,69]. However, the microbiota of their embryos has received less attention than the work on fish.
Most amphibian microbiome research focuses on the potentially beneficial cutaneous bacteria that may ward off infectious diseases in adults [70]. Several studies have investigated the recruitment of this cutaneous microbiota over post-hatching development [71]. It appears that egg-associated bacteria found on the outer surface of the egg capsules differs from the surrounding substrate and may be influenced by an attending parent [72,73,74,75]. However, no studies to date have examined whether bacteria within the extraembryonic fluid have a role in establishing the adult cutaneous microbiome.
The original evidence that extraembryonic fluid of many amphibians is not sterile is attributable to the common presence of the conspicuous green algae, Oophila amblystomatis [76]. To date, it has been found in the fluid of several ranid frog, and ambystomatid and hynobiid salamander species (reviewed in [27]). The stability and persistence of other microbes contained within amphibian egg capsules across evolution and development has not been thoroughly established. However, preliminary investigation of eastern redbacked salamanders using 16s rDNA metabarcoding has also revealed a stable bacterial community inside terrestrial-developing salamander egg capsules (unpublished observation).
It has been known for decades that the fluid of Ambystoma maculatum (spotted salamander) egg capsules also contain ciliates [77], a large number of commensal protists [78], and bacteria [22,33]. The diversity of potential eukaryotic commensals was recently called into question through 18s rDNA metabarcoding: Jurga et al. [79] revealed only a three-taxon eukaryotic microbiota inside A. maculatum egg capsules from Nova Scotia, which includes an occasional chytridiomycete, as well as consistent green algae (Oophila amblystomatis), and cercozoan populations. The closest sequence match to the chytridomycete extracted from the fluid is Rhyzophidium patellarium which is an algal parasite [80], suggesting a potential antagonistic relationship with the algae inside the egg capsule microbiome. Interestingly, the cercozoan and chytridiomycete variants were not found in association with the European agile frog (Rana dalmatina) despite the presence of Oophila amblystomatis in these eggs [27]. Although the sampling techniques for these two studies differed.
The bacteria within the A. maculatum egg capsule was recently identified from two sites in Nova Scotia [33]. Metabarcoding with 16s rDNA primers revealed several phyla conserved between the sites, and bacterial culturing consistently recovered isolates of Herbaspirilium sp. This Gram-negative obligate aerobic species belongs to the Burkholderiales, and has not found to be associated with other vertebrate embryos (Figure 2).
The embryo-associated microbiome in amphibians may have other protective roles. Early speculation that microbes could produce mycosporine-like amino acids that absorb UV irradiation, and are therefore protective to their hosts, has not yet been validated [81]. Additional host physiological and developmental effects from the in ovo microbiota in amphibians remains to be determined.
4.3. Reptiles and Birds
The interior of healthy bird eggs is also thought to be sterile [37,82,83] with neonate microbiome establishment occurring after hatching [84,85,86]. However, several studies have focused on the superficial bacteria found on bird and reptile eggs. The uropygial gland of multiple bird species produces antimicrobial secretions that protect the egg from infection [82,87,88,89]. In the European hoopoe’s (Upupa epops) and red-bellied wood hoopoe (Phoeniculus purpureus), these antibiotics are produced by symbiotic bacteria [90,91]. Unlike several other species surveyed, U. epops’ egg shells have specialized crypts that adhere to uropygial secretions and contain beneficial enterococcal bacteria. This may be unique to hoopoes. There is no evidence of transgenerational bacterial transmission in woodlarks or skylarks, which acquire their eggshell mcrobiome from the environment [84]. Similarly endangered Eretmochelys imbricate (hawksbill sea turtles; [92]) and Lepidochelys olivacea (Olive-Ridley sea turtles; [93]) acquire their eggshell microbiota from the environment. These bacteria aid in pathogen defense and hatching success, respectively.
Unlike birds, crocodiles and turtles, the squamates (lizards, amphisbaenians, and snakes) have a wide range of life history modes: including viviparity, ovoviviparity, and oviparity [94]. A recent study of gut microbiome acquisition in three viviparous leiolaemid lizards from Argentina found conservation of gut bacteria between mothers and their offspring, which was interpreted as evidence for potential transgenerational transmission [95]. Although this was not directly assessed.
The inaccessibility of microorganisms to the developing embryo in oviparous amniotes may also constrain opportunities for microbiota establishment. This is likely driven by adaptations to avoid pathogens. Despite the prevalence of vertical parasite transmission in mammals [96], there is only one example of a matrilineal inheritance of a parasite in egg-laying amniotes. A French population of the European wall lizard (Podarcis muralis) has been found to transmit a transovarial parasitic nematode (Suborder Spirurina), which can enter the extrembryonic membranes and embryonic brain [97]. The rarity of vertical transmission in oviparous amniotes is likely due to the early formation of the egg shell in birds and turtles, which occurs soon after fertilization. Egg shell formation is delayed in lizards and snakes until after the onset of organogenesis [98].
The majority of evidence that bacteria are capable of entering amniote eggs comes from studies of Salmonella enteridis entry in chickens [99]. The routes of entry include maternal diet, systemic spread, or ascending infection through the oviduct. Eggshell penetration can also occur with opportunistic bacteria, including Pseudomonas sp., Alcaligenes sp., Carnobacterium sp. [100], and Campylobacter [101], indicating that the oviparous egg may be susceptible to a wider range of potentially commensal and mutualistic bacteria.
One study to date has used culture-free 16s metabarcoding to look into the bacterial communities inside the egg shell. Trevelline et al. [35] did not specifically target the amniotic fluid, but punctured ethanol-sterilized eggs from eastern sand lizards (Sceloporus undulates) and multiple bird species with a pipette tip and aspirated contents for DNA extraction and 16s rDNA metabarcoding (Figure 2). Interestingly, they found a bacterial community distinct from environmental saline-rinse controls, while the fungal community was not. This study suggests an enrichment for environmental bacteria in the in ovo microbiota in non-diseased embryos.
Given the controversy of the potential “in utero colonization” in humans, it is remarkable that so little research attention has been given to this subject in oviparous amniotes. Unfortunately, the same methodological artefacts from culture-free metabarcoding surveys needs to be considered [31,102], and careful studies would benefit from a combination of culturing and sequencing in order to validate results.
4.4. Mammals
While most of the in utero microbiome research has focused on the human womb, there is a small body of comparative research within mammals that indicates a low-abundance microbiota may exist in some species.
A handful of studies exist for marsupial pouches, which accommodate long developmental periods following relatively short gestations [103]. Earlier work using culture-based methods found early establishment of an intestinal microbiota in pouch-young quokka (Setonix brachyurus; [104]), and brushtail possum (Trichosurus vulpecula; [105]), although healthy koala (Phascolarctos cinereus) pouch cultures lead to no microbial growth [106]. Metabarcoding surveys of pouch microbiota of southern hairy-nosed wombats (Lasiorhinus latifrons; [107]) found similar bacterial taxa with tammar wallaby (Macropus eugenii; [108]), while Tasmanian devil (Sarcophilus harrisii) pouch microbiota was found to be largely similar to the adult skin [109].
Comparative studies in placental mammals differ in their findings of a sterile in utero environment. Mice and humans have a highly invasive hemochorial placenta, where fetal tissue is in direct contact with maternal blood. Ruminants have a less permissive epitheliochorial placenta, where the fetal tissue is separated from maternal circulation by multiple tissue layers. A meta-analysis of placental types found that ruminants and other epitheliochorial placenta-bearing mammals have a higher overall pathogen load than hemochorial species, suggesting that the more restrictive barrier is a by-product of pathogen co-evolution [110]. Subsequent work using 16s amplicon sequencing has found the epitheliochorial lamb amnion, placenta, and fetal intestine [111], as well as the murine placenta and fetal tissues [38], to be sterile. However, the intrauterine transfer of genetically labeled Enterococcus faecium was detected in the amniotic fluid [112] and meconium [113] in preterm mouse pups that were delivered by caesarian section.
There is a long history of research into the potential in utero microbiota encountered by developing humans [114]. The field had largely resolved that the fetal microenvironment is sterile, before modern molecular-based methods of microbial detection [115,116].
Most of the modern in utero microbiome research has focused on the placenta. Aagaard et al. [117] used 16s rDNA metabarcoding to find a human placental microbiome that is similar to the oral microbiome, although subsequent studies raised contamination concerns [118], and later failed to find a stable microbiota once compared to blank controls [119]. Additional placental microbiome studies [120,121,122,123] have been criticized for failing to properly control for environmental contamination [31,115]. However, a very small number of bacteria are detectable with HCR-FISH in human placentas following delivery, without evidence for chorioamnioinitis [23]. Therefore, some independent lines of evidence now suggest some bacteria within the healthy human placenta, although at extremely low abundance.
Additional work has also focused on the human amniotic fluid and neonatal meconium. Early culture-based techniques found healthy newborn meconium to be sterile [124,125] while subsequent work with PCR and FISH found a rapid, potentially prenatal bacterial colonization in healthy infants [126]. Similarly, definitive culture-based approaches found amniotic fluid to be sterile [127], which has largely been confirmed by metagenomic cell-free DNA sequencing with stringent low abundance filtering [4]. However, a recent metabarcoding survey [128] using stringent decontamination and contaminant controls found an amniotic and meconial microbiota which correlated with amniotic cytokine levels and meconium propionate levels [129].
While the establishment of a commensal or even mutualistic in utero human microbial consortia remains controversial, there are many pathogens that are capable of maternal transfer. These include viruses such as rubella, herpes simplex and zika; parasites such as Toxoplasma gondi; and bacteria including syphilis and chlamydia [96]. Therefore, potential transplacental or ascending routes of microbial entry exist for pathogens, which may be employed by non-pathogenic microbes as well.
The implications for in utero colonization of the human microbiome remains a controversial possibility with implications for the formation of the immune system, fetal-microbial interactions, and the establishment of an adult microbiome [130,131]. However, if there is a healthy and stable fetal microbiome, it is an exceptionally sparse community.
The reduced in utero microbiome does not necessarily mean that the developing human embryo/ fetus is insulated from microbial influence. Recent single-cell transcriptomics studies have revealed the surprisingly early differentiation of myeloid and lymphoid components of the human immune system during the first trimester (reviewed in [132]). Maternally derived bacterial metabolites (e.g., lipopolysaccharides or flagellin) from the intestines and breast milk have been shown to influence the biochemical environment and immune system of the developing fetus [133,134]. Gut microbes from pregnant mice can influence offspring metabolic phenotypes in utero [135]. The direct fetal interaction with microbial metabolites is one form of transgenerational immune priming, which can also include transfer of innate and adaptive immune effectors, small RNA’s, histone modifications, and potentially DNA methylation [59].
5. Gnotobiototic Vertebrates
The history of gnotobiotic/axenic research extends back to Louis Pasteur [136] and now spans over 125 years [137]. A large number of gnotobiotic vertebrates have been established, including fish, amphibians, birds, and mammals [138,139,140]. All of the derivation techniques include isolating and sterilizing early embryos, which are then raised in sterile conditions. For instance, axenic mice are routinely derived from an aseptic hysterectomy (mouse caesarian section) of conventional pregnant females followed by a germicidal bath [141]. Axenic zebrafish are similarly derived from antibiotic-treated embryos following in vitro fertilization and later aseptically removed from their chorion [142]. Even human neonates born with severe combined immunodeficiency disorders have been raised “germ free” [143]. Gnotobiotic vertebrates do not necessarily resolve controversies over the phylogenetic distribution of in ovo or in utero microbiota, since the derivation techniques include sterilizing embryos or neonates. However, they do suggest that none of the potential embryo-associated microbiota are essential for survival.
6. Multiciliated Surface Cells
The patchy evidence for a stable in utero microbial community in placental mammals makes it difficult to assess the evidence for either the presence or absence of embryo-associated microbiota in other vertebrates. Received wisdom from fish and oviparous amniote research suggests that their healthy embryonic/fetal microenvironments are typically sterile, although neither of these groups have received the intense level of scrutiny given to humans. The few studies that have looked at amphibian in ovo microbiota corroborate earlier accounts of a diverse assemblage in some species, although most comparative studies focus on surface bacteria. Given the scarcity of evidence, it is impossible to draw any meaningful correlations between microbiota structure, or even presence, with morphology. However, morphology may provide clues as to which vertebrates should be studied further for core microbiota within their extraembryonic fluids.
6.1. Vertebrate Embryos with Multiciliated Surface Cells
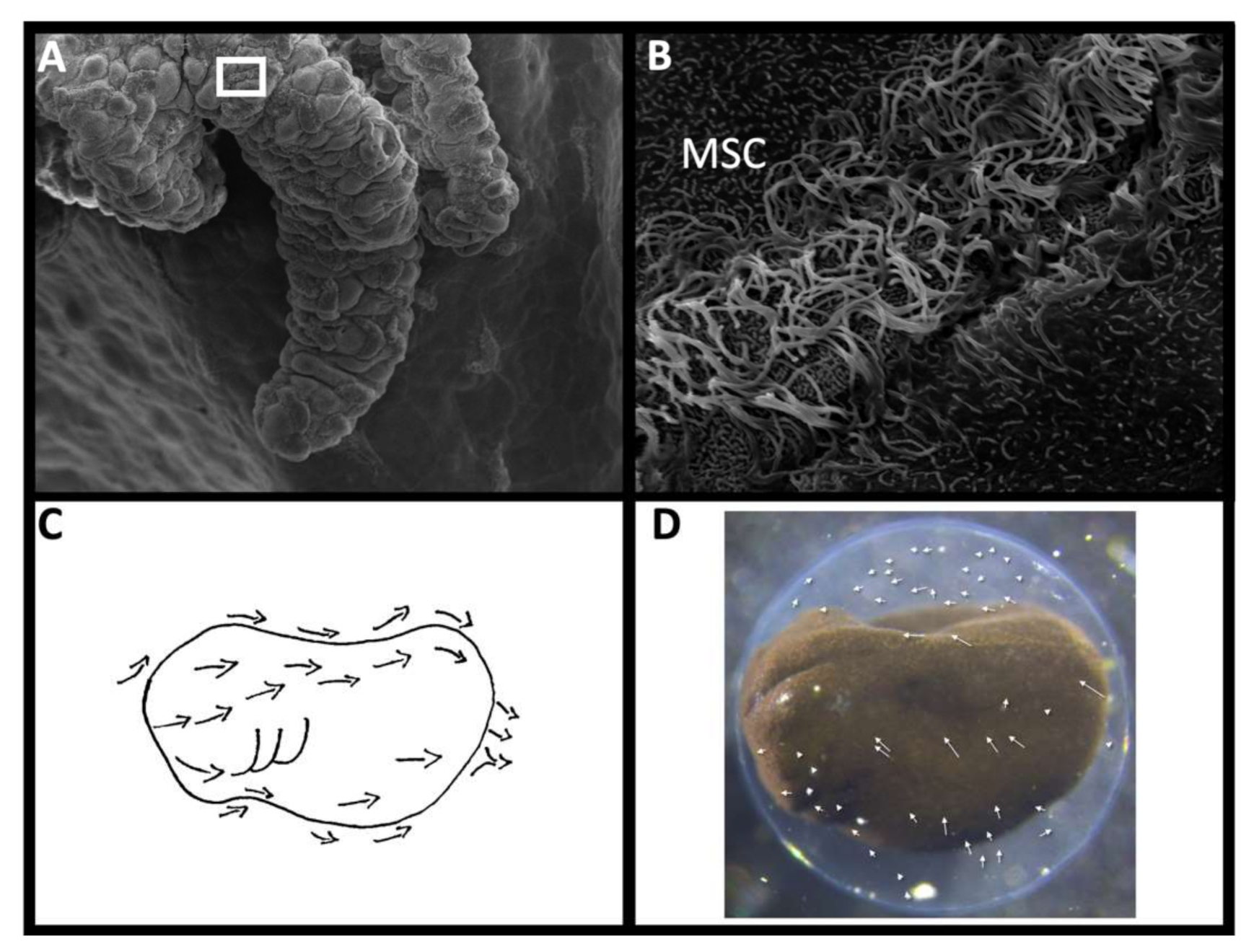
In particular, the multiciliated surface cells (MSC) on the surface of some aquatic embryos may have a role in managing extraembryonic microbes. These widespread epithelial cells create a synchronized metachronal beat which generates current around developing catshark (Scyliorhinus canicula [144]), frog [145,146,147,148]; caecilian [149,150]; salamander [151,152,153,154], Australian lungfish (Neoceratodus fosteri; [3], and multiple invertebrates [155] including amphioxus [156]. A more restrictive MSC population associated with the lateral line system has also been reported for bichir (Polypterus senegalensis, P. palmas; [157,158]), gar (Lepisosteus platyrhincus [159,160]) and sturgeon (Acipenser transmontanus [157]), and they have been found in the olfactory pit of the paddlefish (Polyodon spatula [161]). However, the homology of these localized MSCs in basal actinopterygians with the more dispersed MSCs of lungfish and amphibians has been called into question [157].
MSCs are just one type of multiciliated cell found in eukaryotes [162]. Vertebrates also form multiciliated epithelial cells in oviducts/ fallopian tubes, embryonic and adult kidneys, ependymal lining of the ventricles and spinal cord, and the tracheobronchial tree [162]. These cells differ from the monociliated cells of Henson’s node or Kupfer’s vesicle, which are implicated in cellular signaling [145,162], but do not have the multiple cilia and basal bodies that are formed de novo in multiciliated cells [163]. The presence of embryonic MSCs has been used as a synapomorphy to unite lungfish with tetrapods [164,165], although the presence of similar cells in basal actinopterygians and at least one shark, suggests either convergent gain or loss across the vertebrates [157]. The widespread occurrence of the multiciliated epithelial cell phenotype in a variety of tissues suggests that this convergence is possible.
6.2. Origin and Morphology of MSCs
Traditionally, most research on the function of surface MSCs has been on amphibians [166]. The MSC population of Xenopus laevis (African clawed frog) have been the subject of close scrutiny from a cellular and developmental perspective. These are investigated as a model for understanding cilia formation, dynamics, and ciliopathies in humans (such as Bardet–Biedl syndrome, e.g., [162,167]). Multiciliated surface cells form from a lightly pigmented deep layer of ectoderm, which migrate to the surface by intercalating outer epithelial cells [154]. These kit tyrosine kinase receptor-presenting cells become evenly distributed during the migration process through mutual repulsion, mediated by the soluble Kit ligand Scf, and affinity to the surface cells, which is mediated by membrane-bound Scf [168]. MSC differentiation is dependent on repressing Delta/Notch potentially through miR449 [169], which results in expression of the transcriptional Multicilin/E2f complex (MCIDAS/E2f4-E2f5) that drives expression of downstream ciliogenic regulators and major ciliary proteins [170].
Like the mammalian airway, MSCs are classified as mucociliary cells. The embryonic epithelia of Xenopus includes ionocytes, small secretory cells, goblet cells, and MSCs, which together create and move a complex mucosal barrier to infection [171]. The outer ciliary axonemes of individual MSCs beat at the same frequency, but in a staggered phase-shift that creates a sequential “metachronal” wave and directional current [172]. The planar cell polarity pathway coordinates the MSC directionality across the epithelium by coordinating MSC polarization at the tissue level [173]. The currents collectively generated run from anterior to posterior in lungfish [164] and amphibian [174] embryos (Figure 3C). These currents not only move the cortical mucous, but also rotate the embryos within their egg capsules, and create complex vortices within the capsular fluid (Figure 3D).
6.3. Proposed Functions of MSCs
The proposed functions of MSCs include cutaneous respiration [175,176,177], pre- and post-hatchling movement [147,153,174,178], detecting chemical cues [145,174], and clearing debris [41,174]. In their assessment of potential roles multiciliated surface cells might play Nokhbatolfoghahai et al. [145] assert that these are not mutually exclusive functions. However, the majority of researchers emphasize a primarily respiratory function [145,153,179]. The currents generated by MSCs reduce the “diffusion boundary layer” of respiratory media that surrounds embryos, potentially decreasing stagnation and increasing gas exchange [176].
However, if these cilia are exclusively used in respiration, then why are they not more prevalent in externally-developing teleosts, which presumably contend with similar hypoxic environments [180]? Multiple teleosts have been reported to use cutaneous respiration in early larval, and possibly embryonic stages. However, both the rice eel (Monopterus albus; [181]) and tilapia (Oreochromis mossambicus; [182]) use their fins to generate currents for cutaneous gas exchange. Burggren [175] points out that a cost–benefit analysis has not been calculated for the oxygen demands of ciliary motion against apparent oxygenation benefit (which is still true nearly 40 years later). Kemp [41] asserts that the respiratory function would be minimal in lungfish, and instead emphasizes the clearing of debris and preventing the settlement of pathogens, which can enter these eggs through conspicuous pores prior to hatching in Neoceratodus fosteri.
Assheton [174] and Nokhbatolfoghahai et al. [145] both describe increased ciliary flow surrounding the anuran adhesive glands, which is consistent with trapping debris, although Nokhbatolfoghahai et al. [145] also asserts that any debris in the amphibian egg capsule must be embryonic cells or foreign pathogens, since they assume the disease-free in ovo environment is sterile.
6.4. Boundary Layers in Microbial Recruitment
Multiple independent studies have found the establishment of a boundary layer to be essential in microbiota recruitment and cultivation. The physical management of bacterial settlement depends on localized control of flow [183]. Dancing yeti crabs of deep-sea vents (Kiwa puravuda) selectively recruit chemosynthetic bacteria on their waving setae, which eventually becomes their primary food [184]; lung-associated bacteria and viruses directly affect cilia formation and dynamics in the lung, while mucocilliary clearance winnows the bacterial load [185,186]; and the currents established by cilia of multiple lengths clear mucus and selectively recruit bacteria through the controlled distribution of chemical signals and the physical microenvironment surrounding the bobtailed squid (Euprymna scolopes) light organ [187].
Additionally, microbial influence on host boundary layer formation suggests complex communication between the two. This is clearly revealed in microbial control of spontaneous contractions in Hydra spp. [188], and the increased embryonic rotation associated with algal load (Oophila amblystomatis) inside spotted salamander (A. maculatum) egg capsules [153].
If the surface ciliation of vertebrate embryos was strictly for respiration, then we would anticipate seeing more externally developing species with this feature. However, given the sporadic distribution of MSCs in a handful of vertebrate taxa, and the handful of comparative studies on actual embryonic microbiomes, this review can only suggest that future efforts in elucidating the function of extraembryonic microbial consortia focus on those taxa that are known to have MSCs.
7. Conclusions
There has been a strong emphasis on the potential implications of embryo-associated microbiomes in the recent literature [1,2,6,189,190]. Embryo-associated microbes have been shown to have a role in embryo defense, development, metabolic exchanges, and vertical/ transgenerational transmission [6]. However, as this review illustrates, we are still largely unaware of which vertebrate animals actually develop within a microbial community. The evidence at hand is inconclusive due to lack of targeted sampling as well as potential false-positive signals from PCR-based techniques [31]. However, even if a cultivated extraembryonic microbiota exists within teleosts and amniotes, it is likely not abundant. Limited data suggest amphibians [79] and lungfish [41] encounter heavy microbial loads in healthy wild-caught eggs. Both of these lineages have extensive multicilliated cells on the embryonic and occasional larval integument, suggesting that the currents created by these cells may have an under-appreciated role in embryo microbial consortia establishment and maintenance. Whether this trend holds in additional vertebrate groups with MSCs (e.g., basal actinopterygians and some elasmobrachs), and whether these microbial consortia are voluntary or cultivated, remain to be seen. A targeted approach to establishing which lineages cultivate an extraembryonic microbial community could open up new lines of inquiry into microbial effects on development in an important evolutionary lineage: the vertebrates.
Funding
Funding was provided through the Gordon and Betty Moore Foundation (Grant #GBMF5604; https://doi.org/10.37807/GBMF5604 (accessed on 22 October 2021).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Thanks to Raul Diaz for an invitation to submit to this Special Issue. Special thanks to members of the Carnegie Institution of Washington’s “Symbiosis Journal Club,” especially John Sittmann and Yixian Zheng, for productive discussions on earlier drafts of this work. Additional thanks to Elliana Vickers for researching bacterial phyla detected in the metabarcoding literature.
Conflicts of Interest
The authors declare no conflict of interest.
References
- McFall-Ngai, M.; Bosch, T.C.G. Animal development in the microbial world: The power of experimental model systems. In Current Topics in Developmental Biology; Elsevier: Amsterdam, The Netherlands, 2021; Volume 141, pp. 371–397. ISBN 978-0-12-814968-3. [Google Scholar]
- Bosch, T.C.G.; McFall-Ngai, M. Animal development in the microbial world: Re-thinking the conceptual framework. In Current Topics in Developmental Biology; Elsevier: Amsterdam, The Netherlands, 2021; Volume 141, pp. 399–427. ISBN 978-0-12-814968-3. [Google Scholar]
- Popescu, L.; Cao, Z.-P. From Microscopy to Genomic Approach in Soil Biodiversity Assessment. Curr. Issues Mol. Biol. 2018, 27, 195–198. [Google Scholar] [CrossRef]
- Burnham, P.; Gomez-Lopez, N.; Heyang, M.; Cheng, A.P.; Lenz, J.S.; Dadhania, D.M.; Lee, J.R.; Suthanthiran, M.; Romero, R.; De Vlaminck, I. Separating the Signal from the Noise in Metagenomic Cell-Free DNA Sequencing. Microbiome 2020, 8, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammer, T.J.; Sanders, J.G.; Fierer, N. Not All Animals Need a Microbiome. FEMS Microbiol. Lett. 2019, 366. [Google Scholar] [CrossRef] [Green Version]
- Nyholm, S.V. In the Beginning: Egg–Microbe Interactions and Consequences for Animal Hosts. Philos. Trans. R. Soc. B Biol. Sci. 2020, 375, 20190593. [Google Scholar] [CrossRef]
- Kellam, L.; Pastorelli, L.M.; Bastida, A.M.; Senkbeil, A.; Montgomery, S.; Fishel, S.; Campbell, A. Perivitelline Threads in Cleavage-Stage Human Embryos: Observations Using Time-Lapse Imaging. Reprod. Biomed. Online 2017, 35, 646–656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shu, L.; Suter, M.J.-F.; Räsänen, K. Evolution of Egg Coats: Linking Molecular Biology and Ecology. Mol. Ecol. 2015, 24, 4052–4073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Claw, K.G.; Swanson, W.J. Evolution of the Egg: New Findings and Challenges. Annu. Rev. Genom. Hum. Genet. 2012, 13, 109–125. [Google Scholar] [CrossRef]
- Alteri, A.; Viganò, P.; Maizar, A.A.; Jovine, L.; Giacomini, E.; Rubino, P. Revisiting Embryo Assisted Hatching Approaches: A Systematic Review of the Current Protocols. J. Assist. Reprod. Genet. 2018, 35, 367–391. [Google Scholar] [CrossRef]
- Altig, R.; McDiarmid, R.W. Morphological Diversity and Evolution of Egg and Clutch Structure in Amphibians. Herpetol. Monogr. 2007, 21, 1–32. [Google Scholar] [CrossRef]
- Laale, H.W. The Perivitelline Space and Egg Envelopes of Bony Fishes: A Review. Copeia 1980, 1980, 210–226. [Google Scholar] [CrossRef]
- Kirunda, D.F.K.; McKee, S.R. Relating Quality Characteristics of Aged Eggs and Fresh Eggs to Vitelline Membrane Strength as Determined by a Texture Analyzer. Poult. Sci. 2000, 79, 1189–1193. [Google Scholar] [CrossRef]
- Damaziak, K.; Kieliszek, M.; Bucław, M. Characterization of Structure and Protein of Vitelline Membranes of Precocial (Ring-Necked Pheasant, Gray Partridge) and Superaltricial (Cockatiel Parrot, Domestic Pigeon) Birds. PLoS ONE 2020, 15, e0228310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryder, J. Development of the Silver Gar (Belone Longirostris) with Observations on the Genesis of the Blood in Embryo Fishes and a Comparison of Fish Ova with Those of Other Vertebrates. Bull. U. S. Fish Comm. 1882, 1, 283–301. [Google Scholar]
- Salthe, S.N. The Egg Capsules in the Amphibia. J. Morphol. 1963, 113, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Szarski, H. The Origin of Vertebrate Foetal Membranes. Evolution 1968, 22, 211–214. [Google Scholar] [CrossRef] [PubMed]
- Escherich, T. The Intestinal Bacteria of the Neonate and Breast-Fed Infant. Infect. Dis. 1885, 11, 352–356. [Google Scholar] [CrossRef] [PubMed]
- Reasoner, D.J.; Geldreich, E.E. A New Medium for the Enumeration and Subculture of Bacteria from Potable Water. Appl. Environ. Microbiol. 1985, 49, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Browne, H.P.; Forster, S.C.; Anonye, B.O.; Kumar, N.; Neville, B.A.; Stares, M.D.; Goulding, D.; Lawley, T.D. Culturing of ‘Unculturable’ Human Microbiota Reveals Novel Taxa and Extensive Sporulation. Nature 2016, 533, 543–546. [Google Scholar] [CrossRef] [Green Version]
- Walker, A.W.; Duncan, S.H.; Louis, P.; Flint, H.J. Phylogeny, Culturing, and Metagenomics of the Human Gut Microbiota. Trends Microbiol. 2014, 22, 267–274. [Google Scholar] [CrossRef]
- Kerney, R.; Kim, E.; Hangarter, R.P.; Heiss, A.A.; Bishop, C.D.; Hall, B.K. Intracellular Invasion of Green Algae in a Salamander Host. Proc. Natl. Acad. Sci. USA 2011, 108, 6497–6502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seferovic, M.D.; Pace, R.M.; Carroll, M.; Belfort, B.; Major, A.M.; Chu, D.M.; Racusin, D.A.; Castro, E.C.C.; Muldrew, K.L.; Versalovic, J.; et al. Visualization of Microbes by 16S in Situ Hybridization in Term and Preterm Placentas without Intraamniotic Infection. Am. J. Obstet. Gynecol. 2019, 221, 146.e1–146.e23. [Google Scholar] [CrossRef] [PubMed]
- Borneman, J.; Hartin, R.J. PCR Primers That Amplify Fungal RRNA Genes from Environmental Samples. Appl. Environ. Microbiol. 2000, 66, 4356–4360. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.; Simpson, A.G.; Graham, L.E. Evolutionary Relationships of Apusomonads Inferred from Taxon-Rich Analyses of 6 Nuclear Encoded Genes. Mol. Biol. Evol. 2006, 23, 2455–2466. [Google Scholar] [CrossRef] [Green Version]
- Frank, J.A.; Reich, C.I.; Sharma, S.; Weisbaum, J.S.; Wilson, B.A.; Olsen, G.J. Critical Evaluation of Two Primers Commonly Used for Amplification of Bacterial 16S RRNA Genes. Appl. Environ. Microbiol. 2008, 74, 2461–2470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anslan, S.; Sachs, M.; Rancilhac, L.; Brinkmann, H.; Petersen, J.; Künzel, S.; Schwarz, A.; Arndt, H.; Kerney, R.; Vences, M. Diversity and Substrate-Specificity of Green Algae and Other Micro-Eukaryotes Colonizing Amphibian Clutches in Germany, Revealed by DNA Metabarcoding. Sci. Nat. 2021, 108, 29. [Google Scholar] [CrossRef] [PubMed]
- Collado, M.C.; Rautava, S.; Aakko, J.; Isolauri, E.; Salminen, S. Human Gut Colonization May Be Initiated in utero by Distinct Microbial Communities in the Placenta and Amniotic Fluid. Sci. Rep. 2016, 6, 23129. [Google Scholar] [CrossRef] [Green Version]
- Hazen, T.C.; Dubinsky, E.A.; DeSantis, T.Z.; Andersen, G.L.; Piceno, Y.M.; Singh, N.; Jansson, J.K.; Probst, A.; Borglin, S.E.; Fortney, J.L.; et al. Deep-Sea Oil Plume Enriches Indigenous Oil-Degrading Bacteria. Science 2010, 330, 204–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balvočiūtė, M.; Huson, D.H. SILVA, RDP, Greengenes, NCBI and OTT—How Do These Taxonomies Compare? BMC Genom. 2017, 18, 114. [Google Scholar] [CrossRef] [Green Version]
- Eisenhofer, R.; Minich, J.J.; Marotz, C.; Cooper, A.; Knight, R.; Weyrich, L.S. Contamination in Low Microbial Biomass Microbiome Studies: Issues and Recommendations. Trends Microbiol. 2019, 27, 105–117. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, L.G.E.; Rogivue, A.; Fumagalli, L.; Wedekind, C. Declining Diversity of Egg-Associated Bacteria during Development of Naturally Spawned Whitefish Embryos (Coregonus Spp.). Aquat. Sci. 2015, 77, 481–497. [Google Scholar] [CrossRef]
- Bishop, C.; Jurga, E.; Graham, L. Patterns of Bacterial Diversity in Embryonic Capsules of the Spotted Salamander Ambystoma Maculatum: An Expanding View of a Symbiosis. FEMS Microbiol. Ecol. 2021, 97, fiab128. [Google Scholar] [CrossRef] [PubMed]
- Borghi, E.; Massa, V.; Severgnini, M.; Fazio, G.; Avagliano, L.; Menegola, E.; Bulfamante, G.P.; Morace, G.; Borgo, F. Antenatal Microbial Colonization of Mammalian Gut. Reprod. Sci. 2019, 26, 1045–1053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trevelline, B.K.; MacLeod, K.J.; Knutie, S.A.; Langkilde, T.; Kohl, K.D. In ovo Microbial Communities: A Potential Mechanism for the Initial Acquisition of Gut Microbiota among Oviparous Birds and Lizards. Biol. Lett. 2018, 14. [Google Scholar] [CrossRef] [PubMed]
- Bakke, I.; Attramadal, K.J.K.; Vestrum, R.I.; Vadstein, O. Controlling Factors for Community Assembly in Developing Cod Larvae (Gadus morhua). In Microbial Communities in Aquaculture Ecosystems; Derome, N., Ed.; Springer International Publishing: Cham, Switzerland, 2019; pp. 69–93. ISBN 978-3-030-16189-7. [Google Scholar]
- Cook, M.I.; Beissinger, S.R.; Toranzos, G.A.; Rodriguez, R.A.; Arendt, W.J. Trans–Shell Infection by Pathogenic Micro–Organisms Reduces the Shelf Life of Non–Incubated Bird’s Eggs: A Constraint on the Onset of Incubation? Proc. R. Soc. Lond. B Biol. Sci. 2003, 270, 2233–2240. [Google Scholar] [CrossRef] [Green Version]
- Theis, K.R.; Romero, R.; Greenberg, J.M.; Winters, A.D.; Garcia-Flores, V.; Motomura, K.; Ahmad, M.M.; Galaz, J.; Arenas-Hernandez, M.; Gomez-Lopez, N. No Consistent Evidence for Microbiota in Murine Placental and Fetal Tissues. mSphere 2020, 5. [Google Scholar] [CrossRef] [Green Version]
- Froese, R.; Pauly, D. FishBase. World Wide Web Electronic Publication; ScienceOpen, Inc.: Burlington, MA, USA, 2014. [Google Scholar]
- Balon, E. Epigenesis of an Epigeneticist: The Development of Some Alternative Concepts on the Early Ontogeny and Evolution of Fishes. Guelph Ichthyol. Rev. 1990, 1, 1–48. [Google Scholar]
- Kemp, A. Role of Epidermal Cilia in Development of the Australian Lungfish, Neoceratodus Forsteri (Osteichthyes: Dipnoi). J. Morphol. 1996, 228, 203–221. [Google Scholar] [CrossRef]
- De Schryver, P.; Vadstein, O. Ecological Theory as a Foundation to Control Pathogenic Invasion in Aquaculture. ISME J. 2014, 8, 2360–2368. [Google Scholar] [CrossRef] [Green Version]
- Gauthier, J.; Lavoie, C.; Charette, S.J.; Derome, N. Host-Microbiota Interactions and Their Importance in Promoting Growthand Resistance to Opportunistic Diseases in Salmonids. In Microbial Communities in Aquaculture Ecosystems: Improving Productivity and Sustainability; Derome, N., Ed.; Springer International Publishing: Cham, Switzerland, 2019; ISBN 978-3-030-16189-7. [Google Scholar]
- Derome, N.; Filteau, M. A Continuously Changing Selective Context on Microbial Communities Associated with Fish, from Egg to Fork. Evol. Appl. 2020, 13, 1298–1319. [Google Scholar] [CrossRef] [PubMed]
- Llewellyn, M.S.; Boutin, S.; Hoseinifar, S.H.; Derome, N. Teleost Microbiomes: The State of the Art in Their Characterization, Manipulation and Importance in Aquaculture and Fisheries. Front. Microbiol. 2014, 5. [Google Scholar] [CrossRef] [Green Version]
- Rawls, J.F.; Samuel, B.S.; Gordon, J.I. Gnotobiotic Zebrafish Reveal Evolutionarily Conserved Responses to the Gut Microbiota. Proc. Natl. Acad. Sci. USA 2004, 101, 4596–4601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkins, L.G.E.; Fumagalli, L.; Wedekind, C. Effects of Host Genetics and Environment on Egg-Associated Microbiotas in Brown Trout (Salmo trutta). Mol. Ecol. 2016, 25, 4930–4945. [Google Scholar] [CrossRef] [Green Version]
- Wilkins, L.G.E.; Rogivue, A.; Schütz, F.; Fumagalli, L.; Wedekind, C. Increased Diversity of Egg-Associated Bacteria on Brown Trout (Salmo trutta) at Elevated Temperatures. Sci. Rep. 2015, 5, 17084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; de Bruijn, I.; Jack, A.L.; Drynan, K.; van den Berg, A.H.; Thoen, E.; Sandoval-Sierra, V.; Skaar, I.; van West, P.; Di guez-Uribeondo, J.; et al. Deciphering Microbial Landscapes of Fish Eggs to Mitigate Emerging Diseases. ISME J. 2014, 8, 2002–2014. [Google Scholar] [CrossRef]
- Hansen, G.H.; Olafsen, J.A. Bacterial Colonization of Cod (Gadus morhua L.) and Halibut (Hippoglossus hippoglossus) Eggs in Marine Aquaculture. Appl. Environ. Microbiol. 1989, 55, 1435–1446. [Google Scholar] [CrossRef] [Green Version]
- Stephens, W.Z.; Burns, A.R.; Stagaman, K.; Wong, S.; Rawls, J.F.; Guillemin, K.; Bohannan, B.J.M. The Composition of the Zebrafish Intestinal Microbial Community Varies across Development. ISME J. 2016, 10, 644–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eddy, F.B.; Ward, M.R.; Talbot, C.; Primmett, D. Ionic Movements across the Chorion in Newly Shed Salmon Eggs (Salmo salar L.). J. Comp. Physiol. B 1990, 159, 771–776. [Google Scholar] [CrossRef]
- Groot, E.P.; Alderdice, D.F. Fine Structure of the External Egg Membrane of Five Species of Pacific Salmon and Steelhead Trout. Can. J. Zool. 1985, 63, 552–566. [Google Scholar] [CrossRef]
- Yanagimachi, R.; Harumi, T.; Matsubara, H.; Yan, W.; Yuan, S.; Hirohashi, N.; Iida, T.; Yamaha, E.; Arai, K.; Matsubara, T.; et al. Chemical and Physical Guidance of Fish Spermatozoa into the Egg through the Micropyle. Biol. Reprod. 2017, 96, 780–799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Stockhammer, O.W.; de Boer, L.; Vischer, N.O.E.; Spaink, H.P.; Grijpma, D.W.; Zaat, S.A.J. The Zebrafish Embryo as a Model to Quantify Early Inflammatory Cell Responses to Biomaterials: The Zebrafish Embryo Model. J. Biomed. Mater. Res. A 2017, 105, 2522–2532. [Google Scholar] [CrossRef]
- Benard, E.L.; van der Sar, A.M.; Ellett, F.; Lieschke, G.J.; Spaink, H.P.; Meijer, A.H. Infection of Zebrafish Embryos with Intracellular Bacterial Pathogens. J. Vis. Exp. 2012, 3781. [Google Scholar] [CrossRef] [PubMed]
- van der Sar, A.M.; Spaink, H.P.; Zakrzewska, A.; Bitter, W.; Meijer, A.H. Specificity of the Zebrafish Host Transcriptome Response to Acute and Chronic Mycobacterial Infection and the Role of Innate and Adaptive Immune Components. Mol. Immunol. 2009, 46, 2317–2332. [Google Scholar] [CrossRef] [PubMed]
- De la Paz, J.F.; Anguita-Salinas, C.; Díaz-Celis, C.; Chávez, F.P.; Allende, M.L. The Zebrafish Perivitelline Fluid Provides Maternally-Inherited Defensive Immunity. Biomolecules 2020, 10, 1274. [Google Scholar] [CrossRef] [PubMed]
- Roth, O.; Beemelmanns, A.; Barribeau, S.M.; Sadd, B.M. Recent Advances in Vertebrate and Invertebrate Transgenerational Immunity in the Light of Ecology and Evolution. Heredity 2018, 121, 225–238. [Google Scholar] [CrossRef] [Green Version]
- Gensollen, T.; Iyer, S.S.; Kasper, D.L.; Blumberg, R.S. How Colonization by Microbiota in Early Life Shapes the Immune System. Science 2016, 352, 539–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Archetti, M.; Ubeda, F.; Fudenberg, D.; Green, J.; Pierce, N.E.; Yu, D.W. Let the Right One in: A Microeconomic Approach to Partner Choice in Mutualisms. Am. Nat. 2011, 177, 75–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wedekind, C.; Gessner, M.O.; Vazquez, F.; Maerki, M.; Steiner, D. Elevated Resource Availability Sufficient to Turn Opportunistic into Virulent Fish Pathogens. Ecology 2010, 91, 1251–1256. [Google Scholar] [CrossRef] [Green Version]
- Clark, E.S.; Wilkins, L.G.E.; Wedekind, C. MHC Class I Expression Dependent on Bacterial Infection and Parental Factors in Whitefish Embryos (Salmonidae). Mol. Ecol. 2013, 22, 5256–5269. [Google Scholar] [CrossRef] [Green Version]
- Clark, E.S.; Pompini, M.; Marques da Cunha, L.; Wedekind, C. Maternal and Paternal Contributions to Pathogen Resistance Dependent on Development Stage in a Whitefish (Salmonidae). Funct. Ecol. 2014, 28, 714–723. [Google Scholar] [CrossRef]
- Beemelmanns, A.; Poirier, M.; Bayer, T.; Kuenzel, S.; Roth, O. Microbial Embryonal Colonization during Pipefish Male Pregnancy. Sci. Rep. 2019, 9, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wells, K. The Ecology and Behavior of Amphibians; University of Chicago Press: Chicago, IL, USA, 2007. [Google Scholar]
- Beattie, R.C. A Physico-Chemical Investigation of the Jelly Capsules Surrounding Eggs of the Common Frog (Rana temporaria temporaria). J. Zool. 2009, 190, 1–25. [Google Scholar] [CrossRef]
- Gomez-Mestre, I.; Touchon, J.C.; Warkentin, K.M. Amphibian Embryo and Parental Defenses and a Larval Predator Reduce Egg Mortality from Water Mold. Ecology 2006, 87, 2570–2581. [Google Scholar] [CrossRef]
- Hamdoun, A.; Epel, D. Embryo Stability and Vulnerability in an Always Changing World. Proc. Natl. Acad. Sci. USA 2007, 104, 1745–1750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rebollar, E.A.; Martínez-Ugalde, E.; Orta, A.H. The Amphibian Skin Microbiome and Its Protective Role Against Chytridiomycosis. Herpetologica 2020, 76, 167. [Google Scholar] [CrossRef]
- Loudon, A.H.; Venkataraman, A.; Van Treuren, W.; Woodhams, D.C.; Parfrey, L.W.; McKenzie, V.J.; Knight, R.; Schmidt, T.M.; Harris, R.N. Vertebrate Hosts as Islands: Dynamics of Selection, Immigration, Loss, Persistence, and Potential Function of Bacteria on Salamander Skin. Front. Microbiol. 2016, 7, 333. [Google Scholar] [CrossRef]
- Hughey, M.C.; Delia, J.; Belden, L.K. Diversity and Stability of Egg-Bacterial Assemblages: The Role of Paternal Care in the Glassfrog Hyalinobatrachium colymbiphyllum. Biotropica 2017, 49, 792–802. [Google Scholar] [CrossRef]
- Banning, J.L.; Weddle, A.L.; Wahl III, G.W.; Simon, M.A.; Lauer, A.; Walters, R.L.; Harris, R.N. Antifungal Skin Bacteria, Embryonic Survival, and Communal Nesting in Four-Toed Salamanders, Hemidactylium scutatum. Oecologia 2008, 156, 423–429. [Google Scholar] [CrossRef] [PubMed]
- Walke, J.B.; Becker, M.H.; Loftus, S.C.; House, L.L.; Cormier, G.; Jensen, R.V.; Belden, L.K. Amphibian Skin May Select for Rare Environmental Microbes. ISME J. 2014, 8, 2207–2217. [Google Scholar] [CrossRef] [PubMed]
- Prest, T.L.; Kimball, A.K.; Kueneman, J.G.; McKenzie, V.J. Host-Associated Bacterial Community Succession during Amphibian Development. Mol. Ecol. 2018, 27, 1992–2006. [Google Scholar] [CrossRef]
- Orr, H. Note on the Development of Amphibians, Chiefly Concerning the Central Nervous System; with Additional Observations on the Hypophysis, Mouth, and the Appendages and Skeleton of the Head. J. Cell Sci. 1888, 115, 295–324. [Google Scholar]
- Gilbert, P. Observations on the Eggs of Ambystoma maculatum with Especial Reference to the Green Algae Found Within the Egg Envelopes. Ecology 1942, 23, 215–227. [Google Scholar] [CrossRef] [Green Version]
- Hutchison, V.H.; Hammen, C.S. Oxygen Utilization in the Symbiosis of Embryos of the Salamander, Ambystoma maculatum and the Alga, Oophila amblystomatis. Biol. Bull. 1958, 115, 483–489. [Google Scholar] [CrossRef]
- Jurga, E.; Graham, L.; Bishop, C.D. Oophila Is Monophyletic within a Three-Taxon Eukaryotic Microbiome in Egg Masses of the Salamander Ambystoma maculatum. Symbiosis 2020, 81, 187–199. [Google Scholar] [CrossRef]
- Ibelings, B.W.; De Bruin, A.; Kagami, M.; Rijkeboer, M.; Brehm, M.; Donk, E.V. Host Parasite Interactions between Freshwater Phytoplankton and Chytrid Fungi (Chyridiomycota). J. Phycol. 2004, 40, 437–453. [Google Scholar] [CrossRef]
- Blaustein, A.R.; Belden, L.K. Amphibian Defenses against Ultraviolet-B Radiation. Evol. Dev. 2003, 5, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Cook, M.I.; Beissinger, S.R.; Toranzos, G.A.; Arendt, W.J. Incubation Reduces Microbial Growth on Eggshells and the Opportunity for Trans-Shell Infection. Ecol. Lett. 2005, 8, 532–537. [Google Scholar] [CrossRef]
- Godard, R.D.; Morgan Wilson, C.; Frick, J.W.; Siegel, P.B.; Bowers, B.B. The Effects of Exposure and Microbes on Hatchability of Eggs in Open-Cup and Cavity Nests. J. Avian Biol. 2007, 38, 709–716. [Google Scholar] [CrossRef]
- Van Veelen, H.P.J.; Salles, J.F.; Tieleman, B.I. Microbiome Assembly of Avian Eggshells and Their Potential as Transgenerational Carriers of Maternal Microbiota. ISME J. 2018, 12, 1375–1388. [Google Scholar] [CrossRef]
- Grond, K.; Lanctot, R.B.; Jumpponen, A.; Sandercock, B.K. Recruitment and Establishment of the Gut Microbiome in Arctic Shorebirds. FEMS Microbiol. Ecol. 2017, 93, fix142. [Google Scholar] [CrossRef]
- Wielen, P.W.J.J.; Keuzenkamp, D.A.; Lipman, L.J.A.; Knapen, F.; Biesterveld, S. Spatial and Temporal Variation of the Intestinal Bacterial Community in Commercially Raised Broiler Chickens During Growth. Microb. Ecol. 2002, 44, 286–293. [Google Scholar] [CrossRef]
- Soler, J.J.; Martín-Vivaldi, M.; Peralta-Sánchez, J.M.; Arco, L.; Juárez-García-Pelayo, N. Hoopoes Color Their Eggs with Antimicrobial Uropygial Secretions. Naturwissenschaften 2014, 101, 697–705. [Google Scholar] [CrossRef] [PubMed]
- Soler, J.J.; Martín-Vivaldi, M.; Ruiz-Rodríguez, M.; Valdivia, E.; Martín-Platero, A.M.; Martínez-Bueno, M.; Peralta-Sánchez, J.M.; Méndez, M. Symbiotic Association between Hoopoes and Antibiotic-Producing Bacteria That Live in Their Uropygial Gland. Funct. Ecol. 2008, 22, 864–871. [Google Scholar] [CrossRef] [Green Version]
- Martín-Vivaldi, M.; Soler, J.J.; Peralta-Sánchez, J.M.; Arco, L.; Martín-Platero, A.M.; Martínez-Bueno, M.; Ruiz-Rodríguez, M.; Valdivia, E. Special Structures of Hoopoe Eggshells Enhance the Adhesion of Symbiont-Carrying Uropygial Secretion That Increase Hatching Success. J. Anim. Ecol. 2014, 83, 1289–1301. [Google Scholar] [CrossRef] [PubMed]
- Martín-Platero, A.M.; Valdivia, E.; Ruíz-Rodríguez, M.; Soler, J.J.; Martín-Vivaldi, M.; Maqueda, M.; Martínez-Bueno, M. Characterization of Antimicrobial Substances Produced by Enterococcus faecalis MRR 10-3, Isolated from the Uropygial Gland of the Hoopoe (Upupa epops). Appl. Environ. Microbiol. 2006, 72, 4245–4249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Law-Brown, J.; Meyers, P.R. Enterococcus phoeniculicola Sp. Nov., a Novel Member of the Enterococci Isolated from the Uropygial Gland of the Red-Billed Woodhoopoe, Phoeniculus purpureus. Int. J. Syst. Evol. Microbiol. 2003, 53, 683–685. [Google Scholar] [CrossRef] [PubMed]
- Sarmiento-Ramírez, J.M.; van der Voort, M.; Raaijmakers, J.M.; Diéguez-Uribeondo, J. Unravelling the Microbiome of Eggs of the Endangered Sea Turtle Eretmochelys imbricata Identifies Bacteria with Activity against the Emerging Pathogen Fusarium falciforme. PLoS ONE 2014, 9, e95206. [Google Scholar] [CrossRef]
- Bézy, V.S.; Valverde, R.A.; Plante, C.J. Olive Ridley Sea Turtle Hatching Success as a Function of the Microbial Abundance in Nest Sand at Ostional, Costa Rica. PLoS ONE 2015, 10, e0118579. [Google Scholar] [CrossRef]
- Shine, R. Life-History Evolution in Reptiles. Annu. Rev. Ecol. Evol. Syst. 2005, 36, 23–46. [Google Scholar] [CrossRef] [Green Version]
- Kohl, K.D.; Brun, A.; Magallanes, M.; Brinkerhoff, J.; Laspiur, A.; Acosta, J.C.; Caviedes-Vidal, E.; Bordenstein, S.R. Gut Microbial Ecology of Lizards: Insights into Diversity in the Wild, Effects of Captivity, Variation across Gut Regions and Transmission. Mol. Ecol. 2017, 26, 1175–1189. [Google Scholar] [CrossRef]
- Yockey, L.J.; Lucas, C.; Iwasaki, A. Contributions of Maternal and Fetal Antiviral Immunity in Congenital Disease. Science 2020, 368, 608–612. [Google Scholar] [CrossRef]
- Feiner, N.; de Souza-Lima, S.; Jorge, F.; Naem, S.; Aubret, F.; Uller, T.; Nadler, S.A. Vertical Transmission of a Nematode from Female Lizards to the Brains of Their Offspring. Am. Nat. 2020, 195, 918–926. [Google Scholar] [CrossRef] [PubMed]
- Hubert, J. The embryology of the squamata. In Biology of Reptilia; Wiley: New York, NY, USA, 1985; pp. 1–34. [Google Scholar]
- Gantois, I.; Ducatelle, R.; Pasmans, F.; Haesebrouck, F.; Gast, R.; Humphrey, T.J.; Van Immerseel, F. Mechanisms of Egg Contamination by Salmonella enteritidis. FEMS Microbiol. Rev. 2009, 33, 718–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Reu, K.; Grijspeerdt, K.; Messens, W.; Heyndrickx, M.; Uyttendaele, M.; Debevere, J.; Herman, L. Eggshell Factors Influencing Eggshell Penetration and Whole Egg Contamination by Different Bacteria, Including Salmonella enteritidis. Int. J. Food Microbiol. 2006, 112, 253–260. [Google Scholar] [CrossRef]
- Cox, N.A.; Richardson, L.J.; Maurer, J.J.; Berrang, M.E.; Fedorka-Cray, P.J.; Buhr, R.J.; Byrd, J.A.; Lee, M.D.; Hofacre, C.L.; O’Kane, P.M.; et al. Evidence for Horizontal and Vertical Transmission in Campylobacter Passage from Hen to Her Progeny. J. Food Prot. 2012, 75, 1896–1902. [Google Scholar] [CrossRef]
- Salter, S.J.; Cox, M.J.; Turek, E.M.; Calus, S.T.; Cookson, W.O.; Moffatt, M.F.; Turner, P.; Parkhill, J.; Loman, N.J.; Walker, A.W. Reagent and Laboratory Contamination Can Critically Impact Sequence-Based Microbiome Analyses. BMC Biol. 2014, 12, 87. [Google Scholar] [CrossRef] [Green Version]
- Tyndale-Biscoe, H.; Renfree, M. Breeding biology of marsupials by family. In Reproductive Physiology of Marsupials; Cambridge University Press: Cambridge, UK, 1987; pp. 14–94. [Google Scholar]
- Yadav, M.; Stanley, N.F.; Waring, H. The Microbial Flora of the Gut of the Pouch-Young and the Pouch of a Marsupial, Setonix brachyurus. J. Gen. Microbiol. 1972, 70, 437–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deakin, J.E.; Cooper, D.W. Characterization of and Immunity to the Aerobic Bacteria Found in the Pouch of the Brushtail Possum Trichosurus vulpecula. Comp. Immunol. Microbiol. Infect. Dis. 2004, 27, 33–46. [Google Scholar] [CrossRef]
- Osawa, R.; Blanshard, W.H.; O’Callaghan, P.G. Microflora of the Pouch of the Koala (Phascolarctos cinereus). J. Wildl. Dis. 1992, 28, 276–280. [Google Scholar] [CrossRef]
- Weiss, S.; Taggart, D.; Smith, I.; Helgen, K.M.; Eisenhofer, R. Host Reproductive Cycle Influences the Pouch Microbiota of Wild Southern Hairy-Nosed Wombats (Lasiorhinus latifrons). Anim. Microbiome 2021, 3, 13. [Google Scholar]
- Chhour, K.-L.; Hinds, L.A.; Jacques, N.A.; Deane, E.M. An Observational Study of the Microbiome of the Maternal Pouch and Saliva of the Tammar Wallaby, Macropus Eugenii, and of the Gastrointestinal Tract of the Pouch Young. Microbiology 2010, 156, 798–808. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.; Fox, S.; Pemberton, D.; Hogg, C.; Papenfuss, A.T.; Belov, K. The Tasmanian Devil Microbiome—Implications for Conservation and Management. Microbiome 2015, 3, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capellini, I.; Nunn, C.L.; Barton, R.A. Microparasites and Placental Invasiveness in Eutherian Mammals. PLoS ONE 2015, 10, e0132563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malmuthuge, N.; Griebel, P.J. Fetal Environment and Fetal Intestine Are Sterile during the Third Trimester of Pregnancy. Vet. Immunol. Immunopathol. 2018, 204, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, E.; Fernández, L.; Marín, M.L.; Martín, R.; Odriozola, J.M.; Nueno-Palop, C.; Narbad, A.; Olivares, M.; Xaus, J.; Rodríguez, J.M. Isolation of Commensal Bacteria from Umbilical Cord Blood of Healthy Neonates Born by Cesarean Section. Curr. Microbiol. 2005, 51, 270–274. [Google Scholar] [CrossRef]
- Jiménez, E.; Marín, M.L.; Martín, R.; Odriozola, J.M.; Olivares, M.; Xaus, J.; Fernández, L.; Rodríguez, J.M. Is Meconium from Healthy Newborns Actually Sterile? Res. Microbiol. 2008, 159, 187–193. [Google Scholar] [CrossRef]
- Kustner, O. Beitrag Zur Lehre von Der Puerperalen Infection Der Neugeborenen. Arch Gynakol 1877, 11, 256–263. [Google Scholar] [CrossRef]
- Perez-Muñoz, M.E.; Arrieta, M.-C.; Ramer-Tait, A.E.; Walter, J. A Critical Assessment of the “Sterile Womb” and “in utero Colonization” Hypotheses: Implications for Research on the Pioneer Infant Microbiome. Microbiome 2017, 5, 1–19. [Google Scholar] [CrossRef]
- Funkhouser, L.J.; Bordenstein, S.R. Mom Knows Best: The Universality of Maternal Microbial Transmission. PLoS Biol. 2013, 11, e1001631. [Google Scholar] [CrossRef]
- Aagaard, K.; Ma, J.; Antony, K.M.; Ganu, R.; Petrosino, J.; Versalovic, J. The Placenta Harbors a Unique Microbiome. Sci. Transl. Med. 2014, 6, 237ra65. [Google Scholar] [CrossRef] [Green Version]
- Kliman, H.J. Comment on “The Placenta Harbors a Unique Microbiome”. Sci. Transl. Med. 2014, 6, 254le4. [Google Scholar] [CrossRef]
- Lauder, A.P.; Roche, A.M.; Sherrill-Mix, S.; Bailey, A.; Laughlin, A.L.; Bittinger, K.; Leite, R.; Elovitz, M.A.; Parry, S.; Bushman, F.D. Comparison of Placenta Samples with Contamination Controls Does Not Provide Evidence for a Distinct Placenta Microbiota. Microbiome 2016, 4, 29. [Google Scholar] [CrossRef] [Green Version]
- Antony, K.M.; Ma, J.; Mitchell, K.B.; Racusin, D.A.; Versalovic, J.; Aagaard, K. The Preterm Placental Microbiome Varies in Association with Excess Maternal Gestational Weight Gain. Am. J. Obstet. Gynecol. 2015, 212, 653.e1–653.e16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, J.; Xiao, X.; Zhang, Q.; Mao, L.; Yu, M.; Xu, J. The Placental Microbiome Varies in Association with Low Birth Weight in Full-Term Neonates. Nutrients 2015, 7, 6924–6937. [Google Scholar] [CrossRef] [PubMed]
- Amarasekara, R.; Jayasekara, R.W.; Senanayake, H.; Dissanayake, V.H.W. Microbiome of the Placenta in Pre-Eclampsia Supports the Role of Bacteria in the Multifactorial Cause of Pre-Eclampsia: Pre-Eclampsia and Infectious Agents. J. Obstet. Gynaecol. Res. 2015, 41, 662–669. [Google Scholar] [CrossRef] [PubMed]
- Bassols, J.; Serino, M.; Carreras-Badosa, G.; Burcelin, R.; Blasco-Baque, V.; Lopez-Bermejo, A.; Fernandez-Real, J.-M. Gestational Diabetes Is Associated with Changes in Placental Microbiota and Microbiome. Pediatr. Res. 2016, 80, 777–784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burrage, S. Bacteria in the Supposedly Sterile Meconium. J. Bacteriol. 1927, 13, 47–48. [Google Scholar]
- Hall, I.C. Bacterial Flora of First Specimens of Meconium Passed by Fifty New-Born Infants. Arch. Pediatr. Adolesc. Med. 1934, 47, 1279–1285. [Google Scholar] [CrossRef]
- Hansen, R.; Scott, K.P.; Khan, S.; Martin, J.C.; Berry, S.H.; Stevenson, M.; Okpapi, A.; Munro, M.J.; Hold, G.L. First-Pass Meconium Samples from Healthy Term Vaginally-Delivered Neonates: An Analysis of the Microbiota. PLoS ONE 2015, 10, e0133320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, J.W.; Brown, J.H. The Bacterial Content of the Uterus at Cesarean Section. Am. J. Obstet. Gynecol. 1927, 13, 133–143. [Google Scholar] [CrossRef]
- Stinson, L.F.; Keelan, J.A.; Payne, M.S. Identification and Removal of Contaminating Microbial DNA from PCR Reagents: Impact on Low-biomass Microbiome Analyses. Lett. Appl. Microbiol. 2019, 68, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Stinson, L.F.; Keelan, J.A.; Payne, M.S. Comparison of Meconium DNA Extraction Methods for Use in Microbiome Studies. Front. Microbiol. 2018, 9, 270. [Google Scholar] [CrossRef] [Green Version]
- Collado, M.C.; Segata, N. Initial Exploration of in utero Microbial Colonization. Nat. Med. 2020, 26, 469–470. [Google Scholar] [CrossRef]
- Rackaityte, E.; Halkias, J.; Fukui, E.M.; Mendoza, V.F.; Hayzelden, C.; Crawford, E.D.; Fujimura, K.E.; Burt, T.D.; Lynch, S.V. Viable Bacterial Colonization Is Highly Limited in the Human Intestine in utero. Nat. Med. 2020, 26, 599–607. [Google Scholar] [CrossRef] [PubMed]
- Park, J.-E.; Jardine, L.; Gottgens, B.; Teichmann, S.A.; Haniffa, M. Prenatal Development of Human Immunity. Science 2020, 368, 600–603. [Google Scholar] [CrossRef] [PubMed]
- Ganal-Vonarburg, S.C.; Hornef, M.W.; Macpherson, A.J. Microbial–Host Molecular Exchange and Its Functional Consequences in Early Mammalian Life. Science 2020, 368, 604–607. [Google Scholar] [CrossRef]
- Uchimura, Y.; Fuhrer, T.; Li, H.; Lawson, M.A.; Zimmermann, M.; Yilmaz, B.; Zindel, J.; Ronchi, F.; Sorribas, M.; Hapfelmeier, S.; et al. Antibodies Set Boundaries Limiting Microbial Metabolite Penetration and the Resultant Mammalian Host Response. Immunity 2018, 49, 545–559.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, I.; Miyamoto, J.; Ohue-Kitano, R.; Watanabe, K.; Yamada, T.; Onuki, M.; Aoki, R.; Isobe, Y.; Kashihara, D.; Inoue, D.; et al. Maternal Gut Microbiota in Pregnancy Influences Offspring Metabolic Phenotype in Mice. Science 2020, 367, eaaw8429. [Google Scholar] [CrossRef] [PubMed]
- Pasteur, L. Observations Relatives à La Note Précédente de M. Comptes Rendus Seances Acad. Sci. 1885, 100, 68. [Google Scholar]
- Teah, B.A. Bibliography of Germfree Research 1885–1963; Supplement 1977; Defense Technical Information Center: Fort Belvoir, VA, USA, 1977. [Google Scholar]
- Gordon, H.A. The Germ-Free Animal. Its Use in the Study of “Physiologic” Effects of the Normal Microbial Flora on the Animal Host. Am. J. Dig. Dis. 1960, 5, 841–867. [Google Scholar] [CrossRef] [PubMed]
- Gordon, H.A.; Pesti, L. The Gnotobiotic Animal as a Tool in the Study of Host Microbial Relationships. Bacteriol. Rev. 1971, 35, 390–429. [Google Scholar] [CrossRef] [PubMed]
- Timmons, E.H.; Olmsted, G.M.; Kaplan, H.M. The Germfree Leopard Frog (Rana pipiens): Preliminary Report. Lab. Anim. Sci. 1977, 27, 518–521. [Google Scholar] [PubMed]
- Hedrich, H.J.; Hardy, P. The Laboratory Mouse; Elsevier: London, UK, 2012. [Google Scholar]
- Melancon, E.; De La Torre Canny, S.G.; Sichel, S.; Kelly, M.; Wiles, T.J.; Rawls, J.F.; Eisen, J.S.; Guillemin, K. Best Practices for Germ-Free Derivation and Gnotobiotic Zebrafish Husbandry. Methods Cell Biol. 2017, 138, 61–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chinn, I.K.; Shearer, W.T. Severe Combined Immunodeficiency Disorders. Immunol. Allergy Clin. N. Am. 2015, 35, 671–694. [Google Scholar] [CrossRef] [PubMed]
- Nokhbatolfoghahai, M.; Downie, J.R. Ciliated Cells on the Surface of Embryos of Scyliorhinus canicula. J. Fish Biol. 2003, 63, 523–527. [Google Scholar] [CrossRef]
- Nokhbatolfoghahai, M.; Downie, J.R.; Clelland, A.K.; Rennison, K. The Surface Ciliation of Anuran Amphibian Embryos and Early Larvae: Patterns, Timing Differences and Functions. J. Nat. Hist. 2005, 39, 887–929. [Google Scholar] [CrossRef]
- Nokhbatolfoghahai, M.; Downie, J.R.; Ogilvy, V. Surface Ciliation of Anuran Amphibian Larvae: Persistence to Late Stages in Some Species but Not Others. J. Morphol. 2006, 267, 1248–1256. [Google Scholar] [CrossRef]
- Kessel, R.G.; Beams, H.W.; Shih, C.Y. The Origin, Distribution and Disappearance of Surface Cilia during Embryonic Development of Rana pipiens as Revealed by Scanning Electron Microscopy. Am. J. Anat. 1974, 141, 341–359. [Google Scholar] [CrossRef] [PubMed]
- Morgan, T.H. The Development of the Frog’s Egg; Macmillan and Co.: New York, NY, USA, 1897. [Google Scholar]
- Dünker, N.; Wake, M.H.; Olson, W.M. Embryonic and Larval Development in the Caecilian Ichthyophis Kohtaoensis (Amphibia, Gymnophiona): A Staging Table. J. Morphol. 2000, 243, 3–34. [Google Scholar] [CrossRef]
- Fox, H. Early Development of Caecilian Skin with Special Reference to the Epidermis. J. Herpetol. 1986, 20, 154–167. [Google Scholar] [CrossRef]
- Harrison, R.G. Harrison Stages and Description of the Normal Development of the Spotted Salamander, Amblystoma punctatum. In Organization and Development of the Embryo; Yale University Press: New Haven, CT, USA, 1969; pp. 44–66. [Google Scholar]
- Landström, U. On the Differentiation of Prospective Ectoderm to a Ciliated Cell Pattern in Embryos of Ambystoma mexicanum. J. Embryol. Exp. Morphol. 1977, 41, 23–32. [Google Scholar]
- Tattersall, G.J.; Spiegelaar, N. Embryonic Motility and Hatching Success of Ambystoma maculatum Are Influenced by a Symbiotic Alga. Can. J. Zool. 2008, 86, 1289–1298. [Google Scholar] [CrossRef] [Green Version]
- Lannoo, M.J. Neuromast Topography in Anuran Amphibians. J. Morphol. 1987, 191, 115–129. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, J.I.; Doran, S.A.; Shartau, R.B.; Pon, J.R.; Ali, D.W.; Tam, R.; Kuang, S. Integrative Biology of an Embryonic Respiratory Behaviour in Pond Snails: The ‘embryo Stir-Bar Hypothesis’. J. Exp. Biol. 2008, 211, 1729–1736. [Google Scholar] [CrossRef] [Green Version]
- Wiley, A. Amphioxous and the Ancestry of Vertebrates; Macmillan: New York, NY, USA, 1894. [Google Scholar]
- Webb, J.F.; Northcutt, R.G. Ciliated Epidermal Cells in Non-Teleost Actinopterygian Fish. Acta Zool. 1991, 72, 107–111. [Google Scholar] [CrossRef]
- Bartsch, P.; Gemballa, S.; Piotrowski, T. The Embryonic and Larval Development of Polypterus senegalus Cuvier, 1829: Its Staging with Reference to External and Skeletal Features, Behaviour and Locomotory Habits. Acta Zool. 1997, 78, 309–328. [Google Scholar] [CrossRef]
- Song, J. The Lateral Line System in the Florida Gar, Lepisosteus platyrhincus, de Kay. Ph.D. Thesis, University of Michigan, Ann Arbor, MI, USA, 1989. [Google Scholar]
- Song, J.; Northcutt, R.G. Morphology, Distribution and Innervation of the Lateral-Line Receptors of the Florida Gar, Lepiosteus platyrhinchus. Brain. Behav. Evol. 1991, 37, 10–37. [Google Scholar] [CrossRef]
- Bemis, W.E.; Grande, L. Early Development of the Actinopterygian Head. I. External Development and Staging of the Paddlefish Polyodon spathula. J. Morphol. 1992, 213, 47–83. [Google Scholar] [CrossRef] [PubMed]
- Brooks, E.R.; Wallingford, J.B. Multiciliated Cells. Curr. Biol. 2014, 24, R973–R982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klos Dehring, D.A.; Vladar, E.K.; Werner, M.E.; Mitchell, J.W.; Hwang, P.; Mitchell, B.J. Deuterosome-Mediated Centriole Biogenesis. Dev. Cell 2013, 27, 103–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whiting, H.P.; Bone, Q. Ciliary Cells in the Epidermis of the Larval Australian Dipnoan, Neoceratodus. Zool. J. Linn. Soc. 1980, 68, 125–137. [Google Scholar] [CrossRef]
- Lauder, G.V.; Liem, K.F. The Evolution and Interrelationships of the Actinopterygian Fishes. Bull. Mus. Comp. Zool. Harv. Coll. 1983, 150, 95–197. [Google Scholar]
- Balfour, F.M. Chapter VII—Amphibia. In A Treatise on Comparative Embryology; MacMillan: London, UK, 1881; pp. 99–119. [Google Scholar]
- Walentek, P.; Quigley, I.K. What We Can Learn from a Tadpole about Ciliopathies and Airway Diseases: Using Systems Biology in Xenopus to Study Cilia and Mucociliary Epithelia. Genesis 2017, 55, e23001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chuyen, A.; Rulquin, C.; Daian, F.; Thomé, V.; Clément, R.; Kodjabachian, L.; Pasini, A. The Scf/Kit Pathway Implements Self-Organized Epithelial Patterning. Dev. Cell 2021, 56, 795–810.e7. [Google Scholar] [CrossRef] [PubMed]
- Marcet, B.; Chevalier, B.; Luxardi, G.; Coraux, C.; Zaragosi, L.-E.; Cibois, M.; Robbe-Sermesant, K.; Jolly, T.; Cardinaud, B.; Moreilhon, C.; et al. Control of Vertebrate Multiciliogenesis by MiR-449 through Direct Repression of the Delta/Notch Pathway. Nat. Cell Biol. 2011, 13, 693–699. [Google Scholar] [CrossRef]
- Stubbs, J.L.; Vladar, E.K.; Axelrod, J.D.; Kintner, C. Multicilin Promotes Centriole Assembly and Ciliogenesis during Multiciliate Cell Differentiation. Nat. Cell Biol. 2012, 14, 140–147. [Google Scholar] [CrossRef] [Green Version]
- Dubaissi, E.; Rousseau, K.; Lea, R.; Soto, X.; Nardeosingh, S.; Schweickert, A.; Amaya, E.; Thornton, D.J.; Papalopulu, N. A Secretory Cell Type Develops alongside Multiciliated Cells, Ionocytes and Goblet Cells, and Provides a Protective, Anti-Infective Function in the Frog Embryonic Mucociliary Epidermis. Dev. Camb. Engl. 2014, 141, 1514–1525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elgeti, J.; Gompper, G. Emergence of Metachronal Waves in Cilia Arrays. Proc. Natl. Acad. Sci. USA 2013, 110, 4470–4475. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, B.; Stubbs, J.L.; Huisman, F.; Taborek, P.; Yu, C.; Kintner, C. The PCP Pathway Instructs the Planar Orientation of Ciliated Cells in the Xenopus Larval Skin. Curr. Biol. 2009, 19, 924–929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Assheton, M.A. Notes on the Ciliation of the Ectoderm of the Amphibian Embryo. Q. J. Microsc. Sci. 1896, 38, 465–484. [Google Scholar]
- Burggren, W.W. Gas, Exchange, Metabolism, and “Ventilation”; in Gelatinous Frog Egg Masses. Physiol. Zool. 1985, 58, 503–514. [Google Scholar] [CrossRef]
- Feder, M.E.; Burggren, W.W. Cutaneous Gas Exchange in Vertebrates: Design, Patterns, Control and Implications. Biol. Rev. 1985, 60, 1–45. [Google Scholar] [CrossRef] [PubMed]
- Seymour, R.S.; Bradford, D.F. Gas Exchange through the Jelly Capsule of the Terrestrial Eggs of the Frog, Pseudophryne bibroni. J. Comp. Physiol. B 1987, 157, 477–481. [Google Scholar] [CrossRef]
- Twitty, V.C. Experimental Studies on the Ciliary Action of Amphibian Embryos. J. Exp. Zool. 1928, 50, 319–344. [Google Scholar] [CrossRef]
- Cartwright, J.H.E.; Piro, O.; Tuval, I. Fluid Dynamics in Developmental Biology: Moving Fluids That Shape Ontogeny. HFSP J. 2009, 3, 77–93. [Google Scholar] [CrossRef] [Green Version]
- Ciuhandu, C.S.; Wright, P.A.; Goldberg, J.I.; Stevens, E.D. Parameters Influencing the Dissolved Oxygen in the Boundary Layer of Rainbow Trout (Oncorhynchus mykiss) Embryos and Larvae. J. Exp. Biol. 2007, 210, 1435–1445. [Google Scholar] [CrossRef] [Green Version]
- Liem, K. Larvae of Air-Breathing Fishes as Countercurrent Flow Devices in Hypoxic Environments. Science 1981, 211, 1177–1179. [Google Scholar] [CrossRef] [PubMed]
- Lanzing, W.J.R. A Temporary Respiratory Organ in the Tail of Tilapia mossambica Fry. Copeia 1976, 1976, 800–802. [Google Scholar] [CrossRef]
- Secchi, E.; Vitale, A.; Miño, G.L.; Kantsler, V.; Eberl, L.; Rusconi, R.; Stocker, R. The Effect of Flow on Swimming Bacteria Controls the Initial Colonization of Curved Surfaces. Nat. Commun. 2020, 11, 2851. [Google Scholar] [CrossRef] [PubMed]
- Thurber, A.R.; Jones, W.J.; Schnabel, K. Dancing for Food in the Deep Sea: Bacterial Farming by a New Species of Yeti Crab. PLoS ONE 2011, 6, e26243. [Google Scholar] [CrossRef]
- Nawroth, J.C.; van der Does, A.M.; Ryan (Firth), A.; Kanso, E. Multiscale Mechanics of Mucociliary Clearance in the Lung. Philos. Trans. R. Soc. B Biol. Sci. 2020, 375, 20190160. [Google Scholar] [CrossRef]
- Erb-Downward, J.R.; Thompson, D.L.; Han, M.K.; Freeman, C.M.; McCloskey, L.; Schmidt, L.A.; Young, V.B.; Toews, G.B.; Curtis, J.L.; Sundaram, B.; et al. Analysis of the Lung Microbiome in the “Healthy” Smoker and in COPD. PLoS ONE 2011, 6, e16384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nawroth, J.C.; Guo, H.; Koch, E.; Heath-Heckman, E.A.C.; Hermanson, J.C.; Ruby, E.G.; Dabiri, J.O.; Kanso, E.; McFall-Ngai, M. Motile Cilia Create Fluid-Mechanical Microhabitats for the Active Recruitment of the Host Microbiome. Proc. Natl. Acad. Sci. USA 2017, 114, 9510–9516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murillo-Rincon, A.P.; Klimovich, A.; Pemöller, E.; Taubenheim, J.; Mortzfeld, B.; Augustin, R.; Bosch, T.C.G. Spontaneous Body Contractions Are Modulated by the Microbiome of Hydra. Sci. Rep. 2017, 7, 15937. [Google Scholar] [CrossRef]
- Gilbert, S.F.; Bosch, T.C.G.; Ledón-Rettig, C. Eco-Evo-Devo: Developmental Symbiosis and Developmental Plasticity as Evolutionary Agents. Nat. Rev. Genet. 2015, 16, 611–622. [Google Scholar] [CrossRef]
- Pennisi, E. Mysteries of Development. How Do Microbes Shape Animal Development? Science 2013, 340, 1159–1160. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Vertebrate extraembryonic fluid microenvironments (shaded) and extraembryonic tissues which may harbor embryo-associated microbial life. (A) All vertebrates begin within a maternally-derived zona pellucida, also known as the vitelline membrane. After fertilization, perivitelline fluid surrounds the zygote and early embryo. The in ovo fluid compartment of externally developing fish (B) and amphibians (C) contains extraembryonic capsular fluid or “perivitelline fluid”, although the vitelline membrane usually ruptures during neurulation. Maternally-derived egg capsules (“chorion” in fish) surround the developing embryos and can vary in number. (D) Oviparous amniotes also develop in ovo inside a post-zygotic maternally-derived shell and variable cuticle. The embryo proper develops within the amniotic fluid (shaded). The extraembryonic chorion surrounds yolk, allantois, and amnion, all of which could potentially harbor embryo-associated microbes. (E) These membranes are also part of the in utero microenvironment of developing placental mammals, which also extend fetal circulation to interface maternal circulation in the placenta. Panels D and E are redrawn with permission from Barresi and Gilbert, Developmental Biology, 11th edition. Published by Oxford University Press (2016).
Figure 1.
Vertebrate extraembryonic fluid microenvironments (shaded) and extraembryonic tissues which may harbor embryo-associated microbial life. (A) All vertebrates begin within a maternally-derived zona pellucida, also known as the vitelline membrane. After fertilization, perivitelline fluid surrounds the zygote and early embryo. The in ovo fluid compartment of externally developing fish (B) and amphibians (C) contains extraembryonic capsular fluid or “perivitelline fluid”, although the vitelline membrane usually ruptures during neurulation. Maternally-derived egg capsules (“chorion” in fish) surround the developing embryos and can vary in number. (D) Oviparous amniotes also develop in ovo inside a post-zygotic maternally-derived shell and variable cuticle. The embryo proper develops within the amniotic fluid (shaded). The extraembryonic chorion surrounds yolk, allantois, and amnion, all of which could potentially harbor embryo-associated microbes. (E) These membranes are also part of the in utero microenvironment of developing placental mammals, which also extend fetal circulation to interface maternal circulation in the placenta. Panels D and E are redrawn with permission from Barresi and Gilbert, Developmental Biology, 11th edition. Published by Oxford University Press (2016).

Figure 2.
Venn diagram of bacterial phyla identified inside in ovo and in utero microenvironments of representative vertebrates through 16s rDNA metabarcoding [32,33,34,35]. Whitefish (Corregonus spp.) perivitelline fluid sampling was followed by amplification of the V1–V2 16s regions and taxonomy classification with RDP2.2 and Greengenes 12.10 [32]. Spotted salamander (Ambystoma maculatum) egg capsule fluid sampling was followed by V4–V5 amplification and classification with SILVA132 [33]. Rat (Rattus norvegicus domestica) amniotic fluid extraction was followed by V3–V4 amplification and Greengenes 13.8 classification [34]. Eastern fence lizard (Sceloporus undulatus) and four passerine (Poecile atricapillus, Sialia sialis, Passer domesticus, and Tachycineta bicolor) in ovo contents were extracted followed by V4 amplification and Greengenes 13.8 classification [35]. Other studies have described teleost fish, bird and rodent embryonic microenvironments to be sterile [36,37,38]. “Cyanobacteria” may represent free living prokaryotes or chloroplast DNA. Identification of conserved embryo-associated phyla is preliminary as methodologies, including exclusion of negative control sequences, were inconsistent between studies. Bacterial communities from the rat study were verified through FISH staining in fetal gut [34] and the salamander study cultured multiple bacterial isolates from the same capsular fluid [33].
Figure 2.
Venn diagram of bacterial phyla identified inside in ovo and in utero microenvironments of representative vertebrates through 16s rDNA metabarcoding [32,33,34,35]. Whitefish (Corregonus spp.) perivitelline fluid sampling was followed by amplification of the V1–V2 16s regions and taxonomy classification with RDP2.2 and Greengenes 12.10 [32]. Spotted salamander (Ambystoma maculatum) egg capsule fluid sampling was followed by V4–V5 amplification and classification with SILVA132 [33]. Rat (Rattus norvegicus domestica) amniotic fluid extraction was followed by V3–V4 amplification and Greengenes 13.8 classification [34]. Eastern fence lizard (Sceloporus undulatus) and four passerine (Poecile atricapillus, Sialia sialis, Passer domesticus, and Tachycineta bicolor) in ovo contents were extracted followed by V4 amplification and Greengenes 13.8 classification [35]. Other studies have described teleost fish, bird and rodent embryonic microenvironments to be sterile [36,37,38]. “Cyanobacteria” may represent free living prokaryotes or chloroplast DNA. Identification of conserved embryo-associated phyla is preliminary as methodologies, including exclusion of negative control sequences, were inconsistent between studies. Bacterial communities from the rat study were verified through FISH staining in fetal gut [34] and the salamander study cultured multiple bacterial isolates from the same capsular fluid [33].

Figure 3.
Multiciliated surface cells (MSC) in amphibian embryos. (A,B) Scanning electron micrographs of the eastern redbacked salamander, Plethodon cinereus. (A) external gills 350× magnification; and (B) white box in A, single MSC with tufts of external cilia (500×). (C) Antero-posterior microcurrents created around woodfrog (Rana sylvatica); and (D) Vectors represent detectable particle displacement over two seconds in a timelapse of wild-collected R. sylvatica. MSCs create vortices within the closed environment of the egg capsule that move both the embryo and its extraembryonic fluid. Panel C is redrawn from Assheton [174], reproduced with permission from the Quarterly Journal of Microscopical Sciences (copyright now under The Company of Biologists), (1896).
Figure 3.
Multiciliated surface cells (MSC) in amphibian embryos. (A,B) Scanning electron micrographs of the eastern redbacked salamander, Plethodon cinereus. (A) external gills 350× magnification; and (B) white box in A, single MSC with tufts of external cilia (500×). (C) Antero-posterior microcurrents created around woodfrog (Rana sylvatica); and (D) Vectors represent detectable particle displacement over two seconds in a timelapse of wild-collected R. sylvatica. MSCs create vortices within the closed environment of the egg capsule that move both the embryo and its extraembryonic fluid. Panel C is redrawn from Assheton [174], reproduced with permission from the Quarterly Journal of Microscopical Sciences (copyright now under The Company of Biologists), (1896).

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kerney, R. Developing Inside a Layer of Germs—A Potential Role for Multiciliated Surface Cells in Vertebrate Embryos. Diversity 2021, 13, 527. https://doi.org/10.3390/d13110527
AMA Style
Kerney R. Developing Inside a Layer of Germs—A Potential Role for Multiciliated Surface Cells in Vertebrate Embryos. Diversity. 2021; 13(11):527. https://doi.org/10.3390/d13110527
Chicago/Turabian StyleKerney, Ryan. 2021. "Developing Inside a Layer of Germs—A Potential Role for Multiciliated Surface Cells in Vertebrate Embryos" Diversity 13, no. 11: 527. https://doi.org/10.3390/d13110527
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.