
Lipid Nanoparticles: A Novel Gene Delivery Technique for Clinical Application

1
Department of Clinical Laboratory Medicine, National Center for Child Health and Development, 2-10-1 Okura, Setagaya-ku, Tokyo 157-8535, Japan
2
Department of Systems BioMedicine, National Research Institute for Child Health and Development, 2-10-1 Okura, Setagaya-ku, Tokyo 157-8535, Japan
*
Author to whom correspondence should be addressed.
Curr. Issues Mol. Biol. 2022, 44(10), 5013-5027; https://doi.org/10.3390/cimb44100341
Submission received: 23 September 2022
/
Revised: 8 October 2022
/
Accepted: 17 October 2022
/
Published: 19 October 2022
(This article belongs to the Special Issue Effects of Nanoparticles on Living Organisms)
Abstract
:Lipid nanoparticles (LNPs) are an emerging vehicle for gene delivery that accommodate both nucleic acid and protein. Based on the experience of therapeutic liposomes, current LNPs have been developed based on the chemistry of lipids and RNA and on the biology of human disease. LNPs have been used for the development of Onpattro, an siRNA drug for transthyretin-mediated amyloidosis, in 2018. The subsequent outbreak of COVID-19 required a vaccine for its suppression. LNP-based vaccine production received much attention for this and resulted in great success. In this review, the essential technology of LNP gene delivery has been described according to the chemistry for LNP production and biology for its clinical application.
1. Introduction
Lipid nanoparticles (LNPs) represent a newly emerging type of drug formulation that encapsulates biological molecules such as nucleic acids and proteins, as well as a complex made with both [1,2,3]. LNPs are spherical and are visible in electron microscopy. Therapeutic LNPs are less than 100 nm in diameter, consisting of lipids and payloads such as nucleic acids. The initial idea of LNPs stems from liposome, a much simpler lipid vesicle made with phospholipid and cholesterol and larger in size than the LNP. Liposomes were modeled based on the cell membrane under lipid bilayer theory. Liposomes have been used for research into the physical chemistry of lipids in aqueous solution and have been examined for future clinical use. For the preparation of liposomes, lipids are usually dried by a rotary evaporator, suspended within an aqueous solution, and sonicated to obtain a multi-lamellar vesicle that appears as a milky suspension.
Modern LNPs are more sophisticated and are essentially made with four different species of lipids (Table 1). The preparation procedure for LNPs may be similar to these but optimized by recent findings [4]. For LNP preparation, lipids and RNA are separately dissolved in ethanol and acidic aqueous solution, respectively. Next, they are mixed with an automated microfluidics apparatus for industrial use or a pipette for research use. Then, ethanol is removed by dialysis. In most industrial applications, several chromatographic purification procedures are required to increase the authenticity of the final LNP product. The final LNP composition is examined based on the percentage of RNA encapsulation, the diameter of the LNP, its zeta potential, and other biophysical parameters. Zeta potential represents the stability of the LNP. To optimize these acquired parameters, the polydispersity index (PDI) is used, which measures the heterogeneity of macromolecules including LNP; a value of less than 0.1 has been generally accepted as well-optimized conditions. When LNPs are formulated, lipids are used excessively over RNA (approximately 10:1 in terms of weight ratio).
LNPs in an FDA-approved formulation target the liver [2,8,9]. Upon intravenous administration, the LNPs migrate to the liver through the bloodstream, followed by incorporation into hepatocytes via an LDL receptor or an asialoglycoprotein receptor. During this migration, LNPs may associate with apolipoprotein E. Hepatic sinusoids contain numerous fenestration structures approximately 100 nm in diameter [8]. Thus, the size of the LNP needs to be smaller than this value. Note that the diameter of chylomicron, a dietary lipid droplet taken up by the intestine, is 100–1000 nm; hence, these are too large to be incorporated by the hepatocytes. In contrast, adeno-associated viruses (AAV) has an approximately 25 nm diameter; therefore, even if non-liver directed serotypes are chosen, the hepatotoxicity of AAV may become incidentally apparent.
Lastly, some previously used liposomal components are derived from naturally occurring lipids such as those originating in soy and eggs. These may contain a variety of analogs with different chain lengths and numbers of double bonds as well as isomers with different stereo- and regio-configurations. This composition may vary depending on the source of origin, diet, and even the season of harvest. To eliminate these factors, currently used lipids for LNPs are highly pure.
2. Lipids
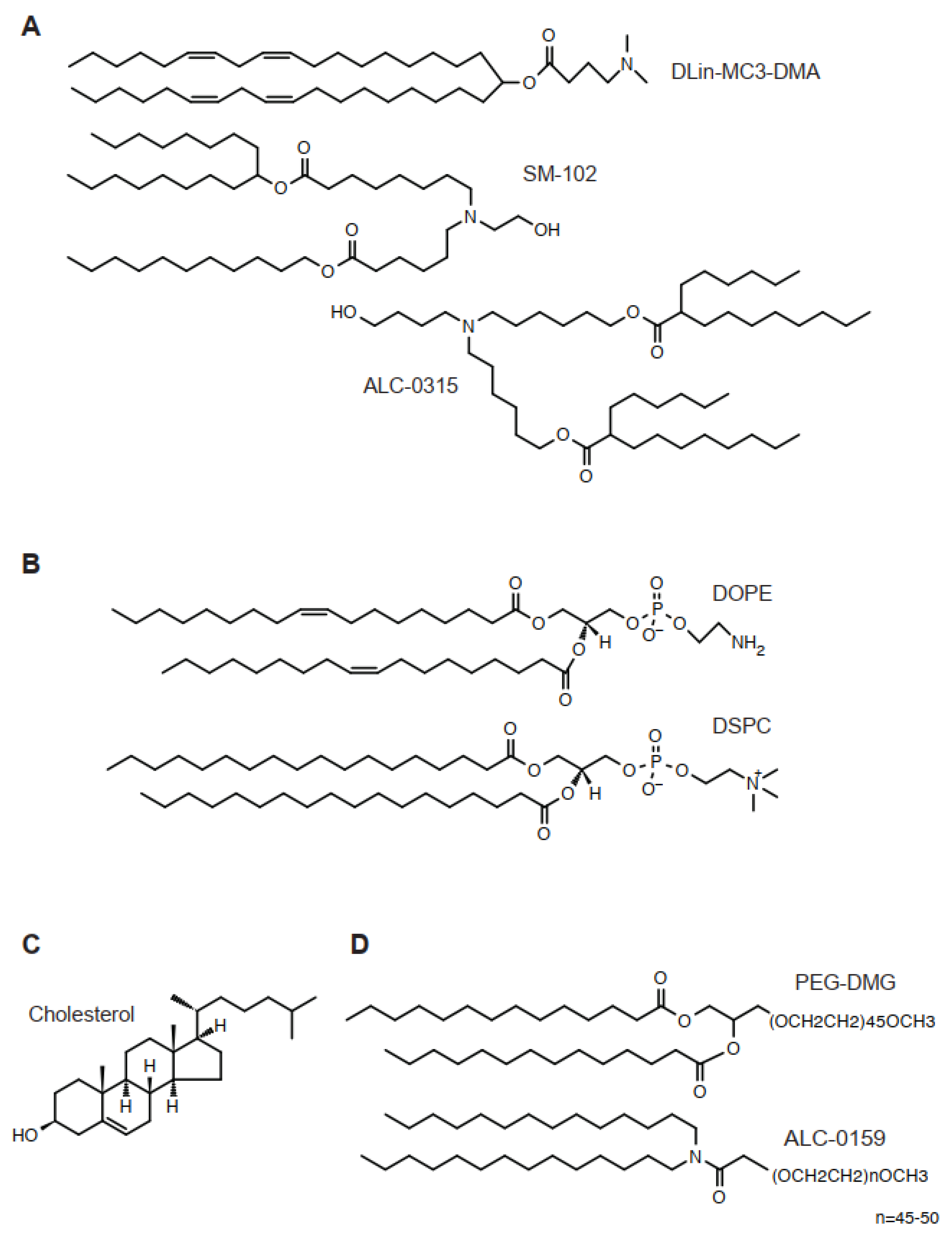
The lipid is an essential component in LNPs, encapsulating nucleic acids and other components [10] (Figure 1). For classical formulation, four lipids are required, such as an ionizable cationic lipid, a helper lipid, cholesterol, and a PEG-lipid. The classical liposome usually required cholesterol and phospholipids, sometimes accompanied by some additional, minor components. In contrast, the molar ratio of an ionizable cationic lipid of LNPs constitutes approximately 50% of the total lipids. The best molar ratio of LNPs depends on diameter, encapsulation of nucleic acids, zeta potential, and the length of encapsulated nucleic acids. The inclusion of novel lipids requires individual optimization. The classical composition of lipids targets the liver; thus, there is a solid strategy when other tissues—such as the spleen, lung, or hematopoietic cells—are to be targeted. Below are essential components for LNPs.
2.1. Ionizable Cationic Lipids
An ionizable cationic lipid, such as DLin-MC3-DMA, is a tertiary amine with hydrophobic tails (Figure 1). In FDA-approved formulation, this lipid constitutes half of all lipids (Table 1). Overall, the difference of structures at the initial stage of development is often found in the linker structure, which connects the amine and hydrophobic tails. In contrast, the number of double bonds, together with their geometry, has not been discussed as a major determinant for the physical properties of LNPs [11]. Biologically degradable ionized cationic lipids tend to have many esters or thioethers.
2.2. Helper Lipids
2.3. Cholesterol
Cholesterol is an essential component of LNP. It is used at a concentration of 30–40% of total lipids depending on the molar ratio of other lipid types. Cholesterol is known to maintain LNP in a rigid form.
2.4. PEG-Lipids
Added PEG-lipids make up 1–5% of total lipids. At these concentrations, PEG lipids make the LNP particle much smaller. PEG lipids localize at the surface of the LNP.
3. RNA
RNA is an essential payload for LNP. Its efficacy is proven by the several vaccine products for COVID-19 with tremendous benefits for humans. RNA has long been thought to be an unstable biomolecule. However, an accumulating knowledge of ribonucleotides, nucleases, and the molecular mechanism of inflammation collectively is establishing an improved understanding on the stability of RNA (Table 2). Uridine is known to induce interferon production through Toll-like receptor-dependent NF-kB activation; thus, its replacement to a variety of modified uridine analogs including N1-methylpseudouridine leads to minimal cell damage in RNA-delivered cells. Among various nucleases with different substrate specificity, exonucleases have been identified as among the key factors for RNA instability. Thus, 5′-capping and/or chemical modification of minimal (usually three) nucleotides from both 5′- and 3′-end is effective for extended RNA stability. When cells are activated by RNA, an activation of RNase L, protein kinase R, and MDA5-MAVS pathways are activated, leading to apoptosis.
3.1. Modification of mRNA
3.1.1. 5′-Capping
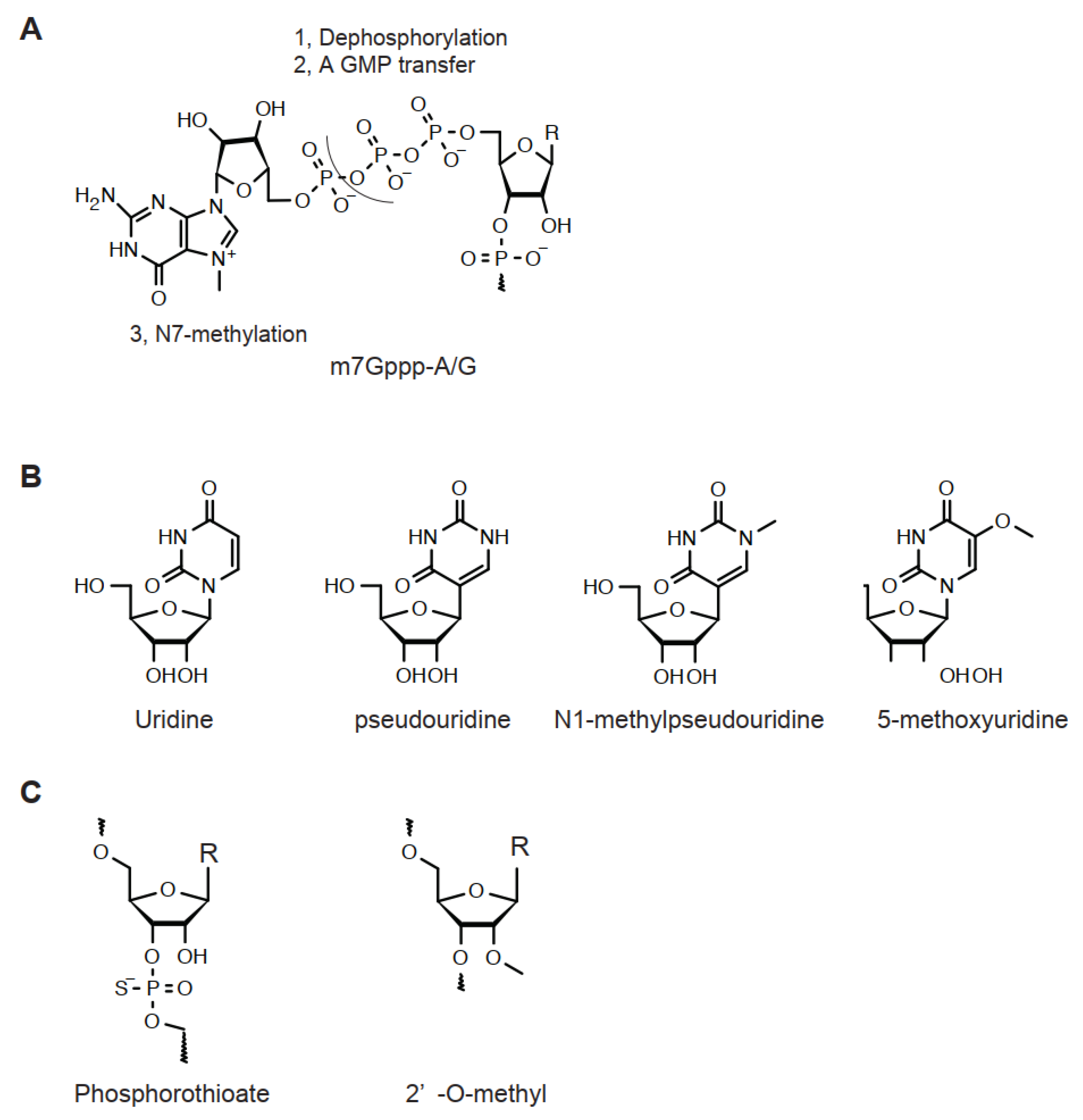
The 5′-end of mRNA is enzymatically modified and known as a 5′-capping structure (Figure 2A) [14,15]. To biochemically achieve this unique structure, a g-phosphate group of first transcription nucleoside is dephosphorylated, followed by transferring a GMP, leading to a unique 5′-Gppp-A/G structure (Figure 2A, Table 3). Then, 2,2,7-trimethylguanosine methyltransferase transfers a methyl group to the N7-position of non-first-transcribed nucleoside guanosine (Cap 0). Subsequent biochemical reactions may occur simultaneously. In Cap 1 and Cap 2 reactions, the 2′-O-position of the ribose of the first and second transcription nucleoside is methylated by 5′-capping methyltransferase 1 and 2, respectively (Cap 1 and Cap 2). In addition, an N2-methylation of guanine of non-first-transcribed nucleoside gives rise to N7, N2-dimethylated and N7, N2,2′-trimethylated guanine by 2,2,7-trimethylguanosine synthase, respectively. In parallel, N2-methylation of adenine is catalyzed by S-adenosylmethionine transferase. Once the Cap 0 structure is generated, this adenine may be further N6-methylated by an enzyme cap-specific adenosine N6-methyltransferase.
3.1.2. Uridine Analog, Codon Optimization, and PolyA
In mRNA, native bases of RNA strongly activate innate immunity [16,17]. Thus, these must be modified by an analog with low immunogenicity for therapeutic purposes [18,19,20]. Uridine is a well-known example. Currently, a series of uridine analogs including pseudouridine, N1-methylpseudouridine, and 5′-methoxyuridine are used for therapeutic mRNA preparation (Figure 2B). For in vivo, N1-methylpseudouridine effectively extends the half-life of mRNA [19]. In addition, to replace uridine to its analog, an in silico procedure to reduce the number of uridines, codon optimization, may be performed. For the polyA tail, its length is correlated with mRNA stability [18].
3.2. Modification of Single Guide RNA (sgRNA)
The CRISPR/Cas9 system requires a guide RNA for the genome editing reaction. An sgRNA for CRISPR/Cas9 has a composed nucleotide sequence of a guide RNA and a trans-acting RNA. Usually these are used as a single strand RNA of approximately 100 nt. When extended half-life is desired, 2′-O-methylation and 5′/3′-phosphorothioation are performed [18,21,22] (Figure 2C). These modifications decrease susceptibility to exonucleases. 2′-O-fluorination is also used as an alternative modification for 2′-O-methylation.
4. Genetic Material
As described earlier, conventional formulation of LNPs has been demonstrated to deliver genetic material to the liver (Table 1). Such a payload includes siRNA and mRNA encoding therapeutic cDNA, Cas9 nuclease, and its related hybrid enzymes including base editor and prime editor. A ribonucleotide-protein particle (RNP) is a complex of protein and RNA with a certain stoichiometry. When the CRISPR/Cas9 system is to be used, a Cas9 nuclease forms an RNP with a guide RNA with a 1:1 molar ratio. To obtain the best results in vivo, it is desirable that the initial amount of RNA is empirically examined.
4.1. siRNA
Transthyretin-mediated amyloidosis is a major disease subtype of amyloidosis, and more than 30 genes have been identified as its cause [5]. While transthyretin circulates as a tetramer under normal conditions, under pathogenic conditions it is no longer metabolized properly and accumulates as amyloids in the liver. Disruption of such an accumulating pathogenic amyloid by genetic techniques is an effective treatment strategy for this disorder. Consistent with this idea, LNP-loaded siRNA for transthyretin effectively removes this [23,24]. These studies showed that, under optimized conditions, LNP-formulated siRNA for transthyretin showed high effectiveness in humans [5]. Based on this evidence, the drug Onpattro was approved by the FDA in 2018 [25]. Another example includes hypercholesterolemia, a metabolic disorder of lipids that occurs with high frequency. Pcsk9 is a serum protein that binds to an LDL receptor. Thus, an elimination of Pcsk9 followed by decreasing LDL in circulation is an effective therapy for hypercholesterolemia. As expected, such treatment effectively lowers serum LDL concentration in LNP-based models [23].
4.2. mRNA
4.2.1. Therapeutic cDNA
4.2.2. Cas9 Nuclease
CRISPR/Cas9 is a leading tool for genome editing in vitro and in vivo [29,30]. Gene delivery of Cas9 in vivo needs to have some vehicle. This is readily applicable to LNP, because the size of Cas9 mRNA is approximately 4 kb. In sharp contrast, in AAV, a mainstream vector for current gene therapy, it is difficult to accommodate a wild-type Cas9 cDNA in a single vector because it is too large; hence, a two-vector system has been developed [29]. Consequently, due to the relative technical ease of Cas9 preparation, several liver disorders can be treated with LNP-mediated gene delivery. Hypercholesterolemia might be involved in one such disorder. In this case, the pathogenic Pcsk9 gene is disrupted, leading to sub-normal serum cholesterol concentration [24,31,32]. LNP-loaded Cas9 for transthyretin effectively removes this, as described above [33].
4.2.3. Base Editor/Prime Editor
Adenine base editor and prime editor is an attractive research toolkit in genome editing [29]. Essentially, these are modified Cas9 with adenosine deaminase Tad7 for an adenosine base editor or reverse transcriptase for a prime editor, respectively. For rational optimization, their tertiary structure has been extensively studied [34]. As discussed above, the size of mRNA for base editors and prime editors is not suitable for a single AAV vector system. Thus, the treatment of some liver disorders has been examined using LNPs. The first application of this technology is hypercholesterolemia by targeting Pcsk9 [35,36]. Similarly, corrections of the pathogenic mutation of phenylalanine hydroxylase for phenylketonuria [37] and fumarylacetoacetate hydrolase for tyrosinemia I [38] have been examined.
5. Preclinical Study
5.1. Liver Disorders
The original formulation of LNPs preferentially target the liver. Based on this fact, liver disorders have been considered as good targets for LNP-mediated therapeutics (Table 4). Among various liver disorders, familial hypercholesterolemia (FH) is a genetic disorder involving multiple genes, such as LDLR, ApoB, and Pcsk9 [39]. LNP-based siRNA therapy has been examined for ApoB [40] and Pcsk9 [23] in murine and primate models. Pathogenic genes have also been corrected using Cas9 nuclease delivered by LNPs [24,31,32].
5.2. Anti-Cancer Therapy
An anti-cancer strategy based on the presentation of tumor peptides on the MHC class II by dendritic cells has been studied for more than a decade [50]. Based on the improvements of LNP delivery, its use in a conventional formulation has been explored [51]. Essentially, encapsulated mRNA for peptides of the tumor antigen was first transfected into dendritic cells, followed by administration to the melanoma model. The replacement of uridine with pseudouridine and cytidine with 5-methylcytidine reduced unselective immune activation. A more recent study reported the use of a novel amino-lipid with bio-degradative properties [52]. This LNP effectively activates CD8+ T-cells of lymph nodes in immunized mice, leading to an improving eradication of antigen-bearing melanoma in the animal model.
Near infrared (NIR) radiation-activatable LNP with a pH-sensitive dye is an emerging technique [53,54]. A recent study reported the synthesis of a novel PEG-BODIPY dye that can be used for LNP [55]. Encapsulated LNPs with this dye are incorporated into cells like other PEG-lipids, followed by the release of mRNA and the dye from the endosomal LNP into the cytosol. The released dye, rather than the PEG lipid, is then selectively activated by the NIR radiation; thus, a therapeutic outcome is expected. In this case, no targeting ligand is required for LNP, because the tumor cells are selectively killed by NIR irradiation under tumorigenic pH where this newly synthesized dye is active. These authors demonstrated that a 50:50 ratio of PEG-DMG to PEG-BODIPY dye gave the best results when examined in a subcutaneous SUM159 breast cancer xenograft model [53,56].
5.3. Congenital Disorders
Lysosomal storage disorders (LSDs) are characterized by an accumulation of unprocessed biochemicals in the lysosome involving 50–60 enzymes [57,58,59]. These disorders present CNS, skeletal, and visceral disorders such as hepatomegaly and splenomegaly. Fabry disease is one such LSD, with renal, cardiac, and CNS manifestations caused by pathogenic mutation of α-galactosidase A (GLA). In 2019, Zhu et al. reported that T7 RNA polymerase-transcribed therapeutic GLA mRNA with 5-methoxyuridine in LNP was administered to mice and primates, with promising results [42]. At the same time, DeRosa et al. reported that an administration of LNP-encapsulated T7 RNA polymerase-transcribed GLA mRNA with a 5′ cap structure (Cap 1) increased GLA enzyme activity more than 1000-fold compared to physiological levels [43]. More recently, Rodríguez-Castejón et al. reported that administration of a complex of plasmid DNA and protamine in LNP also provided a therapeutic effect [60]. These results raise the possibility that visceral manifestation of Fabry disease could be treated with LNP-mediated therapeutic strategies.
5.4. Bleeding Disorder
Factor VII is involved in the clotting process of the blood. Thus, a failure of this leads to bleeding manifestations. In the murine model, an ED50 of 0.005 mg/kg siRNA was achieved in optimized LNP composition.
5.5. Hemophilia A and B
Hemophilia A and B are caused by a defective activation of Factor VIII and IV, respectively, leading to improper activation of thrombin by antithrombin encoded by the SERPINC1 gene. Thus, the degradation of this gene is expected to treat both hemophilia A and B. In a murine model, the inactivation of Serpinc1 gene by LNP-encapsulated siRNA has been examined with a therapeutic outcome.
5.6. Duchenne Muscular Dystrophy (DMD)
DMD is a muscular disorder. DMD in approximately 10% of affected individuals is caused by pathogenic exon skipping. In these cases, an alteration of this followed by restoration of wild-type reading frame is known to be therapeutic in DMD even if some amino acids are still missing. Based on this fact, an originally established murine DMD model harboring human DMD exon 45 by a knock-in strategy was corrected by LNP-encapsulated Cas9 mRNA.
5.7. Human Immunodeficiency Virus (HIV)
HIV is a member of the Retroviridae family, which causes human immunodeficiency. Antibody-mediated neutralization of HIV is one effective therapeutic strategy. N. Pardi et al. administered LNP-encapsulated mRNA for both light and heavy chains of monoclonal antibody against HIV. An administration of 1.4 mg/kg of mRNA into mice resulted in an approximately 170 mg/mL monoclonal antibody. The efficacy of this treatment was further demonstrated with the intravenous HIV-1-challenge in a humanized HIV murine model.
6. Clinical Study
According to the results of preclinical studies, more than 10 clinical studies are now ongoing (Table 5). Most notably, the clinical study of LNP-encapsulated siRNA for transthyretin has been completed, and this agent was approved by the FDA in 2018. Similar to this success, a clinical trial of genome-editing therapy is ongoing. Apart from these, other trials mainly involve anti-cancer therapy. The administration of an LNP-encapsulated payload is delivered into the body either intravenously or intratumorally. Among many cancers, most are liver cancers followed by lung cancers, consistent with the effectiveness of LNP for liver-targeted delivery vehicle.
6.1. Transthyretin
6.1.1. siRNA
Patisiran is an approved RNA-interference therapeutic for hereditary transthyretin amyloidosis, a type of amyloidosis with a high prevalence of known etiology among more than 30 genes. The study was performed with patients with hereditary transthyretin amyloidosis at 0.3 mg/kg once every 3 weeks; [5,62]. Patients aged between 18 and 85 years old with a diagnosis of hATTR were recruited from 44 sites across 19 countries [5,62] and received Patisiran at 0.3 mg/kg intravenously once every 3 weeks for 18 months. Under this condition, the mean changes in serum transthyretin concentration decreased at 3 weeks. At 18 months, Patisiran improved multiple manifestations such as motor, sensory, and autonomic neuropathy collectively measured with an index called the Neuropathy Impairment Score+7 (mNIS + 7). Overall, other parameters including cardiac function and quality of life were improved compared to controls [56,63,64].
6.1.2. Cas9 Nuclease
The results of Phase I of the clinical trial of CRISPR-Cas9-mediated in vivo gene editing for transthyretin amyloidosis (NTLA-2001) was recently reported [61]. In this study, an observation of the first 28 days of treatment at 0.1–0.3mg/kg reported few adverse events with a reduction of mean serum transthyretin concentration at 52% at 0.1 mg/kg (n = 3) and 87% at 0.3 mg/kg (n = 3) in humans (NCT04601051). This clinical study was designed based on the preclinical study in the non-human primates at 1.5–6.0 mg/kg with no adverse events up to 12 months. There were seven putative targeting sites for this therapeutic CRISPR-Cas9 system. The authors reported that all these loci were located in non-coding regions as determined using established software such as Cas-OFFinder, GUIDE-seq, and SITE-seq, and there was no evidence of off-targeting when examined using primary human hepatocytes with a three-fold higher concentration of NTLA-2001.
6.2. Solid Tumor
Checkpoint inhibition through an interaction between PD-1/PD-L1 and CTLA-4 plays a key role in activating the tumor-killing activity of T cells [66]. Therefore, therapeutic cancer vaccines educate the immune system of tumor-bearing patients in response to individual tumor-specific immunity. A clinical study based on this mechanism using LNP as a vehicle for therapeutic mRNA is now ongoing (NCT03739931) [66].
6.3. Hepatic Fibrosis
Hepatic fibrosis is caused by a failure to maintain the balance between a wound process caused by inflammation and the subsequent healing process in the liver. A Phase 1b/2 clinical trial has been performed. In this study, 25 participants with moderate to extensive hepatic fibrosis were enrolled, and the safety and tolerability of LNP-encapsulated siRNA for HSP47 was examined (NCT02227459). The siRNA agent was administered to the subjects aged between 18 to 75 years old once or twice a week for 5 weeks.
6.4. Ornithine Transcarbamylase Deficiency (OTCD)
OTCD is an inherited metabolic disorder in the liver. The deficiency of ornithine transcarbamylase fails to synthesize citrulline at the second step of urea cycle, leading to hyperammonemia. Administration of therapeutic mRNA for ornithine transcarbamylase has been examined (NCT04416126/NCT04442347/NCT05526066). In a Phase 1 trial, 30 subjects aged between 18 to 65 years old were enrolled for a single ascending dose study with no reported adverse events.
7. Tissue Targeting of LNP
Tissue targeting is another issue for LNPs. It is well known that the initial formulation of LNP effectively delivers the payload to the liver (Table 1). Consistently, accumulating examples in preclinical and clinical trial support this evidence (Table 4 and Table 5). For future expansion of LNP as the vehicle of a drug delivery system, numerous studies are been currently performed. Actually, such a systematic project has been known. Selective organ targeting (SORT) is an idea for tissue targeting of LNPs proposed by Siegwart et al. [11,31,67,68,69] (Table 6). These authors hypothesized that: (1) the biodistribution of LNPs to the liver and other organs is likely to be different for uncharacterized reasons, (2) an acid dissociation constant (pKa) close to 6.4 of amine of an ionizable cationic lipid is required for the efficient translocation of LNPs from the endosome to cytosol, and (3) absorption of LNPs by the liver is, at least in part, mediated by apolipoprotein E-mediated modification in the blood [67]. Due to limited data, a series of different lipids has been synthesized, and their tissue targeting has been examined.
7.1. Dendrimers
A dendrimer is a globular synthetic chemical with branched hydrophobic moiety. According to the SORT hypothesis, a series of novel ester-based dendrimers, such as 5A2-SC8, have been synthesized [67,68] (Table 6). During the course of a series of these studies, the authors noticed that an increase in DOTAP enhanced the targeting of LNPs to the lung [31]. Separately, it was also found that an inclusion of C18 phosphatidic acid enhanced LNP targeting to the spleen [67,68]. Currently, these are known as SORT molecules, which modulate the targeting property of LNPs. More recently, researchers have explored the possibility of a novel class of alkenyl-thiols [31] and multi-tailed ionizable phospholipids [69] for expanding the SORT hypothesis. For organ cell specificity, a liver-directed formulation mainly delivered LNP-mediated genes into hepatocytes. In contrast, 66% of pulmonary endothelial cells expressed exogenous genes. In the spleen, LNP-mediated genes were expressed in 20% of macrophages, 12% of B cells, and 10% of T cells [68].
In a separate formulation of LNPs by Dahlman and colleagues, more than 50% of splenic endothelial cells were able to express exogenous genes [70]. In this experiment, each LNP was individually barcoded, and a mixture encapsulating Cre mRNA was administered to Ai14 mice. Then, fluorescence-active cells in the spleen were FACS-sorted based on the expression of CD31, an endothelial marker. The researchers found an accumulation of a lipid 7C3 in these cells, concluding that this formulation provided the selective LNP capable of consistently delivering exogenous genes to the spleen. To estimate the efficiency of gene delivery by this LNP, these authors chose ICAM-2, an abundantly expressed surface protein on endothelial cells, and examined the efficiency of genome editing by CRISPR/Cas9. They found that 13–20% of indel were observed in these splenic endothelial cells, as well as hepatocytes, indicative of a shared mechanism of cellular incorporation between them.
7.2. Polymers
Several studies have tested the possibility of polyesters for extrahepatic targeting of LNPs [71]. Based on the screening of a library with 1200 candidates, the best formulation (PE4k-A17-0.2C8) with an optimal length of polymer, amine, and LNP composition based on the intensity of luciferase was uncovered [72]. Overall, the selected nanoparticle has a neutral surface charge, a diameter of approximately 100 nm, a pKa between 7.5 and 8.0, and more than 90% of RNA-binding activity. This compound exclusively delivered Cre mRNA to the lung, as demonstrated by tdTomato fluorescence mice. Besides its detailed mechanism of pulmonary targeting, this excellent targeting specificity for the lung provides a further starting point for the structure-activity study of LNPs.
7.3. Antibody-Modified LNPs
A study has examined the treatment of cardiac fibrosis by targeting LNPs to T cells [73]. In this study, LNP-encapsulated mRNA for a chimeric antigen receptor (CAR) against the fibrosis activation protein (FAP) was first delivered to T cells via anti-CD5 antibody-modified LNP. The expressed FAP-CAR in the T cells then reacts with FAP in a diseased fibroblast, leading to a therapeutic outcome.
8. Future Perspective
Current LNP technology has an advantage in the encapsulation of larger mRNA for therapeutic use compared to AAV. Furthermore, LNPs are capable of delivering a gene into non-dividing cells. Specifically, for liver disease, there are multiple examples where LNP-based therapy has proven effective in preclinical and clinical studies (Table 4 and Table 5). However, there are also limitations of LNP at this stage discussed below. For example, biodegradation of LNP is one such issue to be addressed. Specifically, one current problem with LNPs is that several lipids are less susceptible to being biologically degradable; thus, these lipids create a concern of hepatotoxicity. In an attempt to accelerate their effective biodegradation, regardless of molecular weight and chemical nature, the introduction of ester and thioester into these compounds may prove beneficial.
Author Contributions
Conceptualization, R.M.; scientific input of genome editing, S.T.; writing—original draft preparation, R.M.; writing—review and editing, R.M. and S.T.; writing—finalization, R.M. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by a grant-in-aid from AMED to R.M. (22ae0201004h0005) and a grant-in-aid from JSPS to R.M. (22K07927).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kulkarni, J.A.; Cullis, P.R.; van der Meel, R. Lipid Nanoparticles Enabling Gene Therapies: From Concepts to Clinical Utility. Nucleic Acid Ther. 2018, 28, 146–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, M.H.Y.; Brimacombe, C.A.; Verbeke, R.; Cullis, P.R. Exciting Times for Lipid Nanoparticles: How Canadian Discoveries Are Enabling Gene Therapies. Mol. Pharm. 2022, 19, 1663–1668. [Google Scholar] [CrossRef]
- Wilbie, D.; Walther, J.; Mastrobattista, E. Delivery Aspects of CRISPR/Cas for in Vivo Genome Editing. Acc. Chem. Res. 2019, 52, 1555–1564. [Google Scholar] [CrossRef] [Green Version]
- Shepherd, S.J.; Warzecha, C.C.; Yadavali, S.; El-Mayta, R.; Alameh, M.G.; Wang, L.; Weissman, D.; Wilson, J.M.; Issadore, D.; Mitchell, M.J. Scalable mRNA and siRNA Lipid Nanoparticle Production Using a Parallelized Microfluidic Device. Nano Lett. 2021, 21, 5671–5680. [Google Scholar] [CrossRef]
- Adams, D.; Gonzalez-Duarte, A.; O’Riordan, W.D.; Yang, C.-C.; Ueda, M.; Kristen, A.V.; Tournev, I.; Schmidt, H.H.; Coelho, T.; Berk, J.L.; et al. Patisiran, an RNAi Therapeutic, for Hereditary Transthyretin Amyloidosis. N. Engl. J. Med. 2018, 379, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Baden, L.R.; el Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.A.; Rouphael, N.; Creech, C.B.; et al. Efficacy and Safety of the mRNA-1273 SARS-CoV-2 Vaccine. N. Engl. J. Med. 2021, 384, 403–416. [Google Scholar] [CrossRef]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Marc, G.P.; Moreira, E.D.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 mRNA COVID-19 Vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef]
- Witzigmann, D.; Kulkarni, J.A.; Leung, J.; Chen, S.; Cullis, P.R.; van der Meel, R. Lipid nanoparticle technology for therapeutic gene regulation in the liver. Adv. Drug Deliv. Rev. 2020, 159, 344–363. [Google Scholar] [CrossRef]
- Francia, V.; Schiffelers, R.M.; Cullis, P.R.; Witzigmann, D. The Biomolecular Corona of Lipid Nanoparticles for Gene Therapy. Bioconjug. Chem. 2020, 31, 2046–2059. [Google Scholar] [CrossRef]
- Kulkarni, J.A.; Witzigmann, D.; Thomson, S.B.; Chen, S.; Leavitt, B.R.; Cullis, P.R.; van der Meel, R. The current landscape of nucleic acid therapeutics. Nat. Nanotechnol. 2021, 16, 630–643. [Google Scholar] [CrossRef]
- Lee, S.M.; Cheng, Q.; Yu, X.; Liu, S.; Johnson, L.T.; Siegwart, D.J. A Systematic Study of Unsaturation in Lipid Nanoparticles Leads to Improved mRNA Transfection In Vivo. Angew. Chem. Int. Ed. Engl. 2021, 60, 5848–5853. [Google Scholar] [CrossRef]
- Álvarez-Benedicto, E.; Farbiak, L.; Ramírez, M.M.; Wang, X.; Johnson, L.T.; Mian, O.; Guerrero, E.D.; Siegwart, D.J. Optimization of phospholipid chemistry for improved lipid nanoparticle (LNP) delivery of messenger RNA (mRNA). Biomater. Sci. 2022, 10, 549–559. [Google Scholar] [CrossRef] [PubMed]
- Du, Z.; Munye, M.M.; Tagalakis, A.D.; Manunta, M.D.I.; Hart, S.L. The role of the helper lipid on the DNA transfection efficiency of lipopolyplex formulations. Sci. Rep. 2014, 4, 7107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bollu, A.; Peters, A.; Rentmeister, A. Chemo-Enzymatic Modification of the 5′ Cap to Study mRNAs. Acc. Chem. Res. 2022, 55, 1249–1261. [Google Scholar] [CrossRef] [PubMed]
- van Dülmen, M.; Muthmann, N.; Rentmeister, A. Chemo-Enzymatic Modification of the 5′ Cap Maintains Translation and Increases Immunogenic Properties of mRNA. Angew. Chem. Int. Ed. Engl. 2021, 60, 13280–13286. [Google Scholar] [CrossRef]
- Chakraborty, C.; Sharma, A.R.; Bhattacharya, M.; Lee, S.S. From COVID-19 to Cancer mRNA Vaccines: Moving from Bench to Clinic in the Vaccine Landscape. Front. Immunol. 2021, 12, 679344. [Google Scholar] [CrossRef]
- Nakahama, T.; Kawahara, Y. Adenosine-to-inosine RNA editing in the immune system: Friend or foe? Cell Mol Life Sci. 2020, 77, 2931–2948. [Google Scholar] [CrossRef]
- Qin, S.; Tang, X.; Chen, Y.; Chen, K.; Fan, N.; Xiao, W.; Zheng, Q.; Li, G.; Teng, Y.; Wu, M.; et al. mRNA-based therapeutics: Powerful and versatile tools to combat diseases. Signal Transduct. Target Ther. 2022, 7, 166. [Google Scholar] [CrossRef]
- Melamed, J.R.; Hajj, K.A.; Chaudhary, N.; Strelkova, D.; Arral, M.L.; Pardi, N.; Alameh, M.G.; Miller, J.B.; Farbiak, L.; Siegwart, D.J.; et al. Lipid nanoparticle chemistry determines how nucleoside base modifications alter mRNA delivery. J. Control. Release 2022, 341, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Loughrey, D.; Dahlman, J.E. Non-liver mRNA Delivery. Acc. Chem. Res. 2022, 55, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Vaidyanathan, D.; Paskaleva, E.; Vargason, T.; Ke, X.; Mccallum, S.A.; Linhardt, R.J.; Dordick, J.S. Elucidating the unusual reaction kinetics of D-glucuronyl C5-epimerase. Glycobiology 2020, 30, 847–858. [Google Scholar] [CrossRef]
- Vargason, A.M.; Anselmo, A.C.; Mitragotri, S. The evolution of commercial drug delivery technologies. Nat. Biomed. Eng. 2021, 5, 951–967. [Google Scholar] [CrossRef] [PubMed]
- Wan, C.; Allen, T.M.; Cullis, P.R. Lipid nanoparticle delivery systems for siRNA-based therapeutics. Drug Deliv. Transl. Res. 2014, 4, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Chang, J.; Jiang, Y.; Meng, X.; Sun, T.; Mao, L.; Xu, Q.; Wang, M. Fast and Efficient CRISPR/Cas9 Genome Editing In Vivo Enabled by Bioreducible Lipid and Messenger RNA Nanoparticles. Adv Mater. 2019, 31, e1902575. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, J.A.; Witzigmann, D.; Chen, S.; Cullis, P.R.; van der Meel, R. Lipid Nanoparticle Technology for Clinical Translation of siRNA Therapeutics. Acc. Chem. Res. 2019, 52, 2435–2444. [Google Scholar] [CrossRef] [PubMed]
- Thess, A.; Grund, S.; Mui, B.L.; Hope, M.J.; Baumhof, P.; Fotin-Mleczek, M.; Schlake, T. Sequence-engineered mRNA Without Chemical Nucleoside Modifications Enables an Effective Protein Therapy in Large Animals. Mol. Ther. 2015, 23, 1456–1464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeRosa, F.; Guild, B.; Karve, S.; Smith, L.; Love, K.; Dorkin, J.R.; Kauffman, K.J.; Zhang, J.; Yahalom, B.; Anderson, D.G.; et al. Therapeutic efficacy in a hemophilia B model using a biosynthetic mRNA liver depot system. Gene Ther. 2016, 23, 699–707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miao, L.; Lin, J.; Huang, Y.; Li, L.; Delcassian, D.; Ge, Y.; Shi, Y.; Anderson, D.G. Synergistic lipid compositions for albumin receptor mediated delivery of mRNA to the liver. Nat. Commun. 2020, 11, 2424. [Google Scholar] [CrossRef]
- Raguram, A.; Banskota, S.; Liu, D.R. Therapeutic in vivo delivery of gene editing agents. Cell 2022, 185, 2806–2827. [Google Scholar] [CrossRef]
- Inui, M.; Miyado, M.; Igarashi, M.; Tamano, M.; Kubo, A.; Yamashita, S.; Asahara, H.; Fukami, M.; Takada, S. Rapid generation of mouse models with defined point mutations by the CRISPR/Cas9 system. Sci. Rep. 2014, 4, 5396. [Google Scholar] [CrossRef] [Green Version]
- Wei, T.; Cheng, Q.; Min, Y.L.; Olson, E.N.; Siegwart, D.J. Systemic nanoparticle delivery of CRISPR-Cas9 ribonucleoproteins for effective tissue specific genome editing. Nat. Commun. 2020, 11, 3232. [Google Scholar] [CrossRef]
- Yin, H.; Song, C.Q.; Suresh, S.; Wu, Q.; Walsh, S.; Rhym, L.H.; Mintzer, E.; Bolukbasi, M.F.; Zhu, L.J.; Kauffman, K.; et al. Structure-guided chemical modification of guide RNA enables potent non-viral in vivo genome editing. Nat. Biotechnol. 2017, 35, 1179–1187. [Google Scholar] [CrossRef] [PubMed]
- Finn, J.D.; Smith, A.R.; Patel, M.C.; Shaw, L.; Youniss, M.R.; van Heteren, J.; Dirstine, T.; Ciullo, C.; Lescarbeau, R.; Seitzer, J.; et al. A Single Administration of CRISPR/Cas9 Lipid Nanoparticles Achieves Robust and Persistent In Vivo Genome Editing. Cell Rep. 2018, 22, 2227–2235. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Yu, W.; Huang, S.; Wu, S.; Li, L.; Zhou, J.; Cao, Y.; Huang, X.; Qiao, Y. Structure-guided engineering of adenine base editor with minimized RNA off-targeting activity. Nat. Commun. 2021, 12, 2287. [Google Scholar] [CrossRef]
- Rothgangl, T.; Dennis, M.K.; Lin, P.J.C.; Oka, R.; Witzigmann, D.; Villiger, L.; Qi, W.; Hruzova, M.; Kissling, L.; Lenggenhager, D.; et al. In vivo adenine base editing of PCSK9 in macaques reduces LDL cholesterol levels. Nat. Biotechnol. 2021, 39, 949–957. [Google Scholar] [CrossRef]
- Musunuru, K.; Chadwick, A.C.; Mizoguchi, T.; Garcia, S.P.; DeNizio, J.E.; Reiss, C.W.; Wang, K.; Iyer, S.; Dutta, C.; Clendaniel, V.; et al. In vivo CRISPR base editing of PCSK9 durably lowers cholesterol in primates. Nature 2021, 593, 429–434. [Google Scholar] [CrossRef] [PubMed]
- Villiger, L.; Rothgangl, T.; Witzigmann, D.; Oka, R.; Lin, P.J.C.; Qi, W.; Janjuha, S.; Berk, C.; Ringnalda, F.; Beattie, M.B.; et al. In vivo cytidine base editing of hepatocytes without detectable off-target mutations in RNA and DNA. Nat. Biomed. Eng. 2021, 5, 179–189. [Google Scholar] [CrossRef]
- Jiang, T.; Henderson, J.M.; Coote, K.; Cheng, Y.; Valley, H.C.; Zhang, X.O.; Wang, Q.; Rhym, L.H.; Cao, Y.; Newby, G.A.; et al. Chemical modifications of adenine base editor mRNA and guide RNA expand its application scope. Nat. Commun. 2020, 11, 1979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grenkowitz, T.; Kassner, U.; Wühle-Demuth, M.; Salewsky, B.; Rosada, A.; Zemojtel, T.; Hopfenmüller, W.; Isermann, B.; Borucki, K.; Heigl, F.; et al. Clinical characterization and mutation spectrum of German patients with familial hypercholesterolemia. Atherosclerosis 2016, 253, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, T.S.; Lee, A.C.H.; Akinc, A.; Bramlage, B.; Bumcrot, D.; Fedoruk, M.N.; Harborth, J.; Heyes, J.A.; Jeffs, L.B.; John, M.; et al. RNAi-mediated gene silencing in non-human primates. Nature 2006, 441, 111–114. [Google Scholar] [CrossRef] [PubMed]
- Jayaraman, M.; Ansell, S.M.; Mui, B.L.; Tam, Y.K.; Chen, J.; Du, X.; Butler, D.; Eltepu, L.; Matsuda, S.; Narayanannair, J.K.; et al. Maximizing the potency of siRNA lipid nanoparticles for hepatic gene silencing in vivo. Angew. Chem. Int. Ed. Engl. 2012, 51, 8529–8533. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Yin, L.; Theisen, M.; Zhuo, J.; Siddiqui, S.; Levy, B.; Presnyak, V.; Frassetto, A.; Milton, J.; Salerno, T.; et al. Systemic mRNA Therapy for the Treatment of Fabry Disease: Preclinical Studies in Wild-Type Mice, Fabry Mouse Model, and Wild-Type Non-human Primates. Am. J. Hum. Genet. 2019, 104, 625–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeRosa, F.; Smith, L.; Shen, Y.; Huang, Y.; Pan, J.; Xie, H.; Yahalom, B.; Heartlein, M.W. Improved Efficacy in a Fabry Disease Model Using a Systemic mRNA Liver Depot System as Compared to Enzyme Replacement Therapy. Mol. Ther. 2019, 27, 878–889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pardi, N.; Secreto, A.J.; Shan, X.; Debonera, F.; Glover, J.; Yi, Y.; Muramatsu, H.; Ni, H.; Mui, B.L.; Tam, Y.K.; et al. Administration of nucleoside-modified mRNA encoding broadly neutralizing antibody protects humanized mice from HIV-1 challenge. Nat. Commun. 2017, 8, 14630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kenjo, E.; Hozumi, H.; Makita, Y.; Iwabuchi, K.A.; Fujimoto, N.; Matsumoto, S.; Kimura, M.; Amano, Y.; Ifuku, M.; Naoe, Y.; et al. Low immunogenicity of LNP allows repeated administrations of CRISPR-Cas9 mRNA into skeletal muscle in mice. Nat. Commun. 2021, 12, 7101. [Google Scholar] [CrossRef]
- Zou, Y.; Sun, X.; Yang, Q.; Zheng, M.; Shimoni, O.; Ruan, W.; Wang, Y.; Zhang, D.; Yin, J.; Huang, X.; et al. Blood-brain barrier-penetrating single CRISPR-Cas9 nanocapsules for effective and safe glioblastoma gene therapy. Sci. Adv. 2022, 8, eabm8011. [Google Scholar] [CrossRef] [PubMed]
- Han, J.P.; Kim, M.J.; Choi, B.S.; Lee, J.H.; Lee, G.S.; Jeong, M.; Lee, Y.; Kim, E.A.; Oh, H.K.; Go, N.; et al. In vivo delivery of CRISPR-Cas9 using lipid nanoparticles enables antithrombin gene editing for sustainable hemophilia A and B therapy. Sci. Adv. 2022, 8, eabj6901. [Google Scholar] [CrossRef]
- Herskovitz, J.; Hasan, M.; Patel, M.; Blomberg, W.R.; Cohen, J.D.; Machhi, J.; Shahjin, F.; Mosley, R.L.; McMillan, J.E.; Kevadiya, B.D.; et al. CRISPR-Cas9 Mediated Exonic Disruption for HIV-1 Elimination. eBioMedicine 2021, 73, 103678. [Google Scholar] [CrossRef]
- Qiu, M.; Glass, Z.; Chen, J.; Haas, M.; Jin, X.; Zhao, X.; Rui, X.; Ye, Z.; Li, Y.; Zhang, F.; et al. Lipid nanoparticle-mediated codelivery of Cas9 mRNA and single-guide RNA achieves liver-specific in vivo genome editing of Angptl3. Proc. Natl. Acad. Sci. USA 2021, 118, e2020401118. [Google Scholar] [CrossRef]
- Rosa, S.S.; Prazeres, D.M.F.; Azevedo, A.M.; Marques, M.P.C. mRNA vaccines manufacturing: Challenges and bottlenecks. Vaccine 2021, 39, 2190–2200. [Google Scholar] [CrossRef]
- Oberli, M.A.; Reichmuth, A.M.; Dorkin, J.R.; Mitchell, M.J.; Fenton, O.S.; Jaklenec, A.; Anderson, D.G.; Langer, R.; Blankschtein, D. Lipid Nanoparticle Assisted mRNA Delivery for Potent Cancer Immunotherapy. Nano Lett. 2017, 17, 1326–1335. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Ye, Z.; Huang, C.; Qiu, M.; Song, D.; Li, Y.; Xu, Q. Lipid nanoparticle-mediated lymph node-targeting delivery of mRNA cancer vaccine elicits robust CD8 + T cell response. Proc. Natl. Acad. Sci. USA 2022, 119, e2207841119. [Google Scholar] [CrossRef] [PubMed]
- Xiong, H.; Zhou, K.; Yan, Y.; Miller, J.B.; Siegwart, D.J. Tumor-Activated Water-Soluble Photosensitizers for Near-Infrared Photodynamic Cancer Therapy. ACS Appl. Mater. Interfaces 2018, 10, 16335–16343. [Google Scholar] [CrossRef] [PubMed]
- Xiong, H.; Kos, P.; Yan, Y.; Zhou, K.; Miller, J.B.; Elkassih, S.; Siegwart, D.J. Activatable Water-Soluble Probes Enhance Tumor Imaging by Responding to Dysregulated pH and Exhibiting High Tumor-to-Liver Fluorescence Emission Contrast. Bioconjug. Chem. 2016, 27, 1737–1744. [Google Scholar] [CrossRef]
- Xiong, H.; Liu, S.; Wei, T.; Cheng, Q.; Siegwart, D.J. Theranostic dendrimer-based lipid nanoparticles containing PEGylated BODIPY dyes for tumor imaging and systemic mRNA delivery in vivo. J. Control. Release 2020, 325, 198–205. [Google Scholar] [CrossRef]
- Solomon, S.D.; Adams, D.; Kristen, A.; Grogan, M.; González-Duarte, A.; Maurer, M.S.; Merlini, G.; Damy, T.; Slama, M.S.; Brannagan, T.H.; et al. Effects of Patisiran, an RNA Interference Therapeutic, on Cardiac Parameters in Patients with Hereditary Transthyretin-Mediated Amyloidosis. Circulation 2019, 139, 431–443. [Google Scholar] [CrossRef]
- Mashima, R.; Okuyama, T.; Ohira, M. Biomarkers for Lysosomal Storage Disorders with an Emphasis on Mass Spectrometry. Int. J. Mol. Sci. 2020, 21, 2704. [Google Scholar] [CrossRef] [Green Version]
- Mashima, R.; Okuyama, T.; Ohira, M. Physiology and Pathophysiology of Heparan Sulfate in Animal Models: Its Biosynthesis and Degradation. Int. J. Mol. Sci. 2022, 23, 1963. [Google Scholar] [CrossRef]
- Mashima, R.; Nakanishi, M. Mammalian Sulfatases: Biochemistry, Disease Manifestation, and Therapy. Int. J. Mol. Sci. 2022, 23, 8153. [Google Scholar] [CrossRef]
- Rodríguez-Castejón, J.; Alarcia-Lacalle, A.; Gómez-Aguado, I.; Vicente-Pascual, M.; Aspiazu, M.Á.S.; del Pozo-Rodríguez, A.; Rodríguez-Gascón, A. α-Galactosidase A Augmentation by Non-Viral Gene Therapy: Evaluation in Fabry Disease Mice. Pharmaceutics 2021, 13, 771. [Google Scholar] [CrossRef]
- Gillmore, J.D.; Gane, E.; Taubel, J.; Kao, J.; Fontana, M.; Maitland, M.L.; Seitzer, J.; O’Connell, D.; Walsh, K.R.; Wood, K.; et al. CRISPR-Cas9 In Vivo Gene Editing for Transthyretin Amyloidosis. N. Engl. J. Med. 2021, 385, 493–502. [Google Scholar] [CrossRef]
- Adams, D.; Suhr, O.B.; Dyck, P.J.; Litchy, W.J.; Leahy, R.G.; Chen, J.; Gollob, J.; Coelho, T. Trial design and rationale for APOLLO, a Phase 3, placebo-controlled study of patisiran in patients with hereditary ATTR amyloidosis with polyneuropathy. BMC Neurol. 2017, 17, 181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minamisawa, M.; Claggett, B.; Adams, D.; Kristen, A.v.; Merlini, G.; Slama, M.S.; Dispenzieri, A.; Shah, A.M.; Falk, R.H.; Karsten, V.; et al. Association of Patisiran, an RNA Interference Therapeutic, with Regional Left Ventricular Myocardial Strain in Hereditary Transthyretin Amyloidosis: The APOLLO Study. JAMA Cardiol. 2019, 4, 466–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obici, L.; Berk, J.L.; González-Duarte, A.; Coelho, T.; Gillmore, J.; Schmidt, H.H.J.; Schilling, M.; Yamashita, T.; Labeyrie, C.; Brannagan, T.H.; et al. Quality of life outcomes in APOLLO, the phase 3 trial of the RNAi therapeutic patisiran in patients with hereditary transthyretin-mediated amyloidosis. Amyloid 2020, 27, 153–162. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Goel, V.; Attarwala, H.; Sweetser, M.T.; Clausen, V.A.; Robbie, G.J. Patisiran Pharmacokinetics, Pharmacodynamics, and Exposure-Response Analyses in the Phase 3 APOLLO Trial in Patients with Hereditary Transthyretin-Mediated (hATTR) Amyloidosis. J. Clin. Pharmacol. 2020, 60, 37–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srikrishna, D.; Sachsenmeier, K. We need to bring R0. Genome Med. 2021, 13, 120. [Google Scholar] [CrossRef]
- Dilliard, S.A.; Cheng, Q.; Siegwart, D.J. On the mechanism of tissue-specific mRNA delivery by selective organ targeting nanoparticles. Proc. Natl. Acad. Sci. USA 2021, 118, e2109256118. [Google Scholar] [CrossRef]
- Cheng, Q.; Wei, T.; Farbiak, L.; Johnson, L.T.; Dilliard, S.A.; Siegwart, D.J. Selective organ targeting (SORT) nanoparticles for tissue-specific mRNA delivery and CRISPR-Cas gene editing. Nat. Nanotechnol. 2020, 15, 313–320. [Google Scholar] [CrossRef]
- Liu, S.; Cheng, Q.; Wei, T.; Yu, X.; Johnson, L.T.; Farbiak, L.; Siegwart, D.J. Membrane-destabilizing ionizable phospholipids for organ-selective mRNA delivery and CRISPR-Cas gene editing. Nat. Mater. 2021, 20, 701–710. [Google Scholar] [CrossRef]
- Sago, C.D.; Lokugamage, M.P.; Paunovska, K.; Vanover, D.A.; Monaco, C.M.; Shah, N.N.; Castro, M.G.; Anderson, S.E.; Rudoltz, T.G.; Lando, G.N.; et al. High-throughput in vivo screen of functional mRNA delivery identifies nanoparticles for endothelial cell gene editing. Proc. Natl. Acad. Sci. USA 2018, 115, E9944–E9952. [Google Scholar] [CrossRef]
- Yan, Y.; Liu, L.; Xiong, H.; Miller, J.B.; Zhou, K.; Kos, P.; Huffman, K.E.; Elkassih, S.; Norman, J.W.; Carstens, R.; et al. Functional polyesters enable selective siRNA delivery to lung cancer over matched normal cells. Proc. Natl. Acad. Sci. USA 2016, 113, E5702–E5710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, X.; Liu, S.; Cheng, Q.; Lee, S.M.; Wei, T.; Wang, X.; Siegwart, D.J.; Zhang, D.; Farbiak, L.; Johnson, L.T. Hydrophobic Optimization of Functional Poly(TPAE-co-suberoyl chloride) for Extrahepatic mRNA Delivery following Intravenous Administration. Pharmaceutics 2021, 13, 1914. [Google Scholar] [CrossRef] [PubMed]
- Rurik, J.G.; Tombácz, I.; Yadegari, A.; Fernández, P.O.M.; Shewale, S.V.; Li, L.; Kimura, T.; Soliman, O.Y.; Papp, T.E.; Tam, Y.K.; et al. CAR T cells produced in vivo to treat cardiac injury. Science 2022, 375, 91–96. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Chemical structure of lipids for LNP. (A) DLin-MC3-DMA, SM-102, and ALC-0315. (B) DOPE and DSPC. (C) Cholesterol. (D) PEG-DMG and ALC-0159.
Figure 1.
Chemical structure of lipids for LNP. (A) DLin-MC3-DMA, SM-102, and ALC-0315. (B) DOPE and DSPC. (C) Cholesterol. (D) PEG-DMG and ALC-0159.

Figure 2.
Chemical structure of ribonucleosides for LNP. (A) Structure of 5′-capping (m7Gppp-A/G). R = A/G. (B) Uridine and a series of its analog. (C) 2′-O-methylation and 3′/5′-O-phosphorothioation. R = A/G/U/C.
Figure 2.
Chemical structure of ribonucleosides for LNP. (A) Structure of 5′-capping (m7Gppp-A/G). R = A/G. (B) Uridine and a series of its analog. (C) 2′-O-methylation and 3′/5′-O-phosphorothioation. R = A/G/U/C.


{kind=link}
{kind=link}
Table 1.
Lipid composition of FDA-approved LNP.
| Product | Lipid Composition | Ref |
|---|---|---|
| Cationic ionizable lipid/helper phospholipid/cholesterol/PEG-lipid | ||
| Onpattro | DLin-MC3-DMA/DSPC/Chol/DMG-PEG2000 = 50/10/38.5/1.5 | [5] |
| mRNA-1273 | SM-102/DSPC/Chol/DMG-PEG2000 = 50/10/38.5/1.5 | [6] |
| BNT126b2 | ALC-0315/DSPC/Chol/ALC-0159 = 46.3/9.4/42.7/1.6 | [7] |
Table 2.
Comparison of preparation and modification procedure for mRNA and sgRNA.
| mRNA | sgRNA | |
|---|---|---|
| Preparation | In vitro transcription | Solid phase synthesis |
| Modification | ||
| 5′-Capping | Enzymatic | Not applicable |
| 2′-O-methylation | Enzymatic | Synthetic |
| 3′/5′-O-phosphorothioation | Not applicable | Synthetic |
| Replacement by uridine analog | Enzymatic | Not applicable |
| Codon optimization | In silico | Not applicable |
Table 3.
5′-Capping of mRNA [14].
Table 3.
5′-Capping of mRNA [14].
| Nomenclature | 5′-5′ Bridge | Base | Enzyme | |||
|---|---|---|---|---|---|---|
| Non-Transcribed Nucleoside | 1st Transcribed Nucleoside | 2nd Transcribed Nucleoside | ||||
| Guanine | Ribose 2′- | Adenine | Ribose 2′- | |||
| Uncapped | GpppA/G | No modification | OH | H | OH | |
| Cap 0 | GpppA/G | N7-Me | OH | H | OH | TGS |
| Cap 1 | GpppA/G | N7-Me | Me | H | OH | CMTR1 |
| Cap 2 | GpppA/G | N7-Me | Me | H | Me | CMTR2 |
| GpppG | N7-Me; N2-Me; N2,2′-di-Me | OH | H | OH | TGS | |
| GpppA | N7-Me | OH | N2-Me | OH | SAMT | |
| GpppA | N7-Me | Me | N6-Me | Me | CAPAM | |
TGS, 2,2,7-trimethylguanosine synthase; CAMT, cap-specific mRNA (nucleoside-2′-O-)-methyltransferase; CAPAM, cap-specific adenosine N6-methyltransferase; SAMT, S-adenosylmethionine transferase.
Table 4.
LNP-based preclinical studies.
| Disorders | Target | Genetic Material | Ref |
|---|---|---|---|
| Bleeding disorder | FVII | siRNA | [41] |
| Hypercholesterolemia | Pcsk9 | siRNA | [23] |
| Hypercholesterolemia | ApoB | siRNA | [40] |
| Transthyretin-mediated amyloidosis | Ttr | siRNA | [24] |
| Transthyretin-mediated amyloidosis | Ttr | siRNA | [23] |
| Anemia | mEPO, pEPO | Therapeutic mRNA | [26] |
| Anemia | hEPO | Therapeutic mRNA | [27] |
| Anemia | hEPO | Therapeutic mRNA | [28] |
| Fabry disease | hGLA | Therapeutic mRNA | [42] |
| Fabry disease | hGLA | Therapeutic mRNA | [43] |
| HIV | anti-HIV-1 antibody | Therapeutic mRNA | [44] |
| DMD | Human DMD exon 45 | Cas9 | [45] |
| Glioblastoma | Plk1 | Cas9 | [46] |
| Hemophilia A and B | Serpinc1 (encoding antithrombin) | Cas9 | [47] |
| HIV | TatDE | Cas9 | [48] |
| Hypercholesterolemia | Angptl3 | Cas9 | [49] |
| Hypercholesterolemia | Pcsk9 | Cas9 | [24] |
| Hypercholesterolemia | Pcsk9 | Cas9 | [31] |
| Hypercholesterolemia | Pcsk9 | Cas9 | [32] |
| Transthyretin-mediated amyloidosis | Ttr | Cas9 | [33] |
| Hypercholesterolemia | Pcsk9 | ABE | [36] |
| Hypercholesterolemia | Pcsk9 | ABE | [35] |
| Phenylketonuria | Pah | CBE | [37] |
| Tyrosinemia I | Fah | ABE | [38] |
Table 5.
Clinical trials of LNP-mediated gene therapy.
| NCT Number | Conditions-1 | Conditions-2 | Target | Payload | Phases | Status | Sponsor/Collaborators | Ref |
|---|---|---|---|---|---|---|---|---|
| NCT04601051 | Amyloidosis | hATTR | Transthyretin | Cas9 mRNA | Phase 1 | Recruiting | Intellia Therapeutics | [61] |
| NCT01960348 | Amyloidosis | hATTR | Transthyretin | siRNA | Phase 3 | Completed | Alnylam Pharmaceuticals | [5,56,62,63,64,65] |
| NCT01437007 | Cancer | Hepatic Metastases | PLK1 | siRNA | Phase 1 | Completed | NCI; NIH Clinical Center | |
| NCT03323398 | Cancer | Solid Tumor | Human OX40L | mRNA | Phase 1/2 | Terminated | ModernaTX, Inc. | |
| NCT03739931 | Cancer | Solid Tumor | Human OX40L, IL-23, and IL-36γ | mRNA | Phase 1 | Recruiting | ModernaTX, Inc.; AstraZeneca | [66] |
| NCT04675996 | Cancer | Solid Tumor | miR-193a-3p | miRNA | Phase 1 | Recruiting | InteRNA | |
| NCT02110563 | Cancer | Solid Tumor | MYC | siRNA | Phase 1 | Terminated | Dicerna Pharmaceuticals, Inc. | |
| NCT02314052 | Cancer | Hepatocellular Carcinoma | MYC | siRNA | Phase 1/2 | Terminated | Dicerna Pharmaceuticals, Inc. | |
| NCT04486833 | Cancer | Carcinoma, Non-Small Cell Lung | TUSC2 | DNA plasmid | Phase 1/2 | Recruiting | Genprex, Inc. | |
| NCT05062980 | Cancer | Non-Small Cell Lung Cancer | TUSC2 | DNA plasmid | Phase 1/2 | Recruiting | Genprex, Inc. | |
| NCT05497453 | Cancer | Solid Tumor | 2 independent epigenomic controllers | biscistronic mRNA | Phase 1/2 | Recruiting | Omega Therapeutics | |
| NCT02227459 | Hepatic Fibrosis | HSP47 | siRNA | Phase 1 | Completed | Bristol-Myers Squibb; Nitto Denko Corporation | ||
| NCT04416126 | OTCD | Ornithine transcarbamylase | Therapeutic mRNA | Phase 1 | Completed | Arcturus Therapeutics, Inc. | ||
| NCT04442347 | OTCD | Ornithine transcarbamylase | Therapeutic mRNA | Phase 1 | Recruiting | Arcturus Therapeutics, Inc. | ||
| NCT05526066 | OTCD | Ornithine transcarbamylase | Therapeutic mRNA | Phase 2 | Recruiting | Arcturus Therapeutics, Inc. |
OTCD, Ornithine transcarbamylase deficiency; hATTR, hereditary transthyretin amyloidosis.
Table 6.
Tissue targeting of LNP.
| Target Organ | Basic Component | Additive | Ref |
|---|---|---|---|
| Cationic ionizable lipid/helper phospholipid/cholesterol/PEG-lipid | (SORT molecule) | ||
| Conventional | LP01/DSPC/Chol/PEG2000-DMG = 45/9/44/2 | Not added | [33] |
| DLin-MC3-DMA/DSPC/Chol/DMG-PEG2000 = 50/10/38.5/1.5 | Not added | [69] | |
| 5A2-SC8/DOPE/Chol/C14PEG2000 = 23.8/23.8/47.8/4.8 | Not added | [67] | |
| Liver | 5A2-SC8/DOPE/Chol/C14PEG2000 = 19/19/38.4/4 | DODAP = 20 | [67,68] |
| 9A1P9/5A2-SC8/Chol/DMG-PEG2000 = 25/30/30/1 | Not added | [69] | |
| Spleen | 5A2-SC8/DOPE/Chol/C14PEG2000 = 16.7/16.7/33.3/3.3 | 18:1 PA = 30 | [67,68] |
| Lung | 5A2-SC8/DOPE/Chol/C14PEG2000 = 11.9/11.9/23.8/2.4 | DOTAP = 50 | [67,68] |
| 9A1P9/DDAB/Chol/DMG-PEG2000 = 60/30/40/0.4 | Not added | [69] | |
| 5A2-SC8/DOPE/Chol/DMG-PEG = 15/15/30/3 | DOTAP = 63 | [31] |
18:1 PA, 1,2-dioleoyl-sn-glycero-3-phosphate; DODAP, 1,2-dioleoyl-3-dimethylammonium propane; DOTAP, 1,2-dioleoyl-3-trimethylammonium-propane; SORT, selective organ targeting.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Mashima, R.; Takada, S. Lipid Nanoparticles: A Novel Gene Delivery Technique for Clinical Application. Curr. Issues Mol. Biol. 2022, 44, 5013-5027. https://doi.org/10.3390/cimb44100341
AMA Style
Mashima R, Takada S. Lipid Nanoparticles: A Novel Gene Delivery Technique for Clinical Application. Current Issues in Molecular Biology. 2022; 44(10):5013-5027. https://doi.org/10.3390/cimb44100341
Chicago/Turabian StyleMashima, Ryuichi, and Shuji Takada. 2022. "Lipid Nanoparticles: A Novel Gene Delivery Technique for Clinical Application" Current Issues in Molecular Biology 44, no. 10: 5013-5027. https://doi.org/10.3390/cimb44100341