
Classifying Integrated Signature Molecules in Macrophages of Rheumatoid Arthritis, Osteoarthritis, and Periodontal Disease: An Omics-Based Study

 , ,
, ,  ,
,
Abstract
:1. Introduction
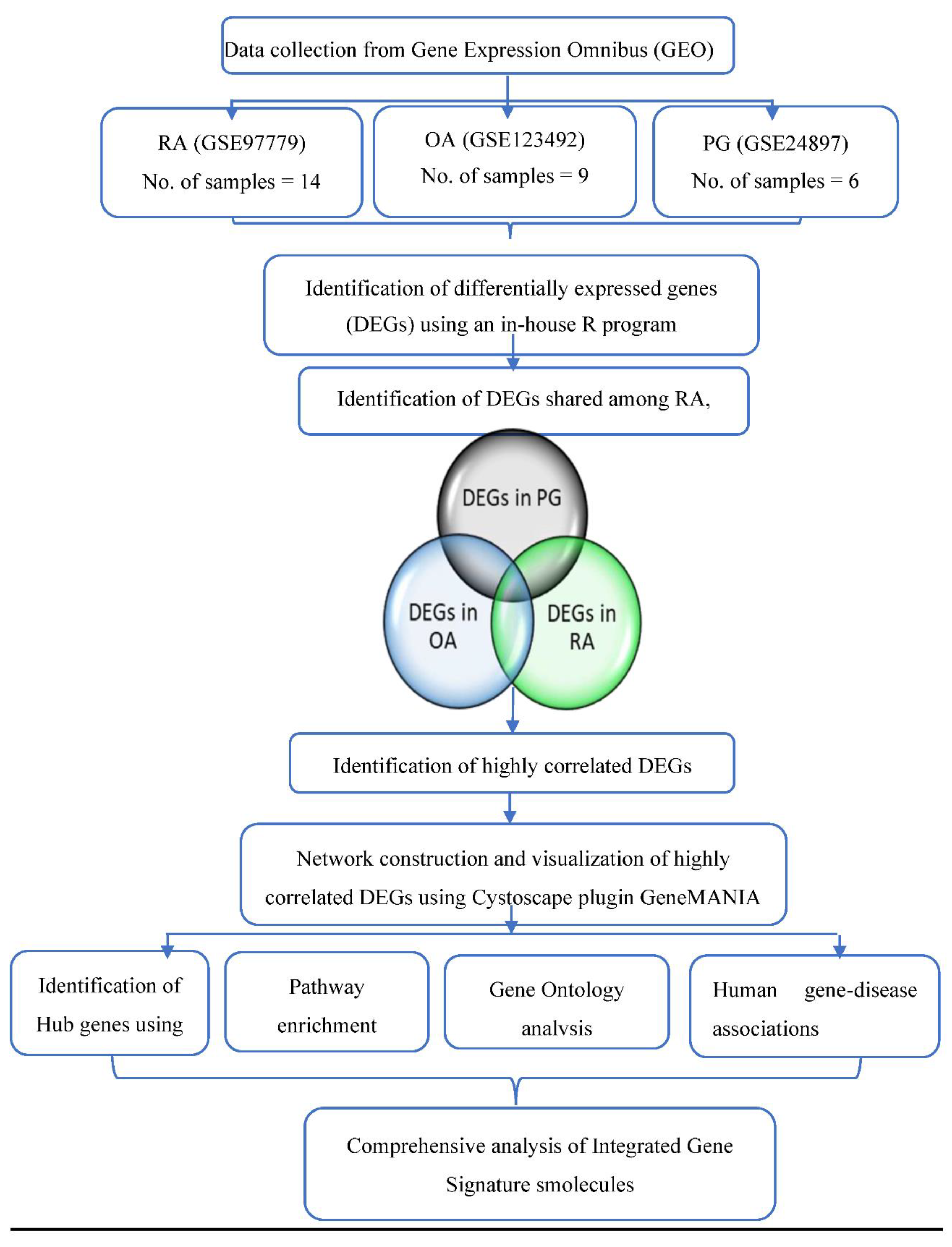
2. Materials and Methods
2.1. Retrieval of Gene Expression Data
2.2. Identification of Differentially Expressed Genes (DEGs)
2.3. Gene–Gene Correlation Network Construction
2.4. Network Construction and Hub Gene Identification
2.4.1. Network Construction
2.4.2. Hub Genes Identification
2.5. Gene Enrichment Analysis
3. Results
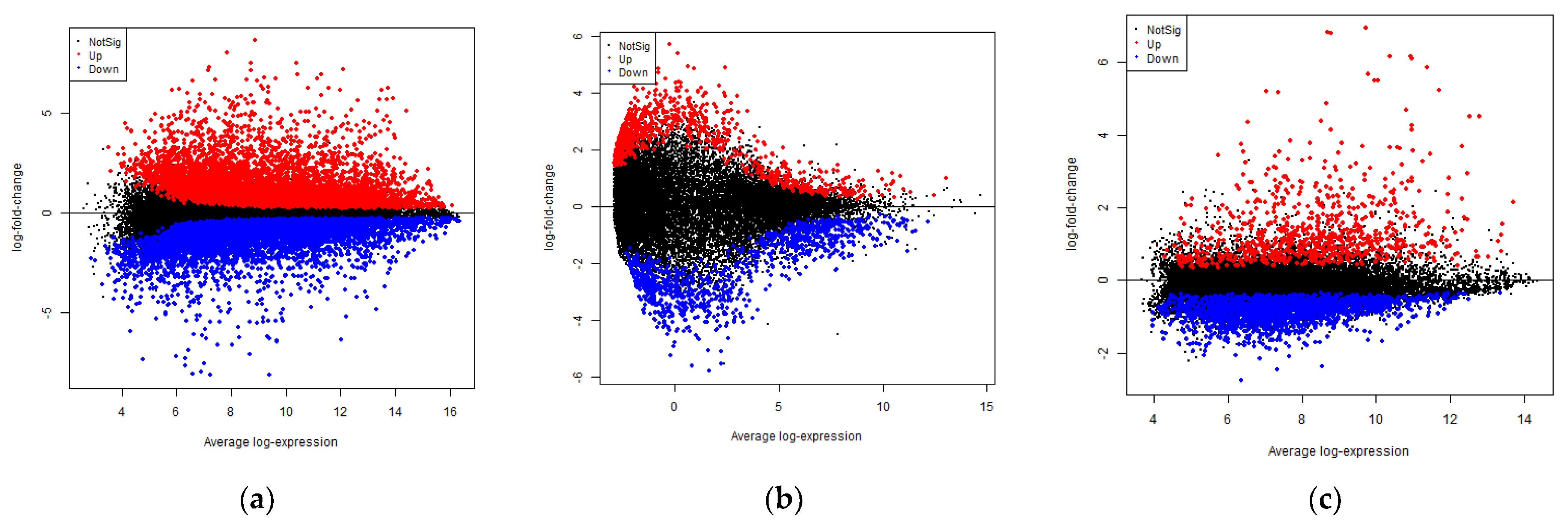
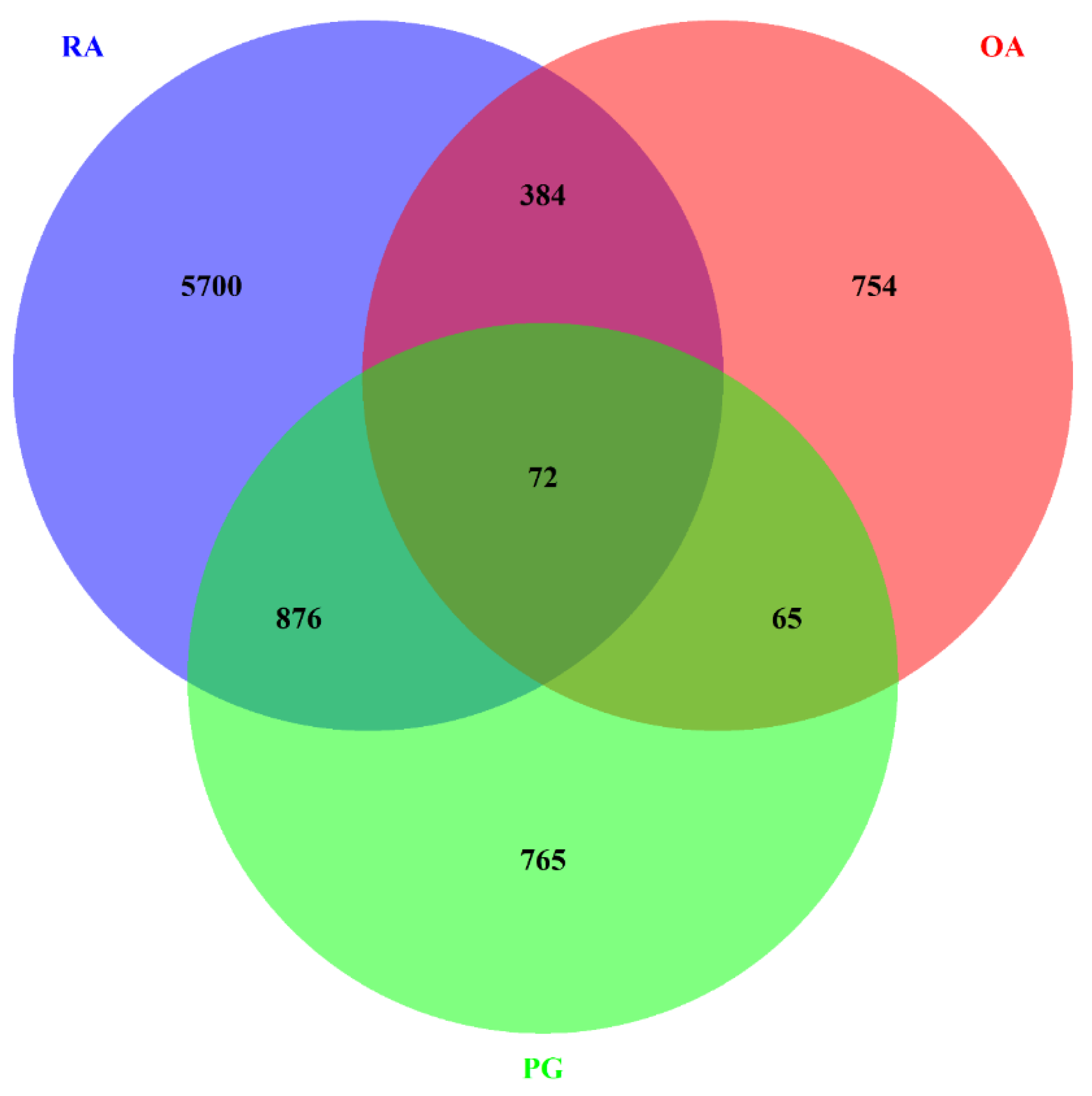
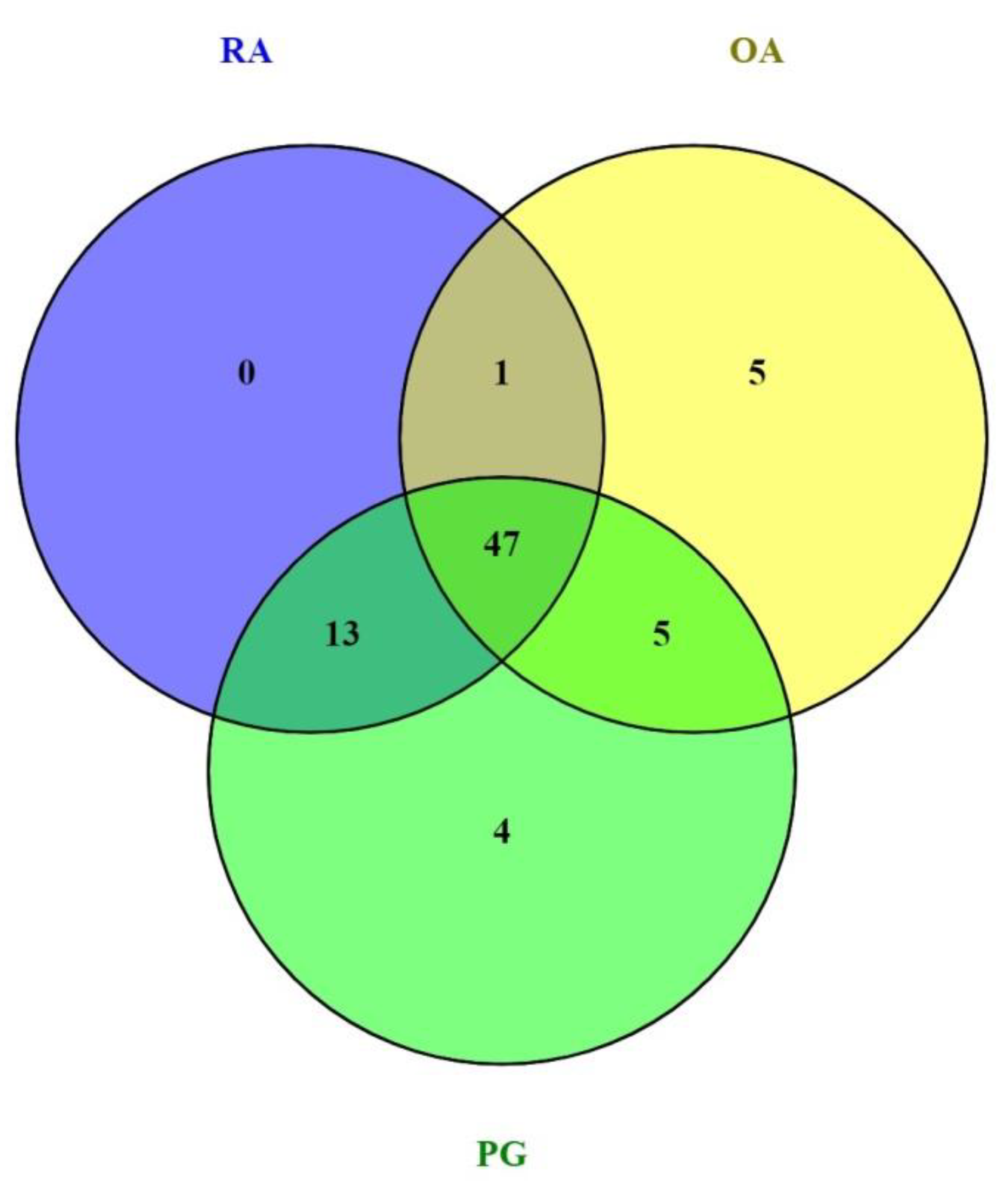
3.1. Identification of DEGs
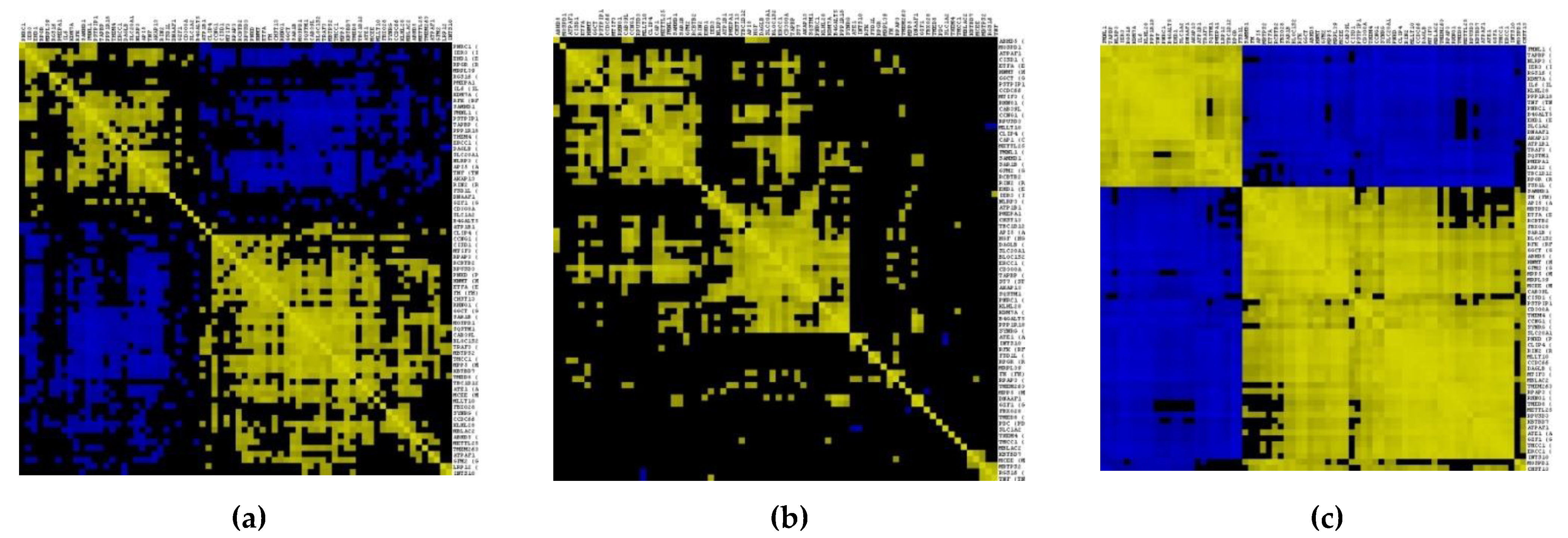
3.2. Gene–Gene Correlation Network Construction
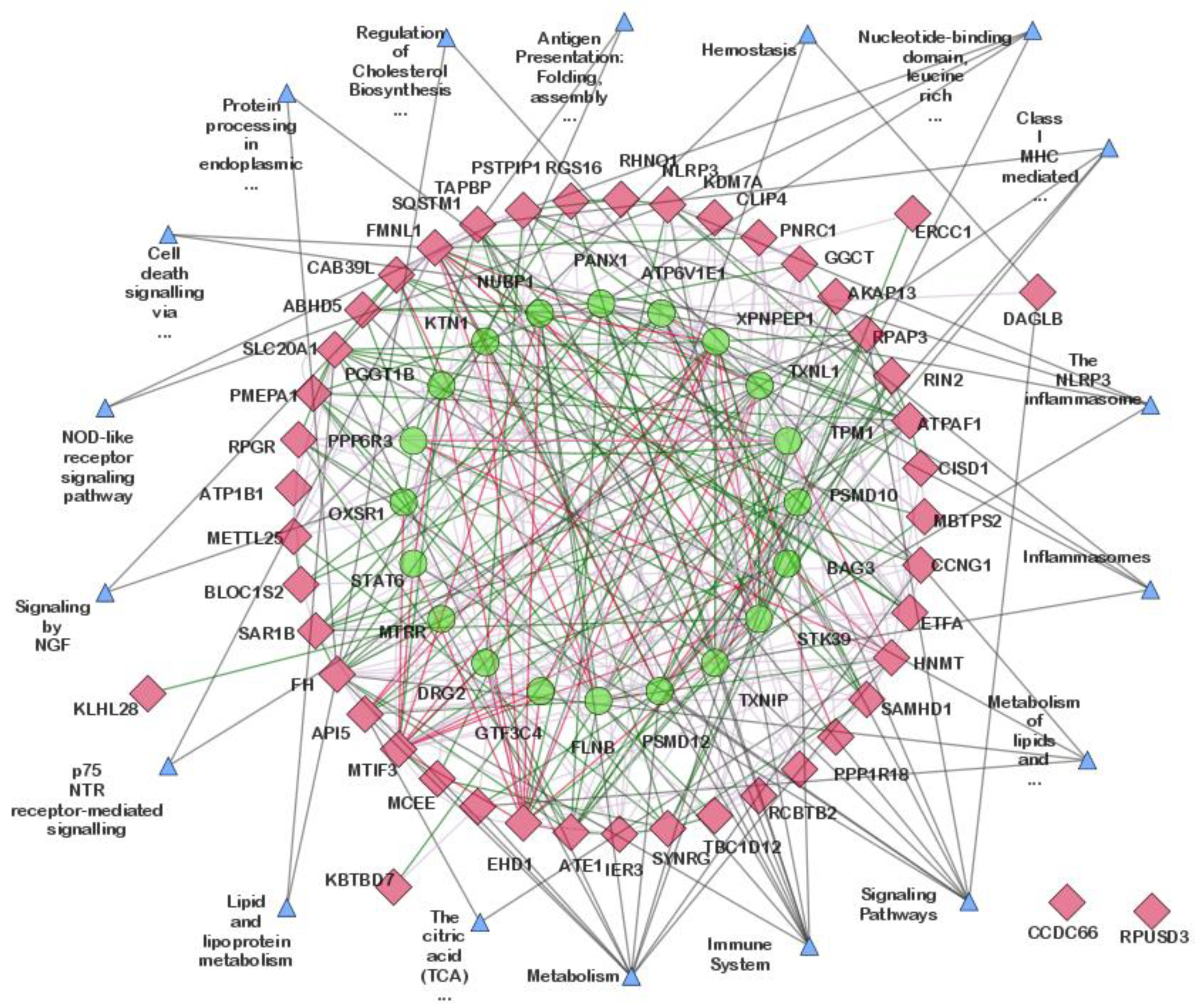
3.3. Network Construction and Hub Genes Identification
3.3.1. Network Construction
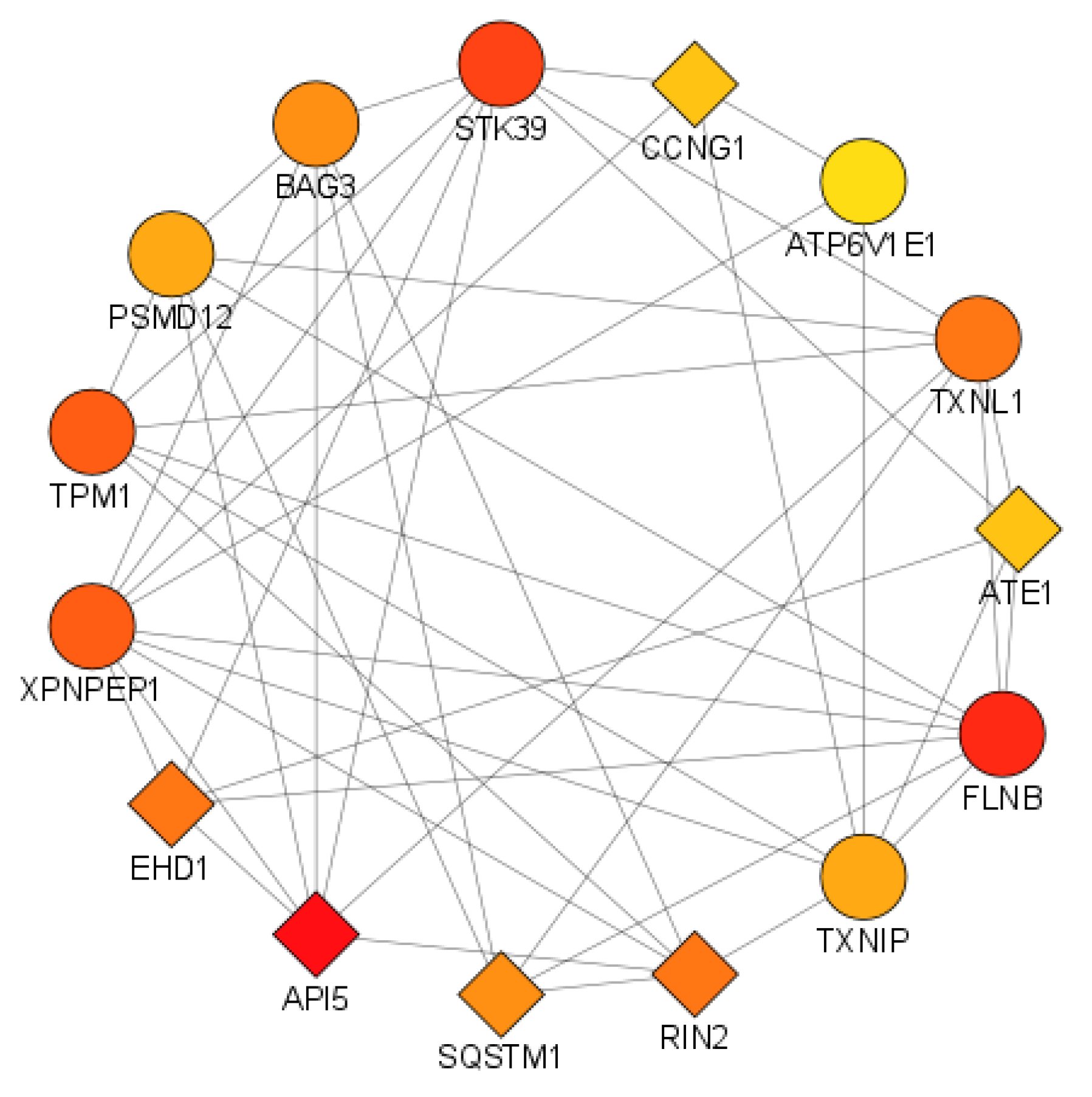
3.3.2. Identification of Hub Genes
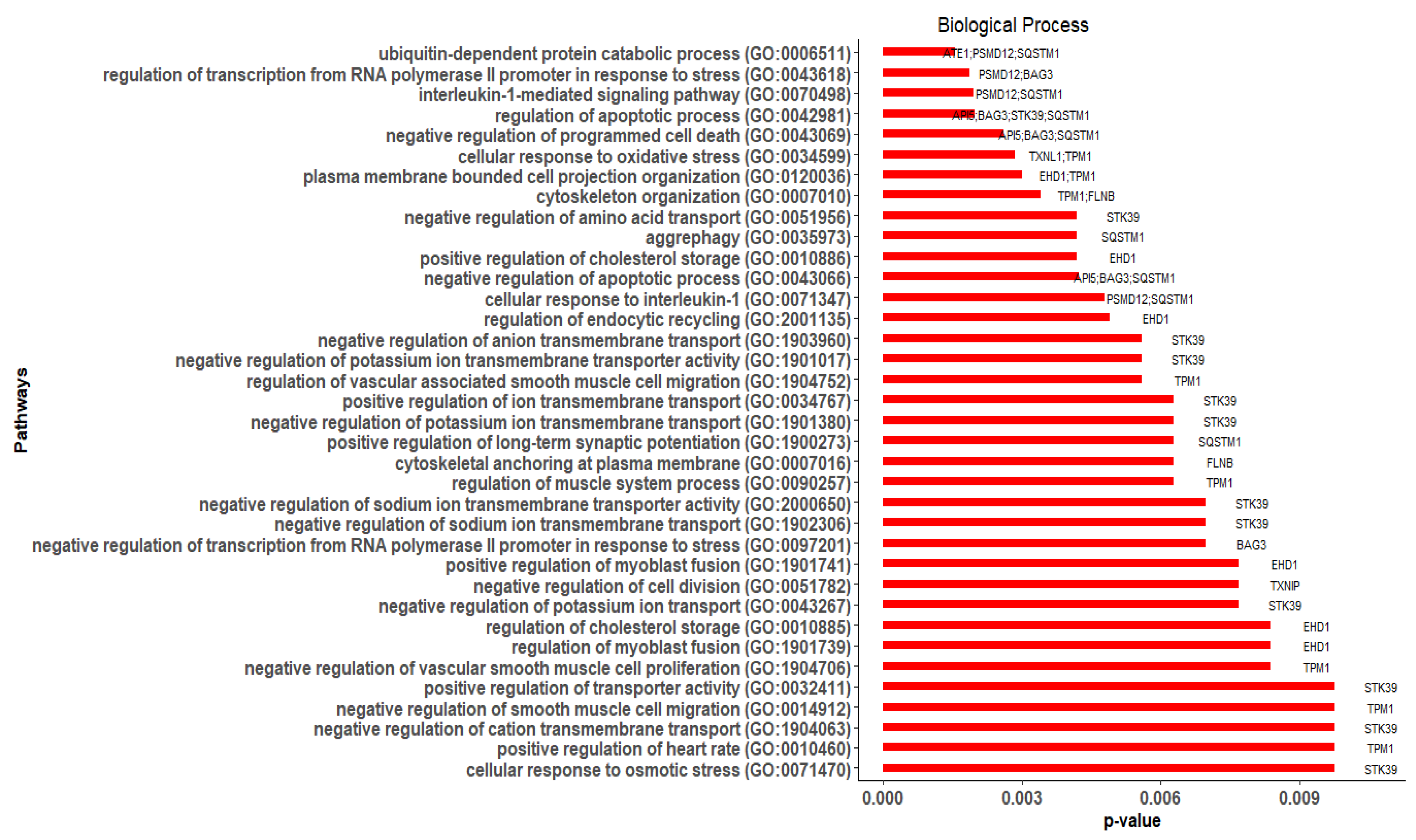
3.4. Gene Ontology (GO) and Pathway Enrichment Analysis of Hub Genes
4. Discussion
5. Conclusion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Deodhar, A.A.; Woolf, A.D. Bone mass measurement and bone metabolism in rheumatoid arthritis: A review. Br. J. Rheumatol. 1996, 35, 309–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Firestein, G.S. Evolving concepts of rheumatoid arthritis. Nature 2003, 423, 356–361. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Zhao, Y.; Li, S.; Yang, L.; Wang, H.; Wang, T.; Bin, S.; Gai, Z.; Heng, X.; Zhang, C.; et al. Variations in oral microbiome profiles in rheumatoid arthritis and osteoarthritis with potential biomarkers for arthritis screening. Sci. Rep. 2018, 8, 17126. [Google Scholar] [CrossRef] [PubMed]
- Favazzo, L.J.; Hendesi, H.; Villani, D.A.; Soniwala, S.; Dar, Q.-A.; Schott, E.M.; Gill, S.R.; Zuscik, M.J. The gut microbiome-joint connection: Implications in osteoarthritis. Curr. Opin. Rheumatol. 2020, 32, 92–101. [Google Scholar] [CrossRef]
- Kim, H.-S.; Park, H.-M.; Kim, H.; Lee, H.S.; Son, D.-H.; Lee, Y.-J. Association Between the Severity of Periodontitis and Osteoarthritis in Middle-Aged and Older Patients With Type 2 Diabetes Mellitus: A Nationwide Population-Based Study. Arthritis Care Res. 2022, 74, 403–409. [Google Scholar] [CrossRef]
- Wood, M.J.; Leckenby, A.; Reynolds, G.; Spiering, R.; Pratt, A.G.; Rankin, K.S.; Isaacs, J.D.; Haniffa, M.A.; Milling, S.; Hilkens, C.M. Macrophage proliferation distinguishes 2 subgroups of knee osteoarthritis patients. JCI Insight 2019, 4, e125325. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.Y.; Stabler, T.; Pei, F.X.; Kraus, V.B. Both systemic and local lipopolysaccharide (LPS) burden are associated with knee OA severity and inflammation. Osteoarthr. Cartil. 2016, 24, 1769–1775. [Google Scholar] [CrossRef] [Green Version]
- Zuo, B.; Zhu, J.; Xiao, F.; Wang, C.; Shen, Y.; Chen, X. Identification of novel biomarkers and candidate small molecule drugs in rheumatoid arthritis and osteoarthritis based on bioinformatics analysis of high-throughput data. Biosci. Rep. 2020, 40, 12. [Google Scholar] [CrossRef]
- Suzuki, A.; Horie, T.; Numabe, Y. Investigation of molecular biomarker candidates for diagnosis and prognosis of chronic periodontitis by bioinformatics analysis of pooled microarray gene expression datasets in Gene Expression Omnibus (GEO). BMC Oral Health 2019, 19, 52. [Google Scholar] [CrossRef] [Green Version]
- Sao, P.; Vats, S.; Singh, S. Porphyromonas gingivalis resistance and virulence: An integrated functional network analysis. Gene 2022, 839, 146734. [Google Scholar] [CrossRef]
- Sao, P.; Chand, Y.; Kumar, A.; Singh, S. Potential Drug Target Identification in Porphyromonas gingivalis using In-silico Subtractive Metabolic Pathway Analysis. Bangladesh J. Med. Sci. 2021, 20, 887–896. [Google Scholar] [CrossRef]
- Beck, J.D.; Papapanou, P.N.; Philips, K.H.; Offenbacher, S. Periodontal Medicine: 100 Years of Progress. J. Dent. Res. 2019, 98, 1053–1062. [Google Scholar] [CrossRef] [PubMed]
- Friedewald, V.E.; Kornman, K.S.; Beck, J.D.; Genco, R.; Goldfine, A.; Libby, P.; Offenbacher, S.; Ridker, P.M.; Dyke, T.E.V.; Roberts, W.C. The American Journal of Cardiology and Journal of Periodontology Editors’ Consensus: Periodontitis and Atherosclerotic Cardiovascular Disease. Am. J. Cardiol. 2009, 104, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Lamster, I.B.; Lalla, E.; Borgnakke, W.S.; Taylor, G.W. The Relationship Between Oral Health and Diabetes Mellitus. J. Am. Dent. Assoc. 2008, 139, 19S–24S. [Google Scholar] [CrossRef] [Green Version]
- Lundmark, A.; Davanian, H.; Båge, T.; Johannsen, G.; Koro, C.; Lundeberg, J.; Yucel-Lindberg, T. Transcriptome analysis reveals mucin 4 to be highly associated with periodontitis and identifies pleckstrin as a link to systemic diseases. Sci. Rep. 2015, 5, 18475. [Google Scholar] [CrossRef]
- Perricone, C.; Ceccarelli, F.; Matteo, S.; Di Carlo, G.; Bogdanos, D.P.; Lucchetti, R.; Pilloni, A.; Valesini, G.; Polimeni, A.; Conti, F. Porphyromonas gingivalis and rheumatoid arthritis. Curr. Opin. Rheumatol. 2019, 31, 517–524. [Google Scholar] [CrossRef]
- Totaro, M.C.; Cattani, P.; Ria, F.; Tolusso, B.; Gremese, E.; Fedele, A.L.; D’Onghia, S.; Marchetti, S.; Sante, G.D.; Canestri, S.; et al. Porphyromonas gingivalis and the pathogenesis of rheumatoid arthritis: Analysis of various compartments including the synovial tissue. Arthritis Res. Ther. 2013, 15, R66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Röhner, E.; Detert, J.; Kolar, P.; Hocke, A.; N’Guessan, P.; Matziolis, G.; Kanitz, V.; Bernimoulin, J.P.; Kielbassa, A.; Burmester, G.R.; et al. Induced apoptosis of chondrocytes by Porphyromonas gingivalis as a possible pathway for cartilage loss in rheumatoid arthritis. Calcif. Tissue Int. 2010, 87, 333–340. [Google Scholar] [CrossRef]
- Blasi, I.; Korostoff, J.; Dhingra, A.; Reyes-Reveles, J.; Shenker, B.J.; Shahabuddin, N.; Alexander, D.; Lally, E.T.; Bragin, A.; Boesze-Battaglia, K. Variants of Porphyromonas gingivalis lipopolysaccharide alter lipidation of autophagic protein, microtubule-associated protein 1 light chain 3, LC3. Mol. Oral Microbiol. 2016, 31, 486–500. [Google Scholar] [CrossRef]
- Yu, W.-H.; Hu, H.; Zhou, Q.; Xia, Y.; Amar, S. Bioinformatics Analysis of Macrophages Exposed to Porphyromonas gingivalis: Implications in Acute vs. Chronic Infections. PLoS ONE 2010, 5, e15613. [Google Scholar] [CrossRef] [Green Version]
- Weinberg, J.B.; Wortham, T.S.; Misukonis, M.A.; Patton, K.L.; Chitneni, S.R. Synovial mononuclear phagocytes in rheumatoid arthritis and osteoarthritis: Quantitative and functional aspects. Immunol. Investig. 1993, 22, 365–374. [Google Scholar] [CrossRef]
- Kennedy, A.; Fearon, U.; Veale, D.J.; Godson, C. Macrophages in Synovial Inflammation. Front. Immunol. 2011, 2, 2011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raggi, F.; Pelassa, S.; Pierobon, D.; Penco, F.; Gattorno, M.; Novelli, F.; Eva, A.; Varesio, L.; Giovarelli, M.; Bosco, M.C. Regulation of Human Macrophage M1–M2 Polarization Balance by Hypoxia and the Triggering Receptor Expressed on Myeloid Cells-1. Front. Immunol. 2017, 8, 1097. [Google Scholar] [CrossRef] [PubMed]
- Werheim, E.R.; Senior, K.G.; Shaffer, C.A.; Cuadra, G.A. Oral Pathogen Porphyromonas gingivalis Can Escape Phagocytosis of Mammalian Macrophages. Microorganisms 2020, 8, 1432. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulos, G.; Shaik-Dasthagirisaheb, Y.B.; Huang, N.; Viglianti, G.A.; Henderson, A.J.; Kantarci, A.; Gibson, F.C. Immunologic Environment Influences Macrophage Response to Porphyromonas gingivalis. Mol. Oral Microbiol. 2017, 32, 250–261. [Google Scholar] [CrossRef]
- Kurowska-Stolarska, M.; Alivernini, S. Synovial tissue macrophages: Friend or foe? RMD Open 2017, 3, e000527. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, A.; Fahy, N.; Hamilton, S.; Eckman, B.; Almeida, L.D.; Ishihara, S.; Mayr, M.G.; Chen, S.Y.; Gadhvi, G.; Cuda, C.; et al. Macrophages drive the inflammatory phase in experimental osteoarthritis. bioRxiv 2020. [Google Scholar] [CrossRef]
- Zhao, Y.; Chen, B.; Li, S.; Yang, L.; Zhu, D.; Wang, Y.; Wang, H.; Wang, T.; Shi, B.; Gai, Z.; et al. Detection and characterization of bacterial nucleic acids in culture-negative synovial tissue and fluid samples from rheumatoid arthritis or osteoarthritis patients. Sci. Rep. 2018, 8, 14305. [Google Scholar] [CrossRef] [PubMed]
- Kassem, A.; Henning, P.; Lundberg, P.; Souza, P.P.C.; Lindholm, C.; Lerner, U.H. Porphyromonas gingivalis Stimulates Bone Resorption by Enhancing RANKL (Receptor Activator of NF-κB Ligand) through Activation of Toll-like Receptor 2 in Osteoblasts. J. Biol. Chem. 2015, 290, 20147–20158. [Google Scholar] [CrossRef] [Green Version]
- Marchesan, J.T.; Gerow, E.A.; Schaff, R.; Taut, A.D.; Shin, S.-Y.; Sugai, J.; Brand, D.; Burberry, A.; Jorns, J.; Lundy, S.K.; et al. Porphyromonas gingivalis oral infection exacerbates the development and severity of collagen-induced arthritis. Arthritis Res. Ther. 2013, 15, R186. [Google Scholar] [CrossRef] [Green Version]
- Snijesh, V.P.; Matchado, M.S.; Singh, S. Classifying Rheumatoid Arthritis gene network signatures for identifying key regulatory molecules and their altered pathways by adopting network biology approach. Gene Rep. 2018, 13, 199–211. [Google Scholar] [CrossRef]
- Baliban, R.C.; Sakellari, D.; Li, Z.; DiMaggio, P.A.; Garcia, B.A.; Floudas, C.A. Novel protein identification methods for biomarker discovery via a proteomic analysis of periodontally healthy and diseased gingival crevicular fluid samples. J. Clin. Periodontol. 2012, 39, 203–212. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Vennila, J.J.; Snijesh, V.P.; George, G.; Sunny, C. Implying Analytic Measures for Unravelling Rheumatoid Arthritis Significant Proteins Through Drug–Target Interaction. Interdiscip. Sci. Comput. Life Sci. 2016, 8, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Barrett, T.; Edgar, R. Mining microarray data at NCBI’s Gene Expression Omnibus (GEO)*. Methods Mol. Biol. Clifton NJ 2006, 338, 175–190. [Google Scholar] [CrossRef] [Green Version]
- Chand, Y.; Sao, P.; Singh, S.; Chandra, N.; Das, S.; Singh, S. Prioritizing potential diagnostic biomarkers of Alzheimer’s disease by investigating gene expression data: A network-based approach. Alzheimers Dement. 2020, 16, e044322. [Google Scholar] [CrossRef]
- Sao, P.; Kishore, I.; Singh, S.; Panneerselvam, K.; Kumar, A.; Chaudhuri, T. Putative Target Identification for Gout; A Network Biology Approach. J. Bionanosci. 2013, 7, 649–653. [Google Scholar] [CrossRef]
- Singh, S. Network Biology Approach for Identifying Significant Drug Targets and Pathways for Rheumatoid Arthritis; Karunya University: Coimbatore, India, 2015; 162p, Available online: http://hdl.handle.net/10603/106794 (accessed on 25 July 2021).
- Taye, B.; Vaz, C.; Tanavde, V.; Kuznetsov, V.A.; Eisenhaber, F.; Sugrue, R.J.; Maurer-Stroh, S. Benchmarking selected computational gene network growing tools in context of virus-host interactions. Sci. Rep. 2017, 7, 5805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, K.; Park, S.H.; Chen, J.; Qiao, Y.; Giannopoulou, E.; Berg, K.; Hanidu, A.; Li, J.; Nabozny, G.; Kang, K.; et al. Interferon-γ Represses M2 Gene Expression in Human Macrophages by Disassembling Enhancers Bound by the Transcription Factor MAF. Immunity 2017, 47, 235–250.e4. [Google Scholar] [CrossRef]
- Davis, S.; Meltzer, P.S. GEOquery: A bridge between the Gene Expression Omnibus (GEO) and BioConductor. Bioinforma. Oxf. Engl. 2007, 23, 1846–1847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. Limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Smyth, G.K. Linear Models and Empirical Bayes Methods for Assessing Differential Expression in Microarray Experiments. Stat. Appl. Genet. Mol. Biol. 2004, 3. [Google Scholar] [CrossRef]
- George, G.; Singh, S.; Lokappa, S.B.; Varkey, J. Gene co-expression network analysis for identifying genetic markers in Parkinson’s disease-a three-way comparative approach. Genomics 2019, 111, 819–830. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Montojo, J.; Zuberi, K.; Rodriguez, H.; Bader, G.D.; Morris, Q. GeneMANIA: Fast gene network construction and function prediction for Cytoscape. F1000Research 2014, 3, 153. [Google Scholar] [CrossRef] [PubMed]
- Blessia, T.F.; Singh, S.; Vennila, J.J. Unwinding the Novel Genes Involved in the Differentiation of Embryonic Stem Cells into Insulin-Producing Cells: A Network-Based Approach. Interdiscip. Sci. Comput. Life Sci. 2017, 9, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Scardoni, G.; Lau, C. Centralities Based Analysis of Complex Networks. In New Frontiers in Graph Theory; Zhang, Y., Ed.; InTech: Hongkong, China, 2012; pp. 323–348. [Google Scholar]
- He, X.; Zhang, J. Why Do Hubs Tend to Be Essential in Protein Networks? PLoS Genet. 2006, 2, e88. [Google Scholar] [CrossRef] [Green Version]
- Chin, C.-H.; Chen, S.-H.; Wu, H.-H.; Ho, C.-W.; Ko, M.-T.; Lin, C.-Y. cytoHubba: Identifying hub objects and sub-networks from complex interactome. BMC Syst. Biol. 2014, 8, S11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, H.; Kim, P.M.; Sprecher, E.; Trifonov, V.; Gerstein, M. The Importance of Bottlenecks in Protein Networks: Correlation with Gene Essentiality and Expression Dynamics. PLoS Comput. Biol. 2007, 3, e59. [Google Scholar] [CrossRef] [PubMed]
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma’ayan, A. Enrichr: Interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinform. 2013, 14, 128. [Google Scholar] [CrossRef] [Green Version]
- Huang, D.W.; Sherman, B.T.; Tan, Q.; Kir, J.; Liu, D.; Bryant, D.; Guo, Y.; Stephens, R.; Baseler, M.W.; Lane, H.C.; et al. DAVID Bioinformatics Resources: Expanded annotation database and novel algorithms to better extract biology from large gene lists. Nucleic Acids Res. 2007, 35, W169–W175. [Google Scholar] [CrossRef] [PubMed]
- Pletscher-Frankild, S.; Pallejà, A.; Tsafou, K.; Binder, J.X.; Jensen, L.J. DISEASES: Text mining and data integration of disease–gene associations. Methods 2015, 74, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo, D.; GianVincenzo, Z.; Carlo Luca, R.; Karan, G.; Villafañe, J.H.; Roberto, M.; Javad, P. Oral–Gut Microbiota and Arthritis: Is There an Evidence-Based Axis? J. Clin. Med. 2019, 8, 1753. [Google Scholar] [CrossRef] [Green Version]
- Tong, Y.; Zheng, L.; Qing, P.; Zhao, H.; Li, Y.; Su, L.; Zhang, Q.; Zhao, Y.; Luo, Y.; Liu, Y. Oral Microbiota Perturbations Are Linked to High Risk for Rheumatoid Arthritis. Front. Cell. Infect. Microbiol. 2020, 9, 475. [Google Scholar] [CrossRef] [PubMed]
- Aihaiti, Y.; Tuerhong, X.; Ye, J.-T.; Ren, X.-Y.; Xu, P. Identification of pivotal genes and pathways in the synovial tissue of patients with rheumatoid arthritis and osteoarthritis through integrated bioinformatic analysis. Mol. Med. Rep. 2020, 22, 3513–3524. [Google Scholar] [CrossRef] [PubMed]
- Puentes-Osorio, Y.; Amariles, P.; Calleja, M.Á.; Merino, V.; Díaz-Coronado, J.C.; Taborda, D. Potential clinical biomarkers in rheumatoid arthritis with an omic approach. Autoimmun. Highlights 2021, 12, 9. [Google Scholar] [CrossRef]
- Trindade, F.; Oppenheim, F.G.; Helmerhorst, E.J.; Amado, F.; Gomes, P.S.; Vitorino, R. Uncovering the molecular networks in periodontitis. Proteom.-Clin. Appl. 2014, 8, 748–761. [Google Scholar] [CrossRef] [Green Version]
- Hamerman, J.A.; Pottle, J.; Ni, M.; He, Y.; Zhang, Z.-Y.; Buckner, J.H. Negative regulation of TLR signaling in myeloid cells—Implications for autoimmune diseases. Immunol. Rev. 2016, 269, 212–227. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.-R.; Xu, Q.; Xiao, J.; Deng, Y.-M.; Tang, Z.-H.; Tang, Y.-L.; Liu, L.-S. Role of oral microbiota in atherosclerosis. Clin. Chim. Acta Int. J. Clin. Chem. 2020, 506, 191–195. [Google Scholar] [CrossRef]
- de Andrade, K.Q.; Almeida-da-Silva, C.L.C.; Coutinho-Silva, R. Immunological Pathways Triggered by Porphyromonas gingivalis and Fusobacterium nucleatum: Therapeutic Possibilities? Mediat. Inflamm. 2019, 2019, e7241312. [Google Scholar] [CrossRef] [Green Version]
- How, K.Y.; Song, K.P.; Chan, K.G. Porphyromonas gingivalis: An Overview of Periodontopathic Pathogen below the Gum Line. Front. Microbiol. 2016, 7, 53. [Google Scholar] [CrossRef]
- Johnson, L.; Atanasova, K.R.; Bui, P.Q.; Lee, J.; Hung, S.-C.; Yilmaz, Ö.; Ojcius, D.M. Porphyromonas gingivalis attenuates ATP-mediated inflammasome activation and HMGB1 release through expression of a nucleoside-diphosphate kinase. Microbes Infect. 2015, 17, 369–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Xie, J.; Wang, C.; Zhong, D.; Xie, L.; Fang, H. Immunomodulatory Properties of Stem Cells in Periodontitis: Current Status and Future Prospective. Stem Cells Int. 2020, 2020. [Google Scholar] [CrossRef]
- Choi, M.-C.; Jo, J.; Park, J.; Kang, H.K.; Park, Y. NF-κB Signaling Pathways in Osteoarthritic Cartilage Destruction. Cells 2019, 8, 734. [Google Scholar] [CrossRef] [Green Version]
- Mathiessen, A.; Conaghan, P.G. Synovitis in osteoarthritis: Current understanding with therapeutic implications. Arthritis Res. Ther. 2017, 19, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, W.; Xu, H.; Sheng, J.; Xu, Z.; Xie, X.; Zhang, C. Comparative evaluation of the effects of platelet-rich plasma formulations on extracellular matrix formation and the NF-κB signaling pathway in human articular chondrocytes. Mol. Med. Rep. 2017, 15, 2940–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mok, C.C.; Lau, C.S. Pathogenesis of systemic lupus erythematosus. J. Clin. Pathol. 2003, 56, 481–490. [Google Scholar] [CrossRef] [Green Version]
- Cornish, J.; Gillespie, M.T.; Callon, K.E.; Horwood, N.J.; Moseley, J.M.; Reid, I.R. Interleukin-18 Is a Novel Mitogen of Osteogenic and Chondrogenic Cells. Endocrinology 2003, 144, 1194–1201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Z.; Liu, P.; Yang, D.; Wang, F.; Yuan, L.; Lin, Z.; Jiang, J. Interleukin-18-induced inflammatory responses in synoviocytes and chondrocytes from osteoarthritic patients. Int. J. Mol. Med. 2012, 30, 805–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Kuang, W.; Li, D.; Li, Y.; Feng, Y.; Lyu, X.; Huang, G.-B.; Lian, J.-Q.; Yang, X.-F.; Hu, C.; et al. Natural Killer-Like B Cells Secreting Interleukin-18 Induces a Proinflammatory Response in Periodontitis. Front. Immunol. 2021, 12, 641562. [Google Scholar] [CrossRef]
- Li, H.; Miao, D.; Zhu, Q.; Huang, J.; Lu, G.; Xu, W. MicroRNA-17-5p contributes to osteoarthritis progression by binding p62/SQSTM1. Exp. Ther. Med. 2018, 15, 1789–1794. [Google Scholar] [CrossRef] [Green Version]
- Mylka, V.; Deckers, J.; Ratman, D.; De Cauwer, L.; Thommis, J.; De Rycke, R.; Impens, F.; Libert, C.; Tavernier, J.; Vanden Berghe, W.; et al. The autophagy receptor SQSTM1/p62 mediates anti-inflammatory actions of the selective NR3C1/glucocorticoid receptor modulator compound A (CpdA) in macrophages. Autophagy 2018, 14, 2049–2064. [Google Scholar] [CrossRef] [Green Version]
- Robertson, S. FLNB Disorders. In GeneReviews®; Adam, M.P., Ardinger, H.H., Pagon, R.A., Wallace, S.E., Bean, L.J., Mirzaa, G., Amemiya, A., Eds.; University of Washington, Seattle: Seattle, WA, USA, 1993. Available online: http://www.ncbi.nlm.nih.gov/books/NBK2534/ (accessed on 20 June 2021).
- Dong, R.; Du, J.; Wang, L.; Wang, J.; Ding, G.; Wang, S.; Fan, Z. Comparison of Long Noncoding RNA and mRNA Expression Profiles in Mesenchymal Stem Cells Derived from Human Periodontal Ligament and Bone Marrow. BioMed Res. Int. 2014, 2014, e317853. [Google Scholar] [CrossRef] [PubMed]
- Lamsoul, I.; Métais, A.; Gouot, E.; Heuzé, M.L.; Lennon-Duménil, A.-M.; Moog-Lutz, C.; Lutz, P.G. ASB2α regulates migration of immature dendritic cells. Blood 2013, 122, 533–541. [Google Scholar] [CrossRef] [Green Version]
- Kumar S., U.; Sankar, S.; Younes, S.; Kumar D., T.; Ahmad, M.N.; Okashah, S.S.; Kamaraj, B.; Al-Subaie, A.M.; Doss C., G.P.; Zayed, H. Deciphering the Role of Filamin B Calponin-Homology Domain in Causing the Larsen Syndrome, Boomerang Dysplasia, and Atelosteogenesis Type I Spectrum Disorders via a Computational Approach. Molecules 2020, 25, 5543. [Google Scholar] [CrossRef]
- Zhao, Y.; Shapiro, S.S.; Eto, M. F-actin clustering and cell dysmotility induced by the pathological W148R missense mutation of filamin B at the actin-binding domain. Am. J. Physiol.-Cell Physiol. 2016, 310, C89–C98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Im, Y.B.; Jee, M.K.; Choi, J.I.; Cho, H.T.; Kwon, O.H.; Kang, S.K. Molecular targeting of NOX4 for neuropathic pain after traumatic injury of the spinal cord. Cell Death Dis. 2012, 3, e426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papadaki, M.; Rinotas, V.; Violitzi, F.; Thireou, T.; Panayotou, G.; Samiotaki, M.; Douni, E. New Insights for RANKL as a Proinflammatory Modulator in Modeled Inflammatory Arthritis. Front. Immunol. 2019, 10, 97. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.S.; Park, H.J.; Park, J.H.; Hong, E.J.; Jang, G.-Y.; Jung, I.D.; Han, H.D.; Lee, S.-H.; Vo, M.-C.; Lee, J.-J.; et al. A novel function of API5 (apoptosis inhibitor 5), TLR4-dependent activation of antigen presenting cells. OncoImmunology 2018, 7, e1472187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.-C.; Xiao, J.; Peng, J.-L.; Chen, J.-W.; Ma, T.; Cheng, G.-Q.; Dong, Y.-Q.; Wang, W.; Liu, Z.-D. Functional Annotation of Rheumatoid Arthritis and Osteoarthritis Associated Genes by Integrative Genome-Wide Gene Expression Profiling Analysis. PLoS ONE 2014, 9, e85784. [Google Scholar] [CrossRef]
- Nejatbakhsh Samimi, L.; Farhadi, E.; Tahmasebi, M.N.; Jamshidi, A.; Sharafat Vaziri, A.; Mahmoudi, M. NF-κB signaling in rheumatoid arthritis with focus on fibroblast-like synoviocytes. Autoimmun. Highlights 2020, 11, 11. [Google Scholar] [CrossRef]
- Lu, M.-C.; Lai, N.-S.; Chen, H.-C.; Yu, H.-C.; Huang, K.-Y.; Tung, C.-H.; Huang, H.-B.; Yu, C.-L. Decreased microRNA(miR)-145 and increased miR-224 expression in T cells from patients with systemic lupus erythematosus involved in lupus immunopathogenesis. Clin. Exp. Immunol. 2013, 171, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Piatkov, K.I.; Brower, C.S.; Varshavsky, A. The N-end rule pathway counteracts cell death by destroying proapoptotic protein fragments. Proc. Natl. Acad. Sci. USA 2012, 109, E1839–E1847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piatkov, K.I.; Oh, J.-H.; Liu, Y.; Varshavsky, A. Calpain-generated natural protein fragments as short-lived substrates of the N-end rule pathway. Proc. Natl. Acad. Sci. USA 2014, 111, E817–E826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cypher, L.R.; Bielecki, T.A.; Adepegba, O.; Huang, L.; An, W.; Iseka, F.; Luan, H.; Tom, E.; Storck, M.D.; Hoppe, A.D.; et al. CSF-1 receptor signalling is governed by pre-requisite EHD1 mediated receptor display on the macrophage cell surface. Cell. Signal. 2016, 28, 1325–1335. [Google Scholar] [CrossRef] [Green Version]
- Mintz, L.; Galperin, E.; Pasmanik-Chor, M.; Tulzinsky, S.; Bromberg, Y.; Kozak, C.A.; Joyner, A.; Fein, A.; Horowitz, M. EHD1—An EH-Domain-Containing Protein with a Specific Expression Pattern. Genomics 1999, 59, 66–76. [Google Scholar] [CrossRef] [PubMed]
- Miyauchi, S.; Maekawa, T.; Aoki, Y.; Miyazawa, H.; Tabeta, K.; Nakajima, T.; Yamazaki, K. Oral infection with Porphyromonas gingivalis and systemic cytokine profile in C57BL/6.KOR-ApoE shl mice. J. Period. Res. 2012, 47, 402–408. [Google Scholar] [CrossRef]
- Gordon, E.M.; Ravicz, J.R.; Liu, S.; Chawla, S.P.; Hall, F.L. Cell cycle checkpoint control: The cyclin G1/Mdm2/p53 axis emerges as a strategic target for broad-spectrum cancer gene therapy-A review of molecular mechanisms for oncologists. Mol. Clin. Oncol. 2018, 9, 115–134. [Google Scholar] [CrossRef] [Green Version]
- Inaba, H.; Kuboniwa, M.; Bainbridge, B.; Yilmaz, Ö.; Katz, J.; Shiverick, K.T.; Amano, A.; Lamont, R.J. Porphyromonas gingivalis invades human trophoblasts and inhibits proliferation by inducing G1 arrest and apoptosis. Cell. Microbiol. 2009, 11, 1517–1532. [Google Scholar] [CrossRef] [Green Version]
- Lin, T.; Ma, Q.; Zhang, Y.; Zhang, H.; Yan, J.; Gao, C. MicroRNA-27a functions as an oncogene in human osteosarcoma by targeting CCNG1. Oncol. Lett. 2018, 15, 1067–1071. [Google Scholar] [CrossRef] [Green Version]
- Velletri, T.; Huang, Y.; Wang, Y.; Li, Q.; Hu, M.; Xie, N.; Yang, Q.; Chen, X.; Chen, Q.; Shou, P.; et al. Loss of p53 in mesenchymal stem cells promotes alteration of bone remodeling through negative regulation of osteoprotegerin. Cell Death Differ. 2021, 28, 156–169. [Google Scholar] [CrossRef]
- Saito, K.; Murai, J.; Kajiho, H.; Kontani, K.; Kurosu, H.; Katada, T. A Novel Binding Protein Composed of Homophilic Tetramer Exhibits Unique Properties for the Small GTPase Rab5. J. Biol. Chem. 2002, 277, 3412–3418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basel-Vanagaite, L.; Sarig, O.; Hershkovitz, D.; Fuchs-Telem, D.; Rapaport, D.; Gat, A.; Isman, G.; Shirazi, I.; Shohat, M.; Enk, C.D.; et al. RIN2 Deficiency Results in Macrocephaly, Alopecia, Cutis Laxa, and Scoliosis: MACS Syndrome. Am. J. Hum. Genet. 2009, 85, 254–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, Y.; Hagiwara, M.; Ishihara, Y.; Isoda, R.; Sugiura, S.; Komatsu, T.; Ishida, N.; Noguchi, T.; Matsushita, K. TNF-α augmented Porphyromonas gingivalis invasion in human gingival epithelial cells through Rab5 and ICAM-1. BMC Microbiol. 2014, 14, 229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hortle, E.; Tran, L.V.; Fontaine, A.R.; Pinello, N.; Wong, J.J.-L.; Britton, W.J.; Oehlers, S.H. OXSR1 inhibits inflammasome activation by limiting potassium efflux during mycobacterial infection. Life Sci. Alliance 2022, 5, e202201476. [Google Scholar] [CrossRef]
- Hung, C.-M.; Peng, C.-K.; Yang, S.-S.; Shui, H.-A.; Huang, K.-L. WNK4-SPAK modulates lipopolysaccharide-induced macrophage activation. Biochem. Pharmacol. 2020, 171, 113738. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Zhou, Y.; Cao, Y.; Tao, J.; Zhou, Z.-H.; Hang, D.-H. STK39, overexpressed in osteosarcoma, regulates osteosarcoma cell invasion and proliferation. Oncol. Lett. 2017, 14, 4599–4604. [Google Scholar] [CrossRef] [PubMed] [Green Version]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
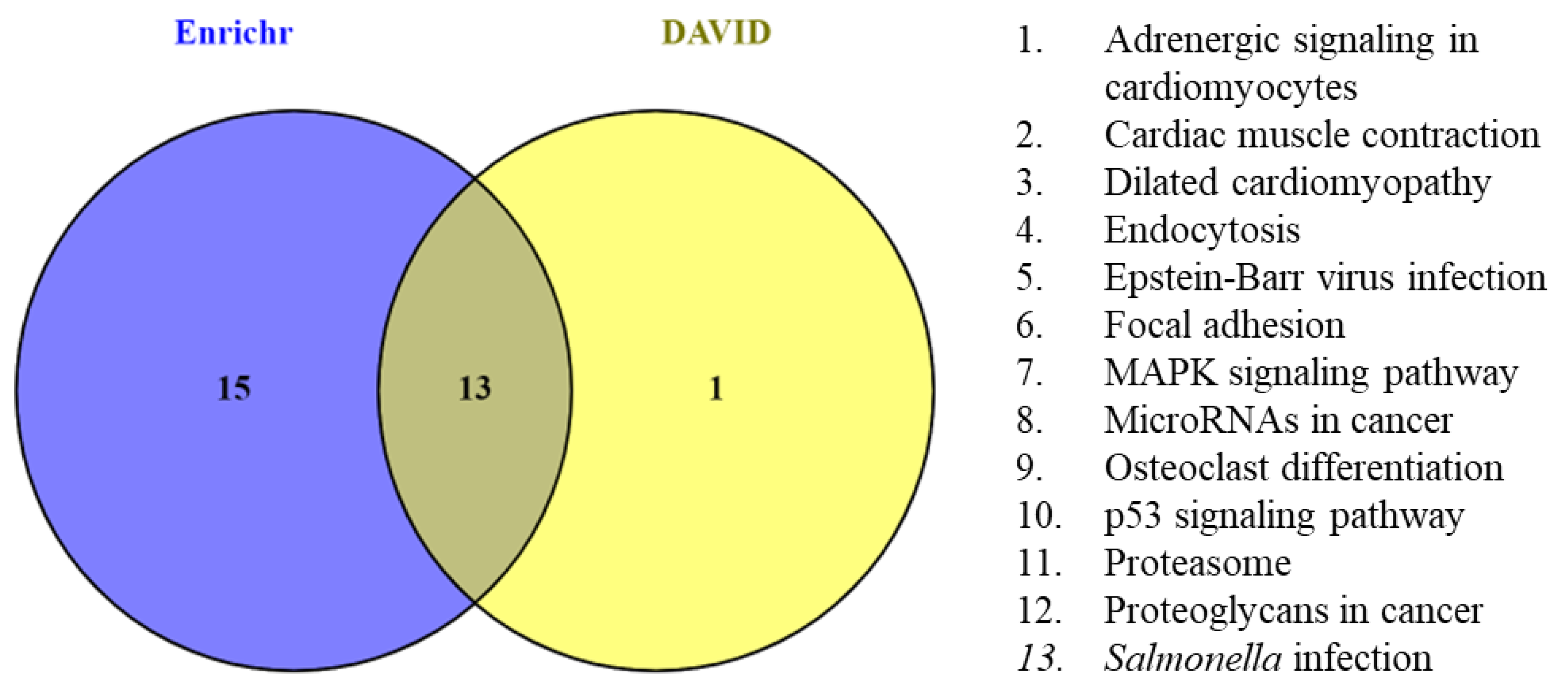{kind=link}
{kind=link}
| Disease | Series ID | Description | No. of Samples | Platform | Year | References |
|---|---|---|---|---|---|---|
| RA | GSE97779 | Expression data from RA synovial macrophages | 14 | GPL570 (Affymetrix Human Genome U133 Plus 2.0 Array) | 2017 | [39] |
| OA | GSE123492 | RNA sequencing of highly pure synovial tissue macrophages reveals two distinct OA subgroups that indicate different disease mechanisms. | 9 | GPL18573 (Illumina NextSeq 500) | 2019 | [6] |
| PG | GSE24897 | Expression data from human macrophages treated with PG and its components. | 6 | GPL570 (Affymetrix Human Genome U133 Plus 2.0 Array) | 2010 | [20] |
| S.No. | Genes | RA | OA | PG | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| r-Value | logFC | p-Value | r-Value | logFC | p-Value | r-Value | logFC | p-Value | ||
| 1 | ABHD5 | 0.88 | −0.566 | 0.002 | 0.9101 | −3.106 | 0.02 | 0.9866 | −0.707 | 0.001 |
| 2 | AKAP13 | 0.839 | 1.093 | 0 | 0.9723 | 2.119 | 0.018 | 0.9253 | 0.925 | 0 |
| 3 | API5 | 0.7908 | 0.949 | 0 | 0.9587 | −1.612 | 0.044 | 0.9681 | −0.838 | 0.004 |
| 4 | ATE1 | 0.8574 | −0.683 | 0 | 0.9346 | −0.909 | 0.011 | 0.9726 | −1.018 | 0.002 |
| 5 | ATP1B1 | 0.7904 | −1.457 | 0.003 | 0.8849 | 3.119 | 0.027 | 0.9819 | 0.777 | 0.001 |
| 6 | ATPAF1 | 0.773 | −0.656 | 0.006 | 0.9537 | −1.314 | 0.004 | 0.9896 | −0.898 | 0.002 |
| 7 | BLOC1S2 | 0.8767 | −1.199 | 0 | 0.9725 | −0.662 | 0.018 | 0.981 | −0.648 | 0 |
| 8 | CAB39L | 0.9258 | −1.924 | 0 | 0.9697 | 4.238 | 0.003 | 0.9658 | −1.231 | 0 |
| 9 | CCDC66 | 0.8811 | −0.65 | 0.002 | 0.8958 | −3.632 | 0.004 | 0.9899 | −0.96 | 0 |
| 10 | CCNG1 | 0.8742 | −0.712 | 0 | 0.923 | −1.613 | 0.009 | 0.9745 | −0.784 | 0.001 |
| 11 | CISD1 | 0.847 | −1.034 | 0.004 | 0.9385 | −2.788 | 0.031 | 0.9263 | −0.809 | 0.002 |
| 12 | CLIP4 | 0.8257 | −0.648 | 0.003 | 0.9052 | −3.548 | 0.011 | 0.9836 | −1.127 | 0 |
| 13 | DAGLB | 0.7146 | 1.117 | 0.002 | 0.9608 | 1.844 | 0.007 | 0.9891 | −0.768 | 0 |
| 14 | EHD1 | 0.8456 | 2.131 | 0 | 0.8832 | −2.591 | 0.041 | 0.9949 | 2.084 | 0 |
| 15 | ERCC1 | 0.8019 | 0.92 | 0.001 | 0.9718 | −2.896 | 0.016 | 0.9864 | −0.647 | 0 |
| 16 | ETFA | 0.9509 | −0.628 | 0 | 0.925 | 1.39 | 0.015 | 0.9891 | −0.528 | 0.003 |
| 17 | FH | 0.8747 | −0.647 | 0 | 0.9047 | −3.761 | 0.007 | 0.9754 | −1.096 | 0.002 |
| 18 | FMNL1 | 0.9714 | 1.529 | 0 | 0.9816 | −2.894 | 0.027 | 0.9806 | 1.036 | 0 |
| 19 | GGCT | 0.8915 | −0.863 | 0 | 0.9109 | 2.605 | 0.012 | 0.9855 | −0.758 | 0 |
| 20 | HNMT | 0.8874 | −1.133 | 0 | 0.8968 | −1.428 | 0.007 | 0.9967 | −0.822 | 0.001 |
| 21 | IER3 | 0.9039 | 1.583 | 0 | 0.8884 | 1.611 | 0.036 | 0.99 | 3.691 | 0 |
| 22 | KBTBD7 | 0.9135 | −3.025 | 0 | 0.8291 | 1.669 | 0.041 | 0.9924 | −0.899 | 0.002 |
| 23 | KDM7A | 0.8482 | 1.082 | 0 | 0.948 | 2.185 | 0.047 | 0.9788 | 0.819 | 0.003 |
| 24 | KLHL28 | 0.8184 | −0.857 | 0 | 0.8628 | 3.24 | 0.035 | 0.9868 | 1.469 | 0 |
| 25 | MBTPS2 | 0.8484 | −0.604 | 0 | 0.8632 | −1.722 | 0 | 0.9795 | −0.779 | 0.003 |
| 26 | MCEE | 0.8159 | −0.739 | 0 | 0.7189 | 0.793 | 0.021 | 0.991 | −1.114 | 0 |
| 27 | METTL25 | 0.7534 | −0.631 | 0.001 | 0.9246 | 0.725 | 0.038 | 0.9649 | −0.708 | 0.003 |
| 28 | MTIF3 | 0.8244 | −0.783 | 0.001 | 0.9307 | −0.618 | 0.046 | 0.9964 | −1.215 | 0 |
| 29 | NLRP3 | 0.8997 | 4.303 | 0 | 0.8854 | −1.11 | 0.017 | 0.9812 | 2.918 | 0 |
| 30 | PMEPA1 | 0.7735 | 1.412 | 0.001 | 0.8759 | −2.231 | 0.042 | 0.9633 | 0.656 | 0.001 |
| 31 | PNRC1 | 0.8301 | 0.597 | 0.007 | 0.7468 | 1.758 | 0.041 | 0.9967 | 1.881 | 0 |
| 32 | PPP1R18 | 0.7838 | 0.598 | 0.001 | 0.9788 | 1.476 | 0.047 | 0.9778 | 1.106 | 0 |
| 33 | PSTPIP1 | 0.8447 | 1.879 | 0 | 0.9272 | 1.495 | 0.045 | 0.9646 | −0.819 | 0 |
| 34 | RCBTB2 | 0.8567 | −0.788 | 0 | 0.8104 | 1.973 | 0.048 | 0.9772 | −1.386 | 0.001 |
| 35 | RGS16 | 0.7793 | 1.141 | 0.006 | 0.8896 | 2.358 | 0.022 | 0.9743 | 1.065 | 0 |
| 36 | RHNO1 | 0.8083 | −0.592 | 0.001 | 0.8469 | 2.126 | 0.031 | 0.9891 | −1.008 | 0.001 |
| 37 | RIN2 | 0.8317 | 3.051 | 0 | 0.7911 | −1.47 | 0.042 | 0.9943 | −1.559 | 0.001 |
| 38 | RPAP3 | 0.8334 | −0.54 | 0.003 | 0.719 | −0.786 | 0.012 | 0.99 | −1.598 | 0 |
| 39 | RPGR | 0.8102 | 0.651 | 0.002 | 0.816 | 2.703 | 0.044 | 0.968 | 0.724 | 0 |
| 40 | RPUSD3 | 0.8895 | −1.102 | 0 | 0.8743 | −0.585 | 0.046 | 0.9632 | −1.082 | 0 |
| 41 | SAMHD1 | 0.8755 | 0.653 | 0 | 0.9581 | −3.903 | 0.017 | 0.9696 | −1.287 | 0.004 |
| 42 | SAR1B | 0.9441 | −0.739 | 0 | 0.9027 | 0.843 | 0 | 0.993 | −0.786 | 0 |
| 43 | SLC20A1 | 0.9401 | 0.572 | 0 | 0.957 | −1.416 | 0.013 | 0.997 | −0.775 | 0.001 |
| 44 | SQSTM1 | 0.9476 | −1.311 | 0 | 0.9383 | −2.452 | 0.03 | 0.9756 | 0.907 | 0.003 |
| 45 | SYNRG | 0.8561 | −0.599 | 0.005 | 0.8443 | −2.746 | 0.007 | 0.9649 | −0.765 | 0 |
| 46 | TAPBP | 0.8511 | 0.913 | 0 | 0.9346 | 2.118 | 0.03 | 0.976 | 0.909 | 0.001 |
| 47 | TBC1D12 | 0.8615 | −0.738 | 0.001 | 0.8791 | 0.655 | 0.004 | 0.9303 | 0.546 | 0.001 |
| S.No. | Gene Name | Node Type | DC | BC | BN | CC |
|---|---|---|---|---|---|---|
| 1 | API5 | query | 21 | 226.67 | 7 | 34.00 |
| 2 | ATE1 | query | 14 | 71.11 | 1 | 32.00 |
| 3 | ATP6V1E1 | result | 13 | 114.54 | 1 | 31.17 |
| 4 | BAG3 | result | 16 | 94.49 | 2 | 32.50 |
| 5 | CCNG1 | query | 14 | 79.93 | 7 | 31.83 |
| 6 | EHD1 | query | 17 | 166.16 | 4 | 33.33 |
| 7 | FLNB | result | 20 | 122.38 | 2 | 34.17 |
| 8 | PSMD12 | result | 15 | 61.81 | 2 | 32.17 |
| 9 | RIN2 | query | 17 | 138.44 | 1 | 33.17 |
| 10 | SQSTM1 | query | 16 | 72.99 | 4 | 31.33 |
| 11 | STK39 | result | 19 | 139.88 | 2 | 34.33 |
| 12 | TPM1 | result | 18 | 99.21 | 1 | 33.17 |
| 13 | TXNIP | result | 15 | 95.66 | 4 | 32.67 |
| 14 | TXNL1 | result | 17 | 155.47 | 2 | 33.50 |
| 15 | XPNPEP1 | result | 18 | 108.06 | 3 | 33.33 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sao, P.; Chand, Y.; Al-Keridis, L.A.; Saeed, M.; Alshammari, N.; Singh, S. Classifying Integrated Signature Molecules in Macrophages of Rheumatoid Arthritis, Osteoarthritis, and Periodontal Disease: An Omics-Based Study. Curr. Issues Mol. Biol. 2022, 44, 3496-3517. https://doi.org/10.3390/cimb44080241
Sao P, Chand Y, Al-Keridis LA, Saeed M, Alshammari N, Singh S. Classifying Integrated Signature Molecules in Macrophages of Rheumatoid Arthritis, Osteoarthritis, and Periodontal Disease: An Omics-Based Study. Current Issues in Molecular Biology. 2022; 44(8):3496-3517. https://doi.org/10.3390/cimb44080241
Chicago/Turabian StyleSao, Prachi, Yamini Chand, Lamya Ahmed Al-Keridis, Mohd Saeed, Nawaf Alshammari, and Sachidanand Singh. 2022. "Classifying Integrated Signature Molecules in Macrophages of Rheumatoid Arthritis, Osteoarthritis, and Periodontal Disease: An Omics-Based Study" Current Issues in Molecular Biology 44, no. 8: 3496-3517. https://doi.org/10.3390/cimb44080241