Protamine-2 Deficiency Initiates a Reactive Oxygen Species (ROS)-Mediated Destruction Cascade during Epididymal Sperm Maturation in Mice

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Protein Isolation from Tissue
2.3. Protein Isolation from Sperm
2.4. Immunoblotting
2.5. Mass Spectrometry
2.6. Immunohistochemistry
2.7. Enzyme-Linked Immunosorbent Assay (ELISA)
2.8. Sperm Vitality Assessment
2.9. Periodic Acid Schiff Staining (PAS)
2.10. Sperm Nuclear Morphology Analysis
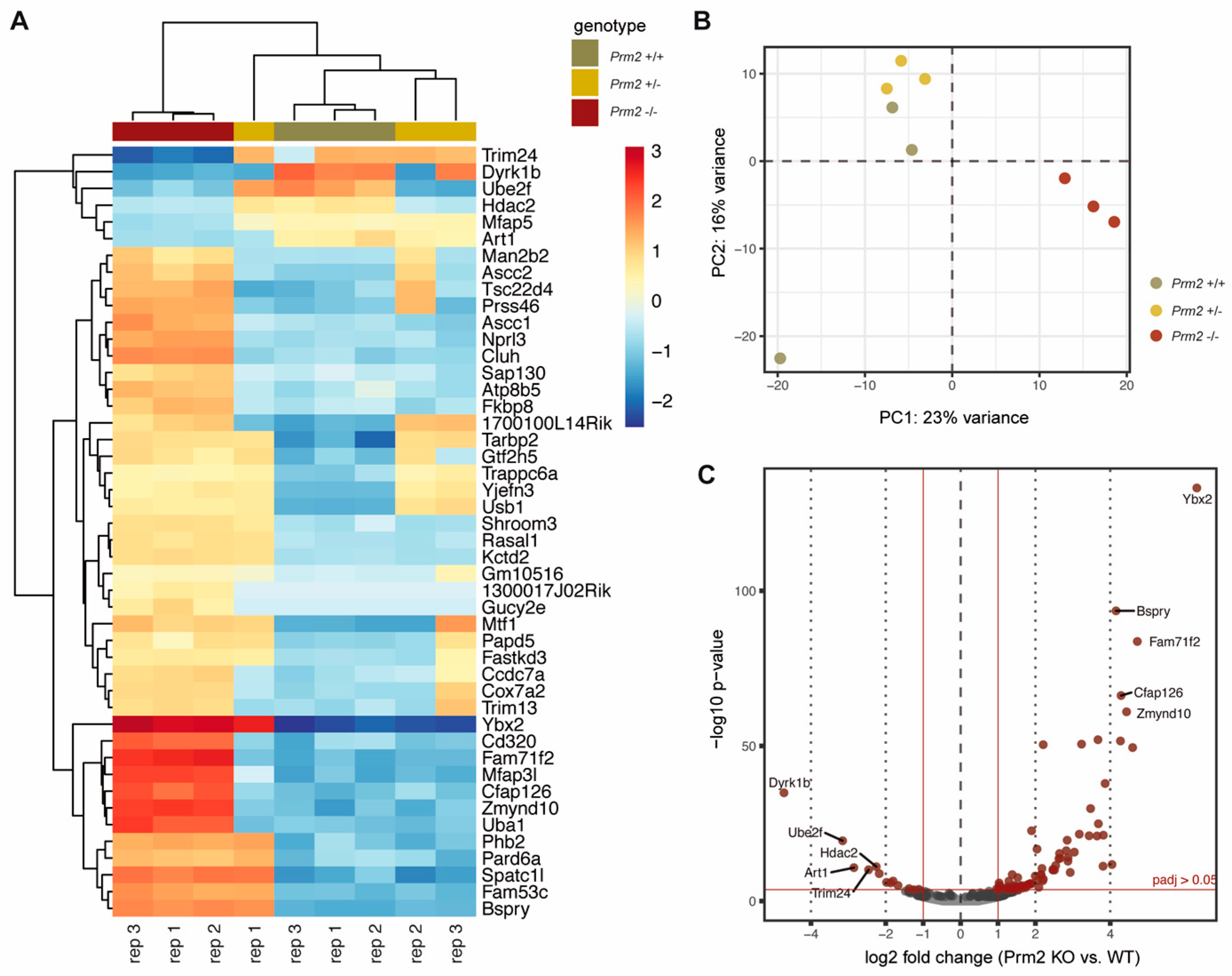
2.11. RNAseq Analysis
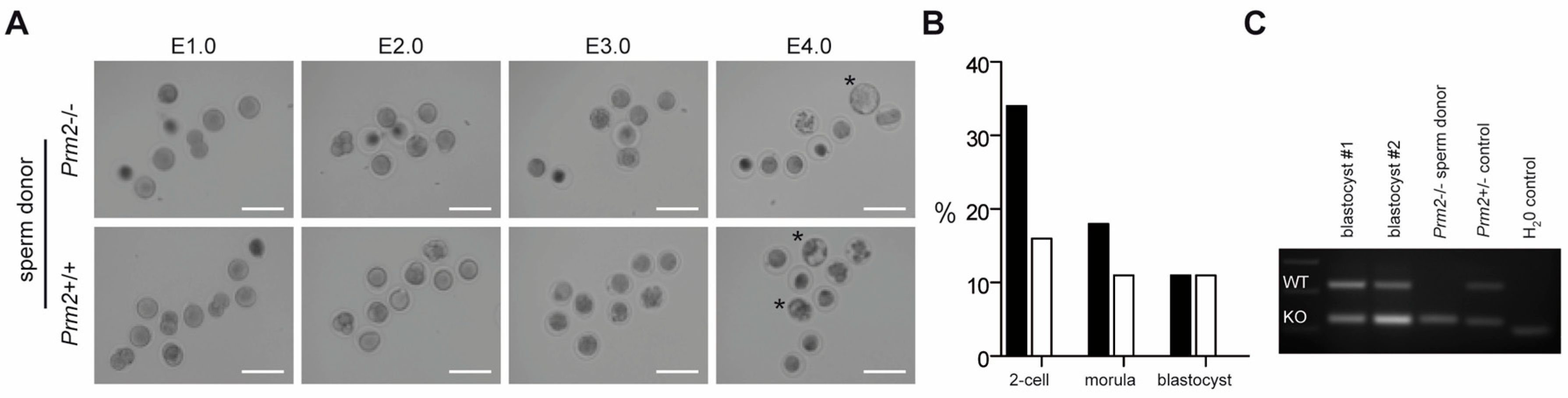
2.12. Intracytoplasmatic Sperm Injection (ICSI)
2.13. Genotyping of Blastocysts
3. Results
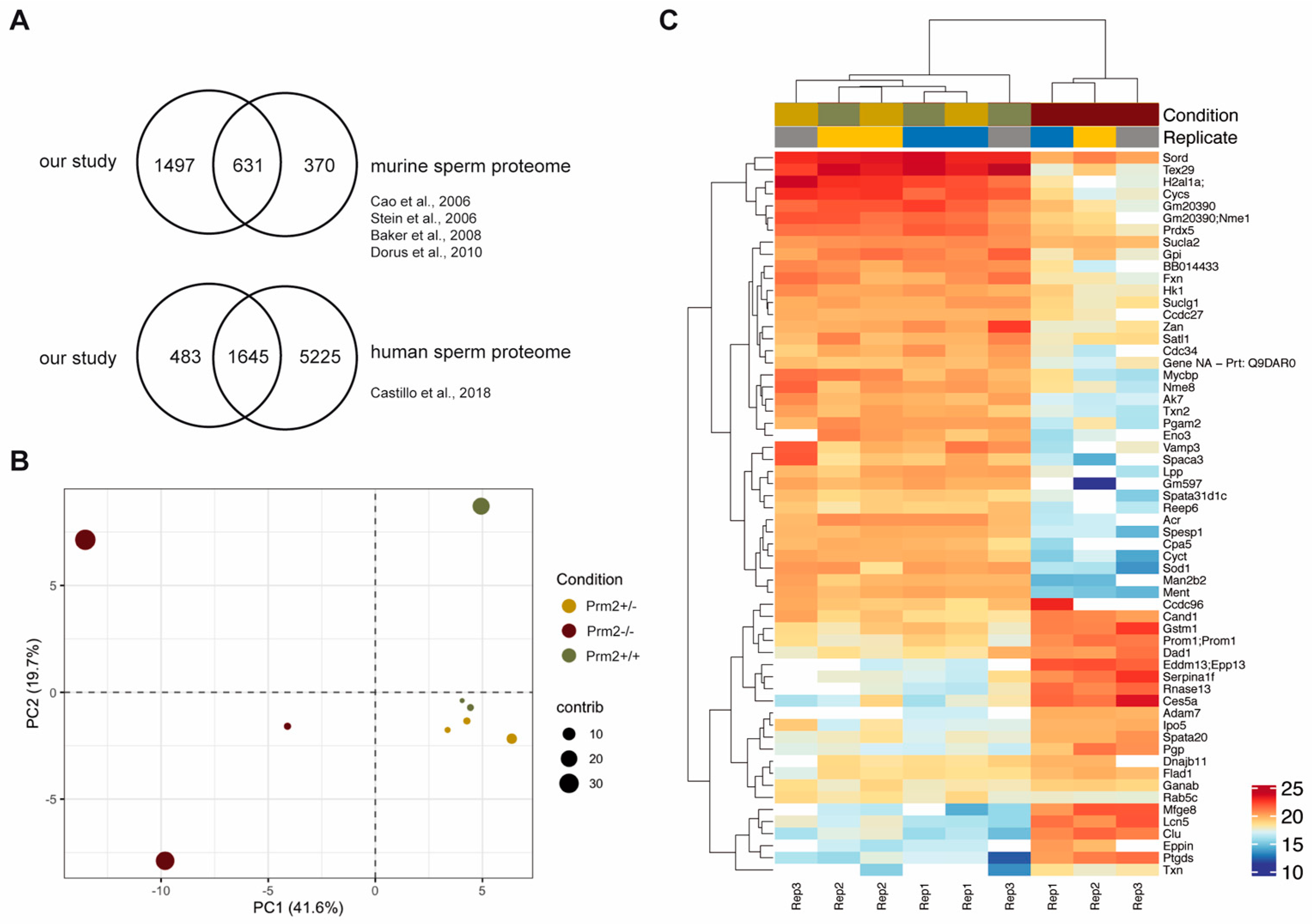
3.1. To the Molecular Origin of Secondary Defects in Prm2-Deficient Sperm
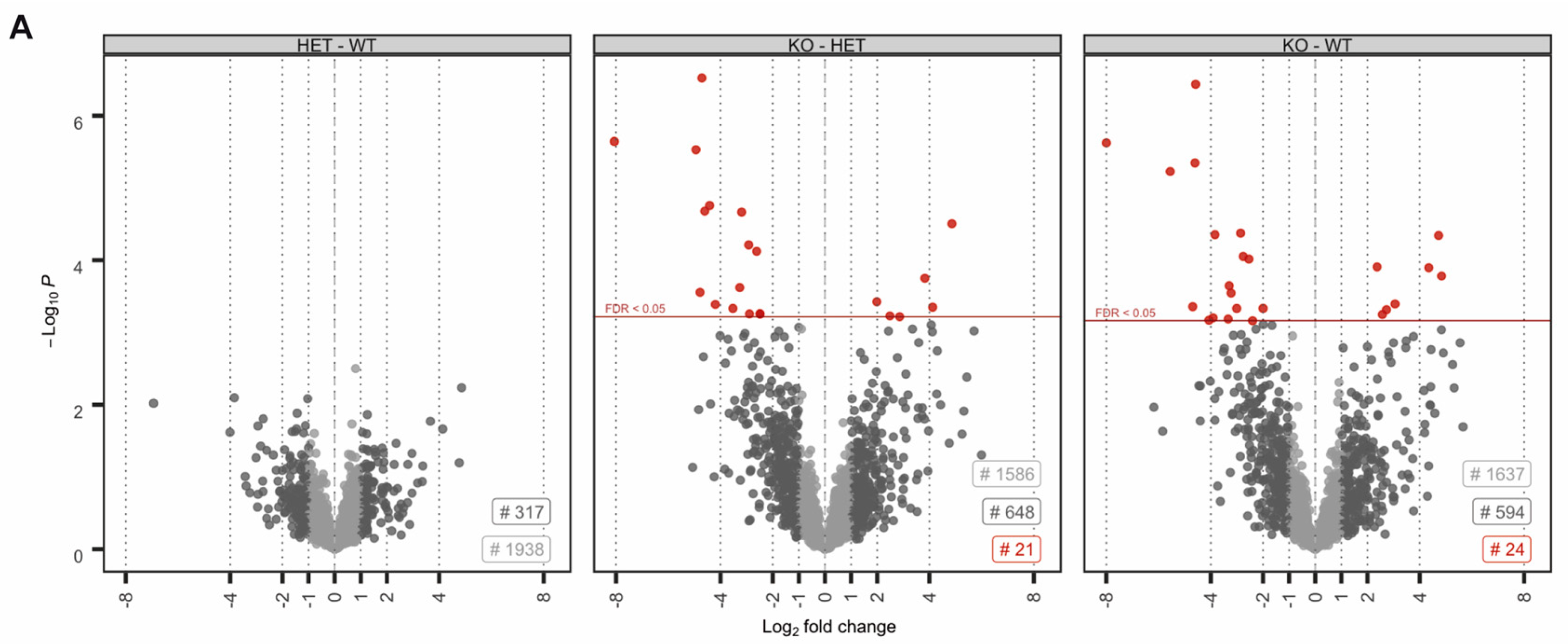
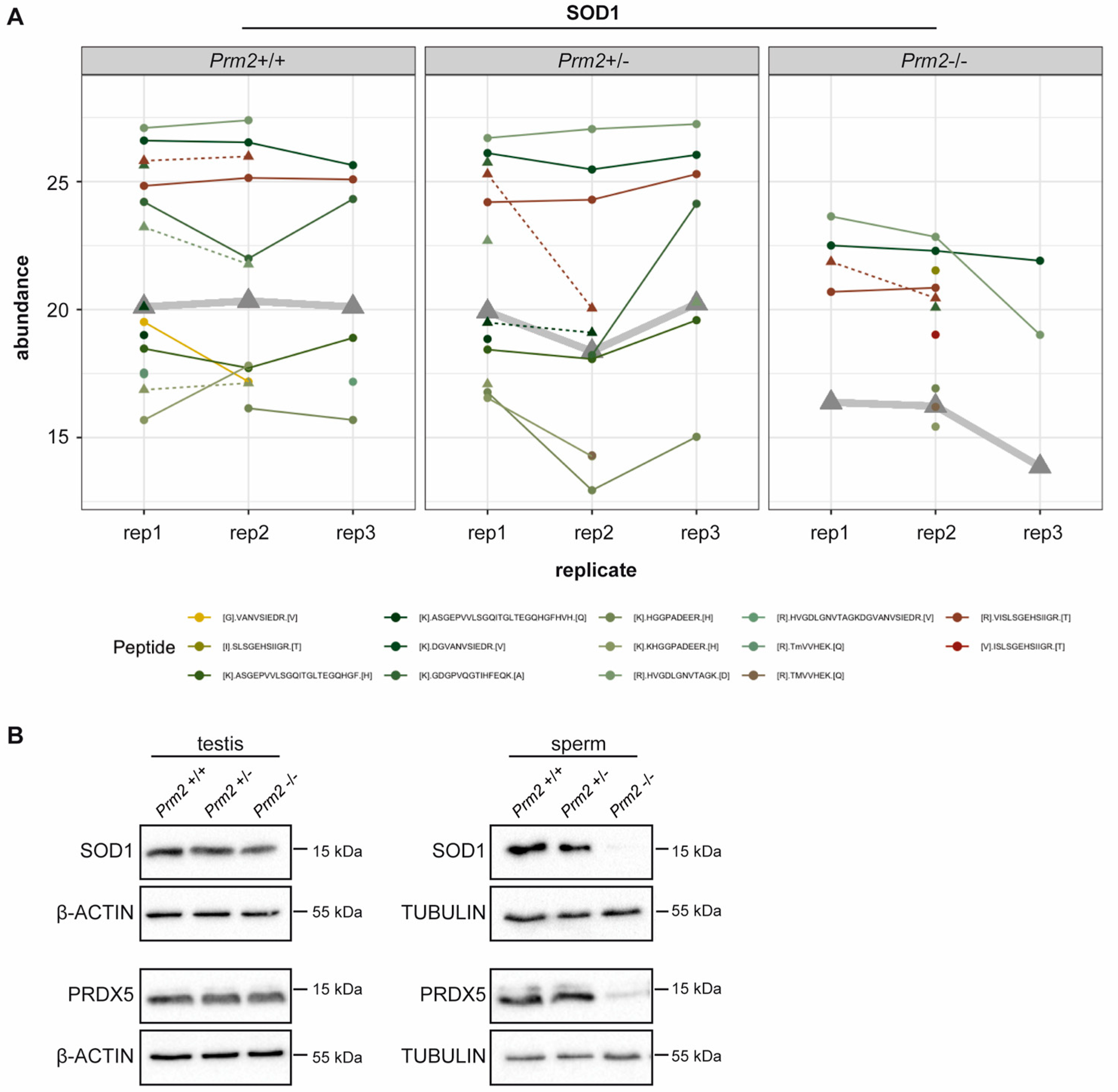
3.2. Prm2 Deficiency Induces a Downregulation of ROS Scavenger Proteins
3.3. Oxidative Stress Causes DNA Damage During Epididymal Sperm Maturation
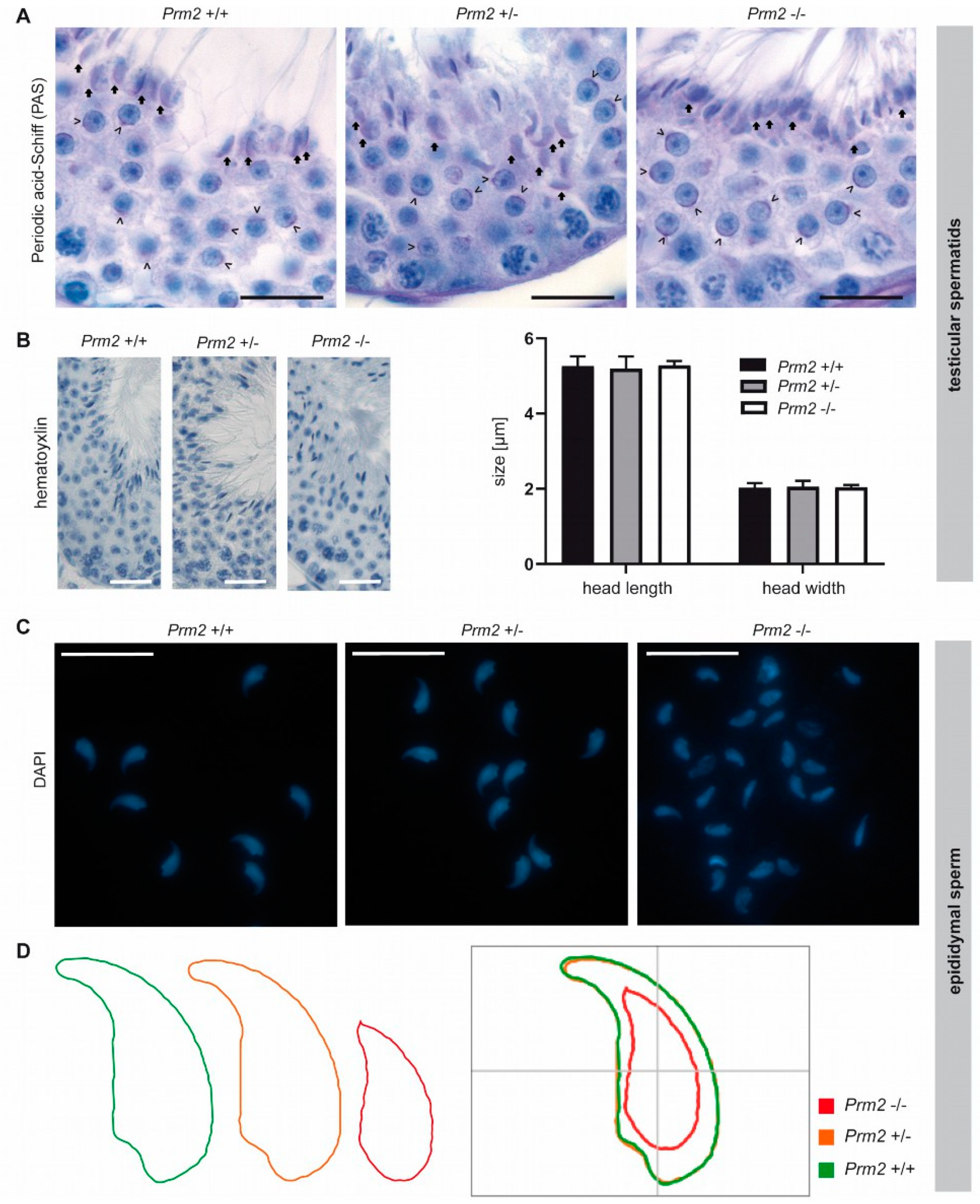
3.4. Secondary Sperm Defects Arise During Epididymal Maturation
3.5. Blastocysts Stage Embryos Can Be Derived from ICSI of Prm2-Deficient Testicular Sperm
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Barral, S.; Morozumi, Y.; Tanaka, H.; Montellier, E.; Govin, J.; De Dieuleveult, M.; Charbonnier, G.; Couté, Y.; Puthier, D.; Buchou, T.; et al. Histone Variant H2A.L.2 Guides Transition Protein-Dependent Protamine Assembly in Male Germ Cells. Mol. Cell 2017, 66, 89–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hud, N.; Allen, M.; Downing, K.; Lee, J.; Balhorn, R. Identification of the Elemental Packing Unit of DNA in Mammalian Sperm Cells by Atomic Force Microscopy. Biochem. Biophys. Res. Commun. 1993, 193, 1347–1354. [Google Scholar] [CrossRef] [PubMed]
- Ward, W.S. Function of sperm chromatin structural elements in fertilization and development. Mol. Hum. Reprod. 2009, 16, 30–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rathke, C.; Baarends, W.M.; Awe, S.; Renkawitz-Pohl, R. Chromatin dynamics during spermiogenesis. Biochim. Biophys. Acta Bioenerg. 2014, 1839, 155–168. [Google Scholar] [CrossRef] [Green Version]
- Balhorn, R. A model for the structure of chromatin in mammalian sperm. J. Cell Boil. 1982, 93, 298–305. [Google Scholar] [CrossRef] [Green Version]
- Balhorn, R. The protamine family of sperm nuclear proteins. Genome Boil. 2007, 8, 227. [Google Scholar] [CrossRef]
- Corzett, M.; Mazrimas, J.; Balhorn, R. Protamine 1: Protamine 2 stoichiometry in the sperm of eutherian mammals. Mol. Reprod. Dev. 2002, 61, 519–527. [Google Scholar] [CrossRef]
- Balhorn, R.; Reed, S.; Tanphaichitr, N. Aberrant protamine 1/protamine 2 ratios in sperm of infertile human males. Experientia 1988, 44, 52–55. [Google Scholar] [CrossRef]
- De Yebra, L.; Ballescà, J.L.; Vanrell, J.A.; Bassas, L.; Oliva, R. Complete selective absence of protamine P2 in humans. J. Boil. Chem. 1993, 268, 10553–10557. [Google Scholar]
- Belokopytova, I.A.; Kostyleva, E.I.; Tomilin, A.N.; Vorob’Ev, V.I. Human male infertility may be due to a decrease of the protamine P2 content in sperm chromatin. Mol. Reprod. Dev. 1993, 34, 53–57. [Google Scholar] [CrossRef]
- Khara, K.K.; Vlad, M.; Griffiths, M.; Kennedy, C.R. Human protamines and male infertility. J. Assist. Reprod. Genet. 1997, 14, 282–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steger, K.; Fink, L.; Failing, K.; Bohle, R.M.; Kliesch, S.; Weidner, W.; Bergmann, M. Decreased protamine-1 transcript levels in testes from infertile men. Mol. Hum. Reprod. 2003, 9, 331–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aoki, V.W.; Liu, L.; Carrell, D.T. Identification and evaluation of a novel sperm protamine abnormality in a population of infertile males. Hum. Reprod. 2005, 20, 1298–1306. [Google Scholar] [CrossRef] [Green Version]
- Oliva, R. Protamines and male infertility. Hum. Reprod. Update 2006, 12, 417–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, K.; Spiess, A.N.; Schuppe, H.C.; Steger, K. The impact of sperm protamine deficiency and sperm DNA damage on human male fertility: A systematic review and meta-analysis. J. Androl. 2016, 4, 789–799. [Google Scholar] [CrossRef] [PubMed]
- Amor, H.; Zeyad, A.; Ben Ali, H.; Hammadeh, M. Protamines ratio (P1/P2) in spermatozoa as biomarker for male infertility. Fertil. Steril. 2017, 108, e142. [Google Scholar] [CrossRef]
- Torregrosa, N.; Domínguez-Fandos, D.; Camejo, M.I.; Shirley, C.R.; Meistrich, M.L.; Ballescà, J.L.; Oliva, R. Protamine 2 precursors, protamine 1/protamine 2 ratio, DNA integrity and other sperm parameters in infertile patients. Hum. Reprod. 2006, 21, 2084–2089. [Google Scholar] [CrossRef]
- Simon, L.; Castillo, J.; Oliva, R.; Lewis, S.E. Relationships between human sperm protamines, DNA damage and assisted reproduction outcomes. Reprod. Biomed. Online 2011, 23, 724–734. [Google Scholar] [CrossRef] [Green Version]
- García-Peiró, A.; Martínez-Heredia, J.; Oliver-Bonet, M.; Abad, C.; Amengual, M.J.; Navarro, J.; Jones, C.; Coward, K.; Gosálvez, J.; Benet, J. Protamine 1 to protamine 2 ratio correlates with dynamic aspects of DNA fragmentation in human sperm. Fertil. Steril. 2011, 95, 105–109. [Google Scholar] [CrossRef]
- Aoki, V.W.; Moskovtsev, S.I.; Willis, J.; Mullen, B.; Carrell, D.T.; Liu, L. DNA Integrity Is Compromised in Protamine-Deficient Human Sperm. J. Androl. 2005, 26, 741–748. [Google Scholar] [CrossRef]
- Aoki, V.W.; Emery, B.R.; Liu, L.; Carrell, D.T. Protamine Levels Vary Between Individual Sperm Cells of Infertile Human Males and Correlate with Viability and DNA Integrity. J. Androl. 2006, 27, 890–898. [Google Scholar] [CrossRef] [PubMed]
- Iranpour, F.G. The effects of protamine deficiency on ultrastructure of human sperm nucleus. Adv. Biomed. Res. 2014, 3, 24. [Google Scholar] [CrossRef] [PubMed]
- Nasr-Esfahani, M.H.; Salehi, M.; Razavi, S.; Mardani, M.; Bahramian, H.; Steger, K.; Oreizi, F. Effect of protamine-2 deficiency on ICSI outcome. Reprod. Biomed. Online 2004, 9, 652–658. [Google Scholar] [CrossRef]
- Aoki, V.W.; Liu, L.; Jones, K.P.; Hatasaka, H.H.; Gibson, M.; Peterson, C.M.; Carrell, D.T. Sperm protamine 1/protamine 2 ratios are related to in vitro fertilization pregnancy rates and predictive of fertilization ability. Fertil. Steril. 2006, 86, 1408–1415. [Google Scholar] [CrossRef]
- Rogenhofer, N.; Dansranjavin, T.; Schorsch, M.; Spiess, A.; Wang, H.; Von Schönfeldt, V.; Cappallo-Obermann, H.; Baukloh, V.; Yang, H.; Paradowska, A.; et al. The sperm protamine mRNA ratio as a clinical parameter to estimate the fertilizing potential of men taking part in an ART programme. Hum. Reprod. 2013, 28, 969–978. [Google Scholar] [CrossRef] [Green Version]
- Cho, C.; Willis, W.D.; Goulding, E.H.; Jungha, H.; Choi, Y.; Hecht, N.B.; Eddy, E.M. Haploinsufficiency of protamine-1 or -2 causes infertility in mice. Nat. Genet. 2001, 28, 82–86. [Google Scholar] [CrossRef]
- Mashiko, D.; Fujihara, Y.; Satouh, Y.; Miyata, H.; Isotani, A.; Ikawa, M. Generation of mutant mice by pronuclear injection of circular plasmid expressing Cas9 and single guided RNA. Sci. Rep. 2013, 3, 3355. [Google Scholar] [CrossRef]
- Takeda, N.; Yoshinaga, K.; Furushima, K.; Takamune, K.; Li, Z.; Abe, S.I.; Aizawa, S.I.; Yamamura, K.I. Viable offspring obtained from Prm1-deficient sperm in mice. Sci. Rep. 2016, 6, 27409. [Google Scholar] [CrossRef] [Green Version]
- Schneider, S.; Balbach, M.; Jikeli, J.F.; Fietz, D.; Nettersheim, D.; Jostes, S.; Schmidt, R.; Kressin, M.; Bergmann, M.; Wachten, D.; et al. Re-visiting the Protamine-2 locus: Deletion, but not haploinsufficiency, renders male mice infertile. Sci. Rep. 2016, 6, 1–13. [Google Scholar] [CrossRef]
- Ren, Z.X.; You, W.; Wu, S.; Poetsch, A.; Xu, C. Secretomic analyses of Ruminiclostridium papyrosolvens reveal its enzymatic basis for lignocellulose degradation. Biotechnol. Biofuels 2019, 12, 183. [Google Scholar] [CrossRef] [Green Version]
- Mellacheruvu, D.; Wright, Z.; Couzens, A.L.; Lambert, J.P.; St-Denis, N.A.; Li, T.; Miteva, Y.V.; Hauri, S.; Sardiu, M.E.; Low, T.Y.; et al. The CRAPome: A contaminant repository for affinity purification–mass spectrometry data. Nat. Methods 2013, 10, 730–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The, M.; MacCoss, M.J.; Noble, W.S.; Käll, L. Fast and Accurate Protein False Discovery Rates on Large-Scale Proteomics Data Sets with Percolator 3.0. J. Am. Soc. Mass Spectrom. 2016, 27, 1719–1727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Llinares, M.B.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res. 2018, 47, D442–D450. [Google Scholar] [CrossRef] [PubMed]
- Anonymous. The R Project for Statistical Computing. Available online: http://www.r-project.org/ (accessed on 13 February 2012).
- Huber, W.; Von Heydebreck, A.; Sültmann, H.; Poustka, A.; Vingron, M. Variance stabilization applied to microarray data calibration and to the quantification of differential expression. Bioinformatics 2002, 18, S96–S104. [Google Scholar] [CrossRef]
- Le, S.; Josse, J.; Husson, F. FactoMineR: An RPackage for Multivariate Analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Gu, Z.; Eils, R.; Schlesner, M. Complex heatmaps reveal patterns and correlations in multidimensional genomic data. Bioinformatics 2016, 32, 2847–2849. [Google Scholar] [CrossRef] [Green Version]
- Nettersheim, D.; Heukamp, L.C.; Fronhoffs, F.; Grewe, M.J.; Haas, N.; Waha, A.; Honecker, F.; Waha, A.; Kristiansen, G.; Schorle, H. Analysis of TET Expression/Activity and 5mC Oxidation during Normal and Malignant Germ Cell Development. PLoS ONE 2013, 8, e82881. [Google Scholar] [CrossRef]
- Weyrich, A. Preparation of Genomic DNA from Mammalian Sperm. Curr. Protoc. Mol. Boil. 2012, 98. [Google Scholar] [CrossRef]
- Moskovtsev, S.I.; Librach, C.L. Methods of sperm vitality assessment. Methods Mol. Biol. 2013, 927, 13–19. [Google Scholar]
- Skinner, B.M.; Rathje, C.C.; Bacon, J.; Johnson, E.E.P.; Larson, E.L.; Kopania, E.E.K.; Good, J.M.; Yousafzai, G.; Affara, N.A.; Ellis, P.J. A high-throughput method for unbiased quantitation and categorisation of nuclear morphology. Biol. Reprod. 2018, 312470. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Langmead, B.; Salzberg, S. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. BioMed Cent. 2014, 15, 002832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durinck, S.; Moreau, Y.; Kasprzyk, A.; Davis, S.; De Moor, B.; Brazma, A.; Huber, W. BioMart and Bioconductor: A powerful link between biological databases and microarray data analysis. Bioinformatics 2005, 21, 3439–3440. [Google Scholar] [CrossRef] [Green Version]
- Bourgon, R.; Gentleman, R.; Huber, W. Reply to Talloen et al.: Independent filtering is a generic approach that needs domain specific adaptation. Proc. Natl. Acad. Sci. USA 2010, 107, E175. [Google Scholar] [CrossRef] [Green Version]
- Mi, H.; Huang, X.; Muruganujan, A.; Tang, H.; Mills, C.; Kang, D.; Thomas, P.D. PANTHER version 11: Expanded annotation data from Gene Ontology and Reactome pathways, and data analysis tool enhancements. Nucleic Acids Res. 2016, 45, D183–D189. [Google Scholar] [CrossRef] [Green Version]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene Ontology: Tool for the unification of biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [Green Version]
- Fabregat, A.; Sidiropoulos, K.; Garapati, P.; Gillespie, M.; Hausmann, K.; Haw, R.; Jassal, B.; Jupe, S.; Korninger, F.; Mckay, S.J.; et al. The Reactome Pathway Knowledgebase. Nucleic Acids Res. 2018, 46, D649–D655. [Google Scholar] [CrossRef]
- Edgar, R.; Domrachev, M.; Lash, A.E. Gene Expression Omnibus: NCBI gene expression and hybridization array data repository. Nucleic Acids Res. 2002, 30, 207–210. [Google Scholar] [CrossRef] [Green Version]
- Kimura, Y.; Yanagimachi, R. Mouse oocytes injected with testicular spermatozoa or round spermatids can develop into normal offspring. Development 1995, 121, 2397–2405. [Google Scholar] [PubMed]
- Yoshida, N.; Perry, A.C. Piezo-actuated mouse intracytoplasmic sperm injection (ICSI). Nat. Protoc. 2007, 2, 296–304. [Google Scholar] [CrossRef] [PubMed]
- Scavizzi, F.; Ryder, E.; Newman, S.; Raspa, M.; Gleeson, D.; Wardle-Jones, H.; Montoliu, L.; Fernández, A.; Dessain, M.L.; Larrigaldie, V.; et al. Blastocyst genotyping for quality control of mouse mutant archives: An ethical and economical approach. Transgenic Res. 2015, 24, 921–927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stein, K.K.; Go, J.C.; Lane, W.S.; Primàkoff, P.; Myles, D.G. Proteomic analysis of sperm regions that mediate sperm-egg interactions. Proteomics 2006, 6, 3533–3543. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.; Gerton, G.L.; Moss, S.B. Proteomic Profiling of Accessory Structures from the Mouse Sperm Flagellum. Mol. Cell. Proteom. 2006, 5, 801–810. [Google Scholar] [CrossRef] [Green Version]
- Baker, M.A.; Hetherington, L.; Reeves, G.M.; Aitken, R.J. The mouse sperm proteome characterized via IPG strip prefractionation and LC-MS/MS identification. Proteomics 2008, 8, 1720–1730. [Google Scholar] [CrossRef]
- Dorus, S.; Wasbrough, E.R.; Busby, J.; Wilkin, E.C.; Karr, T.L. Sperm Proteomics Reveals Intensified Selection on Mouse Sperm Membrane and Acrosome Genes. Mol. Boil. Evol. 2010, 27, 1235–1246. [Google Scholar] [CrossRef] [Green Version]
- Castillo, J.; Jodar, M.; Oliva, R. The contribution of human sperm proteins to the development and epigenome of the preimplantation embryo. Hum. Reprod. Update 2018, 24, 535–555. [Google Scholar] [CrossRef] [Green Version]
- Cao, W.; Aghajanian, H.K.; Haig-Ladewig, L.A.; Gerton, G.L. Sorbitol can fuel mouse sperm motility and protein tyrosine phosphorylation via sorbitol dehydrogenase. Biol. Reprod. 2009, 80, 124–133. [Google Scholar] [CrossRef] [Green Version]
- Klemm, U.; Müller-Esterl, W.; Engel, W. Acrosin, the peculiar sperm-specific serine protease. Qual. Life Res. 1991, 87, 635–641. [Google Scholar] [CrossRef]
- Ryu, D.Y.; Kim, K.U.; Kwon, W.S.; Rahman, S.; Khatun, A.; Pang, M.G. Peroxiredoxin activity is a major landmark of male fertility. Sci. Rep. 2017, 7, 17174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garratt, M.; Bathgate, R.; De Graaf, S.; Brooks, R.C. Copper-zinc superoxide dismutase deficiency impairs sperm motility and in vivo fertility. Reproduction 2013, 146, 297–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walbrecq, G.; Wang, B.; Becker, S.; Hannotiau, A.; Fransen, M.; Knoops, B. Antioxidant cytoprotection by peroxisomal peroxiredoxin-5. Free Radic. Boil. Med. 2015, 84, 215–226. [Google Scholar] [CrossRef] [PubMed]
- Carrell, D.T.; Emery, B.R.; Hammoud, S. Altered protamine expression and diminished spermatogenesis: What is the link? Hum. Reprod. Update 2007, 13, 313–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tremellen, K.P. Oxidative stress and male infertility—A clinical perspective. Hum. Reprod. Update 2008, 14, 243–258. [Google Scholar] [CrossRef] [PubMed]
- Aitken, R.J. Free radicals, lipid peroxidation and sperm function. Reprod. Fertil. Dev. 1995, 7, 659–668. [Google Scholar] [CrossRef]
- Aitken, R.J.; Jones, K.; Robertson, S.A. Reactive Oxygen Species and Sperm Function--In Sickness and In Health. J. Androl. 2012, 33, 1096–1106. [Google Scholar] [CrossRef]
- Alvarez, J.G.; Touchstone, J.C.; Blasco, L.; Storey, B.T. Spontaneous Lipid Peroxidation and Production of Hydrogen Peroxide and Superoxide in Human Spermatozoa Superoxide Dismutase as Major Enzyme Protectant Against Oxygen Toxicity. J. Androl. 1987, 8, 338–348. [Google Scholar] [CrossRef]
- Cho, C.; Jung-Ha, H.; Willis, W.D.; Goulding, E.H.; Stein, P.; Xu, Z.; Schultz, R.M.; Hecht, N.B.; Eddy, E.M. Protamine 2 Deficiency Leads to Sperm DNA Damage and Embryo Death in Mice1. Boil. Reprod. 2003, 69, 211–217. [Google Scholar] [CrossRef] [Green Version]
- Champroux, A.; Damon-Soubeyrand, C.; Goubely, C.; Bravard, S.; Henry-Berger, J.; Guiton, R.; Saez, F.; Drevet, J.R.; Kocer, A. Nuclear Integrity but Not Topology of Mouse Sperm Chromosome is Affected by Oxidative DNA Damage. Genes 2018, 9, 501. [Google Scholar] [CrossRef] [Green Version]
- Ho, Y.S.; Gargano, M.; Cao, J.; Bronson, R.T.; Heimler, I.; Hutz, R.J. Reduced Fertility in Female Mice Lacking Copper-Zinc Superoxide Dismutase. J. Boil. Chem. 1998, 273, 7765–7769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsunoda, S.; Kawano, N.; Miyado, K.; Kimura, N.; Fujii, J. Impaired Fertilizing Ability of Superoxide Dismutase 1-Deficient Mouse Sperm During In Vitro Fertilization1. Boil. Reprod. 2012, 87, 121. [Google Scholar] [CrossRef] [PubMed]
- Murawski, M.; Saczko, J.; Marcinkowska, A.; Chwiłkowska, A.; Gryboś, M.; Banaś, T. Evaluation of superoxide dismutase activity and its impact on semen quality parameters of infertile men. Folia Histochem. Cytobiol. 2007, 45, 23–126. [Google Scholar]
- Yan, L.; Liu, J.; Wu, S.; Zhang, S.; Ji, G.; Gu, A.H. Seminal superoxide dismutase activity and its relationship with semen quality and SOD gene polymorphism. J. Assist. Reprod. Genet. 2014, 31, 549–554. [Google Scholar] [CrossRef] [Green Version]
- Wood, Z.A.; Schröder, E.; Harris, J.R.; Poole, L.B. Structure, mechanism and regulation of peroxiredoxins. Trends Biochem. Sci. 2003, 28, 32–40. [Google Scholar] [CrossRef]
- O’Flaherty, C.; De Souza, A.R. Hydrogen peroxide modifies human sperm peroxiredoxins in a dose-dependent manner. Boil. Reprod. 2010, 84, 238–247. [Google Scholar] [CrossRef] [Green Version]
- Gong, S.; Gabriel, M.C.S.; Zini, A.; Chan, P.; O’Flaherty, C. Low Amounts and High Thiol Oxidation of Peroxiredoxins in Spermatozoa from Infertile Men. J. Androl. 2012, 33, 1342–1351. [Google Scholar] [CrossRef]
- Dias, G.; Retamal, C.; Tobella, L.; Arnholdt, A.; López, M. Nuclear status of immature and mature stallion spermatozoa. Theriogenology 2006, 66, 354–365. [Google Scholar] [CrossRef]
- Ding, F.; Dokholyan, N.V. Dynamical roles of metal ions and the disulfide bond in Cu, Zn superoxide dismutase folding and aggregation. Proc. Natl. Acad. Sci. USA 2008, 105, 19696–19701. [Google Scholar] [CrossRef] [Green Version]
- Miao, L.; Clair, D.K.S. Regulation of superoxide dismutase genes: Implications in disease. Free Radic. Boil. Med. 2009, 47, 344–356. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.; Finelli, R.; Agarwal, A.; Henkel, R. Total antioxidant capacity-Relevance, methods and clinical implications. Andrologia 2020, 2, e13624. [Google Scholar] [CrossRef] [PubMed]
- Guiton, R.; Henry-Berger, J.; Drevet, J.R. The immunobiology of the mammalian epididymis: The black box is now open. Basic Clin. Androl. 2013, 23, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wirleitner, B.; Vanderzwalmen, P.; Stecher, A.; Spitzer, D.; Schuff, M.; Schwerda, D.; Bach, M.; Schechinger, B.; Zech, N.H. Dietary Supplementation of Antioxidants Improves Semen Quality of IVF Patients in Terms of Motility, Sperm Count, and Nuclear Vacuolization. Int. J. Vitam. Nutr. Res. 2012, 82, 391–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.E.; Zhang, Y.; Unni, E.; Shirley, C.R.; Deng, J.M.; Russell, L.D.; Weil, M.M.; Behringer, R.R.; Meistrich, M.L. Abnormal spermatogenesis and reduced fertility in transition nuclear protein 1-deficient mice. Proc. Natl. Acad. Sci. USA 2000, 97, 4683–4688. [Google Scholar] [CrossRef] [Green Version]
- Adham, I.M.; Nayernia, K.; Burkhardt-Göttges, E.; Topaloglu, O.; Dixkens, C.; Holstein, A.F.; Engel, W. Teratozoospermia in mice lacking the transition protein 2 (Tnp2). Mol. Hum. Reprod. 2001, 7, 513–520. [Google Scholar] [CrossRef] [Green Version]
- Suganuma, R.; Yanagimachi, R.; Meistrich, M.L. Decline in fertility of mouse sperm with abnormal chromatin during epididymal passage as revealed by ICSI. Hum. Reprod. 2005, 20, 3101–3108. [Google Scholar] [CrossRef] [Green Version]
- Rogenhofer, N.; Ott, J.; Pilatz, A.; Wolf, J.; Thaler, C.J.; Windischbauer, L.; Schagdarsurengin, U.; Steger, K.; von Schonfeldt, V. Unexplained recurrent miscarriages are associated with an aberrant sperm protamine mRNA content. Hum. Reprod. 2017, 32, 1574–1582. [Google Scholar] [CrossRef]
- Sarasa, J.; Enciso, M.; García, L.; Leza, A.; Steger, K.; Aizpurua, J. Comparison of ART outcomes in men with altered mRNA protamine 1/protamine 2 ratio undergoing intracytoplasmic sperm injection with ejaculated and testicular spermatozoa. Asian J. Androl. 2020. [Google Scholar] [CrossRef]
- Kin, T.; Ono, Y. Idiographica: A general-purpose web application to build idiograms on-demand for human, mouse and rat. Bioinformatics 2007, 23, 2945–2946. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Company | Product-No. | Dilution (IB) |
|---|---|---|---|
| SOD1 | Abcam, Cambridge, UK | ab16831 | 1:1000 |
| PRDX5 | Abcam, Cambridge, UK | ab231892 | 1:500 |
| α-tubulin | Santa Cruz Biotechnology, Dallas, TX, USA | sc-8035 | 1:500 |
| β-actin | Sigma, St. Louis, MO, USA | A5441 | 1:10000 |
| anti-mouse HRP | Dako, Glostrup, Denmark | P0260 | 1:1000 |
| anti-rabbit HRP | Dako, Glostrup, Denmark | P0448 | 1:2000 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schneider, S.; Shakeri, F.; Trötschel, C.; Arévalo, L.; Kruse, A.; Buness, A.; Poetsch, A.; Steger, K.; Schorle, H. Protamine-2 Deficiency Initiates a Reactive Oxygen Species (ROS)-Mediated Destruction Cascade during Epididymal Sperm Maturation in Mice. Cells 2020, 9, 1789. https://doi.org/10.3390/cells9081789
Schneider S, Shakeri F, Trötschel C, Arévalo L, Kruse A, Buness A, Poetsch A, Steger K, Schorle H. Protamine-2 Deficiency Initiates a Reactive Oxygen Species (ROS)-Mediated Destruction Cascade during Epididymal Sperm Maturation in Mice. Cells. 2020; 9(8):1789. https://doi.org/10.3390/cells9081789
Chicago/Turabian StyleSchneider, Simon, Farhad Shakeri, Christian Trötschel, Lena Arévalo, Alexander Kruse, Andreas Buness, Ansgar Poetsch, Klaus Steger, and Hubert Schorle. 2020. "Protamine-2 Deficiency Initiates a Reactive Oxygen Species (ROS)-Mediated Destruction Cascade during Epididymal Sperm Maturation in Mice" Cells 9, no. 8: 1789. https://doi.org/10.3390/cells9081789