
Quercetin Alleviates Lipopolysaccharide-Induced Cell Damage and Inflammation via Regulation of the TLR4/NF-κB Pathway in Bovine Intestinal Epithelial Cells
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bovine Intestinal Epithelial Cell Culture
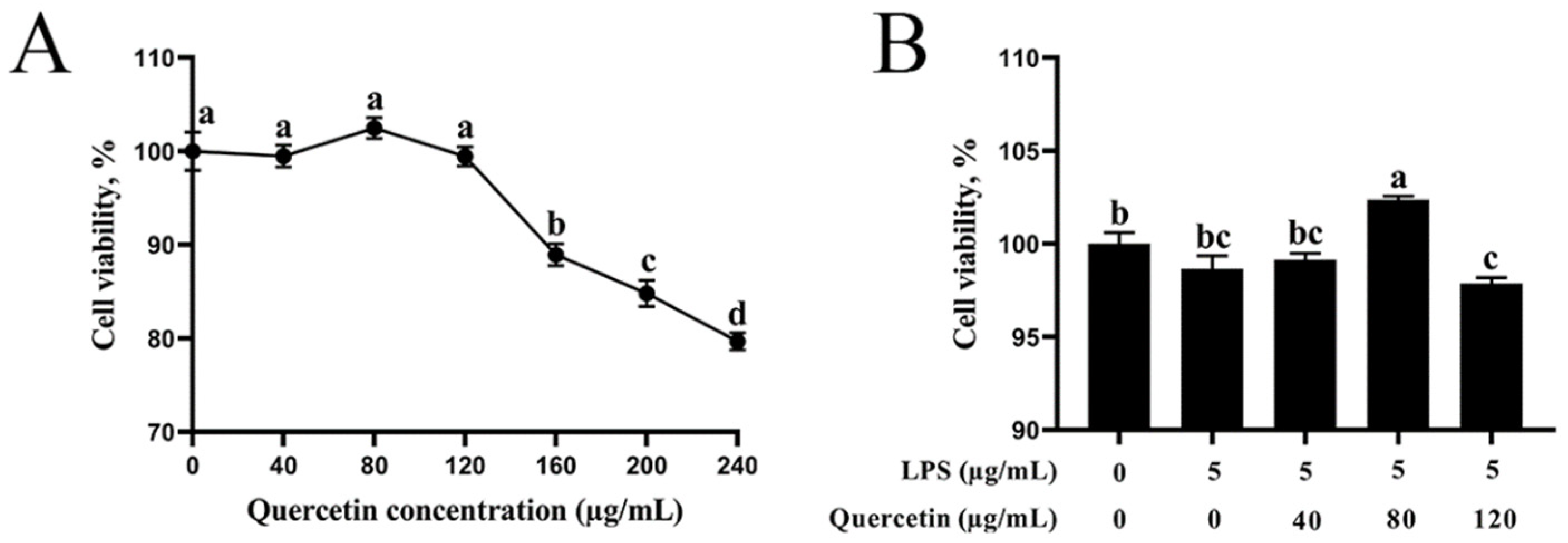
2.2. Effect of Quercetin and LPS on BIECs’ Viability
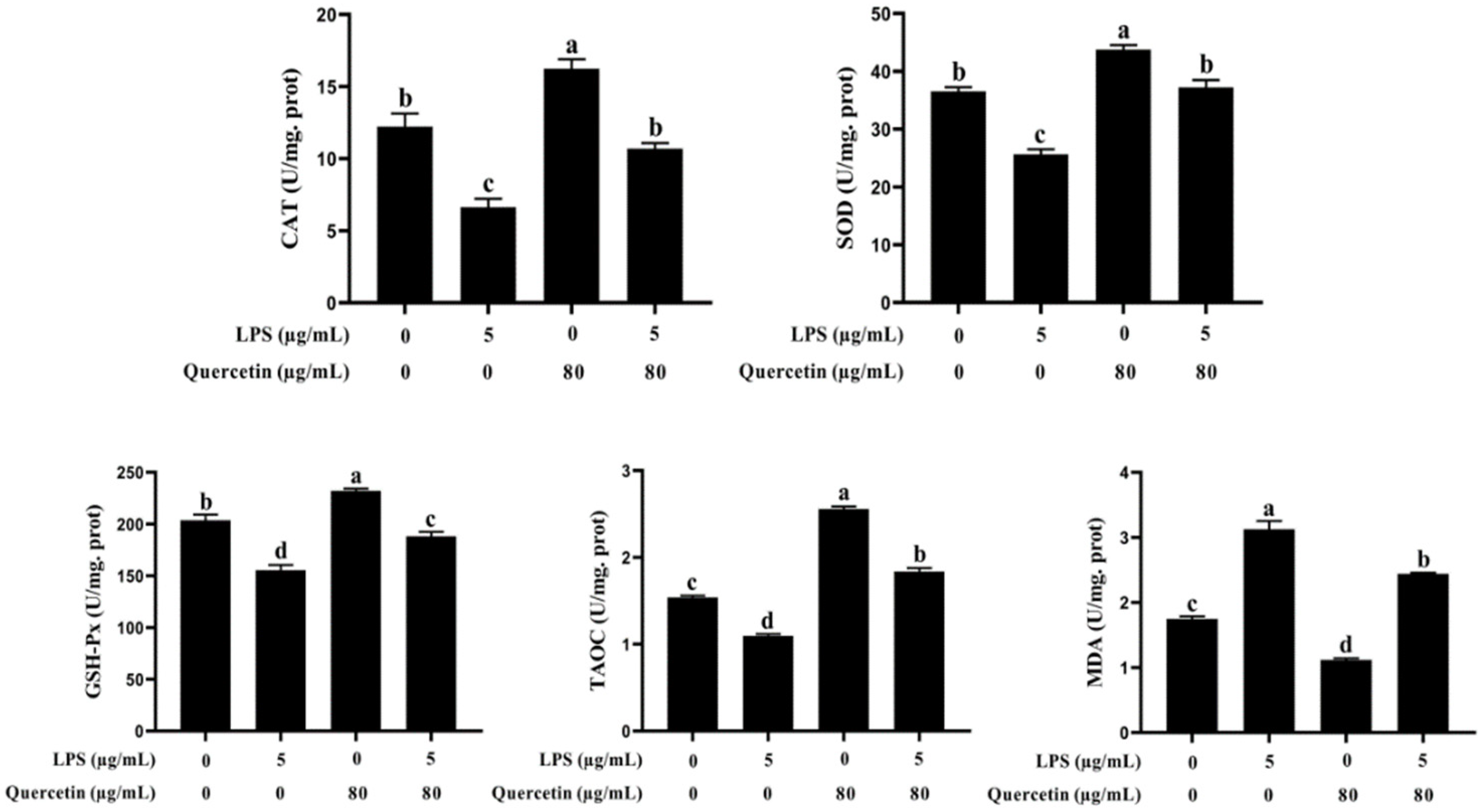
2.3. Antioxidant Analysis
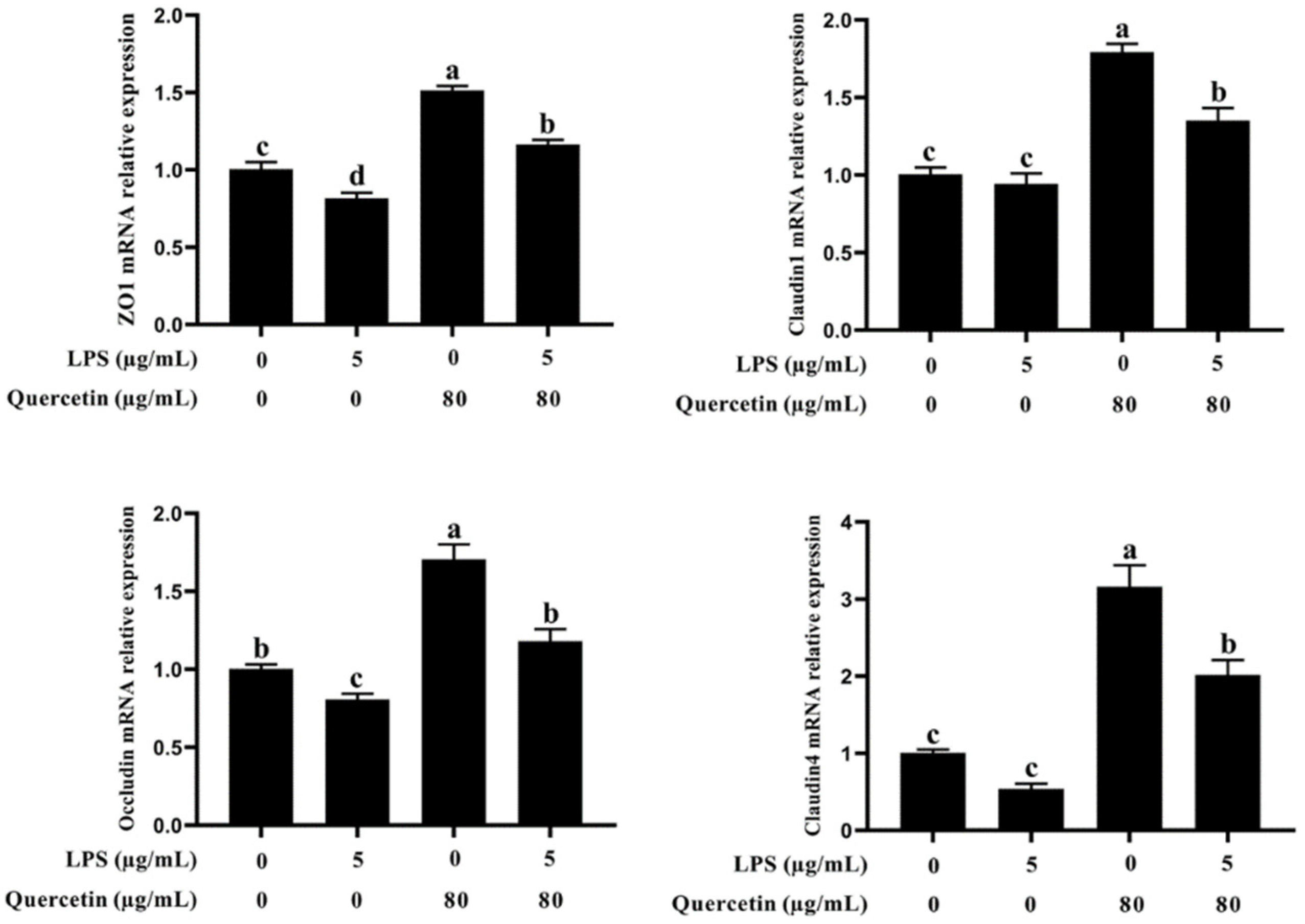
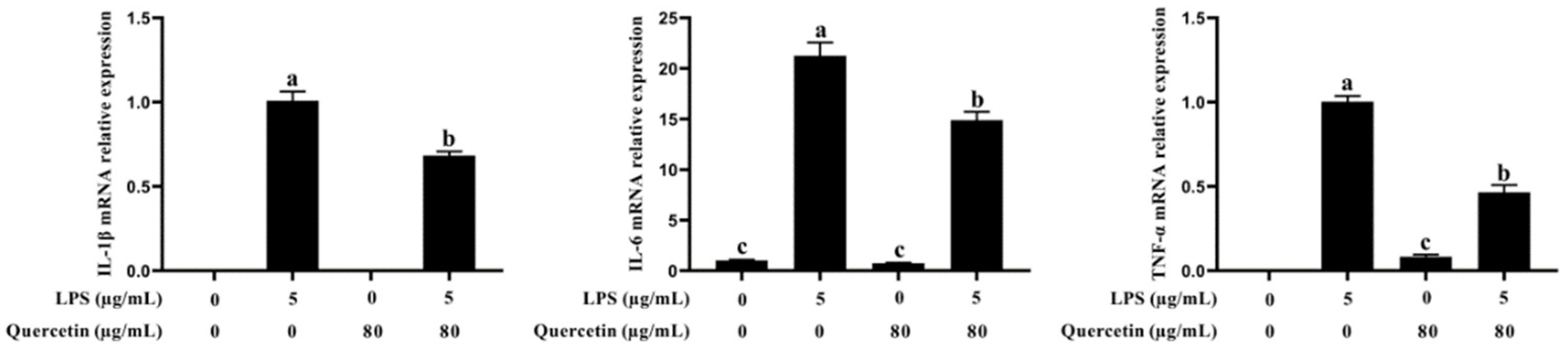
2.4. RNA Extraction, Reverse Transcription, and Real-Time PCR
2.5. Immunofluorescence Analysis
2.6. Western Blotting Analysis
2.7. Statistical Analysis
3. Results
3.1. Impact of Quercetin on the Viability of BIECs
3.2. Effect of Quercetin on Oxidative Features of BIECs Influenced by LPS
3.3. Effect of Quercetin on the Barrier Role of BIECs Caused by LPS
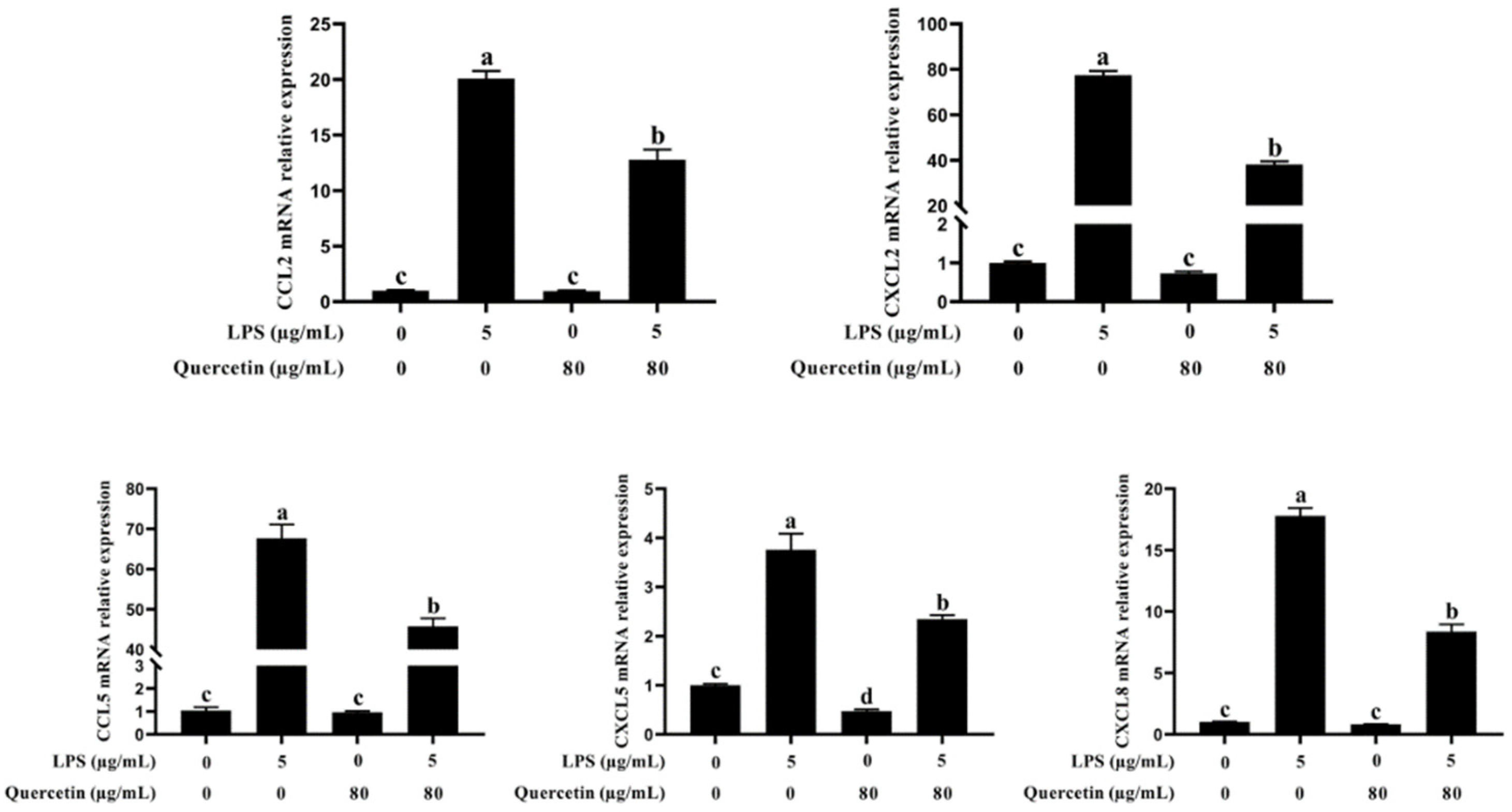
3.4. Effect of Quercetin on Pro-Inflammatory Factors of BIECs Caused by LPS
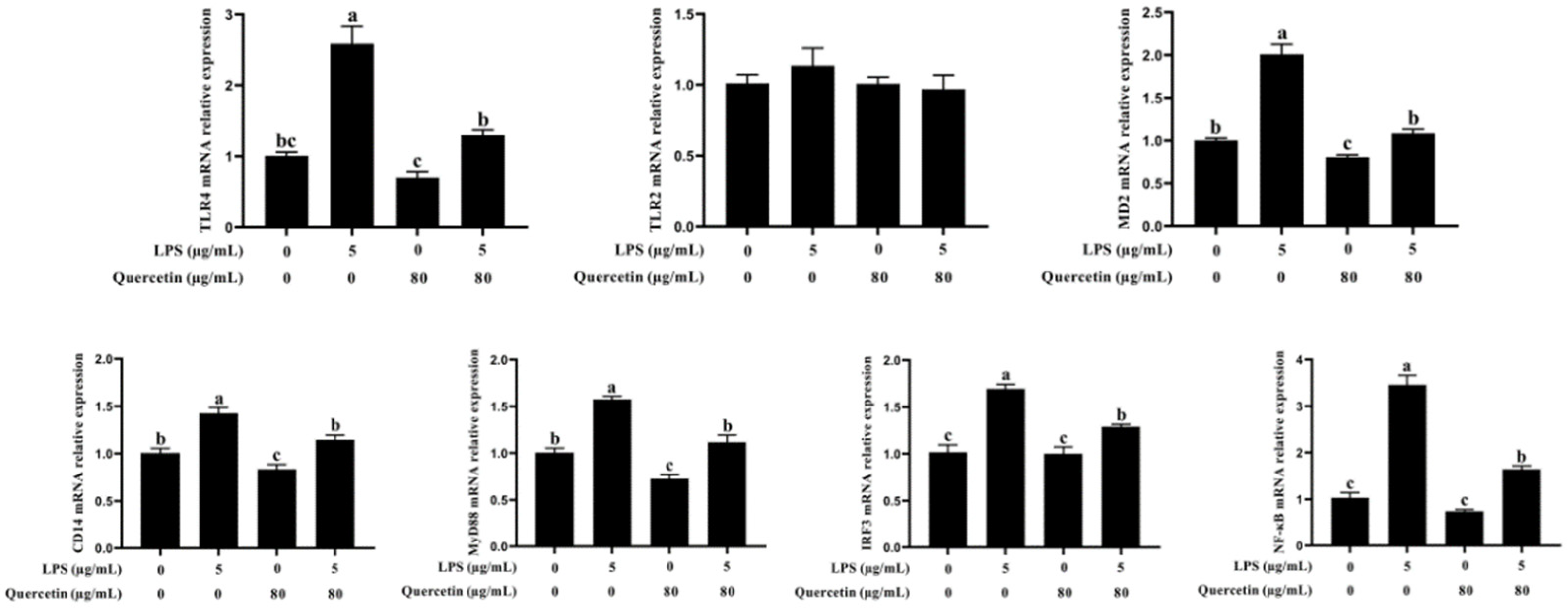
3.5. Effect of Quercetin on the Immune Reaction of BIECs Caused by LPS
3.6. Effect of Quercetin on the TLR4 Signalling Pathway of BIECs Caused by LPS
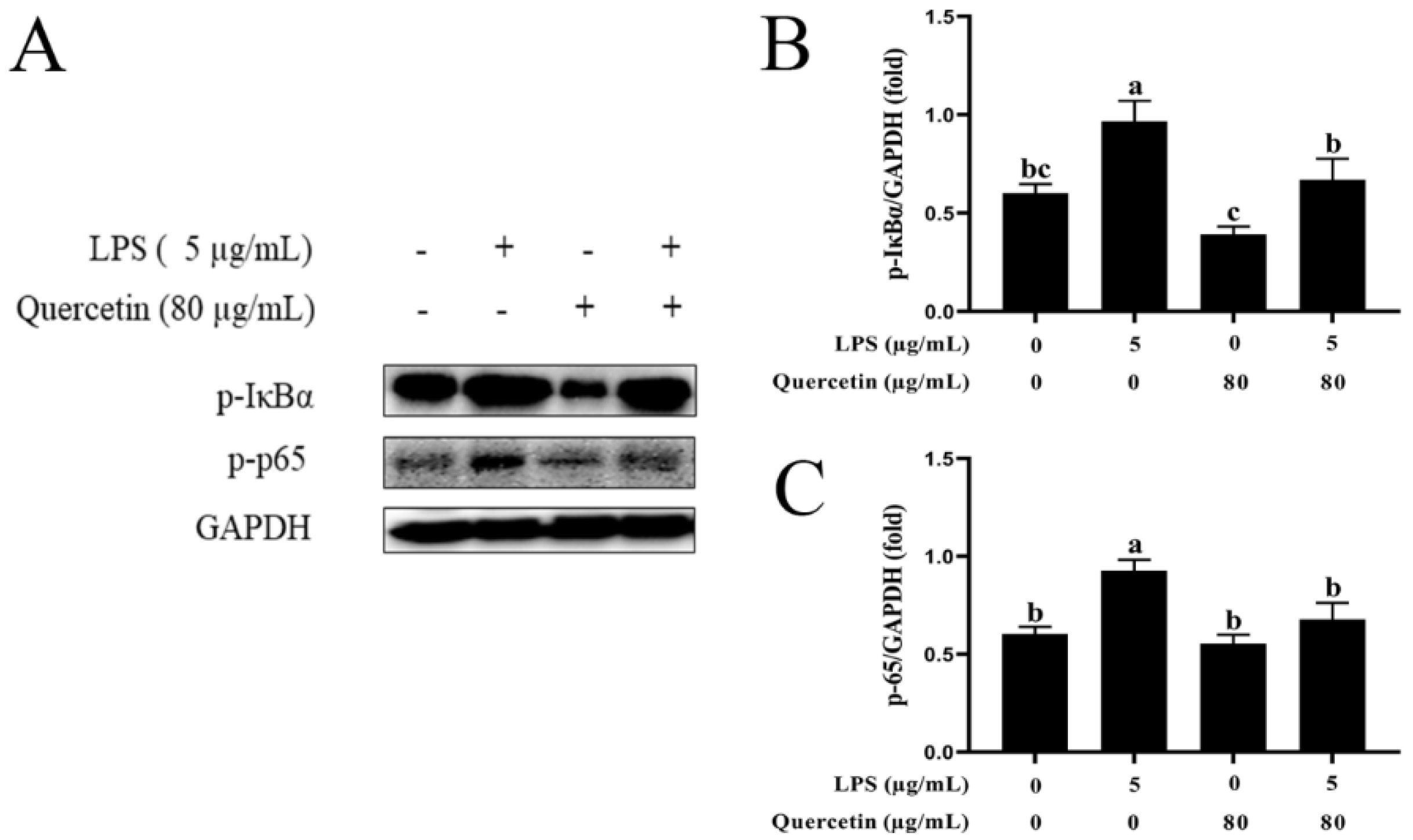
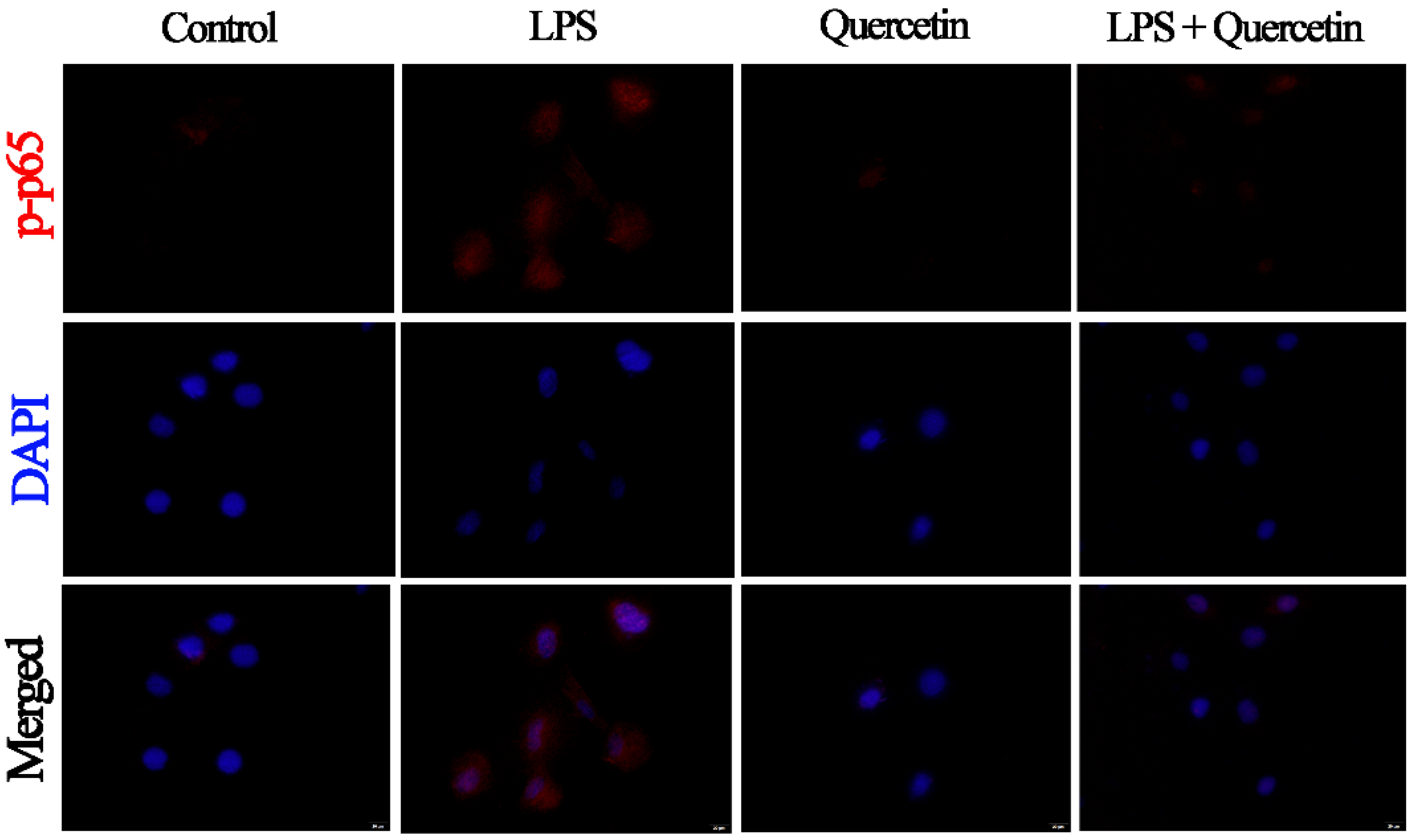
3.7. Effect of Quercetin on the NF-κB Signalling Mechanism of BIECs Caused by LPS
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Takanashi, N.; Tomosada, Y.; Villena, J.; Murata, K.; Takahashi, T.; Chiba, E.; Tohno, M.; Shimazu, T.; Aso, H.; Suda, Y.; et al. Advanced application of bovine intestinal epithelial cell line for evaluating regulatory effect of lactobacilli against heat-killed enterotoxigenic Escherichia coli-mediated inflammation. BMC Microbiol. 2013, 13, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Q.; He, X.; Zhou, S.; Shi, F.; Lu, Y. Role of PEPT1in the transport of cinnabar in Caco-2 cells. Toxicol. Vitr. Int. J. Publ. Assoc. BIBRA 2020, 63, 104747. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Liu, Z.; Jiang, J.; Yu, Y.; Zhang, Q. Differential gene expression profiling of porcine epithelial cells infected with three enterotoxigenic Escherichia coli strains. BMC Genom. 2012, 13, 330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, M.; Zimmermann, A.G.; Roberts, R.A.; Zhang, L.; Swanson, K.V.; Wen, H.T.; Davis, B.K.; Allen, I.C.; Holl, E.K.; Ye, Z.M.; et al. The innate immune sensor NLRC3 attenuates Toll-like receptor signaling via modification of the signaling adaptor TRAF6 and transcription factor NF-kappa B. Nat. Immunol. 2012, 13, 823–831. [Google Scholar] [CrossRef] [Green Version]
- Ondrackova, P.; Alexa, P.; Matiasovic, J.; Volf, J.; Faldyna, M. Interaction of porcine neutrophils with different strains of enterotoxigenic Escherichia coli. Veter-Microbiol. 2012, 160, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Otsuka, M.; Kang, Y.J.; Ren, J.; Jiang, H.; Wang, Y.B.; Omata, M.; Han, J.H. Distinct Effects of p38 alpha Deletion in Myeloid Lineage and Gut Epithelia in Mouse Models of Inflammatory Bowel Disease. Gastroenterology 2010, 138, 1255–1265.e9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fraune, S.; Anton-Erxleben, F.; Augustin, R.; Franzenburg, S.; Knop, M.; Schröder, K.; Willoweit-Ohl, D.; Bosch, T.C.G. Bacteria–bacteria interactions within the microbiota of the ancestral metazoan Hydra contribute to fungal resistance. Isme J. 2015, 9, 1543–1556. [Google Scholar] [CrossRef] [Green Version]
- Aparicio-Domingo, P.; Romera-Hernandez, M.; Karrich, J.J.; Cornelissen, F.; Papazian, N.; Lindenbergh-Kortleve, D.J.; Butler, J.A.; Boon, L.; Coles, M.C.; Samsom, J.N.; et al. Type 3 innate lymphoid cells maintain intestinal epithelial stem cells after tissue damage. J. Exp. Med. 2015, 212, 1783–1791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Citronberg, J.S.; Curtis, K.R.; White, E.; Newcomb, P.A.; Newton, K.; Atkinson, C.; Song, X.L.; Lampe, J.W.; Hullar, M.A.J. Association of gut microbial communities with plasma lipopolysaccharide-binding protein (LBP) in premenopausal women. Isme J. 2018, 12, 1631–1641. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Antony, V.; Wang, Y.; Wu, G.J.; Liang, G. Pattern recognition receptor-mediated inflammation in diabetic vascular complications. Med. Res. Rev. 2020, 40, 2466–2484. [Google Scholar] [CrossRef]
- Hu, X.Y.; Guo, J.; Zhao, C.J.; Jiang, P.; Maimai, T.; Li, Y.Y.; Cao, Y.G.; Fu, Y.H.; Zhang, N.S. The gut microbiota contributes to the development of Staphylococcus aureus-induced mastitis in mice. Isme J. 2020, 14, 1897–1910. [Google Scholar] [CrossRef] [PubMed]
- Ziesch, M.; Wente, N.; Zhang, Y.; Zaremba, W.; Engl, S.; Kromker, V. Noninferiority trial investigating the efficacy of a nonantibiotic intramammary therapy in the treatment of mild-to-moderate clinical mastitis in dairy cows with longer lasting udder diseases. J. Vet. Pharm. Ther. 2017, 41, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Teng, H.; Jia, Z.; Battino, M.; Miron, A.; Yu, Z.L.; Cao, H.; Xiao, J.B. Intracellular signaling pathways of inflammation modulated by dietary flavonoids: The most recent evidence. Crit. Rev. Food Sci. 2018, 58, 2908–2924. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yao, J.Y.; Han, C.Y.; Yang, J.X.; Chaudhry, M.T.; Wang, S.N.; Liu, H.N.; Yin, Y.L. Quercetin, Inflammation and Immunity. Nutrients 2016, 8, 167. [Google Scholar] [CrossRef] [Green Version]
- Xiong, G.; Ji, W.S.; Wang, F.; Zhang, F.X.; Xue, P.; Cheng, M.; Sun, Y.S.; Wang, X.; Zhang, T.L. Quercetin Inhibits Inflammatory Response Induced by LPS from Porphyromonas gingivalis in Human Gingival Fibroblasts via Suppressing NF-kappa B Signaling Pathway. Biomed Res Int. 2019, 2019, 6282635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, H.; Wang, Y.P.; Pan, Q.; Chen, X.; Chen, S.J.; Li, X.H.; Yao, W.G. Quercetin attenuates the proliferation, inflammation, and oxidative stress of high glucose-induced human mesangial cells by regulating the miR-485-5p/YAP1 pathway. Int. J. Immunopathol. Pharmacol. 2022, 36, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.C.; Peng, X.C.; Li, S.T.; Zhang, N.; Wang, Y.; Wei, H. Isolation and Identification of Quercetin Degrading Bacteria from Human Fecal Microbes. PLoS ONE 2014, 9, e90531. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.; Nakashima, S.; Saiki, S.; Myoi, Y.; Abe, N.; Kuwazuru, S.; Zhu, B.W.; Ashida, H.; Murata, Y.; Nakamura, Y. 3,4-Dihydroxyphenylacetic acid is a predominant biologically-active catabolite of quercetin glycosides. Food Res. Int. 2016, 89, 716–723. [Google Scholar] [CrossRef] [PubMed]
- Carrasco-Pozo, C.; Gotteland, M.; Castillo, R.L.; Chen, C. 3,4-dihydroxyphenylacetic acid, a microbiota-derived metabolite of quercetin, protects against pancreatic β-cells dysfunction induced by high cholesterol. Exp. Cell Res. 2015, 334, 270–282. [Google Scholar] [CrossRef]
- Penissi, A.B.; Rudolph, M.I.; Piezzi, R.S. Role of mast cells in gastrointestinal mucosal defense. Biocell 2003, 27, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.W.; Liu, X.Y.; Liu, M.; Chi, H.; Liu, J.R.; Han, H.M. Protective effects of quercetin and taraxasterol against H2O2-induced human umbilical vein endothelial cell injury in vitro. Exp. Ther. Med. 2015, 10, 1253–1260. [Google Scholar] [CrossRef] [Green Version]
- Zhan, K.; Yang, T.Y.; Chen, Y.Y.; Jiang, M.C.; Zhao, G.Q. Propionate enhances the expression of key genes involved in the gluconeogenic pathway in bovine intestinal epithelial cells. J. Dairy Sci. 2020, 103, 5514–5524. [Google Scholar] [CrossRef] [PubMed]
- Rakoff-Nahoum, S.; Kong, Y.; Kleinstein, S.H.; Subramanian, S.; Ahern, P.P.; Gordon, J.I.; Medzhitov, R. Analysis of gene–environment interactions in postnatal development of the mammalian intestine. Proc. Natl. Acad. Sci. USA 2015, 112, 1929–1936. [Google Scholar] [CrossRef] [Green Version]
- Clarke, A.J.; Riffelmacher, T.; Braas, D.; Cornall, R.J.; Simon, A.K. B1a B cells require autophagy for metabolic homeostasis and self-renewal. J. Exp. Med. 2018, 215, 399–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furman, D.; Chang, J.L.; Lartigue, L.; Bolen, C.R.; Haddad, F.; Gaudilliere, B.; Ganio, E.A.; Fragiadakis, G.K.; Spitzer, M.H.; Douchet, I.; et al. Expression of specific inflammasome gene modules stratifies older individuals into two extreme clinical and immunological states. Nat. Med. 2017, 23, 174–184. [Google Scholar] [CrossRef]
- Huang, W.; Wang, G.F.; Lu, S.E.; Kipen, H.; Wang, Y.D.; Hu, M.; Lin, W.W.; Rich, D.; Ohman-Strickland, P.; Diehl, S.R.; et al. Inflammatory and Oxidative Stress Responses of Healthy Young Adults to Changes in Air Quality during the Beijing Olympics. Am. J. Respir. Crit. Care 2012, 186, 1150–1159. [Google Scholar] [CrossRef] [Green Version]
- Xie, S.Z.; Liu, B.; Ye, H.Y.; Li, Q.M.; Pan, L.H.; Zha, X.Q.; Liu, J.; Duan, J.; Luo, J.P. Dendrobium huoshanense polysaccharide regionally regulates intestinal mucosal barrier function and intestinal microbiota in mice. Carbohydr. Polym. 2019, 206, 149–162. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Xiong, X.W.; Ren, K.Q.; Xu, B.; Cheng, M.; Sahu, C.; Wu, K.C.; Nie, Y.Z.; Huang, Z.; Blumberg, R.S.; et al. Deficiency in intestinal epithelial O-GlcNAcylation predisposes to gut inflammation. Embo. Mol. Med. 2018, 10, e8736. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, B.T.; Wang, F.J.; Shen, L.; Clayburgh, D.R.; Su, L.P.; Wang, Y.M.; Fu, Y.X.; Turner, J.R. LIGHT signals directly to intestinal epithelia to cause barrier dysfunction via cytoskeletal and endocytic mechanisms. Gastroenterology. 2007, 132, 2383–2394. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Zhang, M.J.; Feng, J.Q.; Fan, A.Q.; Zhou, Y.L.; Xu, Y.J. The Influence of Quercetin on Maternal Immunity, Oxidative Stress, and Inflammation in Mice with Exposure of Fine Particulate Matter during Gestation. Int. J. Env. Res. Public Health 2017, 14, 592. [Google Scholar] [CrossRef]
- Hu, X.T.; Ding, C.; Zhou, N.; Xu, C. Quercetin protects gastric epithelial cell from oxidative damage in vitro and in vivo. Eur. J. Pharm. 2015, 754, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Zerin, T.; Kim, Y.S.; Hong, S.Y.; Song, H.Y. Quercetin reduces oxidative damage induced by paraquat via modulating expression of antioxidant genes in A549 cells. J. Appl. Toxicol. 2013, 33, 1460–1467. [Google Scholar] [CrossRef] [PubMed]
- Batiha, G.E.-S.; Beshbishy, A.M.; Ikram, M.; Mulla, Z.S.; El-Hack, M.E.A.; Taha, A.E.; Algammal, A.M.; Elewa, Y.H.A. The Pharmacological Activity, Biochemical Properties, and Pharmacokinetics of the Major Natural Polyphenolic Flavonoid: Quercetin. Foods 2020, 9, 374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burmanczuk, A.; Hola, P.; Milczak, A.; Piech, T.; Kowalski, C.; Wojciechowska, B.; Grabowski, T. Quercetin decrease somatic cells count in mastitis of dairy cows. Res. Vet. Sci. 2018, 117, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.L.; Ye, Y.; Cheng, M.; Tao, Y.; Zhang, K.J.; Huang, Q.; Deng, J.W.; Yao, D.N.; Lu, C.J.; Huang, Y. Quercetin Combined with Human Umbilical Cord Mesenchymal Stem Cells Regulated Tumour Necrosis Factor-α/Interferon-γ-Stimulated Peripheral Blood Mononuclear Cells via Activation of Toll-Like Receptor 3 Signalling. Front. Pharmacol. 2020, 11, 499. [Google Scholar] [CrossRef]
- Dicarlo, M.; Teti, G.; Verna, G.; Liso, M.; Cavalcanti, E.; Sila, A.; Raveenthiraraj, S.; Mastronardi, M.; Santino, A.; Serino, G.; et al. Quercetin Exposure Suppresses the Inflammatory Pathway in Intestinal Organoids from Winnie Mice. Int. J. Mol. Sci. 2019, 20, 5771. [Google Scholar] [CrossRef] [Green Version]
- Shen, J.B.; Yang, M.Z.; Ju, D.H.; Jiang, H.; Zheng, J.P.; Xu, Z.H.; Li, L. Disruption of SM22 Promotes Inflammation After Artery Injury via Nuclear Factor kappa B Activation. Circ. Res. 2010, 106, 1351–1362. [Google Scholar] [CrossRef] [Green Version]
- Sun, A.Z.; Nie, S.J.; Xing, D. Nitric Oxide-Mediated Maintenance of Redox Homeostasis Contributes to NPR1-Dependent Plant Innate Immunity Triggered by Lipopolysaccharides. Plant Physiol. 2012, 160, 1081–1096. [Google Scholar] [CrossRef] [Green Version]
- Wlodarska, M.; Luo, C.; Kolde, R.; d′Hennezel, E.; Annand, J.W.; Heim, C.E.; Krastel, P.; Schmitt, E.K.; Omar, A.S.; Creasey, E.A.; et al. Indoleacrylic Acid Produced by Commensal Peptostreptococcus Species Suppresses Inflammation. Cell Host Microbe 2017, 22, 25–37.e26. [Google Scholar] [CrossRef] [Green Version]
- Wen, L.; Javed, T.A.; Yimlamai, D.; Mukherjee, A.; Xiao, X.W.; Husain, S.Z. Transient High Pressure in Pancreatic Ducts Promotes Inflammation and Alters Tight Junctions via Calcineurin Signaling in Mice. Gastroenterology 2018, 155, 1250–1263.e5. [Google Scholar] [CrossRef]
- Holland, W.L.; Bikman, B.T.; Wang, L.P.; Yuguang, G.; Sargent, K.M.; Bulchand, S.; Knotts, T.A.; Shui, G.H.; Clegg, D.J.; Wenk, M.R.; et al. Lipid-induced insulin resistance mediated by the proinflammatory receptor TLR4 requires saturated fatty acid-induced ceramide biosynthesis in mice. J. Clin. Investig. 2011, 121, 1858–1870. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.C.; Yeh, W.C.; Ohashi, P.S. LPS/TLR4 signal transduction pathway. Cytokine 2008, 42, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.L.; Liu, G.L.; Yuan, Y.Y.; Wu, G.T.; Wang, S.L.; Yuan, L.W. NEK7 interacts with NLRP3 to modulate the pyroptosis in inflammatory bowel disease via NF-κB signaling. Cell Death Dis. 2019, 10, 906. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.F.; Zhang, X.; He, X.J.; Yang, B.; Wang, H.; Shan, X.F.; Li, C.Q.; Sun, D.B.; Wu, R. LPS-induced reduction of triglyceride synthesis and secretion in dairy cow mammary epithelial cells via decreased SREBP1 expression and activity. J. Dairy Res. 2018, 85, 439–444. [Google Scholar] [CrossRef] [PubMed]
- Qian, C.; Cao, X.T. Regulation of Toll-like receptor signaling pathways in innate immune responses. Ann. N. Y. Acad. Sci. 2013, 1283, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Woodall, M.C.; Woodall, B.P.; Gao, E.H.; Yuan, A.; Koch, W.J. Cardiac Fibroblast GRK2 Deletion Enhances Contractility and Remodeling Following Ischemia/Reperfusion Injury. Circ. Res. 2016, 119, 1116–1127. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence 1, 5′ to 3′ | Source | Size (bp) |
|---|---|---|---|
| GAPDH | F: GGGTCATCATCTCTGCACCT R: GGTCATAAGTCCCTCCACGA | NM_001034034.2 | 176 |
| IL-1β | F: CAGTGCCTACGCACATGTCT R: AGAGGAGGTGGAGAGCCTTC | NM_174093.1 | 209 |
| IL-6 | F: CACCCCAGGCAGACTACTTC R: TCCTTGCTGCTTTCACACTC | NM_173923.2 | 129 |
| TNF-α | F: GCCCTCTGGTTCAGACACTC R: AGATGAGGTAAAGCCCGTCA | NM_173966.3 | 192 |
| CXCL5 | F: TGAGACTGCTATCCAGCCG R: AGATCACTGACCGTTTTGGG | NM_174300.2 | 193 |
| CCL5 | F: CTGCCTTCGCTGTCCTCCTGATG R: TTCTCTGGGTTGGCGCACACCTG | NM_175827 | 217 |
| CCL2 | F: GCTCGCTCAGCCAGATGCAA R: GGACACTTGCTGCTGGTGACTC R: GCACAACTTGTTTCACCCACT | NM_174006 | 171 |
| CXCL2 | F: CCCGTGGTCAACGAACTGCGCTGC R: CTAGTTTAGCATCTTATCGATGATT | NM_174299.3 | 204 |
| CXCL8 | F: TGGGCCACACTGTGAAAAT R: TCATGGATCTTGCTTCTCAGC | NM_173925.2 | 136 |
| ZO-1 | F: TCTGCAGCAATAAAGCAGCATTTC R: TTAGGGCACAGCATCGTATCACA | XM_010817146.1 | 187 |
| Occludin | F: GAACGAGAAGCGACTGTATC R: CACTGCTGCTGTAATGAGG | NM_001082433.2 | 122 |
| Claudin 1 | F: CGTGCCTTGATGGTGAT R: CTGTGCCTCGTCGTCTT | NM_001001854.2 | 102 |
| Claudin 4 | F: CTTCATCGGCAGCAACATC R: ACAACAGCACGCCAAACA | NM_001014391.2 | 191 |
| TLR2 | F: CAGGCTTCTTCTCTGTCTTGT R: CTGTTGCCGACATAGGTGATA | NM_174197.2 | 140 |
| TLR4 | F: GACCCTTGCGTACAGGTTGT R: GGTCCAGCATCTTGGTTGAT | NM_174198.6 | 103 |
| CD14 | F: CAGTATGCTGACACAATCAA R: AGTTCCTTGAGACGAGAGTA | NM_174008.1 | 122 |
| MD2 | F: GGAGAATCGTTGGGTCTGC R: GCTCAGAACGTATTGAAACAGGA | NM_001046517.1 | 92 |
| MyD88 | F: TCATTGAGAAGAGGTGCCGT R: TGGCTTGTACTTGATGGGGAT | NM_001014382.2 | 146 |
| IRF3 | F: TTGTGAACTCAGGAGTCAGG R: TGGGCTCAAGTCCATGTCAC | NM_001029845.3 | 125 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gong, X.; Huang, Y.; Ma, Q.; Jiang, M.; Zhan, K.; Zhao, G. Quercetin Alleviates Lipopolysaccharide-Induced Cell Damage and Inflammation via Regulation of the TLR4/NF-κB Pathway in Bovine Intestinal Epithelial Cells. Curr. Issues Mol. Biol. 2022, 44, 5234-5246. https://doi.org/10.3390/cimb44110356
Gong X, Huang Y, Ma Q, Jiang M, Zhan K, Zhao G. Quercetin Alleviates Lipopolysaccharide-Induced Cell Damage and Inflammation via Regulation of the TLR4/NF-κB Pathway in Bovine Intestinal Epithelial Cells. Current Issues in Molecular Biology. 2022; 44(11):5234-5246. https://doi.org/10.3390/cimb44110356
Chicago/Turabian StyleGong, Xiaoxiao, Yinghao Huang, Qianbo Ma, Maocheng Jiang, Kang Zhan, and Guoqi Zhao. 2022. "Quercetin Alleviates Lipopolysaccharide-Induced Cell Damage and Inflammation via Regulation of the TLR4/NF-κB Pathway in Bovine Intestinal Epithelial Cells" Current Issues in Molecular Biology 44, no. 11: 5234-5246. https://doi.org/10.3390/cimb44110356