
Disrupted Balance of the Oxidant–Antioxidant System in the Pathophysiology of Female Reproduction: Oxidative Stress and Adverse Pregnancy Outcomes
Abstract
:1. Introduction and Methods
2. Oxidative Stress: Basic Mechanisms
3. Effect of Oxidative Stress on Reproductive Cells
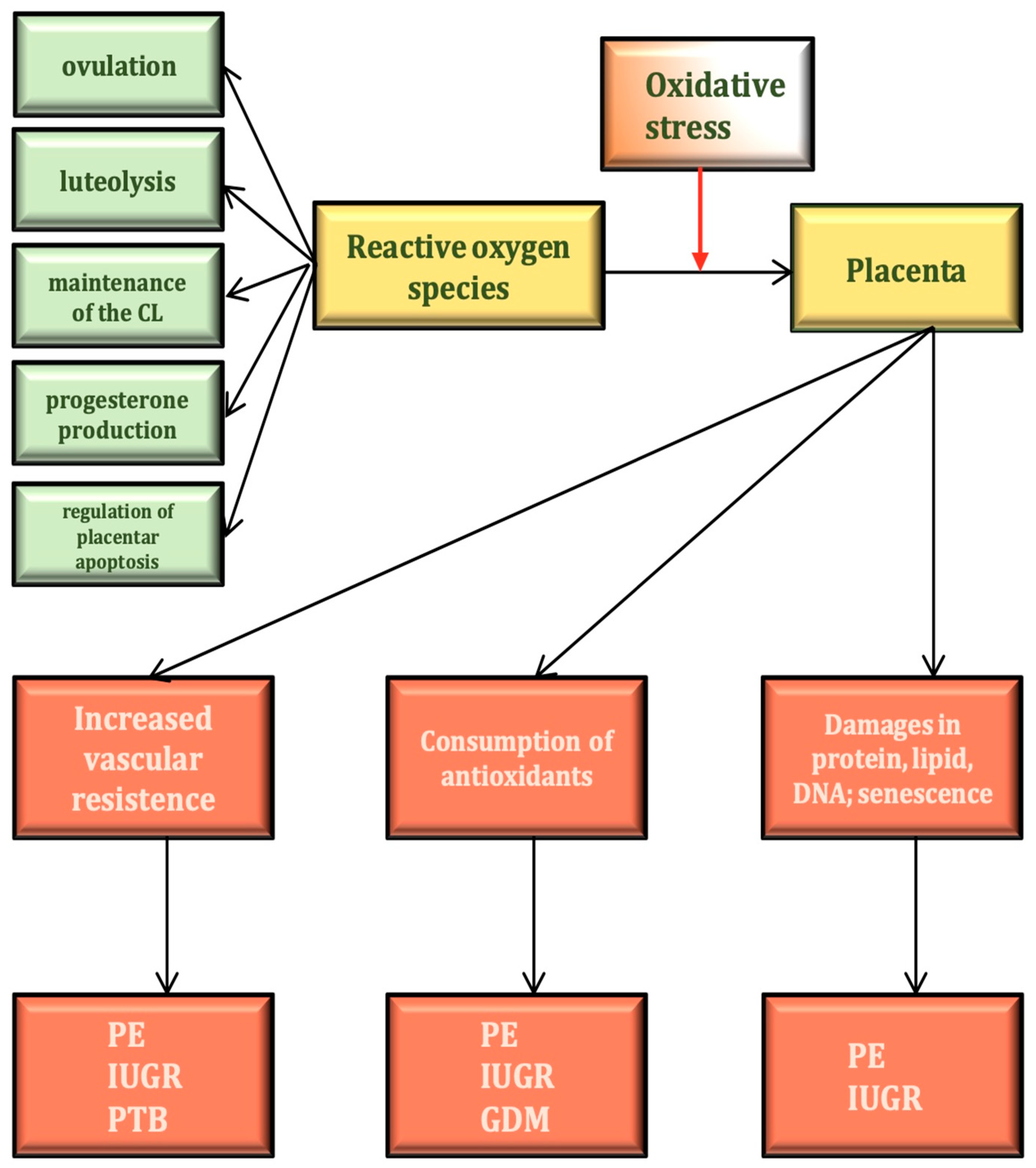
4. The Physiological Role of ROS in the Female Reproductive Process
5. Markers of Oxidative Stress in Obstetrics
6. Regulation of Oxidant–Antioxidant Balance during Physiological Pregnancy
7. Oxidative Stress and Epigenetic Effects
8. Oxidative Stress and the Physiology and Pathophysiology of the Placenta
9. Oxidative Stress and Early Miscarriage
10. Oxidative Stress and Hypertensive Disorders during Pregnancy
11. Oxidative Stress and IUGR
12. Oxidative Stress and Gestational Diabetes Mellitus
13. Oxidative Stress and Premature Delivery
14. Oxidative Stress and Twin Pregnancy
15. Antioxidant Supplementation during Pregnancy
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lu, J.; Wang, Z.; Cao, J.; Chen, Y.; Dong, Y. A novel and compact review on the role of oxidative stress in female reproduction. Reprod. Biol. Endocrinol. 2018, 20, 80–97. [Google Scholar] [CrossRef]
- Agarwal, A.; Gupta, S.; Sharma, R.K. Role of oxidative stress in female reproduction. Reprod. Biol. Endocrinol. 2005, 3, 28–48. [Google Scholar] [CrossRef]
- Ruder, E.H.; Hartman, T.J.; Goldman, M.B. Impact of oxidative stress on female fertility. Curr. Opin. Obstet. Gynecol. 2009, 21, 219–222. [Google Scholar] [CrossRef] [PubMed]
- Hussain, T.; Murtaza, G.; Metwally, E.; Kalhoro, D.H.; Kalhoro, M.S.; Rahu, B.A.; Sahito, R.G.A.; Yin, Y.; Yang, H.; Chughtai, M.I.; et al. The Role of Oxidative Stress and Antioxidant Balance in Pregnancy. Mediat. Inflamm. 2021, 27, 9962860. [Google Scholar] [CrossRef]
- Aitken, R.J. Impact of oxidative stress on male and female germ cells: Implications for fertility. Reproduction 2020, 159, R189–R201. [Google Scholar] [CrossRef]
- Tarín, J.J.; Vendrell, F.J.; Ten, J.; Cano, A. Antioxidant therapy counteracts the disturbing effects of diamide and maternal aging on meiotic division and chromosomal segregation in mouse oocytes. Mol. Hum. Reprod. 1998, 4, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Jeelani, R.; Chatzicharalampous, C.; Kohan-Ghadr, H.R.; Awonuga, A.; Joshi, N.; Morris, R.T.; Abu-Soud, H.M. Acrolein, a commonly found environmental toxin, causes oocyte mitochondrial dysfunction and negatively affects embryo development. Free Radic. Res. 2018, 52, 929–938. [Google Scholar] [CrossRef]
- Petca, A.; Bot, M.; Maru, N.; Calo, I.G.; Borislavschi, A.; Dumitrascu, M.C.; Petca, R.C.; Sandru, F.; Zvanca, M.E. Benefits of α-lipoic acid in high-risk pregnancies (Review). Exp. Ther. Med. 2021, 22, 1232–1239. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, M. Oxygen radicals-superoxide dismutase system and reproduction medicine. Nihon Sanka Fujinka Gakkai Zasshi 1993, 45, 842–848. [Google Scholar]
- Shkolnik, K.; Tadmor, A.; Ben-Dor, S.; Nevo, N.; Galiani, D.; Dekel, N. Reactive oxygen species are indispensable in ovulation. Proc. Natl. Acad. Sci. USA 2011, 108, 1462–1467. [Google Scholar] [CrossRef]
- Sugino, N. Reactive oxygen species in ovarian physiology. Reprod. Med. Biol. 2005, 4, 31–44. [Google Scholar] [CrossRef]
- Agarwal, A.; Aponte-Mellado, A.; Premkumar, B.J.; Shaman, A.; Gupta, S. The effects of oxidative stress on female reproduction: A review. Reprod. Biol. Endocrinol. 2012, 29, 49–79. [Google Scholar] [CrossRef] [PubMed]
- Zarkovic, N. Roles and Functions of ROS and RNS in Cellular Physiology and Pathology. Cells 2020, 9, 767–771. [Google Scholar] [CrossRef]
- Di Meo, S.; Reed, T.T.; Venditti, P.; Victor, V.M. Role of ROS and RNS Sources in Physiological and Pathological Conditions. Oxid. Med. Cell Longev. 2016, 2016, 1245049. [Google Scholar] [CrossRef]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef] [PubMed]
- Aouache, R.; Biquard, L.; Vaiman, D.; Miralles, F. Oxidative Stress in Preeclampsia and Placental Diseases. Int. J. Mol. Sci. 2018, 19, 1496. [Google Scholar] [CrossRef]
- Tenório, M.B.; Ferreira, R.C.; Moura, F.A.; Bueno, N.B.; de Oliveira, A.C.M.; Goulart, M.O.F. Cross-Talk between Oxidative Stress and Inflammation in Preeclampsia. Oxid. Med. Cell Longev. 2019, 2019, 8238727. [Google Scholar] [CrossRef] [PubMed]
- Duhig, K.; Chappell, L.C.; Shennan, A.H. Oxidative stress in pregnancy and reproduction. Obstet. Med. 2016, 9, 113–116. [Google Scholar] [CrossRef]
- Drejza, M.A.; Rylewicz, K.; Majcherek, E.; Gross-Tyrkin, K.; Mizgier, M.; Plagens-Rotman, K.; Wójcik, M.; Panecka-Mysza, K.; Pisarska-Krawczyk, M.; Kędzia, W.; et al. Markers of Oxidative Stress in Obstetrics and Gynaecology-A Systematic Literature Review. Antioxidants 2022, 11, 1477. [Google Scholar] [CrossRef]
- Schoots, M.H.; Bourgonje, M.F.; Bourgonje, A.R.; Prins, J.R.; van Hoorn, E.G.M.; Abdulle, A.E.; Muller Kobold, A.C.; van der Heide, M.; Hillebrands, J.L.; van Goor, H.; et al. Oxidative stress biomarkers in fetal growth restriction with and without preeclampsia. Placenta 2021, 115, 87–96. [Google Scholar] [CrossRef]
- Cortese-Krott, M.M.; Koning, A.; Kuhnle, G.G.C.; Nagy, P.; Bianco, C.L.; Pasch, A.; Wink, D.A.; Fukuto, J.M.; Jackson, A.A.; van Goor, H.; et al. The reactive species interactome: Evolutionary emergence, biological significance, and opportunities for redox metabolomics and personalized medicine. Antioxid. Redox Signal. 2017, 27, 684–712. [Google Scholar] [CrossRef] [PubMed]
- Erol, S.A.; Tanacan, A.; Altinboga, O.; Ozturk, F.H.; Ozgu, B.S.; Tasci, Y.; Neselioglu, S.; Erel, O.; Sahin, D. Evaluation of fetal serum thiol/disulfide homeostasis and ischemia-modified albumin levels in fetal distress. Fetal Pediatr. Pathol. 2022, 41, 426–435. [Google Scholar] [CrossRef]
- Rossi, A.; Bortolotti, N.; Vescovo, S.; Romanello, I.; Forzano, L.; Londero, A.P.; Ambrosini, G.; Marchesoni, D.; Curcio, F. Ischemia-modified albumin in pregnancy. Eur. J. Obstet. Gynecol. Reprod. Biol. 2013, 170, 348–351. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Yang, Y.; Chen, R.; Wei, Y.; Feng, Y.; Zheng, W.; Liao, H.; Zhang, Z. Aberrant expression of oxidative stress related proteins affects the pregnancy outcome of gestational diabetes mellitus patients. Am. J. Transl. Res. 2019, 11, 269–279. [Google Scholar] [PubMed]
- Saifi, H.; Mabrouk, Y.; Saifi, R.; Benabdelkader, M.; Saidi, M. Influence of selenium supplementation on carbohydrate metabolism and oxidative stress in pregnant women with gestational diabetes mellitus. J. Med. Biochem. 2020, 23, 191–198. [Google Scholar] [CrossRef]
- Gong, Y.; Zhang, K.; Xiong, D.; Wei, J.; Tan, H.; Qin, S. Growth hormone alleviates oxidative stress and improves the IVF outcomes of poor ovarian responders: A randomized controlled trial. Reprod. Biol. Endocrinol. 2020, 18, 91. [Google Scholar] [CrossRef] [PubMed]
- Toljic, M.; Egic, A.; Munjas, J.; Karadzov Orlic, N.; Milovanovic, Z.; Radenkovic, A.; Vuceljic, J.; Joksic, I. Increased oxidative stress and cytokinesis-block micronucleus cytome assay parameters in pregnant women with gestational diabetes mellitus and gestational arterial hypertension. Reprod. Toxicol. 2017, 71, 55–62. [Google Scholar] [CrossRef]
- Hajifaraji, M.; Jahanjou, F.; Abbasalizadeh, F.; Aghamohammadzadeh, N.; Abbasi, M.M.; Dolatkhah, N. Effect of probiotic supplements in women with gestational diabetes mellitus on inflammation and oxidative stress biomarkers: A randomized clinical trial. Asia Pac. J. Clin. Nutr. 2018, 27, 581–591. [Google Scholar] [CrossRef]
- Zygula, A.; Kosinski, P.; Zwierzchowska, A.; Sochacka, M.; Wroczynski, P.; Makarewicz-Wujec, M.; Pietrzak, B.; Wielgos, M.; Rzentala, M.; Giebultowicz, J. Oxidative stress markers in saliva and plasma differ between diet-controlled and insulin-controlled gestational diabetes mellitus. Diabetes Res. Clin. Pract. 2019, 148, 72–80. [Google Scholar] [CrossRef]
- Rodrigues, F.; de Lucca, L.; Neme, W.S.; Gonçalves, T.L. Influence of gestational diabetes on the activity of δ-aminolevulinate dehydratase and oxidative stress biomarkers. Redox Rep. 2018, 23, 63–67. [Google Scholar] [CrossRef]
- Li, H.; Dong, A.; Lv, X. Advanced glycation end products and adipocytokines and oxidative stress in placental tissues of pregnant women with gestational diabetes mellitus. Exp. Ther. Med. 2019, 18, 685–691. [Google Scholar] [CrossRef]
- Bulut, A.; Akca, G.; Keskin Aktan, A.; Akbulut, K.G.; Babül, A. The significance of blood and salivary oxidative stress markers and chemerin in gestational diabetes mellitus. Taiwan J. Obstet. Gynecol. 2021, 60, 695–699. [Google Scholar] [CrossRef]
- Gunasegaran, P.; Tahmina, S.; Daniel, M.; Nanda, S.K. Role of vitamin D-calcium supplementation on metabolic profile and oxidative stress in gestational diabetes mellitus: A randomized controlled trial. J. Obstet. Gynaecol. Res. 2021, 47, 1016–1022. [Google Scholar] [CrossRef]
- Soydinc, H.E.; Sak, M.E.; Evliyaoglu, O.; Evsen, M.S.; Turgut, A.; Özler, A.; Yıldız, İ.; Gul, T. Prolidase, matrix metalloproteinases 1 and 13 activity, oxidative-antioxidative status as a marker of preterm premature rupture of membranes and chorioamnionitis in maternal vaginal washing fluids. Int. J. Med. Sci. 2013, 10, 1344–1351. [Google Scholar] [CrossRef] [PubMed]
- Moore, T.A.; Ahmad, I.M.; Zimmerman, M.C. Oxidative Stress and Preterm Birth: An Integrative Review. Biol. Res. Nurs. 2018, 20, 497–512. [Google Scholar] [CrossRef]
- Clerici, G.; Slavescu, C.; Fiengo, S.; Kanninen, T.T.; Romanelli, M.; Biondi, R.; Di Renzo, G.C. Oxidative stress in pathological pregnancies. J. Obstet. Gynaecol. 2012, 32, 124–127. [Google Scholar] [CrossRef]
- Ilhan, N.; Celik, E.; Kumbak, B. Maternal plasma levels of interleukin-6, C-reactive protein, vitamins C, E and A, 8-isoprostane and oxidative status in women with preterm premature rupture of membranes. J. Matern. Fetal Neonatal. Med. 2015, 28, 316–319. [Google Scholar] [CrossRef]
- Bak, A.; Roszkowski, K. Oxidative stress in pregnant women. Arch. Perinat. Med. 2013, 19, 155. [Google Scholar]
- Sultana, Z.; Maiti, K.; Aitken, J.; Morris, J.; Dedman, L.; Smith, R. Oxidative stress, placental aging-related pathologies and adverse pregnancy outcomes. Am. J. Reprod. Immunol. 2017, 77, 969–973. [Google Scholar] [CrossRef]
- Toescu, V.; Nuttall, S.L.; Martin, U.; Kendall, M.J.; Dunne, F. Oxidative stress and normal pregnancy. Clin. Endocrinol. 2002, 57, 609–613. [Google Scholar] [CrossRef] [PubMed]
- Dennery, P.A. Oxidative stress in development: Nature or nurture? Free Radic. Biol. Med. 2010, 49, 1147–1151. [Google Scholar] [CrossRef]
- Hernández-García, D.; Wood, C.D.; Castro-Obregón, S.; Covarrubias, L. Reactive oxygen species: A radical role in development? Free Radic. Biol. Med. 2010, 49, 130–143. [Google Scholar] [CrossRef] [PubMed]
- Mistry, H.D.; Williams, P.J. The importance of antioxidant micronutrients in pregnancy. Oxid. Med. Cell Longev. 2011, 2011, 841749. [Google Scholar] [CrossRef]
- Cho, H.Y.; Reddy, S.P.; Debiase, A.; Yamamoto, M.; Kleeberger, S.R. Gene expression profiling of NRF2-mediated protection against oxidative injury. Free Radic. Biol. Med. 2005, 38, 325–343. [Google Scholar] [CrossRef] [PubMed]
- Itoh, K.; Mochizuki, M.; Ishii, Y.; Ishii, T.; Shibata, T.; Kawamoto, Y.; Kelly, V.; Sekizawa, K.; Uchida, K.; Yamamoto, M. Transcription factor Nrf2 regulates inflammation by mediating the effect of 15-deoxy-Delta(12,14)-prostaglandin j(2). Mol. Cell Biol. 2004, 24, 36–45. [Google Scholar] [CrossRef]
- Cheng, X.; Chapple, S.J.; Patel, B.; Puszyk, W.; Sugden, D.; Yin, X.; Mayr, M.; Siow, R.C.; Mann, G.E. Gestational diabetes mellitus impairs Nrf2-mediated adaptive antioxidant defenses and redox signaling in fetal endothelial cells in utero. Diabetes 2013, 62, 4088–4097. [Google Scholar] [CrossRef] [PubMed]
- Lim, R.; Barker, G.; Lappas, M. The transcription factor Nrf2 is decreased after spontaneous term labour in human fetal membranes where it exerts anti-inflammatory properties. Placenta 2015, 36, 7–17. [Google Scholar] [CrossRef]
- Goto, T.; Takano, M.; Albergaria, A.; Briese, J.; Pomeranz, K.M.; Cloke, B.; Fusi, L.; Feroze-Zaidi, F.; Maywald, N.; Sajin, M.; et al. Mechanism and functional consequences of loss of FOXO1 expression in endometrioid endometrial cancer cells. Oncogene 2008, 27, 9–19. [Google Scholar] [CrossRef]
- Kajihara, T.; Brosens, J.J.; Ishihara, O. The role of FOXO1 in the decidual transformation of the endometrium and early pregnancy. Med. Mol. Morphol. 2013, 46, 61–68, Erratum in Med. Mol. Morphol. 2013, 46, 69. [Google Scholar] [CrossRef]
- Kovács, P.; Joó, J.G.; Tamás, V.; Molnár, Z.; Burik-Hajas, D.; Bódis, J.; Kornya, L. The role of apoptosis in the complex pathogenesis of the most common obstetrics and gynaecology diseases. Physiol. Int. 2020, 107, 106–119. [Google Scholar] [CrossRef]
- Pozharny, Y.; Lambertini, L.; Clunie, G.; Ferrara, L.; Lee, M.J. Epigenetics in women’s health care. Mt. Sinai J. Med. 2010, 77, 225–235. [Google Scholar] [CrossRef]
- Reik, W.; Dean, W.; Walter, J. Epigenetic reprogramming in mammalian development. Science 2001, 293, 1089–1093. [Google Scholar] [CrossRef]
- Barker, D.J. The developmental origins of adult disease. J. Am. Coll. Nutr. 2004, 23 (Suppl. 6), 588S–595S. [Google Scholar] [CrossRef]
- Myatt, L. Review: Reactive oxygen and nitrogen species and functional adaptation of the placenta. Placenta 2010, 31, S66–S69. [Google Scholar] [CrossRef] [PubMed]
- Camm, E.J.; Hansell, J.A.; Kane, A.D.; Herrera, E.A.; Lewis, C.; Wong, S.; Morrell, N.W.; Giussani, D.A. Partial contributions of developmental hypoxia and undernutrition to prenatal alterations in somatic growth and cardiovascular structure and function. Am. J. Obstet. Gynecol. 2010, 203, 495.e24–495.e34. [Google Scholar] [CrossRef] [PubMed]
- Ziech, D.; Franco, R.; Pappa, A.; Panayiotidis, M.I. Reactive oxygen species (ROS)--induced genetic and epigenetic alterations in human carcinogenesis. Mutat. Res. 2011, 711, 167–173. [Google Scholar] [CrossRef]
- Cerda, S.; Weitzman, S.A. Influence of oxygen radical injury on DNA methylation. Mutat. Res. 1997, 386, 141–152. [Google Scholar] [CrossRef]
- Singhal, A.; Lucas, A. Early origins of cardiovascular disease: Is there a unifying hypothesis? Lancet 2004, 363, 1642–1645. [Google Scholar] [CrossRef]
- Peuchant, E.; Brun, J.L.; Rigalleau, V.; Dubourg, L.; Thomas, M.J.; Daniel, J.Y.; Leng, J.J.; Gin, H. Oxidative and antioxidative status in pregnant women with either gestational or type 1 diabetes. Clin. Biochem. 2004, 37, 293–298. [Google Scholar] [CrossRef]
- Block, G.; Dietrich, M.; Norkus, E.P.; Morrow, J.D.; Hudes, M.; Caan, B.; Packer, L. Factors associated with oxidative stress in human populations. Am. J. Epidemiol. 2002, 156, 274–285. [Google Scholar] [CrossRef] [PubMed]
- Mohn, A.; Chiavaroli, V.; Cerruto, M.; Blasetti, A.; Giannini, C.; Bucciarelli, T.; Chiarelli, F. Increased oxidative stress in prepubertal children born small for gestational age. J. Clin. Endocrinol. Metab. 2007, 92, 1372–1378. [Google Scholar] [CrossRef] [PubMed]
- Chiavaroli, V.; Giannini, C.; D’Adamo, E.; de Giorgis, T.; Chiarelli, F.; Mohn, A. Insulin resistance and oxidative stress in children born small and large for gestational age. Pediatrics 2009, 124, 695–702. [Google Scholar] [CrossRef] [PubMed]
- Morton, J.S.; Rueda-Clausen, C.F.; Davidge, S.T. Mechanisms of endothelium-dependent vasodilation in male and female, young and aged offspring born growth restricted. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010, 298, R930–R938. [Google Scholar] [CrossRef]
- Simmons, R.A.; Suponitsky-Kroyter, I.; Selak, M.A. Progressive accumulation of mitochondrial DNA mutations and decline in mitochondrial function lead to beta-cell failure. J. Biol. Chem. 2005, 280, 28785–28791. [Google Scholar] [CrossRef] [PubMed]
- Koek, G.H.; Liedorp, P.R.; Bast, A. The role of oxidative stress in non-alcoholic steatohepatitis. Clin. Chim. Acta. 2011, 412, 1297–1305. [Google Scholar] [CrossRef]
- Thompson, L.P.; Al-Hasan, Y. Impact of oxidative stress in fetal programming. J. Pregnancy 2012, 2012, 582748. [Google Scholar] [CrossRef] [PubMed]
- Schoots, M.H.; Gordijn, S.J.; Scherjon, S.A.; van Goor, H.; Hillebrands, J.L. Oxidative stress in placental pathology. Placenta 2018, 69, 153–161. [Google Scholar] [CrossRef]
- Jauniaux, E.; Watson, A.L.; Hempstock, J.; Bao, Y.P.; Skepper, J.N.; Burton, G.J. Onset of maternal arterial blood flow and placental oxidative stress. A possible factor in human early pregnancy failure. Am. J. Pathol. 2000, 157, 2111–2122. [Google Scholar] [CrossRef]
- Burton, G.J.; Hempstock, J.; Jauniaux, E. Oxygen, early embryonic metabolism and free radical-mediated embryopathies. Reprod. Biomed. Online 2003, 6, 84–96. [Google Scholar] [CrossRef]
- Aris, A.; Benali, S.; Ouellet, A.; Moutquin, J.M.; Leblanc, S. Potential biomarkers of preeclampsia: Inverse correlation between hydrogen peroxide and nitric oxide early in maternal circulation and at term in placenta of women with preeclampsia. Placenta 2009, 30, 342–347. [Google Scholar] [CrossRef]
- Hung, T.H.; Skepper, J.N.; Burton, G.J. In vitro ischemia-reperfusion injury in term human placenta as a model for oxidative stress in pathological pregnancies. Am. J. Pathol. 2001, 159, 1031–1043. [Google Scholar] [CrossRef]
- Watson, A.L.; Skepper, J.N.; Jauniaux, E.; Burton, G.J. Changes in concentration, localization and activity of catalase within the human placenta during early gestation. Placenta 1998, 19, 27–34. [Google Scholar] [CrossRef]
- Watson, A.L.; Palmer, M.E.; Jauniaux, E.; Burton, G.J. Variations in expression of copper/zinc superoxide dismutase in villous trophoblast of the human placenta with gestational age. Placenta 1997, 18, 295–299. [Google Scholar] [CrossRef] [PubMed]
- Sugino, N.; Takiguchi, S.; Umekawa, T.; Heazell, A.; Caniggia, I. Oxidative stress and pregnancy outcome: A workshop report. Placenta 2007, 28 (Suppl. A), S48–S50. [Google Scholar] [CrossRef]
- Kimura, C.; Watanabe, K.; Iwasaki, A.; Mori, T.; Matsushita, H.; Shinohara, K.; Wakatsuki, A. The severity of hypoxic changes and oxidative DNA damage in the placenta of early-onset preeclamptic women and fetal growth restriction. J. Matern. Fetal Neonatal. Med. 2013, 26, 491–496. [Google Scholar] [CrossRef] [PubMed]
- Szentpéteri, I.; Rab, A.; Kornya, L.; Kovács, P.; Joó, J.G. Gene expression patterns of vascular endothelial growth factor (VEGF-A) in human placenta from pregnancies with intrauterine growth restriction. J. Matern. Fetal Neonatal. Med. 2013, 26, 984–989. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Itoh, H.; Kimura, S.; Sugihara, K.; Yaguchi, C.; Kobayashi, Y.; Hirai, K.; Takeuchi, K.; Sugimura, M.; Kanayama, N. Chorangiosis and placental oxygenation. Congenit Anom. 2009, 49, 71–76. [Google Scholar] [CrossRef]
- Ruano, C.S.M.; Miralles, F.; Méhats, C.; Vaiman, D. The impact of oxidative stress of environmental origin on the onset of placental diseases. Antioxidants 2022, 11, 106. [Google Scholar] [CrossRef]
- Sebastiani, G.; Navarro-Tapia, E.; Almeida-Toledano, L.; Serra-Delgado, M.; Paltrinieri, A.L.; García-Algar, Ó.; Andreu-Fernández, V. Effects of antioxidant intake on fetal development and maternal/neonatal health during pregnancy. Antioxidants 2022, 11, 648–684. [Google Scholar] [CrossRef] [PubMed]
- Tasdemir, E.; Chiara Maiuri, M.; Morselli, E.; Criollo, A.; D’Amelio, M.; Djavaheri-Mergny, M.; Cecconi, F.; Tavernarakis, N.; Kroemer, G. A dual role of p53 in the control of autophagy. Autophagy 2008, 4, 810–814. [Google Scholar] [CrossRef]
- Kemse, N.G.; Kale, A.A.; Joshi, S.R. A combined supplementation of omega-3 fatty acids and micronutrients (folic acid, vitamin B12) reduces oxidative stress markers in a rat model of pregnancy induced hypertension. PLoS ONE 2014, 9, e111902. [Google Scholar] [CrossRef]
- Dutta, S.; Kumar, S.; Hyett, J.; Salomon, C. Molecular Targets of aspirin and prevention of preeclampsia and their potential association with circulating extracellular vesicles during pregnancy. Int. J. Mol. Sci. 2019, 20, 4370. [Google Scholar] [CrossRef]
- Khera, A.; Vanderlelie, J.J.; Perkins, A.V. Selenium supplementation protects trophoblast cells from mitochondrial oxidative stress. Placenta 2013, 34, 594–598. [Google Scholar] [CrossRef]
- Johal, T.; Lees, C.C.; Everett, T.R.; Wilkinson, I.B. The nitric oxide pathway and possible therapeutic options in pre-eclampsia. Br. J. Clin. Pharmacol. 2014, 78, 244–257. [Google Scholar] [CrossRef] [PubMed]
- Foisy, M.A.; Marchant, I.; Lessard, L.; Caron, L.; Bujold, E. Aspirin for the prevention of preeclampsia. J. Perinat. Med. 2022, 50, 1271–1272. [Google Scholar] [CrossRef]
- Chen, B.; Longtine, M.S.; Nelson, D.M. Punicalagin, a polyphenol in pomegranate juice, downregulates p53 and attenuates hypoxia-induced apoptosis in cultured human placental syncytiotrophoblasts. Am. J. Physiol. Endocrinol. Metab. 2013, 305, E1274–E1280. [Google Scholar] [CrossRef]
- Wu, F.; Tian, F.J.; Lin, Y. Oxidative stress in placenta: Health and diseases. Biomed. Res. Int. 2015, 2015, 293271. [Google Scholar] [CrossRef] [PubMed]
- Nybo Andersen, A.M.; Wohlfahrt, J.; Christens, P.; Olsen, J.; Melbye, M. Maternal age and fetal loss: Population based register linkage study. BMJ 2000, 320, 1708–1712. [Google Scholar] [CrossRef] [PubMed]
- Zejnullahu, V.A.; Zejnullahu, V.A.; Kosumi, E. The role of oxidative stress in patients with recurrent pregnancy loss: A review. Reprod. Health 2021, 18, 207. [Google Scholar] [CrossRef] [PubMed]
- Fait, V.; Sela, S.; Ophir, E.; Kreutzer, H.; Shnaider, O.; Perri, A.; Khatib, N.; Dourleshter, G.; Tendler, R.; Bornstein, J. Peripheral polymorphonuclear leukocyte priming contributes to oxidative stress in early pregnancy. J. Soc. Gynecol. Investig. 2005, 2, 46–49. [Google Scholar] [CrossRef]
- Hernandez, I.; Fournier, T.; Chissey, A.; Therond, P.; Slama, A.; Beaudeux, J.L.; Zerrad-Saadi, A. NADPH oxidase is the major source of placental superoxide in early pregnancy: Association with MAPK pathway activation. Sci. Rep. 2019, 9, 13962. [Google Scholar] [CrossRef] [PubMed]
- Raijmakers, M.T.; Burton, G.J.; Jauniaux, E.; Seed, P.T.; Peters, W.H.; Steegers, E.A.; Poston, L. Placental NAD(P)H oxidase mediated superoxide generation in early pregnancy. Placenta 2006, 27, 158–163. [Google Scholar] [CrossRef] [PubMed]
- Bedard, K.; Krause, K.H. The NOX family of ROS-generating NADPH oxidases: Physiology and pathophysiology. Physiol. Rev. 2007, 87, 245–313. [Google Scholar] [CrossRef] [PubMed]
- Ghneim, H.K.; Alshebly, M.M. Biochemical Markers of Oxidative Stress in Saudi Women with Recurrent Miscarriage. J. Korean Med. Sci. 2016, 31, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Baban, R.S. Oxidative stress in recurrent pregnancy loss women. Saudi Med. J. 2010, 31, 759–763. [Google Scholar]
- Brown, M.A.; Magee, L.A.; Kenny, L.C.; Karumanchi, S.A.; McCarthy, F.P.; Saito, S.; Hall, D.R.; Warren, C.E.; Adoyi, G.; Ishaku, S. International Society for the Study of Hypertension in Pregnancy (ISSHP). Hypertensive Disorders of Pregnancy: ISSHP Classification, Diagnosis, and Management Recommendations for International Practice. Hypertension 2018, 72, 24–43. [Google Scholar] [CrossRef] [PubMed]
- Baradaran, A.; Nasri, H.; Rafieian-Kopaei, M. Oxidative stress and hypertension: Possibility of hypertension therapy with antioxidants. J. Res. Med. Sci. 2014, 19, 358–367. [Google Scholar] [PubMed]
- Harrison, D.G.; Gongora, M.C. Oxidative stress and hypertension. Med. Clin. N. Am. 2009, 93, 621–635. [Google Scholar] [CrossRef]
- Gathiram, P.; Moodley, J. Pre-eclampsia: Its pathogenesis and pathophysiolgy. Cardiovasc. J. Afr. 2016, 27, 71–78. [Google Scholar] [CrossRef]
- Maynard, S.E.; Min, J.Y.; Merchan, J.; Lim, K.H.; Li, J.; Mondal, S.; Libermann, T.A.; Morgan, J.P.; Sellke, F.W.; Stillman, I.E.; et al. Excess placental soluble fms-like tyrosine kinase 1 (sFlt1) may contribute to endothelial dysfunction, hypertension, and proteinuria in preeclampsia. J. Clin. Investig. 2003, 111, 649–658. [Google Scholar] [CrossRef] [PubMed]
- Roberts, J.M.; Hubel, C.A. The two stage model of preeclampsia: Variations on the theme. Placenta 2009, 30 (Suppl. A), 32–37. [Google Scholar] [CrossRef] [PubMed]
- Redman, C.W.; Sargent, I.L. Placental stress and pre-eclampsia: A revised view. Placenta 2009, 30 (Suppl. A), S38–S42. [Google Scholar] [CrossRef] [PubMed]
- Guerby, P.; Vidal, F.; Garoby-Salom, S.; Vayssiere, C.; Salvayre, R.; Parant, O.; Negre-Salvayre, A. Implication du stress oxydant dans la physiopathologie de la pré-éclampsie: Mise au point [Oxidative stress and preeclampsia: A review]. Gynecol. Obstet. Fertil. 2015, 43, 751–756. (In French) [Google Scholar] [CrossRef]
- Phoswa, W.N.; Khaliq, O.P. The Role of Oxidative Stress in Hypertensive Disorders of Pregnancy (Preeclampsia, Gestational Hypertension) and Metabolic Disorder of Pregnancy (Gestational Diabetes Mellitus). Oxid. Med. Cell Longev. 2021, 31, 5581570. [Google Scholar] [CrossRef] [PubMed]
- Matsubara, K.; Higaki, T.; Matsubara, Y.; Nawa, A. Nitric oxide and reactive oxygen species in the pathogenesis of preeclampsia. Int. J. Mol. Sci. 2015, 16, 4600–4614. [Google Scholar] [CrossRef]
- Sánchez-Aranguren, L.C.; Prada, C.E.; Riaño-Medina, C.E.; Lopez, M. Endothelial dysfunction and preeclampsia: Role of oxidative stress. Front. Physiol. 2014, 5, 372. [Google Scholar] [CrossRef] [PubMed]
- Gielis, J.F.; Lin, J.Y.; Wingler, K.; Van Schil, P.E.; Schmidt, H.H.; Moens, A.L. Pathogenetic role of eNOS uncoupling in cardiopulmonary disorders. Free Radic. Biol. Med. 2011, 50, 765–776. [Google Scholar] [CrossRef]
- Abe, E.; Matsubara, K.; Ochi, H.; Ito, M.; Oka, K.; Kameda, K. Elevated levels of adhesion molecules derived from leukocytes and endothelial cells in patients with pregnancy-induced hypertension. Hypertens. Pregnancy 2003, 22, 31–43. [Google Scholar] [CrossRef]
- Matsunaga, T.; Weihrauch, D.W.; Moniz, M.C.; Tessmer, J.; Warltier, D.C.; Chilian, W.M. Angiostatin inhibits coronary angiogenesis during impaired production of nitric oxide. Circulation 2002, 105, 2185–2191. [Google Scholar] [CrossRef]
- Nabah, Y.N.; Mateo, T.; Cerdá-Nicolás, M.; Alvarez, A.; Martinez, M.; Issekutz, A.C.; Sanz, M.J. L-NAME induces direct arteriolar leukocyte adhesion, which is mainly mediated by angiotensin-II. Microcirculation 2005, 12, 443–453. [Google Scholar] [CrossRef]
- Sanyal, M.; Nag, T.C.; Das, C. Localization of nitric oxide synthase in human trophoblast cells: Role of nitric oxide in trophoblast proliferation and differentiation. Am. J. Reprod. Immunol. 2000, 43, 70–77. [Google Scholar] [CrossRef]
- Buttery, L.D.; McCarthy, A.; Springall, D.R.; Sullivan, M.H.; Elder, M.G.; Michel, T.; Polak, J.M. Endothelial nitric oxide synthase in the human placenta: Regional distribution and proposed regulatory role at the feto-maternal interface. Placenta 1994, 15, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Tamás, P.; Bódis, J.; Sulyok, E.; Kovács, L.G.; Hantosi, E.; Molnár, G.; Martens-Lobenhoffer, J.; Bode-Böger, S.M. L-arginine metabolism in early-onset and late-onset pre-eclamptic pregnancies. Scan J. Clin. Lab. Investig. 2013, 73, 436–443. [Google Scholar] [CrossRef]
- Kurlak, L.O.; Green, A.; Loughna, P.; Broughton Pipkin, F. Oxidative stress markers in hypertensive states of pregnancy: Preterm and term disease. Front. Physiol. 2014, 5, 310. [Google Scholar] [CrossRef]
- Phoswa, W.N. Dopamine in the pathophysiology of preeclampsia and gestational hypertension: Monoamine oxidase (MAO) and catechol-O-methyl transferase (COMT) as Possible Mechanisms. Oxid. Med. Cell Longev. 2019, 28, 3546294. [Google Scholar] [CrossRef]
- Schei, J.; Fuskevåg, O.M.; Stefansson, V.T.N.; Solbu, M.D.; Jenssen, T.G.; Eriksen, B.O.; Melsom, T. Urinary markers of oxidative stress are associated with albuminuria but not GFR decline. Kidney Int. Rep. 2017, 3, 573–582. [Google Scholar] [CrossRef]
- Tan, T.Y.; Yeo, G.S. Intrauterine growth restriction. Curr. Opin. Obstet. Gynecol. 2005, 17, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Bligard, K.H.; Xu, X.; Raghuraman, N.; Dicke, J.M.; Odibo, A.O.; Frolova, A.I. Clinical significance of absent or reversed end diastolic velocity waveforms in umbilical artery. Lancet 1994, 344, 1664–1668. [Google Scholar]
- Krishna, U.; Bhalerao, S. Placental insufficiency and fetal growth restriction. J. Obstet. Gynaecol. India 2011, 61, 505–511. [Google Scholar] [CrossRef]
- Mandò, C.; De Palma, C.; Stampalija, T.; Anelli, G.M.; Figus, M.; Novielli, C.; Parisi, F.; Clementi, E.; Ferrazzi, E.; Cetin, I. Placental mitochondrial content and function in intrauterine growth restriction and preeclampsia. Am. J. Physiol. Endocrinol. Metab. 2014, 306, E404–E413. [Google Scholar] [CrossRef]
- Huppertz, B. Placental origins of preeclampsia: Challenging the current hypothesis. Hypertension 2008, 51, 970–975. [Google Scholar] [CrossRef]
- Joo, E.H.; Kim, Y.R.; Kim, N.; Jung, J.E.; Han, S.H.; Cho, H.Y. Effect of Endogenic and Exogenic Oxidative Stress Triggers on Adverse Pregnancy Outcomes: Preeclampsia, Fetal Growth Restriction, Gestational Diabetes Mellitus and Preterm Birth. Int. J. Mol. Sci. 2021, 22, 10122. [Google Scholar] [CrossRef]
- Biri, A.; Bozkurt, N.; Turp, A.; Kavutcu, M.; Himmetoglu, O.; Durak, I. Role of oxidative stress in intrauterine growth restriction. Gynecol. Obstet. Investig. 2007, 64, 187–192. [Google Scholar] [CrossRef]
- Davy, P.; Nagata, M.; Bullard, P.; Fogelson, N.S.; Allsopp, R. Fetal growth restriction is associated with accelerated telomere shortening and increased expression of cell senescence markers in the placenta. Placenta 2009, 30, 539–542. [Google Scholar] [CrossRef]
- Biron-Shental, T.; Sukenik-Halevy, R.; Sharon, Y.; Laish, I.; Fejgin, M.D.; Amiel, A. Telomere shortening in intra uterine growth restriction placentas. Early Hum. Dev. 2014, 90, 465–469. [Google Scholar] [CrossRef]
- Kim, S.Y.; Lee, S.P.; Lee, J.S.; Yoon, S.J.; Jun, G.; Hwang, Y.J. Telomerase and apoptosis in the placental trophoblasts of growth discordant twins. Yonsei Med. J. 2006, 47, 698–705. [Google Scholar] [CrossRef] [PubMed]
- Kudo, T.; Izutsu, T.; Sato, T. Telomerase activity and apoptosis as indicators of aging in placenta with and without intrauterine growth retardation. Placenta 2000, 21, 493–500. [Google Scholar] [CrossRef] [PubMed]
- Börzsönyi, B.; Demendi, C.; Rigó, J., Jr.; Szentpéteri, I.; Rab, A.; Joó, J.G. The regulation of apoptosis in intrauterine growth restriction: A study of Bcl-2 and Bax gene expression in human placenta. J. Matern. Fetal Neonatal. Med. 2013, 26, 347–350. [Google Scholar] [CrossRef] [PubMed]
- Yudkin, P.L.; Wood, L.; Redman, C.W. Risk of unexplained stillbirth at different gestational ages. Lancet 1987, 1, 1192–1194. [Google Scholar] [CrossRef]
- Ferrari, F.; Facchinetti, F.; Saade, G.; Menon, R. Placental telomere shortening in stillbirth: A sign of premature senescence? J. Matern. Fetal Neonatal. Med. 2016, 29, 1283–1288. [Google Scholar] [CrossRef]
- Levine, R.J.; Maynard, S.E.; Qian, C.; Lim, K.H.; England, L.J.; Yu, K.F.; Schisterman, E.F.; Thadhani, R.; Sachs, B.P.; Epstein, F.H.; et al. Circulating angiogenic factors and the risk of preeclampsia. N. Engl. J. Med. 2004, 350, 672–683. [Google Scholar] [CrossRef] [PubMed]
- Scaife, P.J.; Simpson, A.; Kurlak, L.O.; Briggs, L.V.; Gardner, D.S.; Broughton Pipkin, F.; Jones, C.J.P.; Mistry, H.D. Increased Placental Cell Senescence and Oxidative Stress in Women with Pre-Eclampsia and Normotensive Post-Term Pregnancies. Int. J. Mol. Sci. 2021, 7, 7295. [Google Scholar] [CrossRef]
- Gandhi, P.; Farrell, T. Gestational diabetes mellitus (GDM) screening in morbidly obese pregnant women. Eur. J. Obstet. Gynecol. Reprod. Biol. 2011, 159, 329–332. [Google Scholar] [CrossRef] [PubMed]
- Alberti, K.G.; Zimmet, P.Z. Definition, diagnosis and classification of diabetes mellitus and its complications. Part 1: Diagnosis and classification of diabetes mellitus provisional report of a WHO consultation. Diabet. Med. 1998, 15, 539–553. [Google Scholar] [CrossRef]
- Makgoba, M.; Savvidou, M.D.; Steer, P.J. An analysis of the interrelationship between maternal age, body mass index and racial origin in the development of gestational diabetes mellitus. BJOG 2012, 119, 276–282. [Google Scholar] [CrossRef]
- Carolan, M.; Davey, M.A.; Biro, M.A.; Kealy, M. Maternal age, ethnicity and gestational diabetes mellitus. Midwifery 2012, 28, 778–783. [Google Scholar] [CrossRef]
- Lindsay, C.A.; Huston, L.; Amini, S.B.; Catalano, P.M. Longitudinal changes in the relationship between body mass index and percent body fat in pregnancy. Obstet. Gynecol. 1997, 89, 377–382. [Google Scholar] [CrossRef]
- Buchanan, T.A.; Xiang, A.H. Gestational diabetes mellitus. J. Clin. Investig. 2005, 115, 485–491. [Google Scholar] [CrossRef] [PubMed]
- Oztekin, O. New insights into the pathophysiology of gestational diabetes mellitus: Possible role of human leukocyte antigen-G. Med. Hypotheses 2007, 69, 526–530. [Google Scholar] [CrossRef]
- Christian, L.M.; Porter, K. Longitudinal changes in serum proinflammatory markers across pregnancy and postpartum: Effects of maternal body mass index. Cytokine 2014, 70, 134–140. [Google Scholar] [CrossRef]
- Solis Paredes, J.M.; Perichart Perera, O.; Montoya Estrada, A.; Reyes Muñoz, E.; Espino YSosa, S.; Ortega Castillo, V.; Medina Bastidas, D.; Tolentino Dolores, M.; Sanchez Martinez, M.; Nava Salazar, S.; et al. Gestational Weight Gain Influences the Adipokine-Oxidative Stress Association during Pregnancy. Obes. Facts. 2021, 14, 604–612. [Google Scholar] [CrossRef]
- Zhu, C.; Yang, H.; Geng, Q.; Ma, Q.; Long, Y.; Zhou, C.; Chen, M. Association of oxidative stress biomarkers with gestational diabetes mellitus in pregnant women: A case-control study. PLoS ONE 2015, 10, e0126490. [Google Scholar] [CrossRef]
- Lappas, M.; Hiden, U.; Desoye, G.; Froehlich, J.; Hauguel-de Mouzon, S.; Jawerbaum, A. The role of oxidative stress in the pathophysiology of gestational diabetes mellitus. Antioxid. Redox Signal. 2011, 15, 3061–3100. [Google Scholar] [CrossRef] [PubMed]
- Coughlan, M.T.; Vervaart, P.P.; Permezel, M.; Georgiou, H.M.; Rice, G.E. Altered placental oxidative stress status in gestational diabetes mellitus. Placenta 2004, 25, 78–84. [Google Scholar] [CrossRef]
- Huang, T.T.; Sun, W.J.; Liu, H.Y.; Ma, H.L.; Cui, B.X. p66Shc-mediated oxidative stress is involved in gestational diabetes mellitus. World J. Diabetes 2021, 12, 1894–1907. [Google Scholar] [CrossRef] [PubMed]
- Saiyin, T.; Engineer, A.; Greco, E.R.; Kim, M.Y.; Lu, X.; Jones, D.L.; Feng, Q. Maternal voluntary exercise mitigates oxidative stress and incidence of congenital heart defects in pre-gestational diabetes. J. Cell Mol. Med. 2019, 23, 5553–5565. [Google Scholar] [CrossRef]
- Grissa, O.; Atègbo, J.M.; Yessoufou, A.; Tabka, Z.; Miled, A.; Jerbi, M.; Dramane, K.L.; Moutairou, K.; Prost, J.; Hichami, A.; et al. Antioxidant status and circulating lipids are altered in human gestational diabetes and macrosomia. Transl. Res. 2007, 150, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Saker, M.; Soulimane Mokhtari, N.; Merzouk, S.A.; Merzouk, H.; Belarbi, B.; Narce, M. Oxidant and antioxidant status in mothers and their newborns according to birthweight. Eur. J. Obstet. Gynecol. Reprod. Biol. 2008, 141, 95–99. [Google Scholar] [CrossRef] [PubMed]
- Domellöf, M.; Thorsdottir, I.; Thorstensen, K. Health effects of different dietary iron intakes: A systematic literature review for the 5th Nordic Nutrition Recommendations. Food Nutr. Res. 2013, 12, 57. [Google Scholar] [CrossRef]
- Bowers, K.; Yeung, E.; Williams, M.A.; Qi, L.; Tobias, D.K.; Hu, F.B.; Zhang, C. A prospective study of prepregnancy dietary iron intake and risk for gestational diabetes mellitus. Diabetes Care 2011, 34, 1557–1563. [Google Scholar] [CrossRef]
- Zhuang, T.; Han, H.; Yang, Z. Iron, oxidative stress and gestational diabetes. Nutrients 2014, 25, 3968–3980. [Google Scholar] [CrossRef]
- Pustovrh, M.C.; Jawerbaum, A.; Capobianco, E.; White, V.; Martínez, N.; López-Costa, J.J.; González, E. Oxidative stress promotes the increase of matrix metalloproteinases-2 and -9 activities in the feto-placental unit of diabetic rats. Free Radic. Res. 2005, 39, 1285–1293. [Google Scholar] [CrossRef]
- Delnord, M.; Zeitlin, J. Epidemiology of late preterm and early term births–An international perspective. Semin. Fetal Neonatal. Med. 2019, 24, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Frey, H.A.; Klebanoff, M.A. The epidemiology, etiology, and costs of preterm birth. Semin. Fetal Neonatal. Med. 2016, 21, 68–73. [Google Scholar] [CrossRef] [PubMed]
- Polettini, J.; da Silva, M.G. Telomere-Related Disorders in fetal membranes associated with birth and adverse pregnancy outcomes. Front. Physiol. 2020, 11, 561771. [Google Scholar] [CrossRef]
- Manna, S.; McCarthy, C.; McCarthy, F.P. Placental Aging in adverse pregnancy outcomes: Telomere shortening, cell senescence, and mitochondrial dysfunction. Oxid. Med. Cell Longev. 2019, 2019, 3095383. [Google Scholar] [CrossRef] [PubMed]
- Menon, R. Oxidative stress damage as a detrimental factor in preterm birth pathology. Front. Immunol. 2014, 5, 567. [Google Scholar] [CrossRef] [PubMed]
- Polettini, J.; Dutta, E.H.; Behnia, F.; Saade, G.R.; Torloni, M.R.; Menon, R. Aging of intrauterine tissues in spontaneous preterm birth and preterm premature rupture of the membranes: A systematic review of the literature. Placenta 2015, 36, 969–973. [Google Scholar] [CrossRef]
- Dhobale, M.; Mehendale, S.; Pisal, H.; Nimbargi, V.; Joshi, S. Reduced maternal and cord nerve growth factor levels in preterm deliveries. Int. J. Dev. Neurosci. 2012, 30, 99–103. [Google Scholar] [CrossRef]
- Mustafa, M.D.; Pathak, R.; Ahmed, T.; Ahmed, R.S.; Tripathi, A.K.; Guleria, K.; Banerjee, B.D. Association of glutathione S-transferase M1 and T1 gene polymorphisms and oxidative stress markers in preterm labor. Clin. Biochem. 2010, 43, 1124–1128. [Google Scholar] [CrossRef]
- Cipierre, C.; Haÿs, S.; Maucort-Boulch, D.; Steghens, J.P.; Picaud, J.C. Malondialdehyde adduct to hemoglobin: A new marker of oxidative stress suitable for full-term and preterm neonates. Oxid. Med. Cell Longev. 2013, 2013, 694014. [Google Scholar] [CrossRef]
- Niki, E. Antioxidants: Basic principles, emerging concepts, and problems. Biomed. J. 2014, 37, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Eick, S.M.; Ferguson, K.K.; Milne, G.L.; Rios-McConnell, R.; Vélez-Vega, C.; Rosario, Z.; Alshawabkeh, A.; Cordero, J.F.; Meeker, J.D. Repeated measures of urinary oxidative stress biomarkers during pregnancy and preterm birth. Am. J. Obstet. Gynecol. 2015, 212, e1–e8. [Google Scholar] [CrossRef]
- Wall, P.D.; Pressman, E.K.; Woods, J.R., Jr. Preterm premature rupture of the membranes and antioxidants: The free radical connection. J. Perinat. Med. 2002, 30, 447–457. [Google Scholar] [CrossRef]
- Lázár, R.; Orvos, H.; Szőllősi, R.; Varga, I.S. The quality of the antioxidant defence system in term and preterm twin neonates. Redox Rep. 2015, 20, 103–108. [Google Scholar] [CrossRef]
- Weber, D.; Stuetz, W.; Bernhard, W.; Franz, A.; Raith, M.; Grune, T.; Breusing, N. Oxidative stress markers and micronutrients in maternal and cord blood in relation to neonatal outcome. Eur. J. Clin. Nutr. 2014, 68, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Abiaka, C.; Machado, L. Nitric oxide and antioxidant enzymes in venous and cord blood of late preterm and term omani mothers. Sultan Qaboos Univ. Med. J. 2012, 12, 300–305. [Google Scholar] [CrossRef]
- Isbilen, E.; Kulaksizoglu, S.; Kirmizioglu, M.; Karuserci Komurcu, O.; Tabur, S. Role of prolidase activity and oxidative stress biomarkers in unexplained infertility. Int. J. Gynaecol. Obstet. 2022, 156, 430–435. [Google Scholar] [CrossRef] [PubMed]
- Jeremy, J.Y.; Shukla, N. Ceruloplasmin dysfunction: A key factor in the pathophysiology of atrial fibrillation? J. Intern. Med. 2014, 275, 191–194. [Google Scholar] [CrossRef] [PubMed]
- Herway, C.; Kanninen, T.; Witkin, S.S.; Saade, G.; Fortunato, S.J.; Menon, R. Ethnic disparity in amniotic fluid levels of hyaluronan, histone H2B and superoxide dismutase in spontaneous preterm birth. J. Perinat. Med. 2013, 41, 277–282. [Google Scholar] [CrossRef]
- Song, J.; Dong, X.; Chen, Y.; Chen, G.; Liang, H.; Nakamura, H.; Yodoi, J.; Bai, J. The expression of thioredoxin-1 in premature delivery placenta. Redox Rep. 2012, 17, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Minghetti, L.; Suppiej, A.; Greco, A.; Franzoi, M.; Pascoli, I.; Zanardo, V. Oxidative stress in twin neonates is influenced by birth weight and weight discordance. Clin. Biochem. 2011, 44, 654–658. [Google Scholar] [CrossRef]
- Rogers, S.; Witz, G.; Anwar, M.; Hiatt, M.; Hegyi, T. Antioxidant capacity and oxygen radical diseases in the preterm newborn. Arch. Pediatr. Adolesc. Med. 2000, 154, 544–548. [Google Scholar] [CrossRef] [PubMed]
- Buonocore, G.; Perrone, S.; Longini, M.; Vezzosi, P.; Marzocchi, B.; Paffetti, P.; Bracci, R. Oxidative stress in preterm neonates at birth and on the seventh day of life. Pediatr. Res. 2002, 52, 46–49. [Google Scholar] [CrossRef]
- Hracsko, Z.; Orvos, H.; Novak, Z.; Pal, A.; Varga, I.S. Evaluation of oxidative stress markers in neonates with intra-uterine growth retardation. Redox Rep. 2008, 13, 11–16. [Google Scholar] [CrossRef]
- Simon-Szabo, Z.; Fogarasi, E.; Nemes-Nagy, E.; Denes, L.; Croitoru, M.; Szabo, B. Oxidative stress and peripartum outcomes (Review). Exp. Ther. Med. 2021, 22, 771. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Aggarwal, B.B. Activation of transcription factor NF-kappa B is suppressed by curcumin (diferuloylmethane). J. Biol. Chem. 1995, 270, 24995–25000. [Google Scholar] [CrossRef]
- Walsh, S.W. Preeclampsia: An imbalance in placental prostacyclin and thromboxane production. Am. J. Obstet. Gynecol. 1985, 152, 335–340. [Google Scholar] [CrossRef]
- Shi, D.D.; Guo, J.J.; Zhou, L.; Wang, N. Epigallocatechin gallate enhances treatment efficacy of oral nifedipine against pregnancy-induced severe pre-eclampsia: A double-blind, randomized and placebo-controlled clinical study. J. Clin. Pharm. Ther. 2018, 43, 21–25. [Google Scholar] [CrossRef]
- Ding, J.; Kang, Y.; Fan, Y.; Chen, Q. Efficacy of resveratrol to supplement oral nifedipine treatment in pregnancy-induced preeclampsia. Endocr. Connect. 2017, 6, 595–600. [Google Scholar] [CrossRef]
- Ghomian, N.; Hafizi, L.; Takhti, Z. The role of vitamin C in prevention of preterm premature rupture of membranes. Iran. Red. Crescent Med. J. 2013, 15, 113–116. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Gaikwad, H.S.; Nath, B.; Batra, A. Can vitamin C and interleukin 6 levels predict preterm premature rupture of membranes: Evaluating possibilities in North Indian population. Obstet. Gynecol. Sci. 2020, 63, 432–439. [Google Scholar] [CrossRef]
- Sharma, R.; Mehta, S. Ascorbic Acid concentration and preterm premature rupture of membranes. J. Obstet. Gynaecol. India. 2014, 64, 417–420. [Google Scholar] [CrossRef] [PubMed]
- Hauth, J.C.; Clifton, R.G.; Roberts, J.M.; Spong, C.Y.; Myatt, L.; Leveno, K.J.; Pearson, G.D.; Varner, M.W.; Thorp, J.M., Jr.; Mercer, B.M.; et al. Vitamin C and E supplementation to prevent spontaneous preterm birth: A randomized controlled trial. Obstet. Gynecol. 2010, 116, 653–658. [Google Scholar] [CrossRef] [PubMed]
- Koenig, M.D.; McFarlin, B.L.; Steffen, A.D.; Tussing-Humphreys, L.; Giurgescu, C.; Engeland, C.G.; Kominiarek, M.A.; Ciezczak-Karpiel, C.; O’Brien, W.D., Jr.; White-Traut, R. Decreased nutrient intake Is associated with premature cervical remodeling. J. Obstet. Gynecol. Neonatal. Nurs. 2017, 46, 123–134. [Google Scholar] [CrossRef] [PubMed]
- Naem, N.E.; El-Sayed, N.M.; Nossier, S.A.; Abu Zeid, A.A. Zinc status and dietary intake of pregnant women, Alexandria, Egypt. J. Egypt. Public. Health Assoc. 2014, 89, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Cannavò, L.; Perrone, S.; Viola, V.; Marseglia, L.; Di Rosa, G.; Gitto, E. Oxidative stress and respiratory diseases in preterm newborns. Int. J. Mol. Sci. 2021, 22, 12504. [Google Scholar] [CrossRef]


{kind=link}
| Zn-Cu SOD ⇑ → promotion of the development of follicles [9,10] |
| Synthesis of ovulatory steroids → P450 ⇑–ROS ⇑–blood supply–ovulation [1,9,10] |
| ROS ⇑ → induction of apoptosis in nondominant follicles [11] |
| FSH ⇑ → E2⇑ → CAT and GSH → protection of dominant follicle against apoptosis [10,11] |
| ROS ⇑ → NF-kappaB ⇑ → PGF2alfa → luteolysis [12] |
| Sperm cell–oocyte fusion → ROS ⇑ → maintenance of normal corpus luteum function [12] |
| ROS ⇑ → antioxidant effect ⇑ → progesterone production ⇑ [11] |
| GPx ⇑ → regulation of apoptosis in the placenta [4] |
| CAT, SOD, GPx in fetal tissues ⇑ defense of fetoplacental tissue against oxidative stress [4,9] |
| Condition | Quantitative Markers of Oxidative Stress |
|---|---|
| pre-eclampsia + IUGR | MDA, GSH, CAT, FT leptin, IMA, sRAGE [20,21,22,23,24,25] |
| gestational diabetes | MDA, TAC, GSH, CAT, NO [19,27,28,29] |
| premature delivery | 8-OHdG, GPx, CAT, NO, TAC, TOS, OSI [30,31,32,33,36,37] |
| First Trimester | Second Trimester | Third Trimester |
|---|---|---|
| Enhanced placental oxygen supply increases OS risk [49] | Rapid increase in oxygen delivery increases OS risk [16] | Strong OS effect with risk for lipid, protein and DNA damage [34] |
| Trophoblast invasion inducing development of spiral arteries [12] | Altered perfusion of uterine tissue [20,36] | Fetal DNA damage, impaired fetal development [35] |
| Increased placental arterial resistance reduces uteroplacental flow [12] | Progressive oxidant/antioxidant imbalance [39] | Enhanced placental apoptosis, placental failure [36,37] |
| Ischemia [23] | Reduced production of antioxidants [36] | |
| MISSED ABORTION, IUGR | IUGR | PREMATURE DELIVERY, IUGR, FETAL DEATH IN UTERO |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Joó, J.G.; Sulyok, E.; Bódis, J.; Kornya, L. Disrupted Balance of the Oxidant–Antioxidant System in the Pathophysiology of Female Reproduction: Oxidative Stress and Adverse Pregnancy Outcomes. Curr. Issues Mol. Biol. 2023, 45, 8091-8111. https://doi.org/10.3390/cimb45100511
Joó JG, Sulyok E, Bódis J, Kornya L. Disrupted Balance of the Oxidant–Antioxidant System in the Pathophysiology of Female Reproduction: Oxidative Stress and Adverse Pregnancy Outcomes. Current Issues in Molecular Biology. 2023; 45(10):8091-8111. https://doi.org/10.3390/cimb45100511
Chicago/Turabian StyleJoó, József Gábor, Endre Sulyok, József Bódis, and László Kornya. 2023. "Disrupted Balance of the Oxidant–Antioxidant System in the Pathophysiology of Female Reproduction: Oxidative Stress and Adverse Pregnancy Outcomes" Current Issues in Molecular Biology 45, no. 10: 8091-8111. https://doi.org/10.3390/cimb45100511