

Testosterone Inhibits Secretion of the Pro-Inflammatory Chemokine CXCL1 from Astrocytes
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Viability Assay
2.3. Gene Expression Assay
2.4. Quantification of CXCL1 Secreted to the Medium by Astrocytes
2.5. Statistical Analysis
3. Results
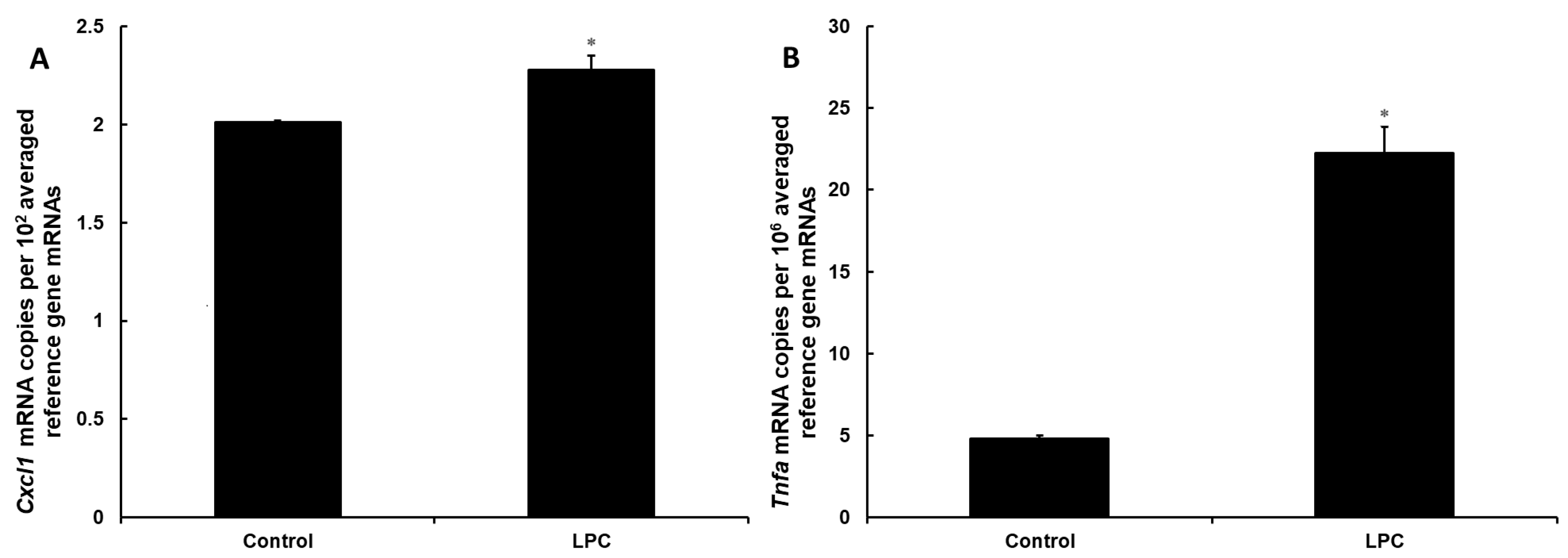
3.1. Lysophosphatidylcholine Is Not Toxic to DI TNC1 Astrocytes but Exerts a Pro-Inflammatory Effect at the Signaling Level
3.2. DI TNC1 Astrocytes Produce an Increased Amount of CXCL1 When Treated with Lysophosphatidylcholine
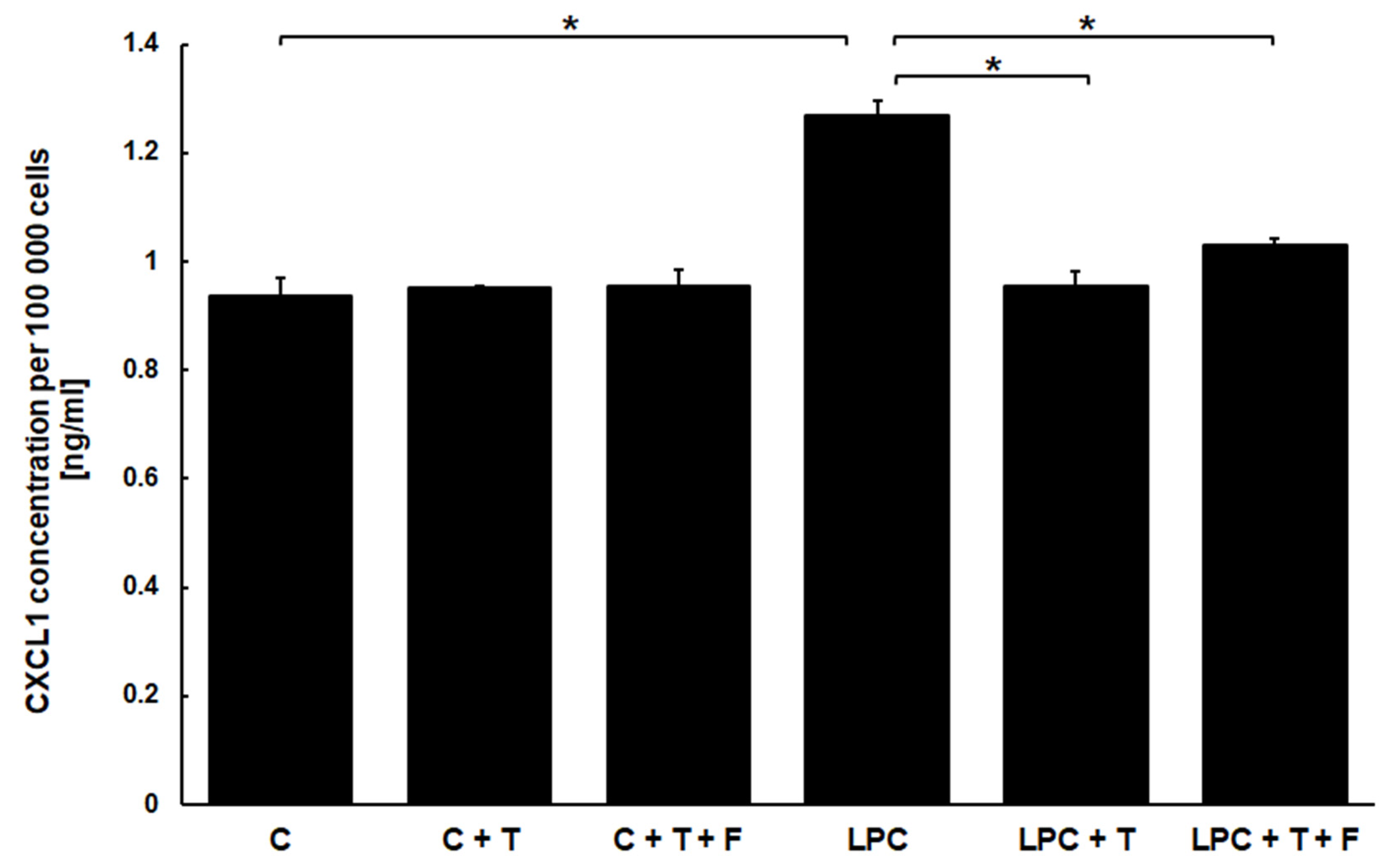
3.3. Testosterone Affects the Production of CXCL1 but Only in Cells Stimulated with Lysophosphatidylcholine
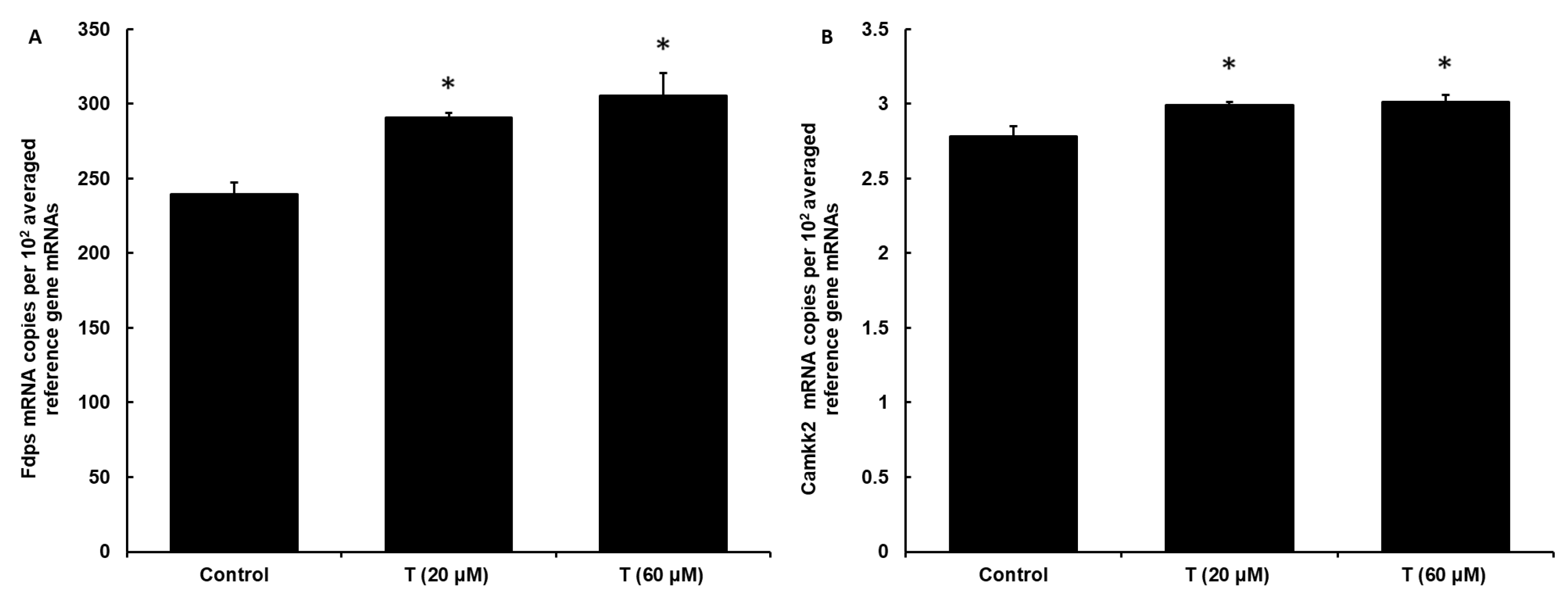
3.4. DI TNC1 Cells Express Functional Androgen Receptor
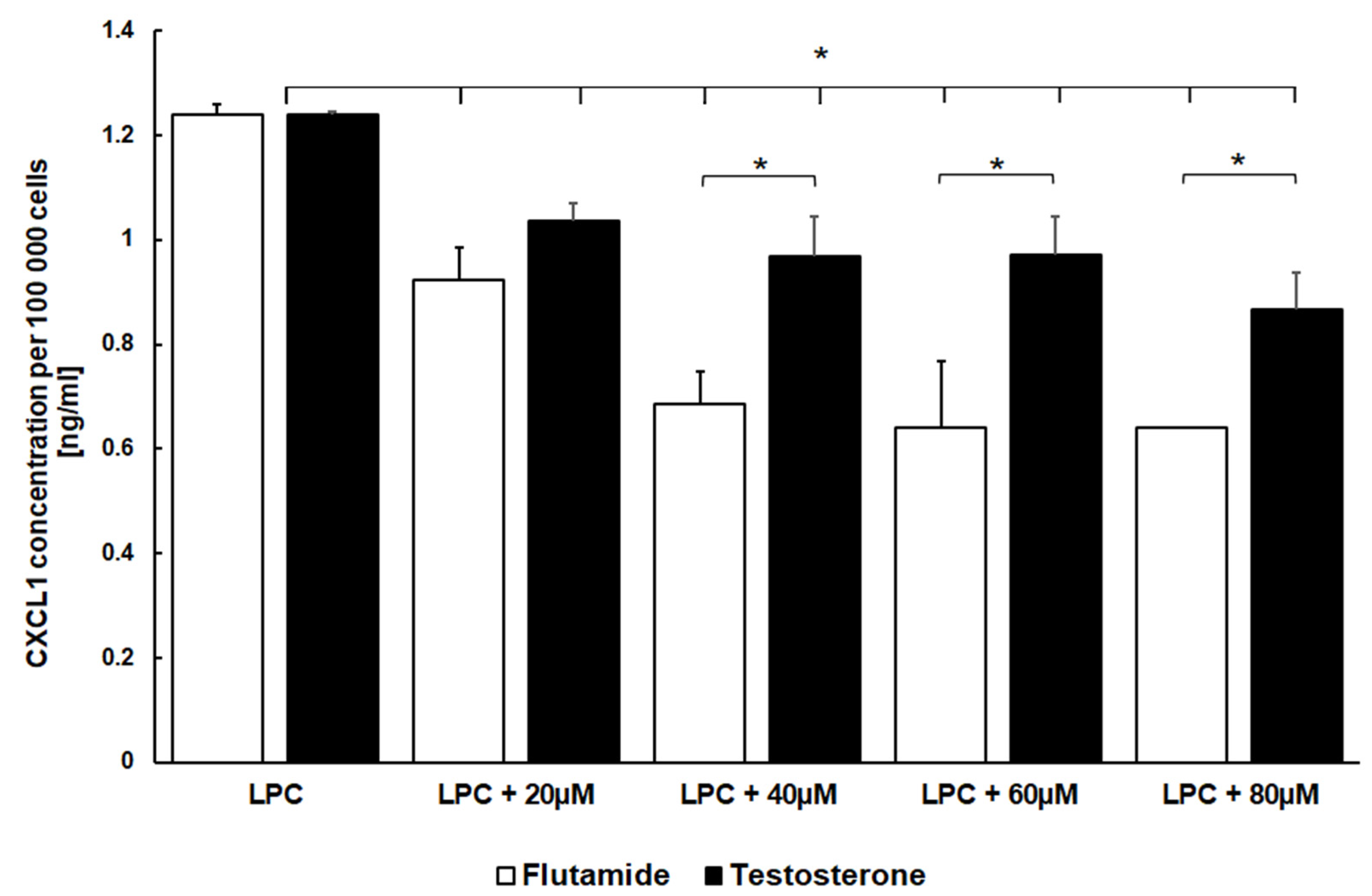
3.5. Flutamide Similarly to Testosterone Reduces the Pro-Inflammatory Stimulus of LPC on CXCL1 Secretion
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Choi, S.S.; Lee, H.J.; Lim, I.; Satoch, J.; Kim, S.U. Human astrocytes: Secretome profiles of cytokines and chemokines. PLoS ONE 2014, 9, e92325. [Google Scholar] [CrossRef]
- Stalder, A.K.; Pagenstecher, A.; Yu, N.C.; Kincaid, C.; Chiang, C.S.; Hobbs, M.V.; Bloom, F.E.; Campbell, I.L. Lipopolysaccharide-induced IL-12 expression in the central nervous system and cultured astrocytes and microglia. J. Immunol. 1997, 159, 1344–1351. [Google Scholar] [CrossRef]
- Van Wagoner, N.J.; Oh, J.W.; Repovic, P.; Benveniste, E.N. Interleukin-6 (IL-6) production by astrocytes: Autocrine regulation by IL-6 and the soluble IL-6 receptor. J. Neurosci. 1999, 19, 5236–5244. [Google Scholar] [CrossRef]
- Sénécal, V.; Deblois, G.; Beauseigle, D.; Schneider, R.; Brandenburg, J.; Newcombe, J.; Moore, C.S.; Prat, A.; Mantel, J.; Arbour, N. Production of IL-27 in multiple sclerosis lesions by astrocytes and myeloid cells: Modulation of local immune responses. Glia 2016, 64, 553–569. [Google Scholar] [CrossRef]
- Wu, Z.; Zhang, J.; Nakanishi, H. Leptomeningeal cells activate microglia and astrocytes to induce IL-10 production by releasing pro-inflammatory cytokines during systemic inflammation. J. Neuroimmunol. 2005, 167, 90–98. [Google Scholar] [CrossRef]
- Cooley, I.D.; Chauhan, V.S.; Donneyz, M.A.; Marriott, I. Astrocytes produce IL-19 in response to bacterial challenge and are sensitive to the immunosuppressive effects of this IL-10 family member. Glia 2014, 62, 818–828. [Google Scholar] [CrossRef]
- Rubio, N.; Sanz-Rodriguez, F.; Lipton, H.L. Theiler’s virus induces the MIP-2 chemokine (CXCL2) in astrocytes from genetically susceptible but not from resistant mouse strains. Cell Immunol. 2006, 239, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Glabinski, A.R.; Tuhoy, V.K.; Ransohoff, R.M. Expression of chemokines RANTES, MIP-1alpha and GRO-alpha correlates with inflammation in acute experimental autoimmune encephalomyelitis. Neuroimmunomodulation 1998, 5, 166–171. [Google Scholar] [CrossRef] [PubMed]
- Blaževski, J.; Petković, F.; Momcilovic, M.; Jevtic, B.; Mostarica-Stojković, M.; Miljkovic, D. Tumor necrosis factor stimulates expression of CXCL12 in astrocytes. Immunobiology 2015, 220, 845–850. [Google Scholar] [CrossRef] [PubMed]
- Sanmarco, L.M.; Polonio, C.M.; Wheeler, M.A.; Quintana, F.J. Functional immune cell-astrocyte interactions. J. Exp. Med. 2021, 218, e20202715. [Google Scholar] [CrossRef] [PubMed]
- Eddleston, M.; Mucke, L. Molecular profile of reactive astrocytes–implications for their role in neurologic disease. Neuroscience 1993, 54, 15–36. [Google Scholar] [CrossRef]
- Verkhratsky, A.; Parpura, V. Recent advances in (patho)physiology of astroglia. Acta Pharmacol. Sin. 2010, 31, 1044–1054. [Google Scholar] [CrossRef]
- Haskill, S.; Peace, A.; Sporn, S.A.; Anisowicz, A.; Lee, S.W.; Smith, T.; Martin, G.; Ralph, P.; Cases, R. Identification of three related human GRO genes encoding cytokine functions. Proc. Natl. Acad. Sci. USA 1990, 87, 7732–7736. [Google Scholar] [CrossRef]
- Iida, N.; Grotendorst, G.R. Cloning and sequencing of a new gro transcript from activated human monocytes: Expression in leukocytes and wound tissue. Mol. Cell. Biol. 1990, 10, 5596–5599. [Google Scholar]
- Becker, S.; Quay, J.; Koren, H.S.; Haskill, J.S. Constitutive and stimulated MCP-1, GRO alpha, beta, and gamma expression in human airway epithelium and bronchoalveolar macrophages. Am. J. Physiol. 1994, 266 Pt 1, L278–L286. [Google Scholar] [CrossRef]
- Omari, K.M.; John, G.; Lango, R.; Raine, C.S. Role for CXCR2 and CXCL1 on glia in multiple sclerosis. Glia 2006, 53, 24–31. [Google Scholar] [CrossRef]
- Katancik, J.A.; Sharma, A.; de Nardin, E. Interleukin 8, neutrophil-activating peptide-2 and GRO-alpha bind to and elicit cell activation via specific and different amino acid residues of CXCR2. Cytokine 2000, 12, 1480–1488. [Google Scholar] [CrossRef]
- Sawant, K.V.; Poluri, K.M.; Dutta, A.K.; Sepuru, K.M.; Troshkina, A.; Garofalo, R.P.; Rajarathnam, K. Chemokine CXCL1 mediated neutrophil recruitment: Role of glycosaminoglycan interactions. Sci. Rep. 2016, 6, 33123. [Google Scholar] [CrossRef]
- Schumacher, C.; Clark-Lewis, I.; Baggiolini, M.; Moser, B. High- and low-affinity binding of GRO alpha and neutrophil-activating peptide 2 to interleukin 8 receptors on human neutrophils. Proc. Natl. Acad. Sci. USA 1992, 89, 10542–10546. [Google Scholar] [CrossRef] [PubMed]
- Serdar, M.; Kempe, K.; Herrmann, R.; Picard, D.; Remke, M.; Herz, J.; Bendix, I.; Felderhoff-Müser, U.; Sabir, H. Involvement of CXCL1/CXCR2 During Microglia Activation Following Inflammation-Sensitized Hypoxic-Ischemic Brain Injury in Neonatal Rats. Front. Neurol. 2020, 11, 540878. [Google Scholar] [CrossRef] [PubMed]
- Robinson, S.; Tani, M.; Strieter, R.M.; Ransohoff, R.M.; Miller, R.H. The chemokine growth-regulated oncogene-alpha promotes spinal cord oligodendrocyte precursor proliferation. J. Neurosci. 1998, 18, 10457–10463. [Google Scholar] [CrossRef]
- Kadi, L.; Selvaraju, R.; de Lys, P.; Proudfoot, A.E.I.; Wells, T.N.C.; Boschert, U. Differential effects of chemokines on oligodendrocyte precursor proliferation and myelin formation in vitro. J. Neuroimmunol. 2006, 174, 133–146. [Google Scholar] [CrossRef]
- Omari, K.M.; John, G.R.; Sealfon, S.C.; Raine, C.S. CXC chemokine receptors on human oligodendrocytes: Implications for multiple sclerosis. Brain 2005, 128 Pt 5, 1003–1015. [Google Scholar] [CrossRef]
- Filipovic, R.; Zecevic, N. The effect of CXCL1 on human fetal oligodendrocyte progenitor cells. Glia 2008, 56, 1–15. [Google Scholar] [CrossRef]
- Wang, L.; Yang, H.; Zang, C.; Dong, Y.; Shang, J.; Chen, J.; Wang, Y.; Liu, H.; Zhang, Z.; Xu, H.; et al. CXCR2 antagonism promotes oligodendrocyte precursor cell differentiation and enhances remyelination in a mouse model of multiple sclerosis. Neurobiol. Dis. 2020, 134, 104630. [Google Scholar] [CrossRef]
- Müller-Ladner, U.; Jones, J.L.; Wetsel, R.A.; Gay, S.; Raine, C.S.; Barnum, S.R. Enhanced expression of chemotactic receptors in multiple sclerosis lesions. J. Neurol. Sci. 1996, 144, 135–141. [Google Scholar] [CrossRef]
- Lustig, R.H. Sex hormone modulation of neural development in vitro. Horm. Behav. 1994, 28, 383–395. [Google Scholar] [CrossRef]
- Zung, A.; Phillip, M.; Chalew, S.A.; Palese, T.; Kowarski, A.A.; Zadik, Z. Testosterone effect on growth and growth mediators of the GH-IGF-I axis in the liver and epiphyseal growth plate of juvenile rats. J. Mol. Endocrinol. 1999, 23, 209–221. [Google Scholar] [CrossRef]
- Chin, K.Y.; Ima-Nirwana, S. The effects of orchidectomy and supraphysiological testosterone administration on trabecular bone structure and gene expression in rats. Aging Male 2015, 18, 60–66. [Google Scholar] [CrossRef]
- Zhang, Y.; Champagne, N.; Beitel, L.K.; Goodyer, C.G.; Trifiro, M.; LeBlanc, A. Estrogen and androgen protection of human neurons against intracellular amyloid beta1-42 toxicity through heat shock protein 70. J. Neurosci. 2004, 24, 5315–5321. [Google Scholar] [CrossRef]
- Białek, M.; Zaremba, P.; Borowicz, K.K.; Czuczwar, S.J. Neuroprotective role of testosterone in the nervous system. Pol. J. Pharmacol. 2004, 56, 509–518. [Google Scholar]
- Ogata, T.; Nakamura, Y.; Tsuji, K.; Shibata, T.; Kataoka, K. Steroid hormones protect spinal cord neurons from glutamate toxicity. Neuroscience 1993, 55, 445–449. [Google Scholar] [CrossRef]
- Asgari, N. Epidemiological, clinical and immunological aspects of neuromyelitis optica (NMO). Dan. Med. J. 2013, 60, B4730. [Google Scholar]
- Wingerchuk, D.M. Neuromyelitis optica: Effect of gender. J. Neurol. Sci. 2009, 286, 18–23. [Google Scholar] [CrossRef]
- Tintoré, M.; Arrambide, G. Early onset multiple sclerosis: The role of gender. J. Neurol. Sci. 2009, 286, 31–34. [Google Scholar] [CrossRef]
- Voumvourakis, K.I.; Tsiodras, S.; Kitsos, D.K.; Stamboulis, E. Gender hormones: Role in the pathogenesis of central nervous system disease and demyelination. Curr. Neurovasc. Res. 2008, 5, 224–235. [Google Scholar] [CrossRef]
- Pugliatti, M.; Rosati, G.; Carton, H.; Riise, T.; Drulovic, J.; Vécsei, L.; Milanov, I. The epidemiology of multiple sclerosis in Europe. Eur. J. Neurol. 2006, 13, 700–722. [Google Scholar] [CrossRef]
- Harbo, H.F.; Gold, R.; Tintoré, M. Sex and gender issues in multiple sclerosis. Ther. Adv. Neurol. Disord. 2013, 6, 237–248. [Google Scholar] [CrossRef]
- Disanto, G.; Ramagopalan, S.V. On the sex ratio of multiple sclerosis. Mult. Scler. J. 2013, 19, 3–4. [Google Scholar] [CrossRef]
- Ribbons, K.A.; McElduff, P.; Boz, C.; Trojano, M.; Izquierdo, G.; Duquette, P.; Girard, M.; Grand’Maison, F.; Hupperts, R.; Grammond, P.; et al. Male Sex Is Independently Associated with Faster Disability Accumulation in Relapse-Onset MS but Not in Primary Progressive MS. PLoS ONE 2015, 10, e0122686. [Google Scholar] [CrossRef]
- Pakpoor, J.; Goldacre, R.; Schmierer, K.; Giovannoni, G.; Goldacre, M.J. Testicular hypofunction and multiple sclerosis risk: A record-linkage study. Ann. Neurol. 2014, 76, 625–628. [Google Scholar] [CrossRef]
- Gold, S.M.; Chalifoux, S.; Giesser, B.S.; Voskuhl, R.R. Immune modulation and increased neurotrophic factor production in multiple sclerosis patients treated with testosterone. J. Neuroinflamm. 2008, 5, 32. [Google Scholar] [CrossRef]
- Bebo, B.F.; Schuster, J.C.; Vandenbark, A.A.; Offner, H. Androgens alter the cytokine profile and reduce encephalitogenicity of myelin-reactive T cells. J. Immunol. 1999, 162, 35–40. [Google Scholar] [CrossRef]
- Liva, S.M.; Voskuhl, R.R. Testosterone acts directly on CD4+ T lymphocytes to increase IL-10 production. J. Immunol. 2001, 167, 2060–2067. [Google Scholar] [CrossRef]
- Johnson, R.T.; Schneider, A.; DonCarlos, L.L.; Breedlove, S.M.; Jordan, C.L. Astrocytes in the rat medial amygdala are responsive to adult androgens. J. Comp. Neurol. 2012, 520, 2531–2544. [Google Scholar] [CrossRef]
- Satriotomo, I.; Miki, T.; Gonzalez, D.; Matsumoto, Y.; Li, H.P.; Gu, H.; Takeuchi, Y. Excessive testosterone treatment and castration induce reactive astrocytes and fos immunoreactivity in suprachiasmatic nucleus of mice. Brain Res. 2004, 1020, 130–139. [Google Scholar] [CrossRef]
- Gu, F.; Hata, R.; Toku, K.; Yang, L.; Ma, Y.J.; Maeda, N.; Sakanaka, M.; Tanaka, J. Testosterone up-regulates aquaporin-4 expression in cultured astrocytes. J. Neurosci. Res. 2003, 72, 709–715. [Google Scholar] [CrossRef]
- Lorenz, B.; Garcia-Segura, L.M.; DonCarlos, L.L. Cellular phenotype of androgen receptor-immunoreactive nuclei in the developing and adult rat brain. J. Comp. Neurol. 2005, 492, 456–468. [Google Scholar] [CrossRef]
- Mhaouty-Kodja, S. Role of the androgen receptor in the central nervous system. Mol. Cell. Endocrinol. 2018, 465, 103–112. [Google Scholar] [CrossRef]
- Sygnecka, K.; Hein, C.; Scherf, N.; Fasold, M.; Binder, H.; Scheller, C.; Franke, H. Nimodipine enhances neurite outgrowth in dopaminergic brain slice co-cultures. Int. J. Dev. Neurosci. 2015, 40, 1–11. [Google Scholar] [CrossRef]
- Klimas, J.; Olvedy, M.; Ochodnicka-Mackovicova, K.; Kruzliak, P.; Cacanyiova, S.; Kristek, F.; Krenek, P.; Ochodnicky, P. Perinatally administered losartan augments renal ACE2 expression but not cardiac or renal Mas receptor in spontaneously hypertensive rats. J. Cell. Mol. Med. 2015, 19, 1965–1974. [Google Scholar] [CrossRef]
- Chen, Z.; Chintagari, N.R.; Guo, Y.; Bhaskaran, M.; Chen, J.; Gao, L.; Jin, N.; Weng, T.; Liu, L. Gene expression of rat alveolar type II cells during hyperoxia exposure and early recovery. Free Radic. Biol. Med. 2007, 43, 628–642. [Google Scholar] [CrossRef]
- Yu, S.; Li, Y.; Zhao, H.; Wang, Q.; Chen, P. The Histone Demethylase JMJD1C Regulates CAMKK2-AMPK Signaling to Participate in Cardiac Hypertrophy. Front. Physiol. 2020, 11, 539. [Google Scholar] [CrossRef]
- Schludi, B.; Moin, A.S.M.; Montemurro, C.; Gurlo, T.; Matveyenko, A.V.; Kirakossian, D.; Dawson, D.W.; Dry, S.M.; Butler, P.C.; Butler, A.E. Islet inflammation and ductal proliferation may be linked to increased pancreatitis risk in type 2 diabetes. JCI Insight 2017, 2, 5499368. [Google Scholar] [CrossRef]
- Gujral, J.S.; Liu, J.; Farhood, A.; Jaeschke, H. Reduced oncotic necrosis in Fas receptor-deficient C57BL/6J-lpr mice after bile duct ligation. Hepatology 2004, 40, 998–1007. [Google Scholar] [CrossRef]
- Omari, K.M.; Lutz, S.E.; Santambrogio, L.; Lira, S.A.; Raine, C.S. Neuroprotection and remyelination after autoimmune demyelination in mice that inducibly overexpress CXCL1. Am. J. Pathol. 2009, 174, 164–176. [Google Scholar] [CrossRef]
- Hosking, M.P.; Tirotta, E.; Ransohoff, R.M.; Lane, T.E. CXCR2 signaling protects oligodendrocytes and restricts demyelination in a mouse model of viral-induced demyelination. PLoS ONE 2010, 5, e11340. [Google Scholar] [CrossRef]
- Grist, J.J.; Marro, B.S.; Skinner, D.D.; Syage, A.R.; Worne, C.; Doty, D.J.; Fujinami, R.S.; Lane, T.E. Induced CNS expression of CXCL1 augments neurologic disease in a murine model of multiple sclerosis via enhanced neutrophil recruitment. Eur. J. Immunol. 2018, 48, 1199–1210. [Google Scholar] [CrossRef]
- Zhang, J.; Harada, Y.; Hayashi, Y. A TLR-CXCL1 pathway in DRG neurons induces neutrophil accumulation in the DRG and mechanical allodynia in EAE mice. Sci. Rep. 2019, 9, 12003. [Google Scholar] [CrossRef]
- Kerstetter, A.E.; Padovani-Claudio, D.A.; Bai, L.; Miller, R.H. Inhibition of CXCR2 signaling promotes recovery in models of multiple sclerosis. Exp. Neurol. 2009, 220, 44–56. [Google Scholar] [CrossRef]
- Kalincik, T.; Vivek, V.; Jokubaitis, V.; Lechner-Scott, J.; Trojano, M.; Izquierdo, G.; Lugaresi, A.; Grand’Maison, F.; Hupperts, R.; Oreja-Guevara, C.; et al. Sex as a determinant of relapse incidence and progressive course of multiple sclerosis. Brain 2013, 136 Pt 12, 3609–3617. [Google Scholar] [CrossRef]
- Pozzilli, C.; Tomassini, V.; Marinelli, F.; Paolillo, A.; Gasperini, C.; Bastianello, S. ‘Gender gap’ in multiple sclerosis: Magnetic resonance imaging evidence. Eur. J. Neurol. 2003, 10, 95–97. [Google Scholar] [CrossRef]
- Khalid, R. Contributing factors in multiple sclerosis and the female sex bias. Immunol. Lett. 2014, 162 Pt A, 223–232. [Google Scholar] [CrossRef]
- Fazekas, F.; Enzinger, C.; Wallner-Blazek, M.; Ropele, S.; Pluta-Fuerst, A.; Fox, S. Gender differences in MRI studies on multiple sclerosis. J. Neurol. Sci. 2009, 286, 28–30. [Google Scholar] [CrossRef]
- Bove, R.M.; Healy, B.; Augustine, A.; Musallam, A.; Gholipour, T.; Chitnis, T. Effect of gender on late-onset multiple sclerosis. Mult. Scler. J. 2012, 18, 1472–1479. [Google Scholar] [CrossRef]
- Bielecki, B.; Matters, C.; Ghoumari, A.M.; Javaid, S.; Smietanka, K.; Abi Ghanem, C.; Mhaouty-Kodja, S.; Ghandour, M.S.; Baulieu, E.E.; Franklin, R.J.M.; et al. Unexpected central role of the androgen receptor in the spontaneous regeneration of myelin. Proc. Natl. Acad. Sci. USA 2016, 113, 14829–14834. [Google Scholar] [CrossRef]
- Ghasemlou, N.; Jeong, S.Y.; Lacroix, S.; David, S. T cells contribute to lysophosphatidylcholine-induced macrophage activation and demyelination in the CNS. Glia 2007, 55, 294–302. [Google Scholar] [CrossRef]
- Plemel, J.R.; Michaels, N.J.; Weishaupt, N.; Caprariello, A.V.; Keough, M.B.; Rogers, J.A.; Yukseloglu, A.; Lim, J.; Patel, V.V.; Rawji, K.S.; et al. Mechanisms of lysophosphatidylcholine-induced demyelination: A primary lipid disrupting myelinopathy. Glia 2018, 66, 327–347. [Google Scholar] [CrossRef]
- Hung, N.D.; Sok, D.E.; Kim, M.R. Prevention of 1-palmitoyl lysophosphatidylcholine-induced inflammation by polyunsaturated acyl lysophosphatidylcholine. Inflamm. Res. 2012, 61, 473–483. [Google Scholar] [CrossRef]
- Bach, G.; Perrin-Cocon, L.; Gerossier, E.; Guironnet-Paquet, A.; Lotteau, V.; Inchauspé, G.; Fournillier, A. Singlelysophosphatidylcholine components exhibit adjuvant activities in vitro and in vivo. Clin. Vaccine Immunol. 2010, 17, 429–438. [Google Scholar] [CrossRef]
- Sheikh, A.M.; Nagai, A.; Ryu, J.K.; McLarnon, J.G.; Kim, S.U.; Masuda, J. Lysophosphatidylcholine induces glial cell activation: Role of rho kinase. Glia 2009, 57, 898–907. [Google Scholar] [CrossRef]
- Inose, Y.; Kato, Y.; Kitagawa, K.; Uchiyama, S.; Shibata, N. Activated microglia in ischemic stroke penumbra upregulate MCP-1 and CCR2 expression in response to lysophosphatidylcholine derived from adjacent neurons and astrocytes. Neuropathology 2015, 35, 209–223. [Google Scholar] [CrossRef]
- Hussain, R.; Ghoumari, A.M.; Bielecki, B.; Steibel, J.; Bohem, N.; Liere, P.; Macklin, W.B.; Kumar, N.; Habert, R.; Mhaouty-Kodja, S.; et al. The neural androgen receptor: A therapeutic target for myelin repair in chronic demyelination. Brain 2013, 136 Pt 1, 132–146. [Google Scholar] [CrossRef]
- Sellau, J.; Groneberg, M.; Fehling, H.; Thye, T.; Hoenow, S.; Marggraff, C.; Weskamm, M.; Hansen, C.; Stanelle-Bertram, S.; Kuhl, S.; et al. Androgens predispose males to monocyte-mediated immunopathology by inducing the expression of leukocyte recruitment factor CXCL1. Nat. Commun. 2020, 11, 3459. [Google Scholar] [CrossRef]
- Itoh, N.; Itoh, Y.; Tassoni, A.; Ren, E.; Kaito, M.; Ohno, A.; Ao, Y.; Farkhondeh, V.; Johnsonbaugh, H.; Burda, J.; et al. Cell-specific and region-specific transcriptomics in the multiple sclerosis model: Focus on astrocytes. Proc. Natl. Acad. Sci. USA 2018, 115, E302–E309. [Google Scholar] [CrossRef]
- Masiello, D.; Cheng, S.; Bubley, G.J.; Lu, M.L.; Balk, S.P. Bicalutamide functions as an androgen receptor antagonist by assembly of a transcriptionally inactive receptor. J. Biol. Chem. 2002, 277, 26321–26326. [Google Scholar] [CrossRef]
- Nguyen, T.V.; Yao, M.; Pike, C.J. Flutamide and cyproterone acetate exert agonist effects: Induction of androgen receptor-dependent neuroprotection. Endocrinology 2007, 148, 2936–2943. [Google Scholar] [CrossRef]
- Prekovic, S.; van Royen, M.E.; Voet, A.R.D.; Geverts, B.; Houtman, R.; Melchers, D.; Zhang, K.Y.J.; Van den Broeck, T.; Smeets, E.; Spans, L.; et al. The Effect of F877L and T878A Mutations on Androgen Receptor Response to Enzalutamide. Mol. Cancer Ther. 2016, 15, 1702–1712. [Google Scholar] [CrossRef]
- Kata, D.; Gróf, I.; Hoyk, Z.; Ducza, E.; Deli, M.A.; Zupkó, I.; Földesi, I. Immunofluorescent Evidence for Nuclear Localization of Aromatase in Astrocytes in the Rat Central Nervous System. Int. J. Mol. Sci. 2022, 23, 8946. [Google Scholar] [CrossRef]
- Thiel, G.; Welck, J.; Wissenbach, U.; Rössler, O.G. Dihydrotestosterone activates AP-1 in LNCaP prostate cancer cells. Int. J. Biochem. 2019, 110, 9–20. [Google Scholar] [CrossRef]
- Shieh, J.-M.; Tsai, Y.-J.; Tsou, C.-J.; Wu, W.-B. CXCL1 Regulation in Human Pulmonary Epithelial Cells by Tumor Necrosis Factor. Cell. Physiol. Biochem. 2014, 34, 1373–1384. [Google Scholar] [CrossRef]
- Karim, H.; Kim, S.H.; Lapato, A.S.; Yasui, N.; Katzenellenbogen, J.A.; Tiwari-Woodruff, S.K. Increase in chemokine CXCL1 by ERβ ligand treatment is a key mediator in promoting axon myelination. Proc. Natl. Acad. Sci. USA 2018, 115, 6291–6296. [Google Scholar] [CrossRef] [PubMed]
- Salinas-Muñoz, L.; Campos-Fernández, R.; Olivera-Valle, I.; Mercader, E.; Fernandez-Pacheco, C.; Lasarte, S.; Pérez-Martín, L.; Navarro-González, M.T.; Sánchez-Mateos, P.; Samaniego, R.; et al. Estradiol impairs epithelial CXCL1 gradient in the cervix to delay neutrophil transepithelial migration during insemination. J. Reprod. Immunol. 2019, 132, 9–15. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward and Reverse Sequences (5′-3′) | Source |
|---|---|---|
| Ywhaz | Fw: AACTTGACATTGTGGACATCGG Rv: AAAGGTTGGAAGGCCGGTTA | this study |
| Ubc | Fw: ACACCAAGAAGGTCAAACAGGA Rv: CACCTCCCCATCAAACCCAA | [50] |
| B2m | Fw: GTCACCTGGGACCGAGACAT Rv: AGAAGATGGTGTGCTCATTGC | [51] |
| Ar | Fw: CTTATGGGGACATGCGTTTGG Rv: GCTCCGTAGTGACAACCAGA | this study |
| Fdps | Fw: GCAGACTCTCGACCTCATCACA Rv: CCCATCAATTCCAGCCATG | [52] |
| Camkk2 | Fw: AGAACTGCACACTGGTCGAG Rv: CCGGCTACCTTCAAATGGGT | [53] |
| Cxcl1 | Fw: GCCACACTCAAGAATGGTCG Rv: TGGGGACACCCTTTAGCATC | [54] |
| Tnfa | Fw: GACCCTCACACTCAGATCATCTTCT Rv: TGCTACGACGTGGGCTACG | [55] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Turniak-Kusy, M.; Studzian, M.; Szpakowski, P.; Kuchta, P.; Smietanka, K.; Mattern, C.; Pulaski, L.; Bielecki, B. Testosterone Inhibits Secretion of the Pro-Inflammatory Chemokine CXCL1 from Astrocytes. Curr. Issues Mol. Biol. 2024, 46, 2105-2118. https://doi.org/10.3390/cimb46030135
Turniak-Kusy M, Studzian M, Szpakowski P, Kuchta P, Smietanka K, Mattern C, Pulaski L, Bielecki B. Testosterone Inhibits Secretion of the Pro-Inflammatory Chemokine CXCL1 from Astrocytes. Current Issues in Molecular Biology. 2024; 46(3):2105-2118. https://doi.org/10.3390/cimb46030135
Chicago/Turabian StyleTurniak-Kusy, Malgorzata, Maciej Studzian, Piotr Szpakowski, Piotr Kuchta, Kaja Smietanka, Claudia Mattern, Lukasz Pulaski, and Bartosz Bielecki. 2024. "Testosterone Inhibits Secretion of the Pro-Inflammatory Chemokine CXCL1 from Astrocytes" Current Issues in Molecular Biology 46, no. 3: 2105-2118. https://doi.org/10.3390/cimb46030135