Molecular Diversity of Tidal Swamp Rice (Oryza sativa L.) in South Kalimantan, Indonesia


1
Faculty of Mathematics and Natural Sciences, Universitas Lambung Mangkurat, South Kalimantan 70714, Indonesia
2
Faculty of Biology, Universitas Gadjah Mada, Yogyakarta 55281, Indonesia
*
Author to whom correspondence should be addressed.
Diversity 2018, 10(2), 22; https://doi.org/10.3390/d10020022
Submission received: 15 March 2018
/
Revised: 1 April 2018
/
Accepted: 2 April 2018
/
Published: 9 April 2018
Abstract
:Tidal swamp rice has long been cultivated by the local people of the South Kalimantan, Indonesia. This germplasm possess some important traits for adapted to a wide range of abiotic and biotic stresses. In this study, a total of 16 cultivars of the tidal swamp rice, consisting of 15 from the South Kalimantan Provinces and 1 from South Sumatera, Indonesia (an outgroup) were analyzed phylogenetically based on the chloroplast trnL-F and nuclear intergenic spacer region (IGS). The results showed that this germplasm has a relatively more extraordinary genetic diversity than other local rice. On a nucleotide level, the tidal swamp rice showed a genetic diversity of 0.61 for nuclear IGS and 0.58 for trnL-F. The phylogenetic reconstruction also exhibited that the tidal swamp rice has the unique phylogenetic trees, particularly for the combined sequence datasets. This information would be useful for the rice conservation and breeding programs in the future.
1. Introduction
Tidal swamps are a potential area to be developed for future agricultural activity in many Asian countries with a total of 20.1 million hectares still available [1]. While such regions are considered the marginal areas, primarily due to natural problems, they support an excessive amount of indigenous rice cultivars [1]. Hundreds of the tidal swamp rice cultivars have been found in the region, and they show some beneficial agronomic traits. As a result of the domestication process and through long-term natural selection, this germplasm shows better adaptation to the local conditions and cultures [2]. Tidal swamp rice also exhibited both a tremendous genetic diversity and substantial genes for promoting the development of new rice cultivars in the future [3,4,5].
In South Kalimantan for example, one of the biggest rice producing regions in Indonesia, there are about 70 tidal swamp rice cultivars which are still found and preserved by the local farmers [6]. Some of them show important traits for the future breeding, such as Siam Pandak which has a high level of Fe content [6]. Nevertheless, the genetics of these germplasms have not yet been adequately studied and incorporated optimally into the rice breeding programs [7,8]. In general, characterization of rice germplasm has been undertaken based on morphological markers, such as grain shape and color, as well as the plant height and culm diameter [9,10]. However, these markers were strongly influenced by the environmental factors [11].
The molecular marker application may contribute in-depth information on the potential and genetic diversity of the rice germplasm [12]. These markers are more effective, efficient, accurate, and stable to environmental changes than morphological markers [11]. DNA sequencing is a part of molecular applications that constitutes a substantial contribution to the study of phylogenetics, in plants with both distant and close relationships [13,14]. In this case, the chloroplast trnL-F genes and nuclear intergenic spacer (IGS) region sequences could be used for that purpose [15,16]. It is because both have an elevated mutation rate, as well as the genetic diversity [17,18,19,20]. Some plant germplasms have been revealed by these markers, such as Brassica [21], Oryza [14,22], tomato [23], and Annonaceae [24].
This study was directed to investigate and determine the genetic diversity and relationship of the tidal swamp rice of the South Kalimantan Province, Indonesia using the chloroplast trnL-F and nuclear IGS sequences. In a local scale, such studies could reveal the complex interaction between the germplasm and farmers cultivation practices in good view [25]. Hence, the results of this study are expected to be used as a reference nationally, even internationally, in the genetic improvement and conservation programs of rice in the future.
2. Materials and Methods
2.1. Plant Materials
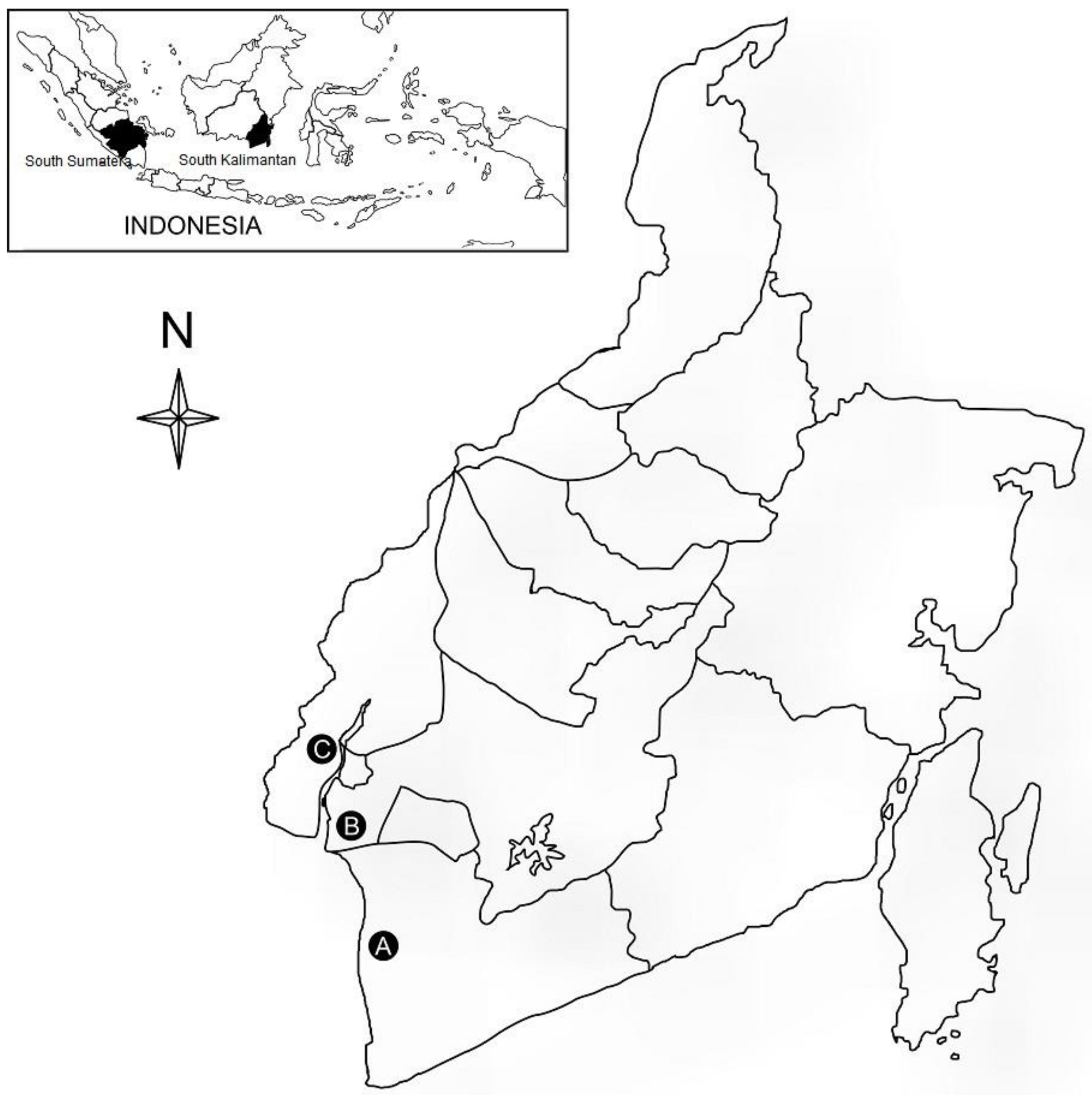
A total of 16 cultivars of tidal swamp rice, consisting of 15 from South Kalimantan and 1 from South Sumatera, Indonesia (an outgroup) were sampled for this study (Table 1, Figure 1). An outgroup cultivar was obtained from the Indonesian Agriculture of the Tidal Swamp Research Institute, South Kalimantan, Indonesia. Seeds of different cultivars were germinated after being soaked in a fungicide solution and rinsed thoroughly in warm water (30–35 °C). Seeds then were transplanted into pots, 30 cm in diameter and 20 cm in height, with soil collected from a paddy field and placed in a greenhouse. Leaf samples of, at least, three individual seedlings of each rice cultivar were collected for DNA assay.
2.2. DNA Isolation, Amplification, and Sequencing
Genomic DNAs were isolated using three-weeks-old of each rice leaf sample, following the Mini Kit of DNA Extraction protocol (Geneaid, New Taipei City, Taiwan). The isolated DNAs were measured using UV–vis spectrophotometer (NanoVue, GE Healthcare, Little Chalfont, UK), at wavelengths of 260 nm and 280 nm [26]. The isolated DNA sequences were then amplified using the chloroplast trnL-F and nuclear IGS primers (Table 2). The PCR was carried out at a total volume of 25 μL, consisted of 1.25 μL of 10 ng DNA template, 4.75 μL of the PCR grade water, 1.25 μL each primer (0.5 μM), and 16.50 μL master mix PCR (Kapa Biosystems). This master mix PCR contains 50 mmol KCl, 10 mmol Tris-HCl pH 8.8, 0.1% Triton-X, 1.5 mmol MgCl2, 200 μmol dNTP, and 0.5 U Taq DNA polymerase (Kapa Biosystems, Wilmington, MA, USA). All amplification reactions were performed in a PCR thermocycler (Biorad T100, Hercules, CA, USA) for 35 cycles, and programmed with the following conditions [14]: (1) An initial denaturation for 3 min at 95 °C; (2) denaturation for 30 s at 95 °C, (3) annealing for 30 s at 55 °C for trnL-F and 53 °C for IGS; (4) extension for 1 min at 72 °C; and (5) final extension for 5 min at 72 °C. PCR products were separated on 1.5% agarose gels. The amplified DNA fragments which corresponding to the expected size (Table 2) were then collected from the gels, purified by the QIAquick gel extraction kit (Qiagen, Germantown, MD, USA) and sequenced using Sanger dideoxy sequencing methods (Integrated DNA Technologies, Singapore). All sequences can be accessed in GenBank database with the accession numbers of MH150976 to MH151008.
2.3. Data Analysis
The DNA sequences from the two datasets were separately aligned using the Clustal X version 2.0 [28] and refined manually. Indels (insertions or deletions) were introduced into the alignment coded in the following ways. Shared indels were treated as single characters. Indels of uniform length were coded as absence (1) or presence (0) characters independent of the indel length. The gapped regions in the alignment were excluded from subsequent analysis unless some positions included nucleotide diversity. Gaps were treated as missing entries [29]. Maximum likelihood (ML) analysis was performed by heuristic search with nearest neighbor interchange (NNI) options, strong branch swap filter, and initial tree automatically (maximum parsimony). The sequence data were also analyzed with a neighbor-joining method using the Kimura two-parameter distance estimates [30]. Topological robustness was assessed by bootstrap analysis with 1000 replicates using simple taxon addition [31]. The characters were weighted equally in all phylogenetic analyses. The phylogenetic analyses were conducted by a software of MEGA7 [32].
3. Results
3.1. Sequence Characteristics and Genetic Diversity
Approximately 0.89 kilobase pairs (kb) of two sets of DNA fragments from the chloroplast trnL-F and nuclear IGS genomes were sequenced separately from 16 tidal swamp rice cultivars. The phylogenetic information was generated by both sequences given in Table 3. In general, the aligned sequences of nuclear IGS indicate a more extraordinary value of the number of segregating sites, Bayesian information criterion (BIC), and Akaike information criterion (AICc) than chloroplast trnL-F. Similarly, GC content and nucleotide diversity were higher in IGS sequences than the chloroplast trnL-F. However, both sequences have a similar value of Ti/Tv ratio (1.13) (see Table 3). Moreover, the Tajima’s test of neutrality shows that these sequences have a progressive value of D greater than zero (D > 0).
3.2. Phylogenetic Relationship Based on the Chloroplast trnL-F Spacer
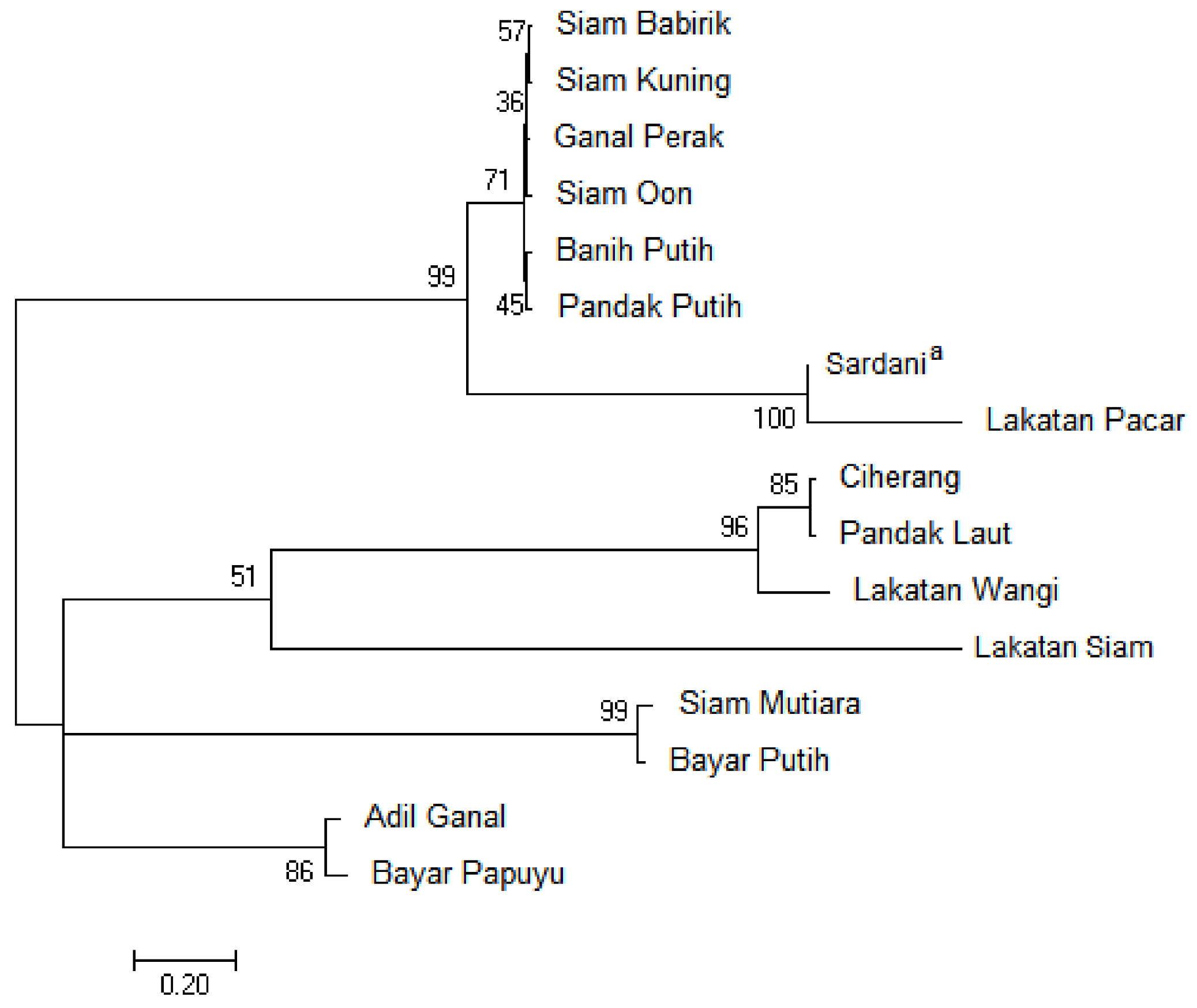
The phylogenetic tree was obtained for the chloroplast trnL-F sequences from the tidal swamp rice (Figure 2). In general, four separate groups with relatively high values of bootstrap support (bs) formed on this tree. Based on this tree, the Lakatan Pacar was closely related to Sardani, an outgroup, with a bootstrap value of 100%. Similarly, the Pandak Laut was closely related to Ciherang, an improved cultivar, at bs = 85%. However, the Sardani has far related to Pandak Putih (bs = 99%).
3.3. Phylogenetic Relationship Based on the Nuclear IGS Region
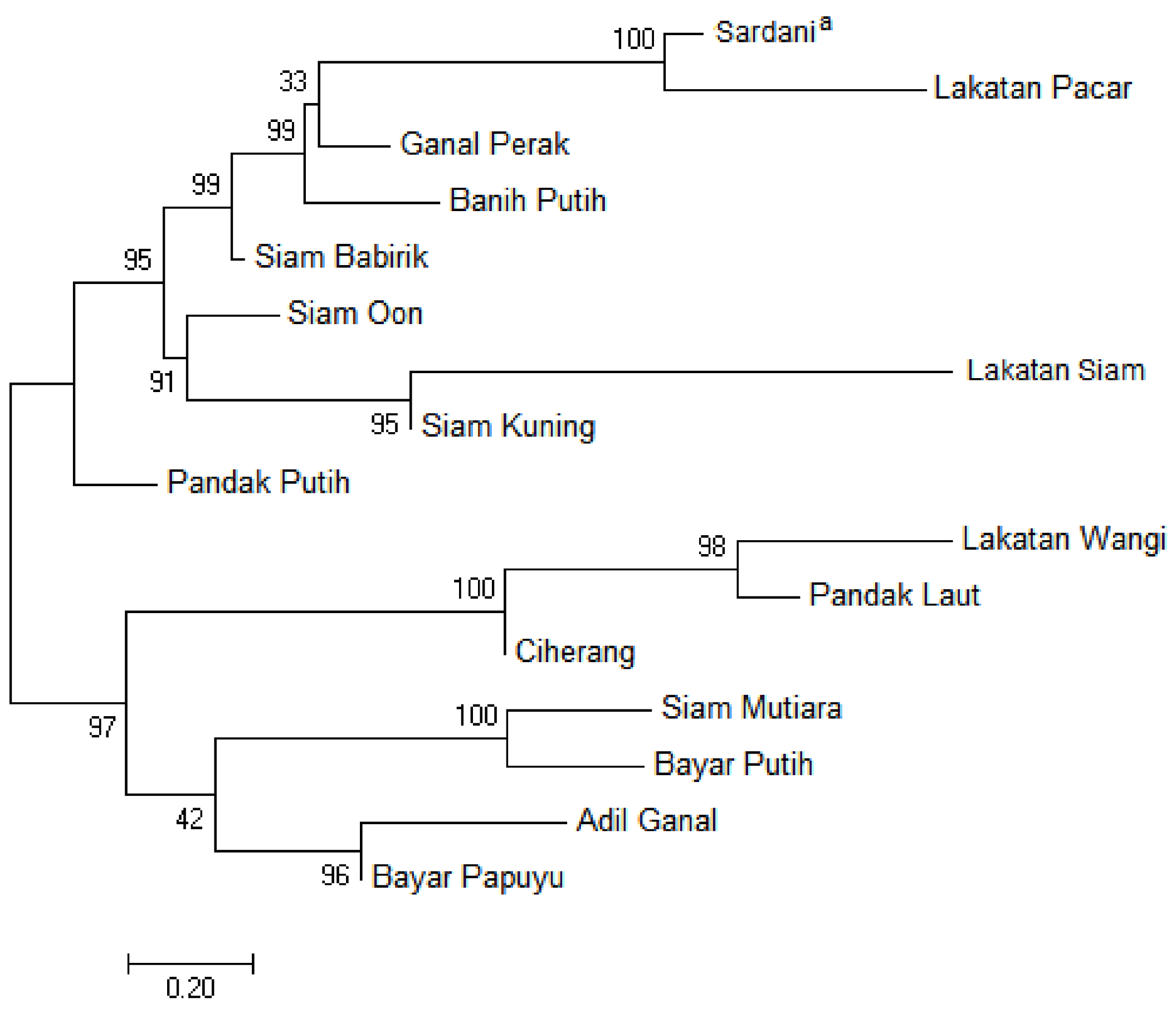
The phylogenetic relationship was obtained for the IGS sequences from the tidal swamp rice. The IGS tree was shown in Figure 3. In general, two independent groups of these germplasms were resolved by the IGS sequences. Differ from the trnL-F, the IGS was able to combine the Sardani with three tidal swamp rice of the South Kalimantan, such as Lakatan Pacar, Banih Putih, and Ganal Perak, at a bootstrap value of 96%. Whereas the Ciherang clustered with Pandak Putih, not with Pandak Laut, in a similar group (bs = 100%) as well. In this tree, the distant relationship is shown by Lakatan Pacar and Sardani (Figure 3).
3.4. Phylogenetic Relationship of Combined Chloroplast and Nuclear Datasets
One phylogenetic tree was reconstructed based on the combination of two individual sequencing datasets (Figure 4). This combined dataset produced many changes in a phylogenetic tree of the tidal swamp rice, including their bootstrap values. In general, two independent groups formed on this tree, and relatively similar to the topology of the IGS phylogenetic tree. In this case, Lakatan Pacar has closely related to Sardani at 100% bootstrap support, whereas Ciherang to Lakatan Wangi and Pandak Laut (bs = 100%). Based on this tree, Sardani and Ganal Perak were two cultivars with the most distant relationship.
4. Discussion
Genetic diversity was manifested by differences in many characters, such as morphology, anatomy, and in the proteins; enzymes; and DNA sequences of almost all organisms [33]. It was required for populations to adapt the environmental changes [33]. In our results, the aligned sequences of nuclear IGS show a relatively higher genetic diversity than chloroplast trnL-F, indicated both by nucleotide diversity and the number of segregating sites (Table 1). Moreover, this region strongly influenced by some mutations, like deletions and substitutions, as reported by Cordesse et al. [27]. In the Oryza genus, the IGS shows a high genetic diversity as well [27]. According to Chung et al. [34], nuclear IGS have been changing quickly, both in the number of copies and its chromosome segments. Furthermore, Chung et al. [34] reported that this region on some chromosomes of 10 genera of Oryza has an excessive number of repeated elements, located at the end of the short arm of chromosome 9 [34].
Based on the result, while the chloroplast trnL-F has relatively lower genetic diversity than IGS, it would be interesting for future studies [20]. Drábková et al. [35] reported that trnL-F has a higher substitution rate than an insertion–deletion (indel), making it a valuable tool for studying short-term evolutionary processes. Furthermore, Bayer and Starr [36] confirmed that this region is a primary tool for phylogenetic studies because of mutation rates of insertion–deletion. Thus, both sequences (trnL-F and IGS) have a good chance to apply in the future rice breeding program or developing new rice cultivars.
Overall, the tidal swamp rice of the South Kalimantan, Indonesia demonstrated a relatively more extraordinary genetic diversity than other local rice germplasm. For instance, in this study, our rice germplasm showed a genetic diversity by 0.61 for nuclear IGS and 0.58 for trnL-F, whereas the South Korean local rice, using SSR markers, only had 0.035 [22]. Thus, this indicates that the tidal swamp rice germplasm experienced a long-term period of domestication process [37]. In fact, this process may generate the changes of an individual genome [38]. Furthermore, the Tajima test of neutrality indicated that selection balance (there are no rare alleles) has occurred in this rice population [39]. Thus, the more accurate application of other molecular markers, such as single nucleotide polymorphism (SNP), to study the local rice population is profoundly important to do.
Furthermore, studies on phylogenetic relationships are indispensable to support rice breeding programs in the future [40], particularly in choosing the parents for developing new rice cultivars. These studies are extensively conducted by many researchers—such as Kim et al. [22], Choudhury et al. [41], Liu et al. [11], Zhang et al. [42], etc.—using different molecular markers, both in a local and a global scale. Nevertheless, such studies are still limited to the tidal swamp rice germplasm.
Our results revealed the phylogenetic relationships of the tidal swamp rice germplasm of the South Kalimantan, Indonesia based on the chloroplast trnL-F and nuclear IGS sequences. Both reflected the differences in clustering when analyzed separately, four groups for trnL-F (Figure 2) and two for IGS (Figure 3). While these trees were generated by two aligned sequences, both have a low resolution, indicated by the average of bootstrap values for each tree. Such a condition can be caused by the weakness of sequence variation used [14]. For this reason, Anderberg et al. [43] suggested the combined sequence data use in increasing the phylogenetic tree resolution.
In this study, the combined sequences of both regions provided a better resolution of a phylogenetic tree than sequence data used separately (Figure 4) but generated a similar pattern of a tree by IGS. This tree, however, could not show the germplasm groupings based on the geographical origin as well. Using the combined sequences data of chloroplast trnL-F, nuclear ITS, and mitochondrial nad1 of the Oryza genus, Duan et al. [14] reported a different result of this study, where the phylogenetic tree has a high-resolution with a geographical origin associated.
Regardless of there being no association of geographical origin, the phylogenetic trees provided a unique illustration, where the tidal swamp rice has a monophyletic divergence (derived from a common ancestor). Using the chloroplast trnL-F sequences, Hall et al. [44] succeeded to show and reconstruct the genetic relationships of Capparaceae and Brassicaceae and concluded that their genetic divergences are monophyletic. Such cases are also reported on Diseae (Orchidoideae: Orchidaceae) [45]. However, in Annonaceae, Pirie et al. [24] revealed a different phylogenetic signal to this marker. According to Pirie et al. [24] this was probably due to the gene paralogy, caused by duplicating events, in the past, where the second copy of trnL-F diverged from the first trnL-F directly from Annonaceae ancestor.
Using the chloroplasts and nuclear genome on clustering, Kim et al. [22] succeeded in classifying and reconstructing the phylogenetic relationships of 9 species of Oryza into 3 different groups. The same grouping is also shown by Duan et al. [14] when analyzing the Oryza genus using a combined sequence data of chloroplast trnL-F and nuclear ITS, as well as mitochondrial nad1. Thus, the markers can be used in other phylogenetic studies [46]. Whereas, this result may provide a particular opportunity to explore phylogenetic relationships of the tidal swamp rice from different angles in the future.
5. Conclusions
The tidal swamp rice cultivars of South Kalimantan, Indonesia demonstrated a relatively more extraordinary genetic diversity than other local rice cultivars. In this study, these germplasms showed a genetic polymorphism of 0.61 for nuclear IGS and 0.58 for trnL-F. The phylogenetic reconstruction also exhibited that the tidal swamp rice has the unique phylogenetic trees, particularly for the combined sequence datasets. Thus, this information would be useful for the rice conservation and breeding programs in the future.
Acknowledgments
We thank the Director of Indonesian Swampland Agriculture Research Institute (South Kalimantan, Indonesia) for providing Sardani (an outgroup cultivar) for this study. This study was funded by The Ministry of Research, Technology and Higher Education of the Republic of Indonesia (grant no. 061/SP2H/LT/DRPM/2018).
Author Contributions
D.H.M., P.P., I.S., and B.S.D. conceived and designed the experiments; D.H.M. and B.S.D. did the fieldwork; D.H.M. performed the experiments; D.H.M. and B.S.D. analyzed the data; I.S., P.P., and B.S.D. contributed materials; and D.H.M. and B.S.D. wrote the paper.
Conflicts of Interest
The authors declare no conflicts of interest. The funding sponsor had no role in the design of the experiments, in the collection, analysis, or interpretation of data; in the writing of the manuscript; nor in the decision to publish the results.
References
- Sinaga, P.H.; Trikoesoemaningtyas; Sopandie, D.; Aswidinnoor, H. Screening of rice genotypes and evaluation of their ratooning ability in tidal swamp area. Asian J. Agric. Res. 2014, 8, 218–233. [Google Scholar] [CrossRef]
- Ray, A.; Deb, D.; Ray, R.; Chattopadhayay, B. Phenotypic characters of rice landraces reveal independent lineages of short-grain aromatic indica rice. AOB Plants 2013, 5, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Chakravorty, A.; Ghosh, P.D.; Sahu, P.K. Multivariate analysis of phenotypic diversity of landraces of rice of West Bengal. Am. J. Exp. Agric. 2013, 3, 110–123. [Google Scholar] [CrossRef]
- Divya, K. Study of genetic diversity in Karnataka rice (Oryza Sativa) landraces using trait specific simple sequence repeat (SSR) markers. Int. J. Thesis Proj. Diss. 2013, 1, 45–70. [Google Scholar]
- Das, B.; Sengupta, S.; Parida, S.K.; Roy, B.; Ghosh, M.; Prasad, M.; Ghose, T.K. Genetic diversity and population structure of rice landraces from Eastern and North Eastern States of India. BMC Genet. 2013, 14, 71. [Google Scholar] [CrossRef] [PubMed]
- Mursyidin, D.H.; Nazari, Y.A.; Daryono, B.S. Tidal swamp rice cultivars of South Kalimantan Province, Indonesia: A case study of diversity and local culture. Biodivers. J. Biol. Divers. 2017, 18, 427–432. [Google Scholar] [CrossRef]
- Thomson, M.J.; Septiningsih, E.M.; Suwardjo, F.; Santoso, T.J.; Silitonga, T.S.; McCouch, S.R. Genetic diversity analysis of traditional and improved Indonesian rice (Oryza sativa L.) germplasm using microsatellite markers. Theor. Appl. Genet. 2007, 114, 559–568. [Google Scholar] [CrossRef] [PubMed]
- Parikh, M.; Sharma, B.; Chaudhari, P.R.; Bhandarkar, S.; Sharma, D. Collection and conservation of extinct land races of rice from Raigarh district of Chhattisgarh. Recent Res. Sci. Technol. 2014, 6, 111–112. [Google Scholar]
- Semwal, D.P.; Pandey, A.; Bhandari, D.C.; Dhariwal, O.P.; Sharma, S.K. Variability study in seed morphology and uses of indigenous rice landraces (Oryza sativa L.) collected from West Bengal, India. Aust. J. Crop Sci. 2014, 8, 460–467. [Google Scholar]
- Anumalla, M.; Roychowdhury, R.; Geda, C.K.; Mazid, M.; Rathoure, A.K. Utilization of plant genetic resources and diversity analysis tools for sustainable crop improvement with special emphasis on rice. Int. J. Adv. Res. 2015, 3, 1155–1175. [Google Scholar]
- Liu, P.; Cai, X.X.; Lu, B.R. Single-seeded InDel fingerprints in rice: An effective tool for indica-japonica rice classification and evolutionary studies. J. Syst. Evol. 2012, 50, 1–11. [Google Scholar] [CrossRef]
- Pervaiz, Z.H.; Ashiq Rabbani, M.; Khaliq, I.; Pearce, S.R.; Malik, S.A. Genetic diversity associated with agronomic traits using microsatellite markers in Pakistani rice landraces. Electron. J. Biotechnol. 2010, 13, 4–5. [Google Scholar] [CrossRef]
- Soltis, E.D.; Soltis, P.S. Contributions of plant molecular systematics to studies of molecular evolution. Plant Mol. Biol. 2000, 42, 45–75. [Google Scholar] [CrossRef] [PubMed]
- Duan, S.; Lu, B.; Li, Z.; Tong, J.; Kong, J.; Yao, W.; Li, S.; Zhu, Y. Phylogenetic analysis of AA-genome Oryza species (Poaceae) based on chloroplast, mitochondrial, and nuclear DNA sequences. Biochem. Genet. 2007, 45, 113–129. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, B.G.; Sanderson, M.J.; Porter, J.M.; Wojciechowski, M.F.; Campbell, C.S.; Donoghue, M.J. The ITS region of nuclear ribosomal DNA—A valuable source of evidence on angiosperm phylogeny. Ann. Mo. Bot. Gard. 1995, 82, 247–277. [Google Scholar] [CrossRef]
- Soltis, D.E.; Soltis, P.S. Choosing an approach and an appropriate gene for phylogenetic anlysis. In Molecular Systematics of Plants II; Springer: Boston, MA, USA, 1998; pp. 1–42. ISBN 0412111314. [Google Scholar]
- Mort, M.E.; Archibald, J.K.; Randle, C.P.; Levsen, N.D.; Ryan O’Leary, T.; Topalov, K.; Wiegand, C.M.; Crawford, D.J. Inferring phylogeny at low taxonomic levels: Utility of rapidly evolving cpDNA and nuclear ITS loci. Am. J. Bot. 2007, 94, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Rogers, S.O.; Bendich, A.J. Ribosomal RNA genes in plants: Variability in copy number and in the intergenic spacer. Plant Mol. Biol. 1987, 9, 509–520. [Google Scholar] [CrossRef] [PubMed]
- Poczai, P.; Hyvönen, J. Nuclear ribosomal spacer regions in plant phylogenetics: Problems and prospects. Mol. Biol. Rep. 2010, 37, 1897–1912. [Google Scholar] [CrossRef] [PubMed]
- Taberlet, P.; Gielly, L.; Bouvet, J.; Pautou, G. Universal primer for amplification of three non-coding regions of chloroplast DNA. Plant Mol. Biol. 1991, 17, 1105–1109. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Robin, A.H.K.; Yi, G.E.; Lee, J.; Chung, M.Y.; Yang, T.J.; Nou, I.S. Diversity and inheritance of intergenic spacer sequences of 45s ribosomal DNA among accessions of Brassica oleracea L. var. capitata. Int. J. Mol. Sci. 2015, 16, 28783–28799. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Jeong, E.G.; Ahn, S.-N.; Doyle, J.; Singh, N.; Greenberg, A.J.; Won, Y.J.; McCouch, S.R. Nuclear and chloroplast diversity and phenotypic distribution of rice (Oryza sativa L.) germplasm from the democratic People’s Republic of Korea (DPRK; North Korea). Rice 2014, 7, 7. [Google Scholar] [CrossRef] [PubMed]
- Jo, S.-H.; Koo, D.-H.; Kim, J.F.; Hur, C.-G.; Lee, S.; Yang, T.; Kwon, S.-Y.; Choi, D. Evolution of ribosomal DNA-derived satellite repeat in tomato genome. BMC Plant Biol. 2009, 9, 42. [Google Scholar] [CrossRef] [PubMed]
- Pirie, M.D.; Vargas, M.P.B.; Botermans, M.; Bakker, F.T.; Chatrou, L.W. Ancient paralogy in the cpDNA trnL-F region in Annonaceae: Implications for plant molecular systematics. Am. J. Bot. 2007, 94, 1003–1016. [Google Scholar] [CrossRef] [PubMed]
- Thomson, M.J.; Polato, N.R.; Prasetiyono, J.; Trijatmiko, K.R.; Silitonga, T.S.; McCouch, S.R. Genetic diversity of isolated populations of Indonesian landraces of rice (Oryza sativa L.) collected in East Kalimantan on the Island of Borneo. Rice 2009, 2, 80–92. [Google Scholar] [CrossRef]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1989; ISBN 13 978-0879693091. [Google Scholar]
- Cordesse, F.; Cooke, R.; Tremousaygue, D.; Grellet, F.; Delseny, M. Fine structure and evolution of the rDNA intergenic spacer in rice and other cereals. J. Mol. Evol. 1993, 36, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; Mcgettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed]
- Petersen, G.; Seberg, O. Molecular evolution and phylogenetic application of DMC1. Mol. Phylogenet. Evol. 2002, 22, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide-sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. Confidence-Limits on phylogenies—An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Data sets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Frankham, R.; Ballou, J.D.; Briscoe, D.A. A Primer of Conservation Genetics; Cambridge University Press: New York, NY, USA, 2004; Volume 39, ISBN 0521538270. [Google Scholar]
- Chung, M.C.; Lee, Y.I.; Cheng, Y.Y.; Chou, Y.J.; Lu, C.F. Chromosomal polymorphism of ribosomal genes in the genus Oryza. Theor. Appl. Genet. 2008, 116, 745–753. [Google Scholar] [CrossRef] [PubMed]
- Drábková, L.; Kirschner, J.; Vlcek, C.; Paces, V. TrnL-trnF intergenic spacer and trnL intron define major clades within Luzula and Juncus (Juncaceae): Importance of structural mutations. J. Mol. Evol. 2004, 59, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Bayer, R.; Starr, J. Tribal phylogeny of the Asteraceae based on two non-coding chloroplast sequences, the trnL intron and trnL/trnF intergenic spacer. Ann. Mo. Bot. Gard. 1998, 85, 242–256. [Google Scholar] [CrossRef]
- Guo, L.; Gao, Z.; Qian, Q. Application of resequencing to rice genomics, functional genomics and evolutionary analysis. Rice 2014, 7, 4. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, D.A.; Lu, B.R.; Tomooka, N. Was Asian rice (Oryza sativa) domesticated more than once? Rice 2008, 1, 16–24. [Google Scholar] [CrossRef]
- Tajima, F. Statistical method for testing the neutral mutation hypothesis by DNA polymorphism. Genetics 1989, 123, 585–595. [Google Scholar] [PubMed]
- Chakhonkaen, S.; Pitnjam, K.; Saisuk, W.; Ukoskit, K.; Muangprom, A. Genetic structure of Thai rice and rice accessions obtained from the International Rice Research Institute. Rice 2012, 5, 19. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, D.R.; Singh, N.; Singh, A.K.; Kumar, S.; Srinivasan, K. Analysis of genetic diversity and population structure of rice germplasm from North-Eastern region of India and development of a core germplasm set. PLoS ONE 2014, 9, e113094. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Hu, S.; Zhang, G.; Pan, L.; Zhang, X.; Al-Mssallem, I.S.; Yu, J. The organelle genomes of Hassawi rice (Oryza sativa L.) and its hybrid in Saudi Arabia: Genome variation, rearrangement, and origins. PLoS ONE 2012, 7, e42041. [Google Scholar] [CrossRef] [PubMed]
- Anderberg, A.A.; Rydin, C.; Källersjö, M. Phylogenetic relationships in the order Ericales s.l.: Analyses of molecular data from five genes from the plastid and mitochondrial genomes. Am. J. Bot. 2002, 89, 677–687. [Google Scholar] [CrossRef] [PubMed]
- Hall, J.C.; Sytsma, K.J.; Iltis, H.H. Phylogeny of Capparaceae and Brassicaceae based on chloroplast sequence data. Am. J. Bot. 2002, 89, 1826–1842. [Google Scholar] [CrossRef] [PubMed]
- Bellstedt, D.U.; Linder, H.P.; Harley, E.H. Phylogenetic relationships in Disa based on non-coding Trn L-Trn F chloroplast sequences: Evidence of numerous repeat region. Am. J. Bot. 2001, 88, 2088–2100. [Google Scholar] [CrossRef] [PubMed]
- Wambugu, P.W.; Brozynska, M.; Furtado, A.; Waters, D.L.; Henry, R.J. Relationships of wild and domesticated rices (Oryza AA genome species) based upon whole chloroplast genome sequences. Sci. Rep. 2015, 5, 13957. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
A map of Indonesia showing three collecting sites of the tidal swamp rice in the South Kalimantan Province, Indonesia: Tanah Laut (A), Banjar (B), and Barito Kuala (C).
Figure 1.
A map of Indonesia showing three collecting sites of the tidal swamp rice in the South Kalimantan Province, Indonesia: Tanah Laut (A), Banjar (B), and Barito Kuala (C).

Figure 2.
Phylogenetic tree of the tidal swamp rice based on the chloroplast trnL-F sequences. The numbers above branches indicate bootstrap values inferred from 1000 replicates (a an outgroup).
Figure 2.
Phylogenetic tree of the tidal swamp rice based on the chloroplast trnL-F sequences. The numbers above branches indicate bootstrap values inferred from 1000 replicates (a an outgroup).

Figure 3.
Phylogenetic tree of the tidal swamp rice based on the nuclear IGS sequences. The numbers above branches indicate bootstrap values inferred from 1000 replicates (a an outgroup).
Figure 3.
Phylogenetic tree of the tidal swamp rice based on the nuclear IGS sequences. The numbers above branches indicate bootstrap values inferred from 1000 replicates (a an outgroup).

Figure 4.
Phylogenetic tree of the tidal swamp rice inferred from the combined of chloroplast trnL-F and nuclear IGS sequences. The numbers above branches indicate bootstrap values inferred from 1000 replicates (a an outgroup).
Figure 4.
Phylogenetic tree of the tidal swamp rice inferred from the combined of chloroplast trnL-F and nuclear IGS sequences. The numbers above branches indicate bootstrap values inferred from 1000 replicates (a an outgroup).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
List of the tidal swamp rice employed in this study.
| Name of Cultivars | Genetic Status | Grain Type | Sub-Species | Origin |
|---|---|---|---|---|
| Adil Ganal | Landrace | Non-glutinous | Javanica | Banjar, South Kalimantan |
| Banih Putih | Landrace | Non-glutinous | Javanica | Banjar, South Kalimantan |
| Bayar Papuyu | Landrace | Non-glutinous | Close to Japonica | Banjar, South Kalimantan |
| Bayar Putih | Landrace | Non-glutinous | Javanica | Banjar, South Kalimantan |
| Ciherang | Improved | Non-glutinous | Javanica | Tanah Laut, South Kalimantan |
| Ganal Perak | Landrace | Non-glutinous | Javanica | Banjar, South Kalimantan |
| Lakatan Siam | Landrace | Glutinous | Javanica | Banjar, South Kalimantan |
| Lakatan Pacar | Landrace | Glutinous | Javanica | Barito Kuala, South Kalimantan |
| Lakatan Wangi | Landrace | Glutinous | Javanica | Banjar, South Kalimantan |
| Pandak Laut | Landrace | Non-glutinous | Intermediate | Banjar, South Kalimantan |
| Pandak Putih | Landrace | Non-glutinous | Javanica | Banjar, South Kalimantan |
| Siam Babirik | Landrace | Non-glutinous | Intermediate | Tanah Laut, South Kalimantan |
| Siam Kuning | Landrace | Non-glutinous | Javanica | Tanah Laut, South Kalimantan |
| Siam Mutiara | Landrace | Non-glutinous | Indica | Barito Kuala, South Kalimantan |
| Siam Oon | Landrace | Non-glutinous | Indica | Banjar, South Kalimantan |
| Sardani1 | Landrace | Non-glutinous | Close to Indica | South Sumatera |
1 An outgroup.
Table 2.
Primers used for PCR amplification and for cycle sequencing.
| Marker | Primer | Sequences (5′ to 3′) | Expected Size (bp) | Ta/Tm (°C) | Reference |
|---|---|---|---|---|---|
| trnL-F | Forward | CGAAATCGGTAGACGCTACG | 950 | 55 | Taberlet et al. [20] |
| Reverse | ATTTGAACTGGTGACACGAG | ||||
| IGS | Forward | TAGGCCTCTCTAAGTCCGCT | 1612 | 53 | Cordesse et al. [27] |
| Reverse | GTCCAAAGCAGCTCATACGC |
Table 3.
Comparison of phylogenetic information for the tidal swamp rice genome from two datasets 1.
Table 3.
Comparison of phylogenetic information for the tidal swamp rice genome from two datasets 1.
| Parameter | Chloroplast trnL-F | Nuclear IGS | Combined |
|---|---|---|---|
| Range of sequence length (bp) | 936–953 | 849–1405 | 1796–2352 |
| Total number of sites (n) | 936 | 849 | 1780 |
| Number of polymorphic sites (S) | 929 | 845 | 266 |
| Parameters | 30 | 30 | 30 |
| Bayesian information criterion (BIC) | 20,221.72 | 29,081.93 | 60,694.01 |
| Akaike information criterion (AICc) | 19,993.42 | 28,856.57 | 60,446.29 |
| Maximum likelihood value (lnL) | −9966.65 | −14,398.22 | −30,193.11 |
| Transition/transversion bias value (R) | 0.58 | 0.56 | 0.68 |
| Ti/Tv ratio | 1.13 | 1.13 | 0.89 |
| GC content (%) | 35.71 | 69.10 | 51.60 |
| Nucleotide diversity (π) | 0.58 | 0.61 | 0.04 |
| Tajima’s test of neutrality (D) | 4.12 | 4.57 | −0.54 |
1 Based on Kimura two-parameter model.
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Mursyidin, D.H.; Purnomo, P.; Sumardi, I.; Daryono, B.S. Molecular Diversity of Tidal Swamp Rice (Oryza sativa L.) in South Kalimantan, Indonesia. Diversity 2018, 10, 22. https://doi.org/10.3390/d10020022
AMA Style
Mursyidin DH, Purnomo P, Sumardi I, Daryono BS. Molecular Diversity of Tidal Swamp Rice (Oryza sativa L.) in South Kalimantan, Indonesia. Diversity. 2018; 10(2):22. https://doi.org/10.3390/d10020022
Chicago/Turabian StyleMursyidin, Dindin H., Purnomo Purnomo, Issirep Sumardi, and Budi S. Daryono. 2018. "Molecular Diversity of Tidal Swamp Rice (Oryza sativa L.) in South Kalimantan, Indonesia" Diversity 10, no. 2: 22. https://doi.org/10.3390/d10020022
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.