
Construction of Three High-Density Genetic Linkage Maps and Dynamic QTL Mapping of Growth Traits in Yellow River Carp (Cyprinus carpio haematopterus)

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mapping Family and Measuring Phenotype
2.2. GBS Sequencing and Creation of Maternal (HA), Paternal (AH), and Both Parent (HH) SNP Datasets
2.3. Map Construction and Estimation of Genome Size
2.4. QTL Location and Consensus QTL Analysis
2.5. Screening for Growth-Related Candidate Genes
3. Results
3.1. Phenotype Data
3.2. GBS Tags
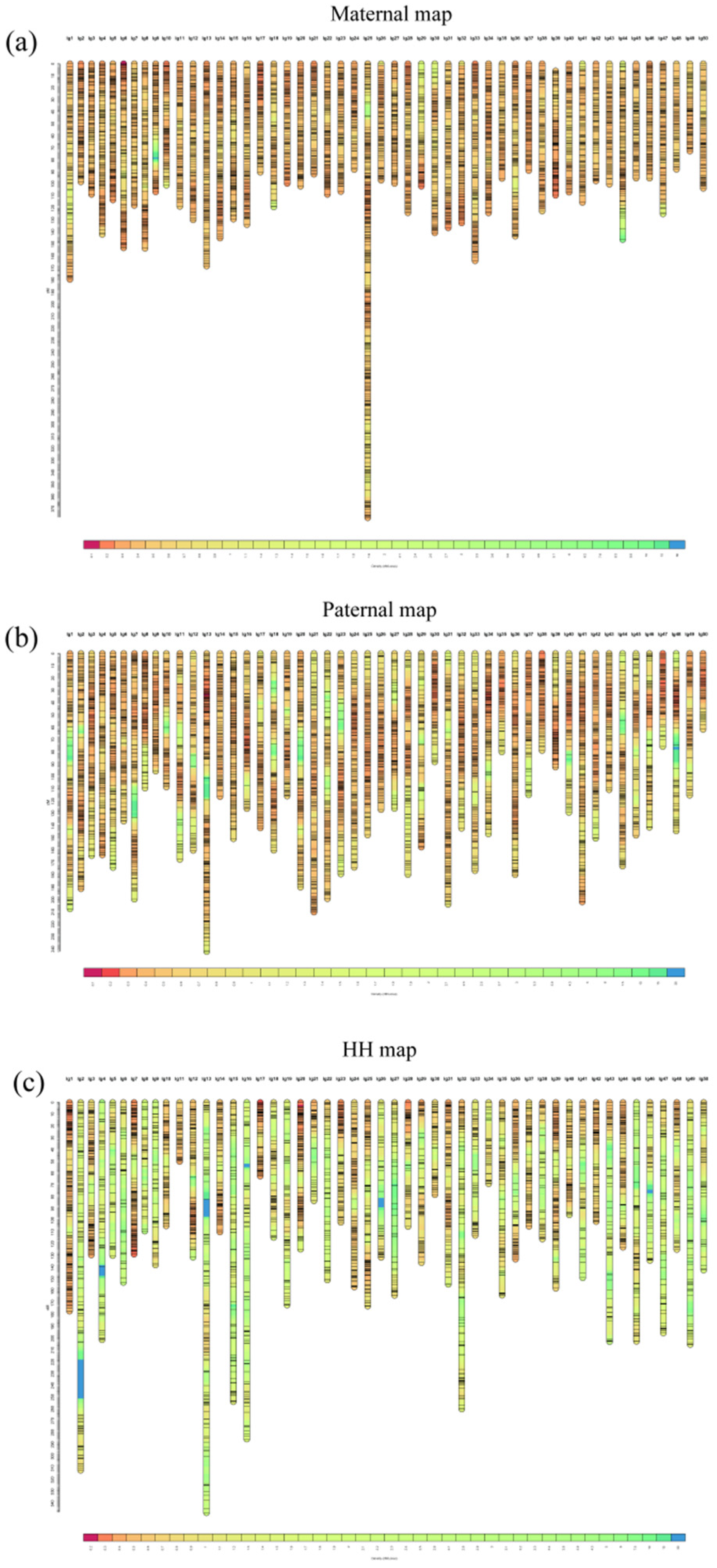
3.3. Construction of the Genetic Linkage Map
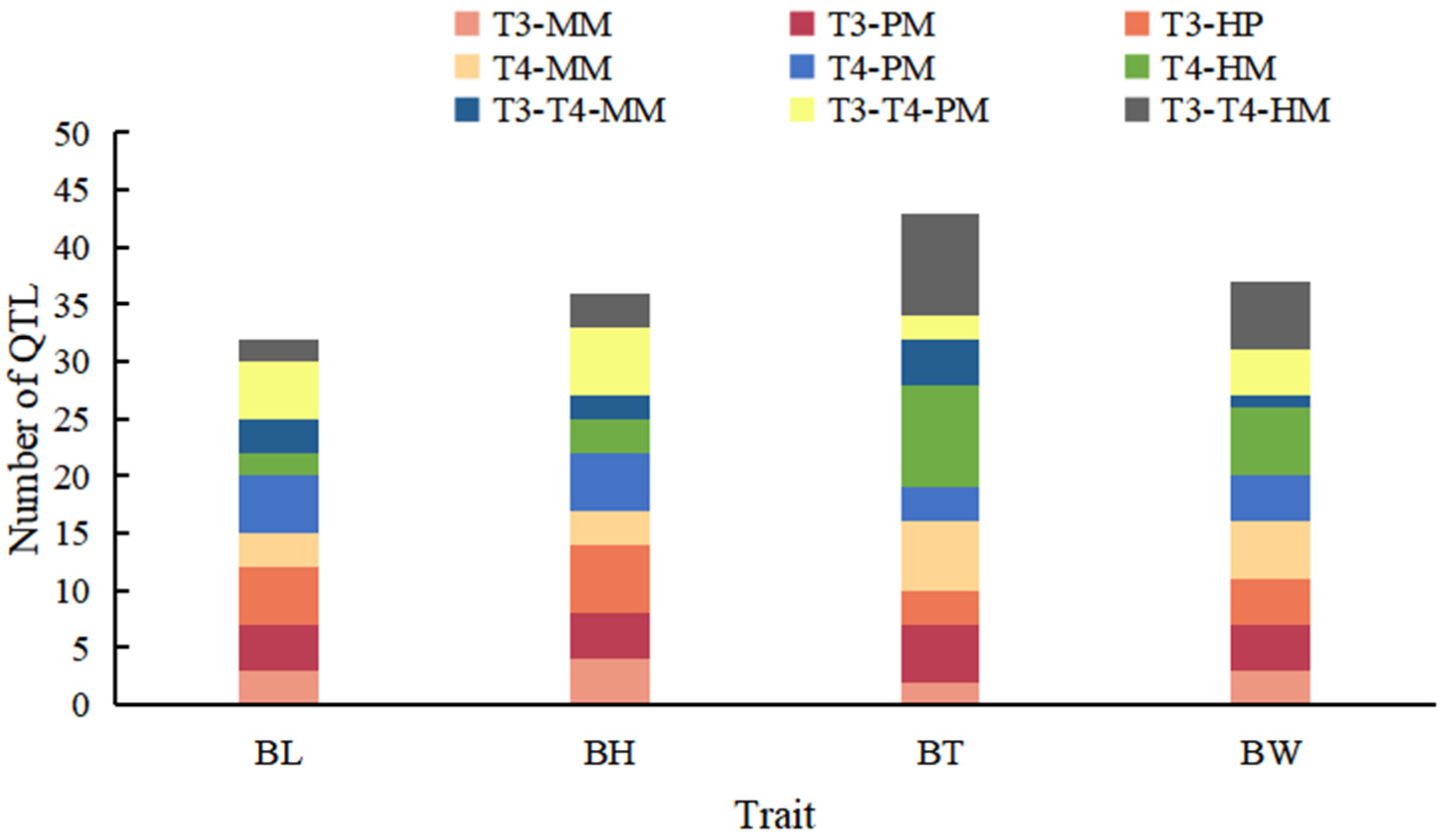
3.4. QTLs for Growth Traits
3.5. Analysis of Consensus QTLs (Cqtls)
3.6. Candidate Growth-Related Genes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Sun, X.; Liang, L. A genetic linkage map of common carp. J. Fish. Sci. China 2000, 7, 1–5. (In Chinese) [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, P.; Lu, C.; Kuang, Y.; Zhang, X.; Cao, D.; Li, C.; Chang, Y.; Hou, N.; Li, H.; et al. Genetic linkage mapping and analysis of muscle fiber-related QTLs in common carp (Cyprinus carpio L.). Mar. Biotechnol. 2011, 13, 376–392. [Google Scholar] [CrossRef]
- Zheng, X.; Kuang, Y.; Zhang, X.; Lu, C.; Cao, D.; Li, C.; Sun, X. A genetic linkage map and comparative genome analysis of common carp (Cyprinus carpio L.) using microsatellites and SNPs. Mol. Genet. Genomics 2011, 286, 261–277. [Google Scholar] [CrossRef]
- Wang, X.; Fu, B.; Yu, X.; Qu, C.; Zhang, Q.; Tong, J. Fine mapping of growth-related quantitative trait loci in Yellow River carp (Cyprinus carpio haematoperus). Aquaculture 2018, 484, 277–285. [Google Scholar] [CrossRef]
- Peng, W.; Xu, J.; Zhang, Y.; Feng, J.; Dong, C.; Jiang, L.; Feng, J.; Chen, B.; Gong, Y.; Chen, L.; et al. An ultra-high density linkage map and QTL mapping for sex and growth-related traits of common carp (Cyprinus carpio). Sci. Rep. 2016, 6, 26693. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Peng, W.; Kong, S.; Pu, F.; Chen, B.; Zhou, Z.; Feng, J.; Li, X.; Xu, P. Genetic Mapping of Head Size Related Traits in Common Carp (Cyprinus carpio). Front. Genet. 2018, 9, 448. [Google Scholar] [CrossRef] [PubMed]
- Li, J.T.; Wang, Q.; Huang, Y.M.; Li, Q.S.; Cui, M.S.; Dong, Z.J.; Wang, H.W.; Yu, J.H.; Zhao, Y.J.; Yang, C.R.; et al. Parallel subgenome structure and divergent expression evolution of allo-tetraploid common carp and goldfish. Nat. Genet. 2021, 53, 1493–1503. [Google Scholar] [CrossRef] [PubMed]
- Laghari, M.Y.; Zhang, Y.; Lashari, P.; Zhang, X.; Xu, P.; Xin, B.; Sun, X. Quantitative trait loci (QTL) associated with growth rate trait in common carp (Cyprinus carpio). Aquacult. Int. 2013, 21, 1373–1379. [Google Scholar] [CrossRef]
- Xu, P.; Zhang, X.; Wang, X.; Li, J.; Liu, G.; Kuang, Y.; Xu, J.; Zheng, X.; Ren, L.; Wang, G.; et al. Genome sequence and genetic diversity of the common carp, Cyprinus carpio. Nat. Genet. 2014, 46, 1212–1219. [Google Scholar] [CrossRef] [Green Version]
- Feng, X.; Yu, X.; Fu, B.; Wang, X.; Liu, H.; Pang, M.; Tong, J. A high-resolution genetic linkage map and QTL fine mapping for growth-related traits and sex in the Yangtze River common carp (Cyprinus carpio haematopterus). BMC Genomics 2018, 19, 230. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J. Analysis of conditional genetic effects and variance components in developmental genetics. Genetics 1995, 141, 1633–1639. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Li, W.; Lu, H. Strategy of dynamic mapping of quantitative trait loci. J. Biomath. 1997, 12, 490–495. (In Chinese) [Google Scholar]
- Li, J.; Lindqvist-Kreuze, H.; Tian, Z.; Liu, J.; Song, B.; Landeo, J.; Portal, L.; Gastelo, M.; Frisancho, J.; Sanchez, L.; et al. Conditional QTL underlying resistance to late blight in a diploid potato population. Theor. Appl. Genet. 2012, 124, 1339–1350. [Google Scholar] [CrossRef]
- Wang, Z.; Wu, X.; Ren, Q.; Chang, X.; Li, R.; Jing, R. QTL mapping for developmental behavior of plant height in wheat (Triticum aestivum L.). Euphytica 2010, 174, 447–458. [Google Scholar] [CrossRef]
- Liu, L.; Lai, Y.; Cheng, J.; Wang, L.; Du, W.; Wang, Z.; Zhang, H.; Wu, R. Dynamic quantitative trait locus analysis of seed vigor at three maturity stages in rice. PLoS ONE 2014, 9, e115732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, Q.; Li, P.; Hu, C.; Hua, H.; Li, Z.; Rong, Y.; Wang, K.; Hua, J. Dynamic QTL and epistasis analysis on seedling root traits in upland cotton. J. Genet. 2014, 93, 63–78. [Google Scholar] [CrossRef] [PubMed]
- McClelland, E.K.; Naish, K.A. Quantitative trait locus analysis of hatch timing, weight, length and growth rate in coho salmon, Oncorhynchus kisutch. Heredity 2010, 105, 562–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutierrez, A.P.; Lubieniecki, K.P.; Davidson, E.A.; Lien, S.; Kent, M.P.; Fukui, S.; Withler, R.E.; Swift, B.; Davidson, W.S. Genetic mapping of quantitative trait loci (QTL) for body-weight in Atlantic salmon (Salmo salar) using a 6.5K SNP array. Aquaculture 2012, 358–359, 61–70. [Google Scholar] [CrossRef]
- Loukovitis, D.; Chatziplis, D.; Batargias, C. Age-dependent QTL affecting body weight in gilthead seabream (Sparus aurata L.). Mediterr. Mar. Sci. 2016, 17, 666–669. [Google Scholar] [CrossRef]
- Qi, P.; Gimode, D.; Saha, D.; Schröder, S.; Chakraborty, D.; Wang, X.; Dida, M.M.; Malmberg, R.L.; Devos, K.M. UGbS-Flex, a novel bioinformatics pipeline for imputation-free SNP discovery in polyploids without a reference genome: Finger millet as a case study. BMC Plant Biol. 2018, 18, 117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, P.; Eudy, D.; Schnable, J.C.; Schmutz, J.; Raymer, P.L.; Devos, K.M. High Density Genetic Maps of Seashore Paspalum Using Genotyping-By-Sequencing and Their Relationship to The Sorghum Bicolor Genome. Sci. Rep. 2019, 9, 12183. [Google Scholar] [CrossRef]
- Wu, Y.; Bhat, P.R.; Close, T.J.; Lonardi, S. Efficient and accurate construction of genetic linkage maps from the minimum spanning tree of a graph. PLoS Genet. 2008, 4, e1000212. [Google Scholar] [CrossRef]
- Lander, E.S.; Green, P.; Abrahamson, J.; Barlow, A.; Daly, M.J.; Lincoln, S.E.; Newberg, L.A. MAPMAKER: An interactive computer package for constructing primary genetic linkage maps of experimental and natural populations. Genomics 1987, 1, 174–181. [Google Scholar] [CrossRef]
- Voorrips, R.E. MapChart: Software for the graphical presentation of linkage maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Basten, C.J.; Zeng, Z.B. Windows QTL Cartographer 2.5; Department of Statistics, North Carolina State University: Raleigh, NC, USA, 2007. [Google Scholar]
- Liu, H.; Fu, B.; Pang, M.; Feng, X.; Wang, X.; Yu, X.; Tong, J. QTL fine mapping and identification of candidate genes for growth-related traits in bighead carp (Hypophthalmichehys nobilis). Aquaculture 2016, 465, 134–143. [Google Scholar] [CrossRef]
- Wang, D.; Zhang, G.; Wei, K.; Ji, W.; Gardner, J.P.A.; Yang, R.; Chen, K. Molecular identification and expression of the Foxl2 gene during gonadal sex differentiation in northern snakehead Channa argus. Fish Physiol. Biochem. 2015, 41, 1419–1433. [Google Scholar] [CrossRef] [PubMed]
- Andrews, K.R.; Good, J.M.; Miller, M.R.; Luikart, G.; Hohenlohe, P.A. Harnessing the power of RADseq for ecological and evolutionary genomics. Nat. Rev. Genet. 2016, 17, 81–92. [Google Scholar] [CrossRef] [Green Version]
- Elshire, R.J.; Glaubitz, J.C.; Sun, Q.; Poland, J.A.; Kawamoto, K.; Buckler, E.S.; Mitchell, S.E. A robust, simple genotyping-by-sequencing (GBS) approach for high diversity species. PLoS ONE 2011, 6, e19379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peterson, B.K.; Weber, J.N.; Kay, E.H.; Fisher, H.S.; Hoekstra, H.E. Double digest RADseq: An inexpensive method for de novo SNP discovery and genotyping in model and non-model species. PLoS ONE 2012, 7, e37135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.; Liu, D.; Zhang, X.; Li, W.; Liu, H.; Hong, W.; Jiang, C.; Guan, N.; Ma, C.; Zeng, H.; et al. SLAF-seq: An efficient method of large-scale de novo SNP discovery and genotyping using high-throughput sequencing. PLoS ONE 2013, 8, e58700. [Google Scholar] [CrossRef]
- Wang, S.; Meyer, E.; McKay, J.K.; Matz, M.V. 2b-RAD: A simple and flexible method for genome-wide genotyping. Nat. Methods 2012, 9, 808–810. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Li, C.; Teng, T.; Shen, F.; Chen, Y.; Wang, Y.; Pan, C.; Ling, Q. Construction of the first high-density genetic linkage map of pikeperch (Sander lucioperca) using specific length amplified fragment (SLAF) sequencing and QTL analysis of growth-related traits. Aquaculture 2018, 497, 299–305. [Google Scholar] [CrossRef]
- Kong, S.; Ke, Q.; Chen, L.; Zhou, Z.; Pu, F.; Zhao, J.; Bai, H.; Peng, W.; Xu, P. Constructing a High-Density Genetic Linkage Map for Large Yellow Croaker (Larimichthys crocea) and Mapping Resistance Trait Against Ciliate Parasite Cryptocaryon irritans. Mar. Biotechnol. 2019, 21, 262–275. [Google Scholar] [CrossRef]
- Zhu, C.; Liu, H.; Pan, Z.; Chang, G.; Wang, H.; Wu, N.; Ding, H.; Yu, X. Construction of a high-density genetic linkage map and QTL mapping for growth traits in Pseudobagrus ussuriensis. Aquaculture 2019, 511, 734213. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, Z.; Guo, X.; Wu, J.; Lu, W.; Zhou, L.; Gui, J. Construction of a high-density genetic linkage map and fine mapping of QTLs for growth and sex-related traits in red-tail catfish (Hemibagrus wyckioides). Aquaculture 2021, 531, 735892. [Google Scholar] [CrossRef]
- Laghari, M.Y.; Lashari, P.; Zhang, X.; Xu, P.; Narejo, N.T.; Xin, B.; Zhang, Y.; Sun, X. QTL mapping for economically important traits of common carp (Cyprinus carpio L.). J. Appl. Genet. 2015, 56, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Everett, M.V.; Seeb, J.E. Detection and mapping of QTL for temperature tolerance and body size in Chinook salmon (Oncorhynchus tshawytscha) using genotyping by sequencing. Evol. Appl. 2014, 7, 480–492. [Google Scholar] [CrossRef]
- Falica, B.K.; Lehnert, S.J.; Pitcher, T.E.; Heath, D.D.; Higgs, D.M. Ontogentic shifts in genetic and maternal effects on length and survival in Chinook salmon (Oncorhynchus tshawytscha). Aquaculture 2017, 468, 218–225. [Google Scholar] [CrossRef]
- Green, B.S. Maternal effects in fish populations. Adv. Mar. Biol. 2008, 54, 1–105. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, A.P.; Lubieniecki, K.P.; Fukui, S.; Withler, R.E.; Swift, B.; Davidson, W.S. Detection of Quantitative Trait Loci (QTL) Related to Grilsing and Late Sexual Maturation in Atlantic Salmon (Salmo salar). Mar. Biotechnol. 2014, 16, 103–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, B.; Wang, Q.; Sun, G.; Ren, X.; Cheng, Y.; Wang, Y.; Gao, S.; Li, C.; Sun, D. Mapping dynamic QTL dissects the genetic architecture of grain size and grain filling rate at different grain-filling stages in barley. Sci. Rep. 2019, 9, 18823. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Trait | T1 | T2 | T1T2 |
|---|---|---|---|
| Full length/cm | 14.3 ± 3.5 | 20.6 ± 4.5 | 6.4 ± 1.6 |
| Body height/cm | 4.4 ± 1.0 | 6.2 ± 1.4 | 1.8 ± 0.6 |
| Body thickness/cm | 2.5 ± 0.6 | 3.7 ± 0.9 | 1.3 ± 0.4 |
| Body weight/g | 82.2 ± 56.8 | 243.1 ± 155.4 | 161.7 ± 102.5 |
| T1-BL | T1-BH | T1-BT | T1-BW | T2-BL | T2-BH | T2-BT | T2-BW | T1T2-BL | T1T2-BH | T1T2-BT | T1T2-BW | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| T1-BL | 1 | 0.946 ** | 0.948 ** | 0.964 ** | 0.945 ** | 0.923 ** | 0.894 ** | 0.929 ** | 0.445 ** | 0.537 ** | 0.441 ** | 0.875 ** |
| T1-BH | 1 | 0.943 ** | 0.943 ** | 0.912 ** | 0.922 ** | 0.881 ** | 0.913 ** | 0.471 ** | 0.442 ** | 0.424 ** | 0.862 ** | |
| T1-BT | 1 | 0.941 ** | 0.909 ** | 0.912 ** | 0.884 ** | 0.899 ** | 0.457 ** | 0.516 ** | 0.348 ** | 0.842 ** | ||
| T1-BW | 1 | 0.914 ** | 0.911 ** | 0.885 ** | 0.956 ** | 0.439 ** | 0.513 ** | 0.434 ** | 0.895 ** | |||
| T2-BL | 1 | 0.951 ** | 0.909 ** | 0.953 ** | 0.713 ** | 0.657 ** | 0.527 ** | 0.938 ** | ||||
| T2-BH | 1 | 0.918 ** | 0.937 ** | 0.625 ** | 0.755 ** | 0.541 ** | 0.916 ** | |||||
| T2-BT | 1 | 0.903 ** | 0.573 ** | 0.633 ** | 0.746 ** | 0.878 ** | ||||||
| T2-BW | 1 | 0.618 ** | 0.624 ** | 0.529 ** | 0.987 ** | |||||||
| T1T2-BL | 1 | 0.649 ** | 0.499 ** | 0.693 ** | ||||||||
| T1T2-BH | 1 | 0.535 ** | 0.663 ** | |||||||||
| T1T2-BT | 1 | 0.562 ** | ||||||||||
| T1T2-BW | 1 |
| Consencus QTL | QTL | LOD | PVE (%) | Marker Position | Marker Chromosome | No. of QTLs |
|---|---|---|---|---|---|---|
| Cqtl-1 | BH-MM-T1T2-2 | 6.33 | 11.69 | 24,895,873 | 1 | 2 |
| BL-MM-T2-1 | 3.42 | 5.78 | ||||
| Cqtl-2 | BT-HM-T2-2 | 3.22 | 4.20 | 29,195,166 | 1 | 2 |
| BT-HM-T1T2-2 | 3.22 | 4.20 | ||||
| Cqtl-3 | BT-HM-T2-1 | 3.09 | 0.67 | 1,774,640 | 3 | 2 |
| BT-HM-T1T2-1 | 3.09 | 0.67 | ||||
| Cqtl-4 | BT-HM-T1-1 | 3.31 | 3.13 | 17,045,253 | 4 | 3 |
| BT-HM-T2-3 | 3.15 | 0.26 | ||||
| BT-HM-T1T2-3 | 3.15 | 0.26 | ||||
| Cqtl-5 | BL-HM-T2-2 | 4.42 | 4.15 | 22,794,002 | 12 | 3 |
| BL-HM-T1T2-2 | 4.42 | 4.15 | ||||
| BW-PM-T2-1 | 5.52 | 9.34 | 23,575,083 | |||
| Cqtl-6 | BL-PM-T1-1 | 7.95 | 13.20 | 24,258,680 | 12 | 2 |
| BT-PM-T1-1 | 7.60 | 12.20 | ||||
| Cqtl-7 | BH-PM-T2-1 | 7.02 | 11.70 | 24,883,274 | 12 | 27 |
| BL-PM-T1-2 | 6.79 | 11.43 | 25,754,793 | |||
| BL-PM-T2-1 | 6.11 | 9.61 | 24,883,274 | |||
| BL-PM-T2-2 | 5.63 | 8.90 | 25,754,793 | |||
| BL-PM-T1T2-3 | 5.34 | 9.24 | 24,980,556 | |||
| BL-HM-T1-1 | 4.60 | 6.91 | 25,680,671 | |||
| BL-HM-T2-1 | 3.93 | 2.78 | 25,507,539 | |||
| BL-HM-T1T2-1 | 3.93 | 2.78 | ||||
| BH-PM-T1-1 | 7.91 | 12.43 | 24,883,274 | |||
| BH-PM-T1-2 | 7.18 | 11.38 | 25,754,793 | |||
| BH-PM-T2-2 | 6.65 | 11.13 | ||||
| BH-HM-T1-2 | 5.89 | 11.20 | 25,680,671 | |||
| BH-HM-T2-2 | 3.18 | 0.62 | 25,735,909 | |||
| BH-HM-T1T2-2 | 3.18 | 0.62 | ||||
| BT-PM-T1-2 | 6.62 | 10.75 | 25,754,793 | |||
| BT-PM-T2-2 | 6.16 | 10.43 | 24,883,274 | |||
| BT-PM-T2-3 | 5.31 | 9.07 | 25,754,793 | |||
| BT-HM-T1-2 | 3.79 | 7.26 | 25,680,671 | |||
| BW-PM-T1-1 | 5.70 | 9.92 | 24,883,274 | |||
| BW-PM-T1-2 | 4.86 | 8.55 | 25,754,793 | |||
| BW-PM-T2-2 | 6.77 | 11.30 | 24,883,274 | |||
| BW-PM-T2-3 | 5.64 | 9.54 | 25,754,793 | |||
| BW-PM-T1T2-1 | 6.97 | 13.06 | 24,883,274 | |||
| BW-PM-T1T2-2 | 6.53 | 12.28 | 25,754,793 | |||
| BW-HM-T1-1 | 5.58 | 5.00 | 25,680,671 | |||
| BW-HM-T2-2 | 5.33 | 4.77 | ||||
| BW-HM-T1T2-2 | 5.57 | 5.11 | ||||
| Cqtl-8 | BH-PM-T1T2-5 | 3.59 | 5.95 | 33,896,198 | 13 | 2 |
| BH-HM-T1-3 | 4.70 | 6.87 | 33,051,888 | |||
| Cqtl-9 | BH-HM-T2-1 | 3.60 | 5.24 | 6,651,371 | 17 | 2 |
| BH-HM-T1T2-1 | 3.60 | 5.24 | ||||
| Cqtl-10 | BT-HM-T2-6 | 3.12 | 0.01 | 9,979,524 | 18 | 2 |
| BT-HM-T1T2-6 | 3.12 | 0.01 | ||||
| Cqtl-11 | BT-HM-T2-4 | 3.80 | 0.13 | 2,672,269 | 19 | 2 |
| BT-HM-T1T2-4 | 3.80 | 0.13 | ||||
| Cqtl-12 | BT-HM-T2-5 | 3.71 | 0.10 | 6,033,807 | 19 | 2 |
| BT-HM-T1T2-5 | 3.71 | 0.10 | ||||
| Cqtl-13 | BT-HM-T2-7 | 3.22 | 1.61 | 8,855,465 | 28 | 2 |
| BT-HM-T1T2-7 | 3.22 | 1.61 | ||||
| Cqtl-14 | BW-HM-T1-2 | 3.03 | 4.90 | 14,096,508 | 29 | 3 |
| BW-HM-T2-3 | 3.06 | 4.99 | ||||
| BW-HM-T1T2-4 | 3.04 | 4.87 | ||||
| Cqtl-15 | BH-HM-T2-3 | 3.30 | 6.27 | 18,356,606 | 29 | 4 |
| BH-HM-T1T2-3 | 3.30 | 6.27 | ||||
| BW-HM-T2-4 | 3.02 | 4.78 | 18,202,561 | |||
| BH-HM-T1-5 | 3.14 | 1.83 | 19,199,891 | |||
| Cqtl-16 | BL-MM-T1-3 | 4.77 | 8.32 | 5,507,998 | 34 | 2 |
| BT-MM-T2-3 | 4.76 | 8.37 | 6,379,096 | |||
| Cqtl-17 | BT-HM-T2-8 | 3.02 | 1.32 | 10,669,401 | 34 | 2 |
| BT-HM-T1T2-8 | 3.02 | 1.32 | ||||
| Cqtl-18 | BW-HM-T1-3 | 3.42 | 3.20 | 4,996,409 | 39 | 3 |
| BW-HM-T2-5 | 3.33 | 3.10 | ||||
| BW-HM-T1T2-5 | 3.58 | 3.44 | ||||
| Cqtl-19 | BL-PM-T2-3 | 3.92 | 6.06 | 9,244,863 | 39 | 6 |
| BL-HM-T1-5 | 3.57 | 0.10 | 9,548,459 | |||
| BH-PM-T1-3 | 3.88 | 5.85 | 8,819,887 | |||
| BH-PM-T2-3 | 3.72 | 5.70 | ||||
| BT-PM-T1-3 | 3.00 | 4.69 | 9,244,863 | |||
| BW-PM-T1T2-3 | 4.36 | 6.43 | 8,819,887 | |||
| Cqtl-20 | BL-PM-T1-3 | 4.78 | 7.34 | 10,941,579 | 39 | 4 |
| BL-PM-T2-4 | 4.44 | 6.83 | 11,297,467 | |||
| BH-PM-T2-4 | 4.12 | 6.28 | ||||
| BW-PM-T1-4 | 3.56 | 5.29 | 10,941,579 | |||
| Cqtl-21 | BL-PM-T1-4 | 4.69 | 7.21 | 16,555,037 | 39 | 7 |
| BL-PM-T2-5 | 4.00 | 6.18 | ||||
| BL-HM-T1-4 | 3.58 | 0.01 | 16,165,443 | |||
| BH-PM-T1-4 | 4.01 | 6.04 | 16,555,037 | |||
| BH-PM-T2-5 | 3.91 | 5.98 | ||||
| BW-PM-T1T2-4 | 3.88 | 5.76 | ||||
| BT-PM-T1-4 | 3.28 | 5.10 | ||||
| Cqtl-22 | BW-PM-T2-4 | 3.17 | 4.61 | 19,390,144 | 39 | 4 |
| BW-HM-T1-4 | 3.69 | 4.17 | 18,795,619 | |||
| BW-HM-T2-6 | 3.62 | 3.98 | ||||
| BW-HM-T1T2-6 | 3.71 | 4.29 | ||||
| Cqtl-23 | BT-HM-T2-9 | 3.07 | 2.48 | 12,218,984 | 40 | 2 |
| BT-HM-T1T2-9 | 3.07 | 2.48 | ||||
| Cqtl-24 | BH-MM-T1-4 | 3.67 | 6.43 | 2,458,218 | 44 | 6 |
| BH-MM-T2-3 | 4.08 | 7.15 | ||||
| BT-MM-T2-6 | 3.82 | 6.55 | ||||
| BT-MM-T1-2 | 4.09 | 7.37 | 3,318,682 | |||
| BW-MM-T1-3 | 3.63 | 6.37 | ||||
| BW-MM-T2-3 | 3.01 | 5.81 | ||||
| Cqtl-25 | BT-MM-T1-1 | 4.41 | 7.92 | 5,304,386 | 44 | 2 |
| BW-MM-T1-2 | 3.46 | 6.09 | ||||
| Cqtl-26 | BW-MM-T2-4 | 3.01 | 5.16 | 17,239,666 | 50 | 3 |
| BW-MM-T2-5 | 3.21 | 5.49 | 16,482,578 | |||
| BL-MM-T2-3 | 3.15 | 5.24 | 16,055,240 |
| Number | Gene Name | Annotation | Gene_Start | Gene_End | Chr | QTL |
|---|---|---|---|---|---|---|
| 1 | ank2 | Ankyrin-2 | 24,866,644 | 24,926,743 | LG1 | BH-MM-T1T2-2 |
| 2 | cd027 | UPF0609 protein C4orf27 homolog | 24,860,171 | 24,864,690 | ||
| 3 | camk2d | Calcium/calmodulin-dependent protein kinase type II delta | 24,930,027 | 24,966,141 | ||
| 4 | syt1 | Synaptotagmin-1 | 24,858,871 | 24,869,140 | LG12 | BW-PM-T1T2-1; BW-PM-T2-2; BT-PM-T2-2; BH-PM-T2-1; BH-PM-T1-1 |
| 5 | manbl | Protein MANBAL | 24,873,472 | 24,874,795 | LG12 | BH-PM-T1-1; BH-PM-T2-1; BT-PM-T2-2; BW-PM-T2-2; BW-PM-T1T2-1 |
| 6 | nfatc2 | Nuclear factor of activated T-cells, cytoplasmic 2 | 24,876,144 | 24,887,937 | LG12 | |
| 7 | rnf182 | E3 ubiquitin-protein ligase RNF182 | 24,897,900 | 24,898,634 | LG12 | |
| 8 | rgs9bp | Regulator of G-protein signaling 9-binding protein | 24,900,978 | 24,901,894 | LG12 | |
| 9 | ube2c | Ubiquitin-conjugating enzyme E2 C | 24,902,429 | 24,904,315 | LG12 | |
| 10 | dnttip1 | Deoxynucleotidyltransferase terminal-interacting protein 1 | 24,905,995 | 24,909,672 | LG12 | |
| 11 | pcif1 | Phosphorylated CTD-interacting factor 1 | 24,913,041 | 24,921,314 | LG12 | |
| 12 | pltp | Phospholipid transfer protein | 24,928,567 | 24,932,818 | LG12 | |
| 13 | znfx1 | Zinc finger NFX1-type containing 1 | 25,678,811 | 25,690,871 | LG12 | BH-HM-T1-2 |
| 14 | snrpb | Small nuclear ribonucleoprotein-associated protein B′ | 25,708,066 | 25,711,568 | LG12 | BL-PM-T1-2; BH-PM-T1-2; BH-PM-T2-2; BH-HM-T1-2; BT-PM-T1-2; BW-PM-T1T2-2 |
| 15 | itch | Itchy E3 ubiquitin protein ligase | 25,714,170 | 25,726,476 | LG12 | BH-PM-T1-2; BH-PM-T2-2; BH-HM-T1-2; BT-PM-T1-2; BW-PM-T1T2-2 |
| 16 | neca1 | N-terminal EF-hand calcium-binding protein 1 | 25,732,452 | 25,764,815 | LG12 | BL-PM-T1-2; BH-PM-T1-2; BH-PM-T2-2; BT-PM-T1-2; BW-PM-T1T2-2 |
| 17 | cbfa2t2 | Protein CBFA2T2 | 25,771,196 | 25,808,027 | LG12 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Jia, S.; Zhang, Y.; Jiang, S.; Chen, Y.; Chen, J.; Yu, M.; Zhang, L.; Qiao, Z.; Li, X. Construction of Three High-Density Genetic Linkage Maps and Dynamic QTL Mapping of Growth Traits in Yellow River Carp (Cyprinus carpio haematopterus). Curr. Issues Mol. Biol. 2021, 43, 2276-2288. https://doi.org/10.3390/cimb43030160
Wang L, Jia S, Zhang Y, Jiang S, Chen Y, Chen J, Yu M, Zhang L, Qiao Z, Li X. Construction of Three High-Density Genetic Linkage Maps and Dynamic QTL Mapping of Growth Traits in Yellow River Carp (Cyprinus carpio haematopterus). Current Issues in Molecular Biology. 2021; 43(3):2276-2288. https://doi.org/10.3390/cimb43030160
Chicago/Turabian StyleWang, Lei, Songpeng Jia, Yuxuan Zhang, Shuhong Jiang, Yuhan Chen, Junping Chen, Miao Yu, Lan Zhang, Zhigang Qiao, and Xuejun Li. 2021. "Construction of Three High-Density Genetic Linkage Maps and Dynamic QTL Mapping of Growth Traits in Yellow River Carp (Cyprinus carpio haematopterus)" Current Issues in Molecular Biology 43, no. 3: 2276-2288. https://doi.org/10.3390/cimb43030160