
Identification of m6A Modification Regulated by Dysregulated circRNAs in Decidua of Recurrent Pregnancy Loss


Abstract
:1. Introduction
2. Materials and Methods
2.1. Human Sample
2.2. CircRNA-Seq
2.3. CircRNA–miRNA–mRNA Network Construction
2.4. RT-PCR
2.5. Statistical Analysis
3. Results
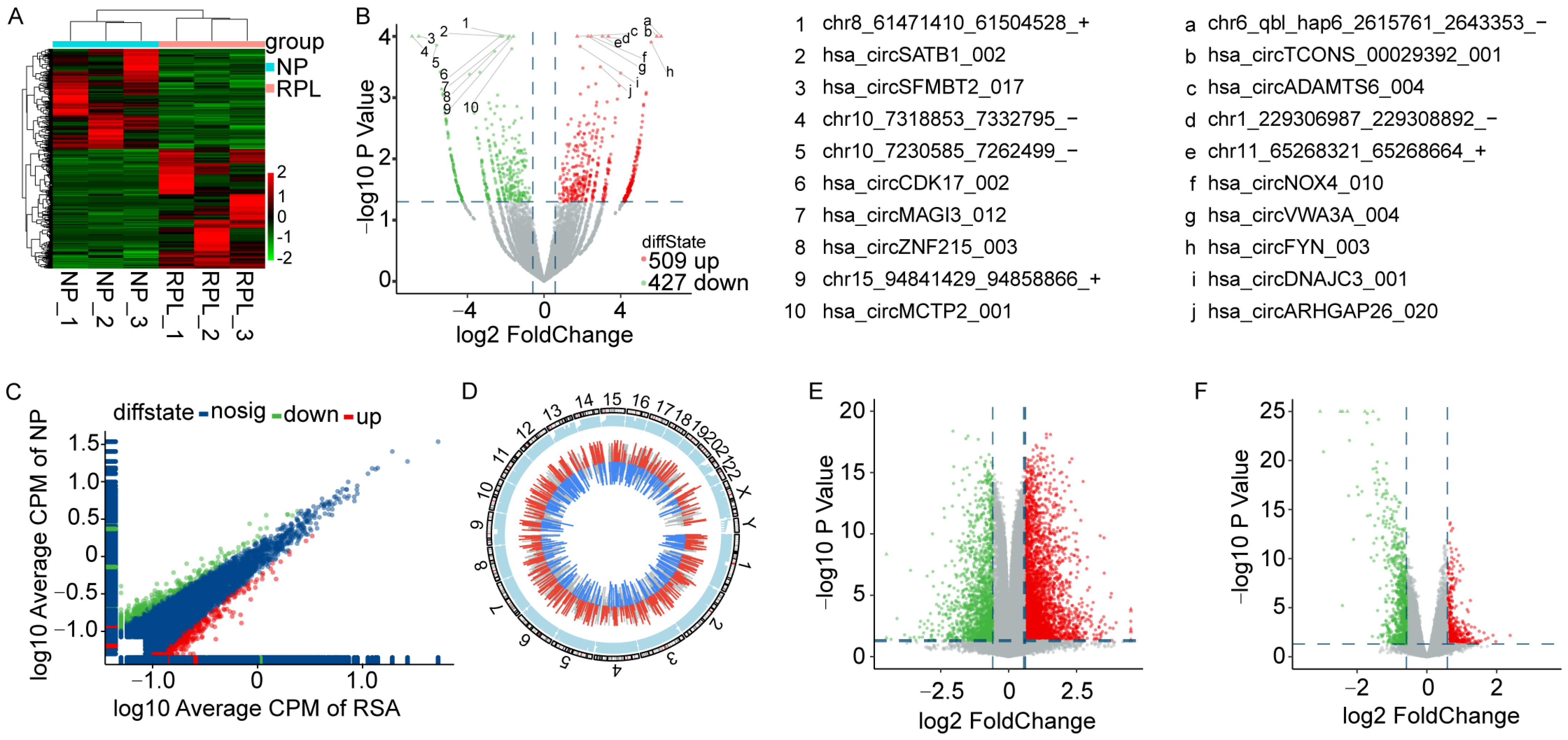
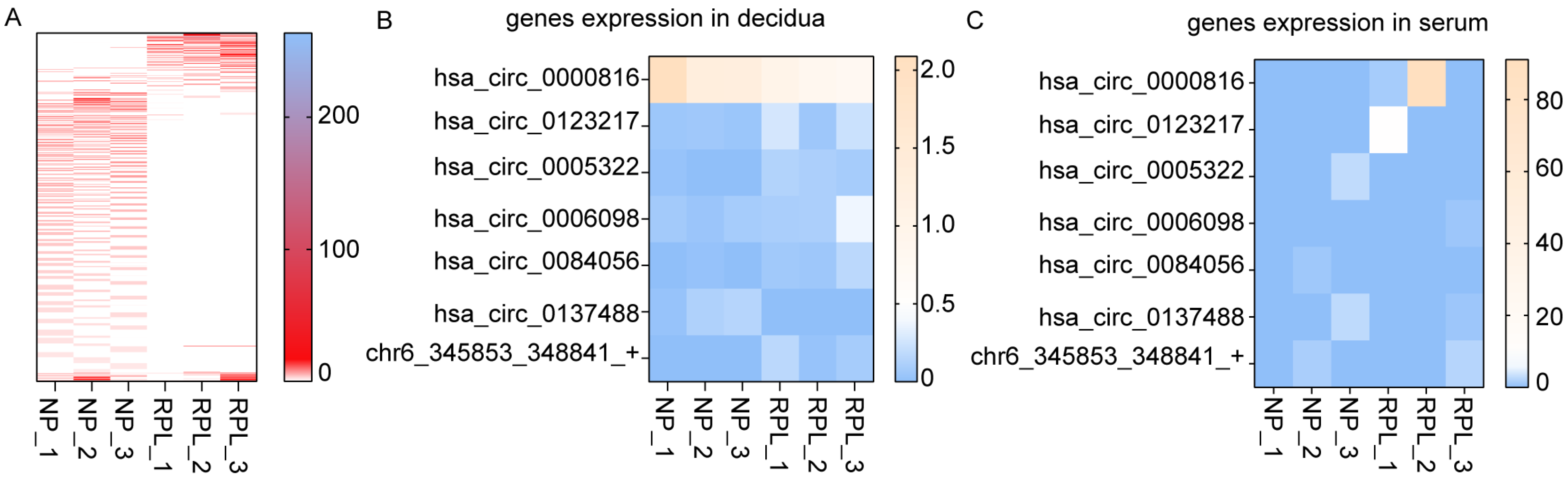
3.1. Expression Profiles of circRNAs and mRNAs in Decidua from RPL Patients
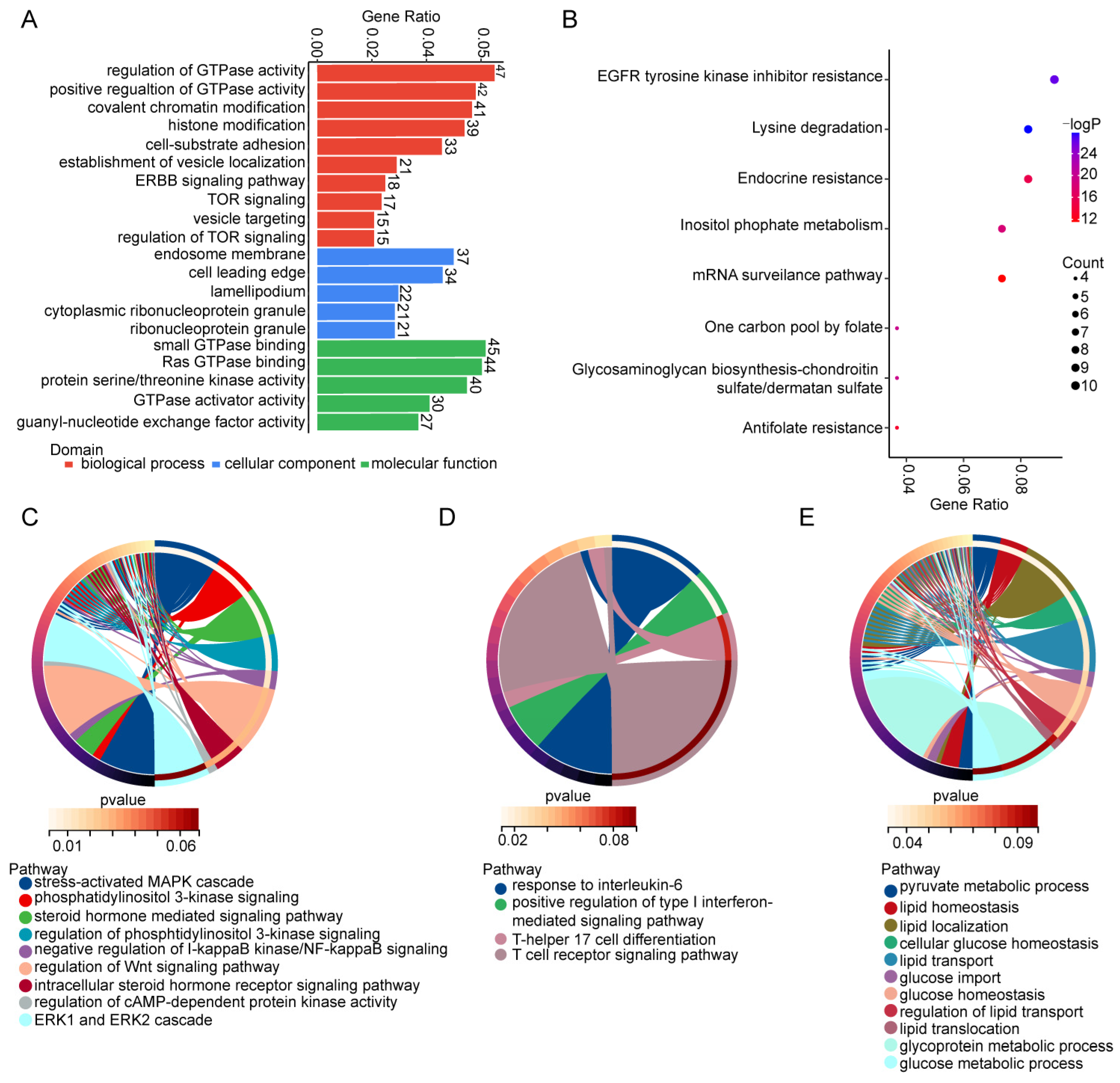
3.2. GO and KEGG Pathway Analyses of Parental Genes of Differentially Expressed circRNAs in Decidua
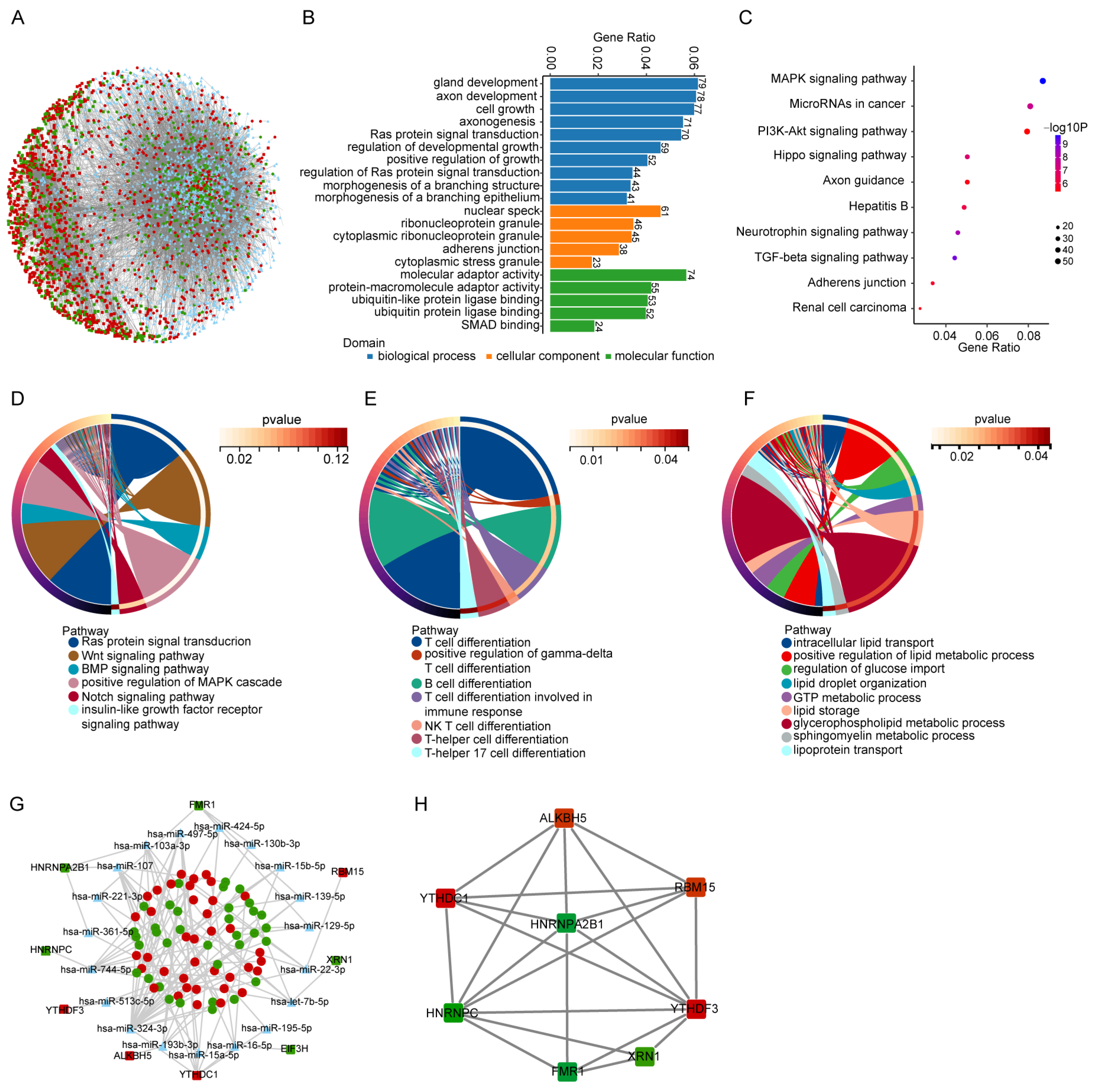
3.3. Prediction of ceRNA Network and Functional Analyses of m6A Modification in This Network
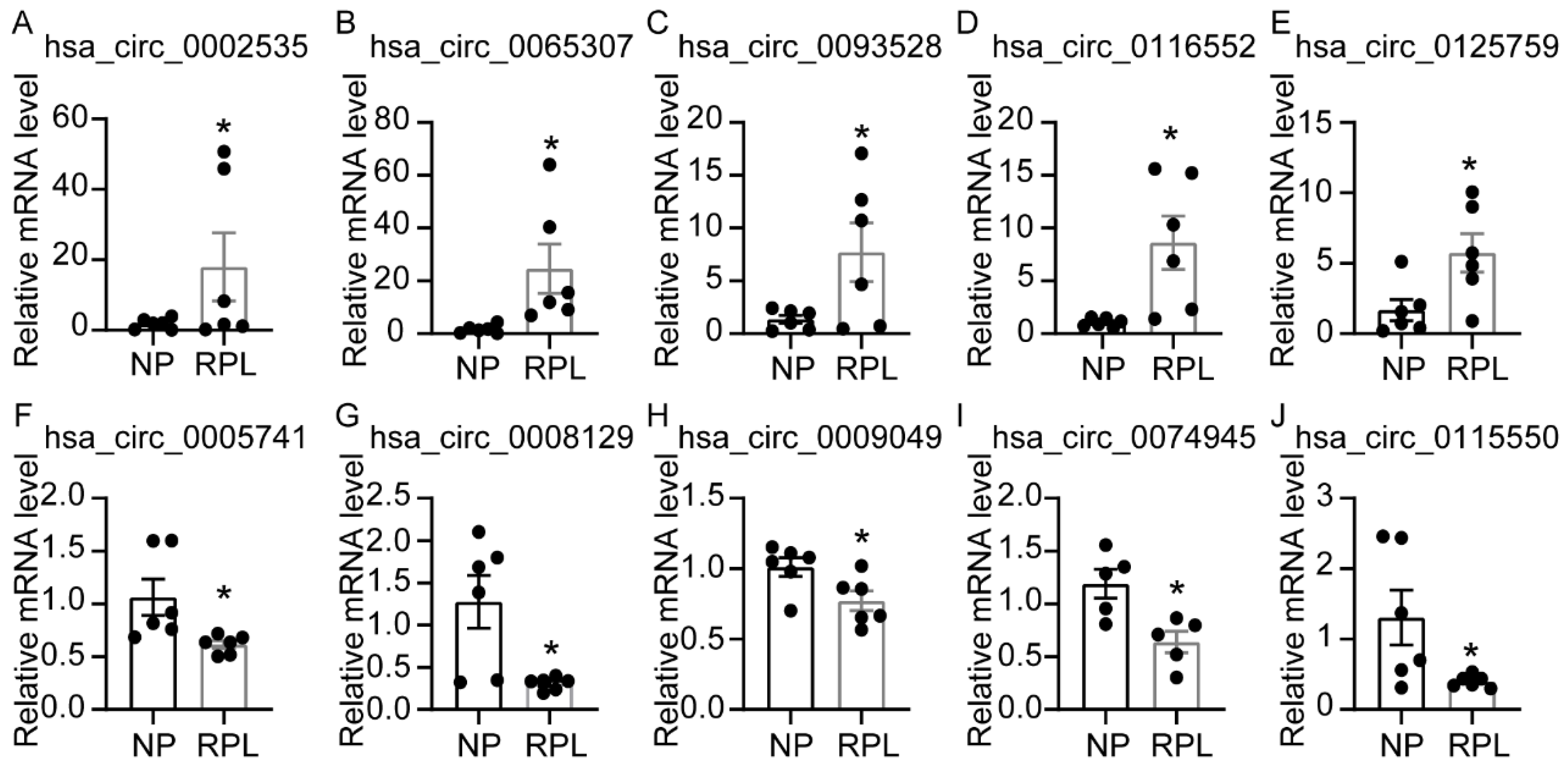
3.4. Validation of Differentially Expressed circRNAs
3.5. Prediction of Biomarker in Serum of Peripheral Blood
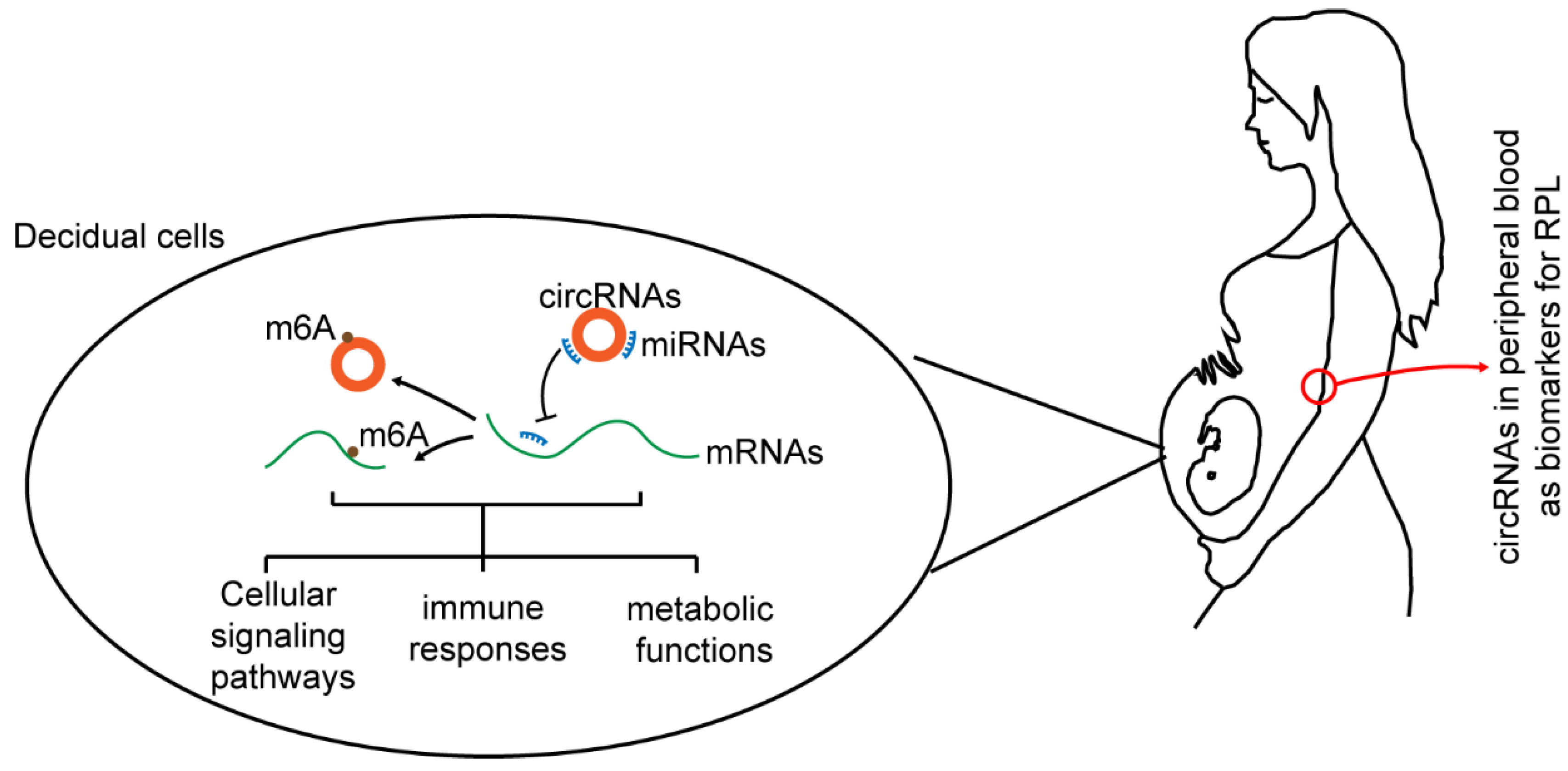
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dimitriadis, E.; Menkhorst, E.; Saito, S.; Kutteh, W.H.; Brosens, J.J. Recurrent pregnancy loss. Nat. Rev. Dis. Primers 2020, 6, 98. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Hou, C.C.; Luo, L.F.; Hu, Y.J.; Yang, W.X. Endometrial stromal cells and decidualized stromal cells: Origins, transformation and functions. Gene 2014, 551, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.J.; Sung, N.Y.; Gilman-Sachs, A.; Kwak-Kim, J. T Helper (Th) Cell Profiles in Pregnancy and Recurrent Pregnancy Losses: Th1/Th2/Th9/Th17/Th22/Tfh Cells. Front. Immunol. 2020, 11, 2025. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Xu, Q.H.; Ma, L.N.; Luo, J.; Muyayalo, K.P.; Wang, L.L.; Huang, D.H.; Xiao, X.J.; Cheng, S.B.; Mor, G.; et al. Trophoblast-derived Lactic Acid Orchestrates Decidual Macrophage Differentiation via SRC/LDHA Signaling in Early Pregnancy. Int. J. Biol. Sci. 2022, 18, 599–616. [Google Scholar] [CrossRef]
- Meng, X.; Chen, C.; Qian, J.; Cui, L.; Wang, S. Energy metabolism and maternal-fetal tolerance working in decidualization. Front. Immunol. 2023, 14, 1203719. [Google Scholar] [CrossRef]
- Li, C.; Chen, X.; Liu, X.; Liu, X.; He, J.; Ding, Y.; Tong, C.; Peng, C.; Wang, Y.; Gao, R. CircRNA expression profiles in decidual tissue of patients with early recurrent miscarriage. Genes Dis. 2020, 7, 414–423. [Google Scholar] [CrossRef]
- Quan, G.B.; Li, J.L. Circular RNAs: Biogenesis, expression and their potential roles in reproduction. J. Ovarian Res. 2018, 11, 9. [Google Scholar] [CrossRef]
- Li, Y.; Zheng, Q.P.; Bao, C.Y.; Li, S.Y.; Guo, W.J.; Zhao, J.; Chen, D.; Gu, J.R.; He, X.H.; Huang, S.L. Circular RNA is enriched and stable in exosomes: A promising biomarker for cancer diagnosis. Cell Res. 2015, 25, 981–984. [Google Scholar] [CrossRef]
- Gao, Q.Y.; Wang, T.J.; Pan, L.X.; Qian, C.; Wang, J.; Xin, Q.; Liu, Y.J.; Zhang, Z.G.; Xu, Y.P.; He, X.J.; et al. Circular RNAs: Novel potential regulators in embryogenesis, female infertility, and pregnancy-related diseases. J. Cell Physiol. 2021, 236, 7223–7241. [Google Scholar] [CrossRef]
- Arthurs, A.L.; Jankovic-Karasoulos, T.; Smith, M.D.; Roberts, C.T. Circular RNAs in Pregnancy and the Placenta. Int. J. Mol. Sci. 2022, 23, 4551. [Google Scholar] [CrossRef]
- Zhou, W.B.; Wang, H.Y.; Wu, X.; Long, W.; Zheng, F.X.; Kong, J.; Yu, B. The profile analysis of circular RNAs in human placenta of preeclampsia. Exp. Biol. Med. 2018, 243, 1109–1117. [Google Scholar] [CrossRef] [PubMed]
- Fan, Z.Y.; Wang, Q.M.; Deng, H. Circ_0011460 upregulates HTRA1 expression by sponging miR-762 to suppress HTR8/SVneo cell growth, migration, and invasion. Am. J. Reprod. Immunol. 2021, 86, e13485. [Google Scholar] [CrossRef]
- Yao, P.; Hu, G.H.; Niu, H.L. Hsa_circ_0074371 Regulates Proliferation, Apoptosis, Migration, and Invasion via the miR-582-3p/LRP6 Axis in Trophoblast Cells. Biochem. Genet. 2022, 60, 267–285. [Google Scholar] [CrossRef] [PubMed]
- Zou, H.; Mao, Q.H. Circ_0037078 promotes trophoblast cell proliferation, migration, invasion and angiogenesis by miR-576-5p/IL1RAP axis. Am. J. Reprod. Immunol. 2022, 87, e13507. [Google Scholar] [CrossRef]
- Liu, B.W.; Liu, J.; Qiu, Y.; Chen, J.; Yang, J. MITA Promotes Macrophage Proinflammatory Polarization and Its circRNA-Related Regulatory Mechanism in Recurrent Miscarriage. Int. J. Mol. Sci. 2023, 24, 9545. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Do, X.Y.; Chen, C.Y.; Chen, C.; Liu, C.; Xu, M.M.; Zhao, S.Q.; Shen, B.; Gao, Y.W.; Han, D.L.; et al. N-6-methyladenosine of chromosome-associated regulatory RNA regulates chromatin state and transcription. Science 2020, 367, 580–586. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zheng, J.; Liao, A. The regulation and potential roles of m6A modifications in early embryonic development and immune tolerance at the maternal-fetal interface. Front. Immunol. 2022, 13, 988130. [Google Scholar] [CrossRef]
- Huang, N.; Gao, Y.; Zhang, M.; Guo, L.; Qin, L.; Liao, S.; Wang, H. METTL3-Mediated m6A RNA Methylation of ZBTB4 Interferes With Trophoblast Invasion and Maybe Involved in RSA. Front. Cell Dev. Biol. 2022, 10, 894810. [Google Scholar] [CrossRef]
- Luo, Y.; Chen, J.; Cui, Y.; Fang, F.; Zhang, Z.; Hu, L.; Chen, X.; Li, Z.; Li, L.; Chen, L. Transcriptome-wide high-throughput m6A sequencing of differential m6A methylation patterns in the decidual tissues from RSA patients. FASEB J. 2023, 37, e22802. [Google Scholar] [CrossRef]
- Yang, J.; Tong, J.; Li, H.; Lv, S.; Li, W.; Zhang, C.; Di, W. Decreased expression of m6A demethylase ALKBH5 in decidua contributes to preeclampsia via m6A-CORIN-HuR pathway. Genes Dis. 2023, 10, 650–653. [Google Scholar] [CrossRef]
- Ledee, N.; Munaut, C.; Aubert, J.; Serazin, V.; Rahmati, M.; Chaouat, G.; Sandra, O.; Foidart, J.M. Specific and extensive endometrial deregulation is present before conception in IVF/ICSI repeated implantation failures (IF) or recurrent miscarriages. J. Pathol. 2011, 225, 554–564. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Liu, M.; Nie, X.; Zhang, Y.; Chen, Y.; Zhu, L.; Chen, X.; Chen, L.; Chen, H.; Zhang, J. NOD1 and NOD2 control the invasiveness of trophoblast cells via the MAPK/p38 signaling pathway in human first-trimester pregnancy. Placenta 2015, 36, 652–660. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Ding, J.; Wang, J.; Yin, T.; Zhang, Y.; Yang, J. CXCL5 Downregulation in Villous Tissue Is Correlated with Recurrent Spontaneous Abortion. Front. Immunol. 2021, 12, 717483. [Google Scholar] [CrossRef] [PubMed]
- Lou, Y.; Fu, Z.; Tian, Y.; Hu, M.; Wang, Q.; Zhou, Y.; Wang, N.; Zhang, Q.; Jin, F. Estrogen-sensitive activation of SGK1 induces M2 macrophages with anti-inflammatory properties and a Th2 response at the maternal–fetal interface. Reprod. Biol. Endocrinol. 2023, 21, 50. [Google Scholar] [CrossRef] [PubMed]
- Shukla, V.; Kaushal, J.B.; Sankhwar, P.; Manohar, M.; Dwivedi, A. Inhibition of TPPP3 attenuates β-catenin/NF-κB/COX-2 signaling in endometrial stromal cells and impairs decidualization. J. Endocrinol. 2019, 240, 417–429. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Gomez, T.; Castillo-Marco, N.; Clemente-Ciscar, M.; Cordero, T.; Munoz-Blat, I.; Amadoz, A.; Jimenez-Almazan, J.; Monfort-Ortiz, R.; Climent, R.; Perales-Marin, A.; et al. Disrupted PGR-B and ESR1 signaling underlies defective decidualization linked to severe preeclampsia. eLife 2021, 10, e70753. [Google Scholar] [CrossRef]
- Li, B.; Yan, Y.-P.; Liang, C.; He, Y.-Y.; Wang, Y.; Li, M.-Y.; Chen, S.-T.; Li, Y.; Liu, A.-X.; Yan, G.-J.; et al. Primary Cilia Restrain PI3K-AKT Signaling to Orchestrate Human Decidualization. Int. J. Mol. Sci. 2022, 23, 15573. [Google Scholar] [CrossRef]
- Yang, D.; Dai, F.; Wang, L.; Cai, S.; Zhang, Y.; Diao, L.; Cheng, Y. HSP70 regulates lipid metabolism of decidual macrophages to maintain normal pregnancy. J. Reprod. Immunol. 2023, 156, 103829. [Google Scholar] [CrossRef]
- D’Ippolito, S.; Capozzi, A.; Scambia, G.; Sorge, R.; Lello, S.; Simone, N.D. Glucose/insulin metabolism and vitamin D in women with recurrent pregnancy loss. Am. J. Reprod. Immunol. 2021, 87, e13505. [Google Scholar] [CrossRef]
- Jiang, M.; Lash, G.E.; Zhao, X.; Long, Y.; Guo, C.; Yang, H. CircRNA-0004904, CircRNA-0001855, and PAPP-A: Potential Novel Biomarkers for the Prediction of Preeclampsia. Cell. Physiol. Biochem. 2018, 46, 2576–2586. [Google Scholar] [CrossRef]
- Hu, X.P.; Ao, J.P.; Li, X.Y.; Zhang, H.J.; Wu, J.; Cheng, W.W. Competing endogenous RNA expression profiling in pre-eclampsia identifies hsa_circ_0036877 as a potential novel blood biomarker for early pre-eclampsia. Clin. Epigenetics 2018, 10, 48. [Google Scholar] [CrossRef] [PubMed]
- Haim-Vilmovsky, L.; Henriksson, J.; Walker, J.A.; Miao, Z.C.; Natan, E.; Kar, G.; Clare, S.; Barlow, J.L.; Charidemou, E.; Mamanova, L.; et al. Mapping Rora expression in resting and activated CD4+ T cells. PLoS ONE 2021, 16, e0251233. [Google Scholar] [CrossRef] [PubMed]
- Cohen, C.J.; Crome, S.Q.; MacDonald, K.G.; Dai, E.L.; Mager, D.L.; Levings, M.K. Human Th1 and Th17 Cells Exhibit Epigenetic Stability at Signature Cytokine and Transcription Factor Loci. J. Immunol. 2011, 187, 5615–5626. [Google Scholar] [CrossRef] [PubMed]
- Sendinc, E.; Shi, Y. RNA m6A methylation across the transcriptome. Mol. Cell 2023, 83, 428–441. [Google Scholar] [CrossRef]
- Huo, J.H.; Chen, Q.; Zhang, Y.T.; Li, N.; Fu, Z.Y.; Ma, N.; Zheng, N.; Cui, N.; Li, L. Molecular subtype identification and predictive power of N6-methyladenosine regulator in unexplained recurrent pregnancy loss. Front. Genet. 2022, 13, 925652. [Google Scholar] [CrossRef]
- Li, A.; Chen, Y.-S.; Ping, X.-L.; Yang, X.; Xiao, W.; Yang, Y.; Sun, H.-Y.; Zhu, Q.; Baidya, P.; Wang, X.; et al. Cytoplasmic m6A reader YTHDF3 promotes mRNA translation. Cell Res. 2017, 27, 444–447. [Google Scholar] [CrossRef]
- Prefumo, F. Circular RNA and pre-eclampsia: On the long road from the laboratory to the bedside. BJOG Int. J. Obstet. Gynaecol. 2016, 123, 2119. [Google Scholar] [CrossRef]
- Cui, Y.M.; Liu, J.W.; Liu, L.N.; Ma, X.Z.; Gui, Y.; Liu, H.; Zhao, W. m(6)A-modified circFOXK2 targets GLUT1 to accelerate oral squamous cell carcinoma aerobic glycolysis. Cancer Gene Ther. 2023, 30, 163–171. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subjects b | NP (n = 9) | RPL (n = 9) | p-Value |
|---|---|---|---|
| Age mean (years) a | 29.89 ± 2.43 | 30.33 ± 1.63 | 0.88 |
| Age range (years) | 22–40 | 23–36 | - |
| Previous pregnancy loss a | 0 | 2.778 ± 0.28 | <0.0001 |
| Previous normal births a | 1.444 ± 0.24 | 0 | <0.0001 |
| Pregnancy week (sample collected) a | 6.21 ± 0.24 | 6.43 ± 0.14 | 0.44 |
| Primer Name | Forward | Reverse |
|---|---|---|
| hsa_circ_0002535 | CTCCTTGGCACTGTGCTTCC | GAGCCCAGCAGTTGCCC |
| hsa_circ_0065307 | CAAACGACATCAGGCAAAGTGT | GTAGGTCAAACCTCCGCCAT |
| hsa_circ_0093528 | ACAGATGGCGTAATGGGGTG | AACTGCTACATGTCTGTTGGA |
| hsa_circ_0116552 | CTCGAAGGATGCGCAGAGAT | GCAGATCTCAACACCATTAAGTACC |
| hsa_circ_0125759 | GGAAGTGAAGAGGCTGACATGA | GGAAGTGAAGAGGCTGACATGA |
| hsa_circ_0005741 | GGTCCATTGCTATCAGCCCA | CCTTCACTGGGACACTGGTC |
| hsa_circ_0008129 | AGCTACTTGTAGAGGCTTATTGTGT | AGCTACTTGTAGAGGCTTATTGTGT |
| hsa_circ_0009049 | CCTTCTTCTCTGGCCATGC | ACTCAAGAAGTGAGGACGCA |
| hsa_circ_0074945 | TCTACGACCCCAACAAGCAA | AGCTCAAGGATTCGTGAAGACC |
| hsa_circ_0115550 | GAAACACAGGACATCGCTGC | AGACACTGAAGATCAGGCCAAC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, L.; Shi, M.; Meng, X.; Qian, J.; Wang, S. Identification of m6A Modification Regulated by Dysregulated circRNAs in Decidua of Recurrent Pregnancy Loss. Curr. Issues Mol. Biol. 2023, 45, 8767-8779. https://doi.org/10.3390/cimb45110551
Cui L, Shi M, Meng X, Qian J, Wang S. Identification of m6A Modification Regulated by Dysregulated circRNAs in Decidua of Recurrent Pregnancy Loss. Current Issues in Molecular Biology. 2023; 45(11):8767-8779. https://doi.org/10.3390/cimb45110551
Chicago/Turabian StyleCui, Liyuan, Minfeng Shi, Xinhang Meng, Jinfeng Qian, and Songcun Wang. 2023. "Identification of m6A Modification Regulated by Dysregulated circRNAs in Decidua of Recurrent Pregnancy Loss" Current Issues in Molecular Biology 45, no. 11: 8767-8779. https://doi.org/10.3390/cimb45110551