Transthyretin Maintains Muscle Homeostasis through the Novel Shuttle Pathway of Thyroid Hormones during Myoblast Differentiation

 ,
,  ,
,
Abstract
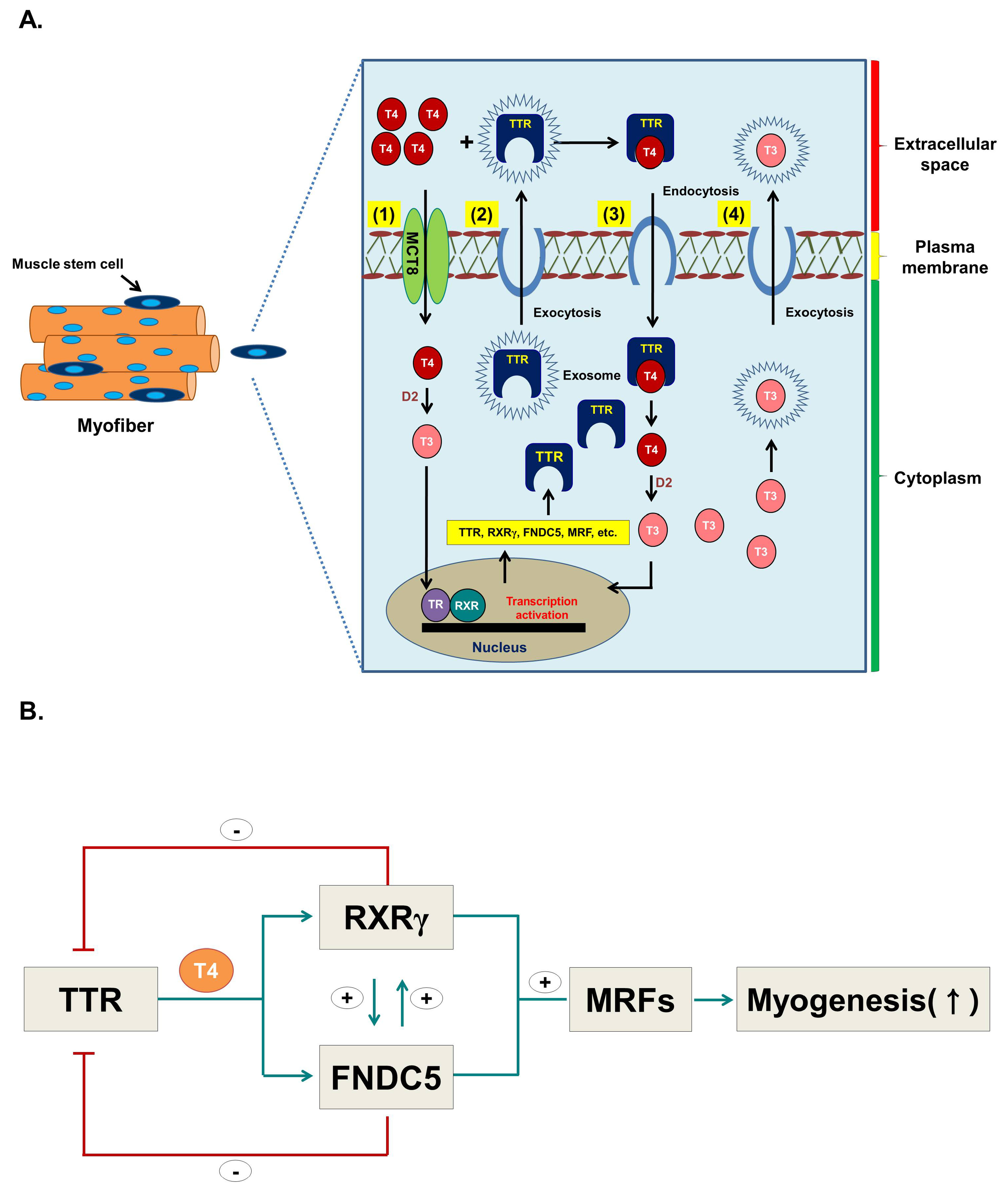
:1. Introduction
2. Materials and Methods
2.1. Animal Experiments
2.2. C2C12 Cell Culture
2.3. Mouse MSCs Culture
2.4. MTT Assay
2.5. Immunoneutralization
2.6. Exosomes Isolation
2.7. T4 and T3 Concentration Measurement
2.8. Gene Knockdown
2.9. RNA Isolation, cDNA Synthesis and RealTime RT-PCR
2.10. RT-PCR
2.11. Protein Isolation from Culture Media
2.12. Western Blot
2.13. Fusion Index
2.14. TTR Protein Labeling with Fluorescence
2.15. TTR Overexpression Vector
2.16. Immunocytochemistry
2.17. Immunohistochemistry
2.18. Microarray Analysis
2.19. DAVID Analysis
2.20. Statistical Analysis
3. Results
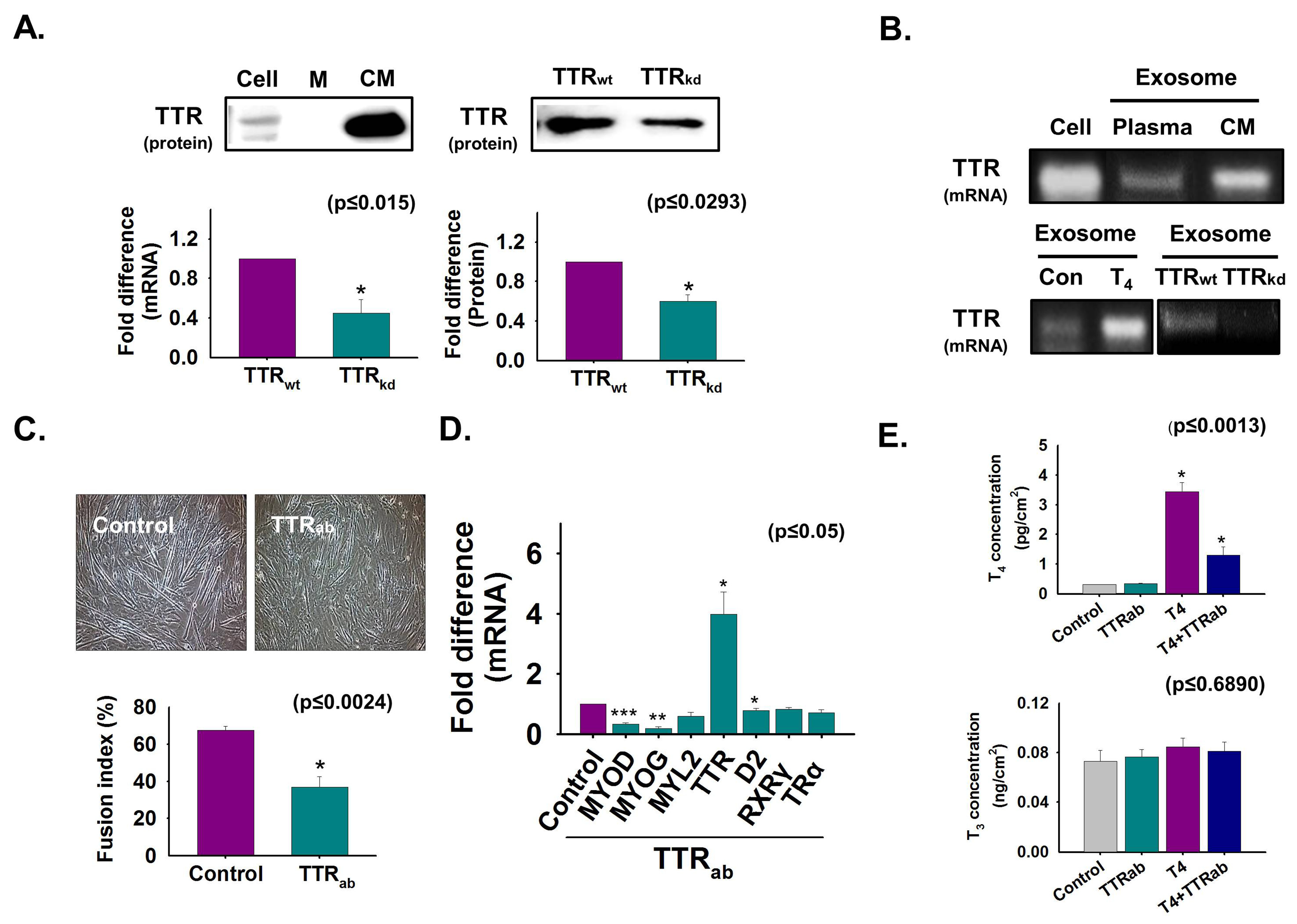
3.1. TTR Secretion During Myoblast Differentiation
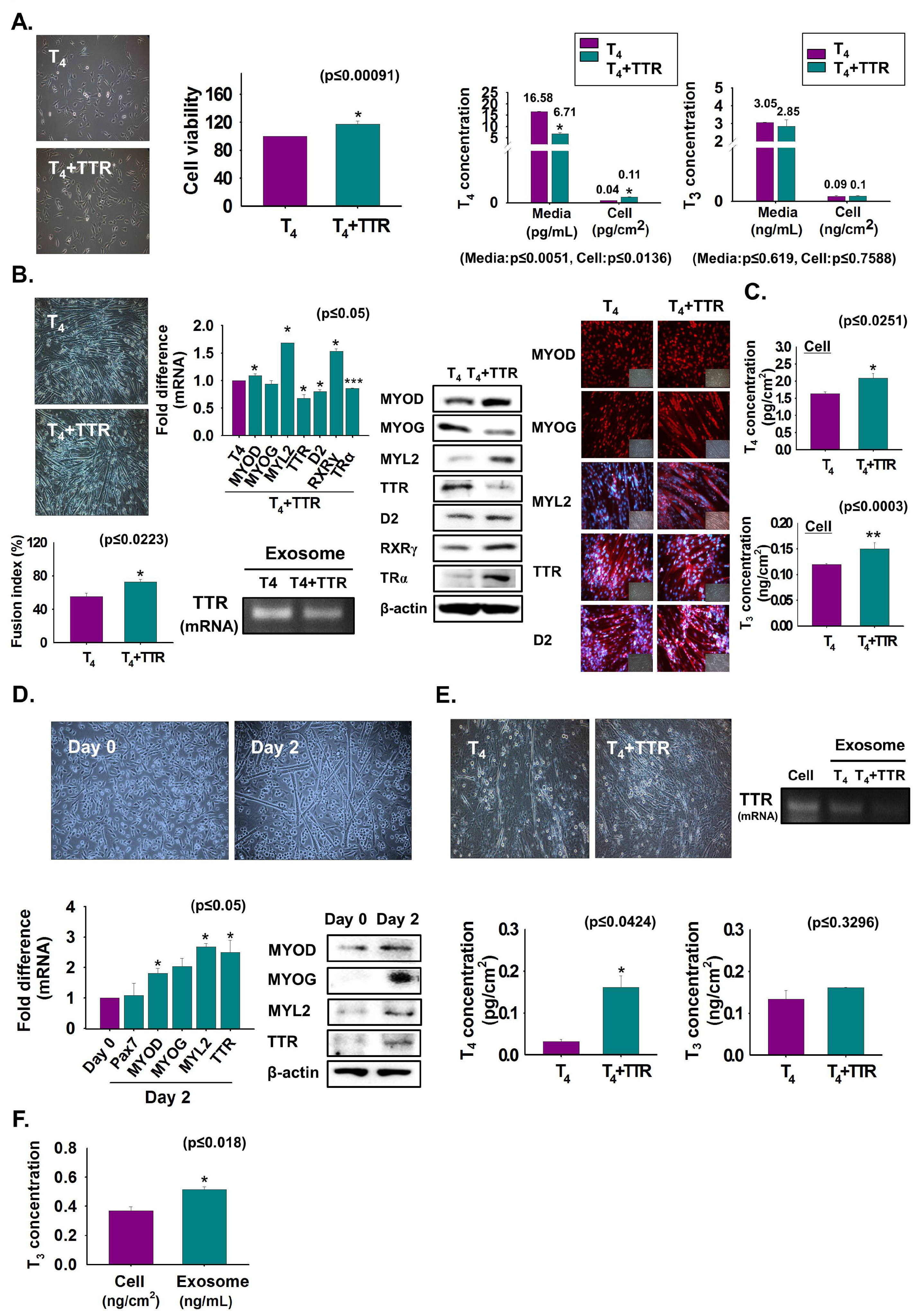
3.2. Enhancement of Myoblast Viability and Differentiation by TTR with T4
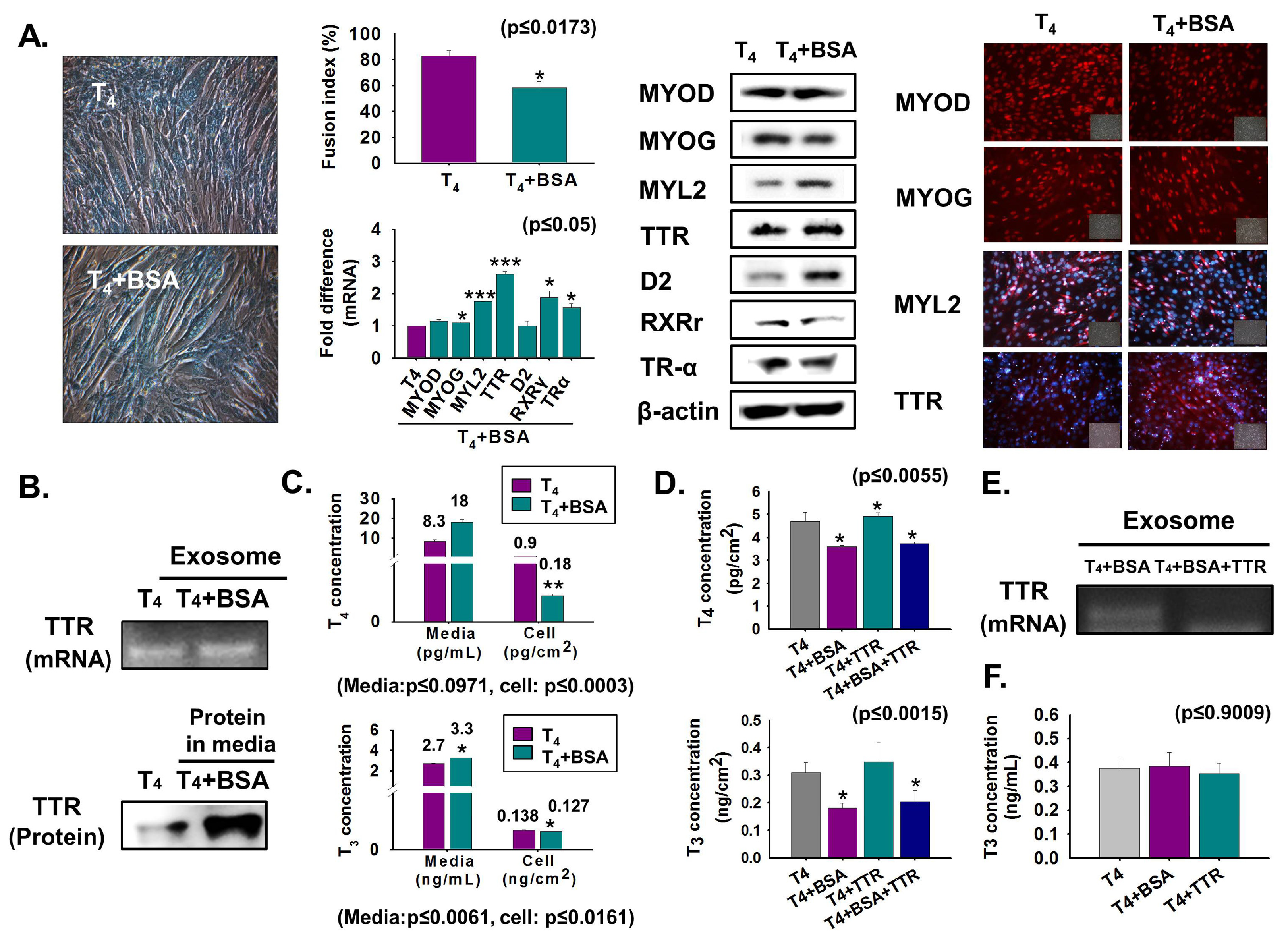
3.3. Reduction of T4 Concentration Inside Cells and Myoblast Differentiation by Bovine Albumin Serum (BSA) Treatment
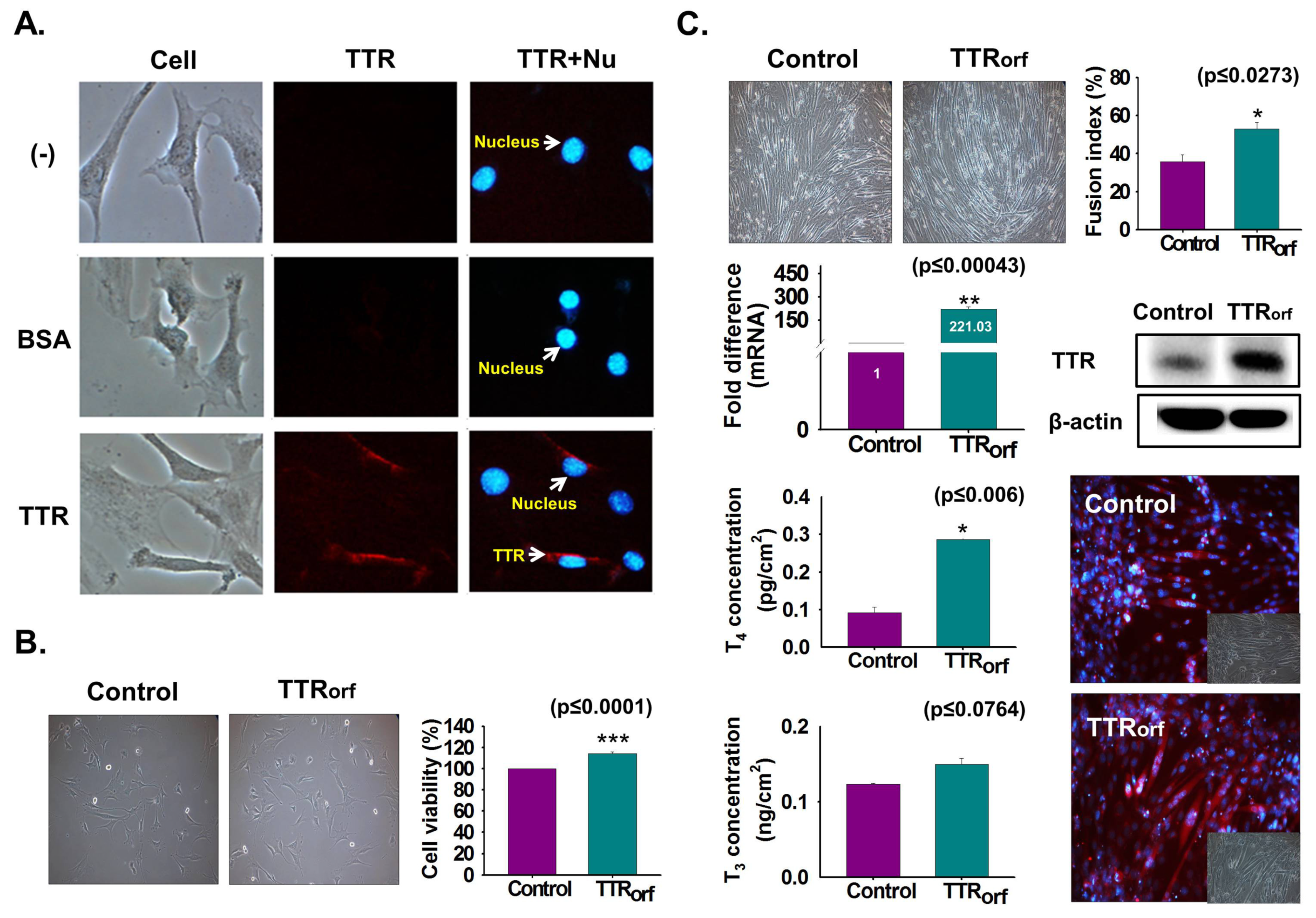
3.4. TTR Internalization Into Myoblast
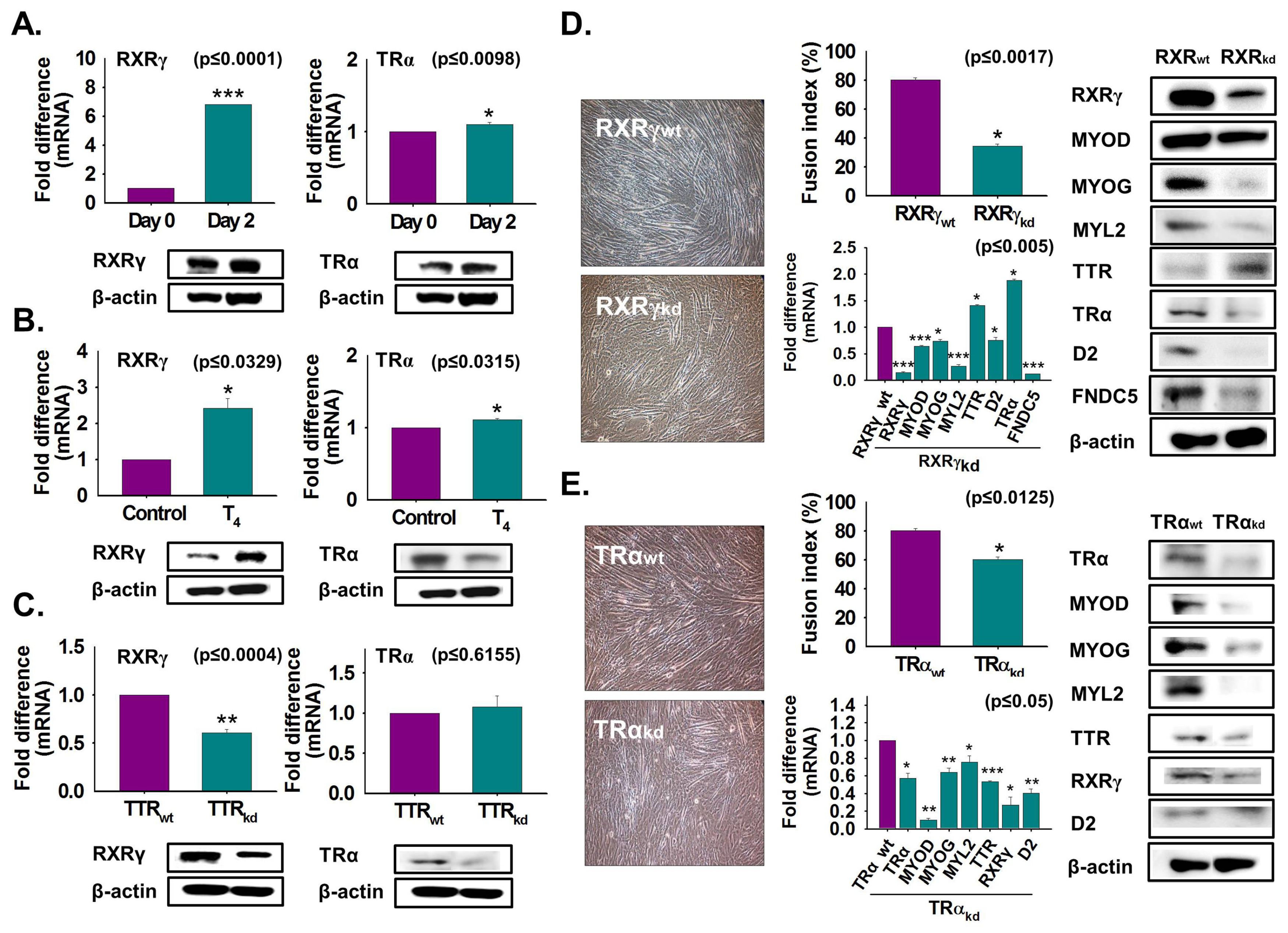
3.5. Regulation of RXRγ and TRα Expression by TTR During Myoblast Differentiation
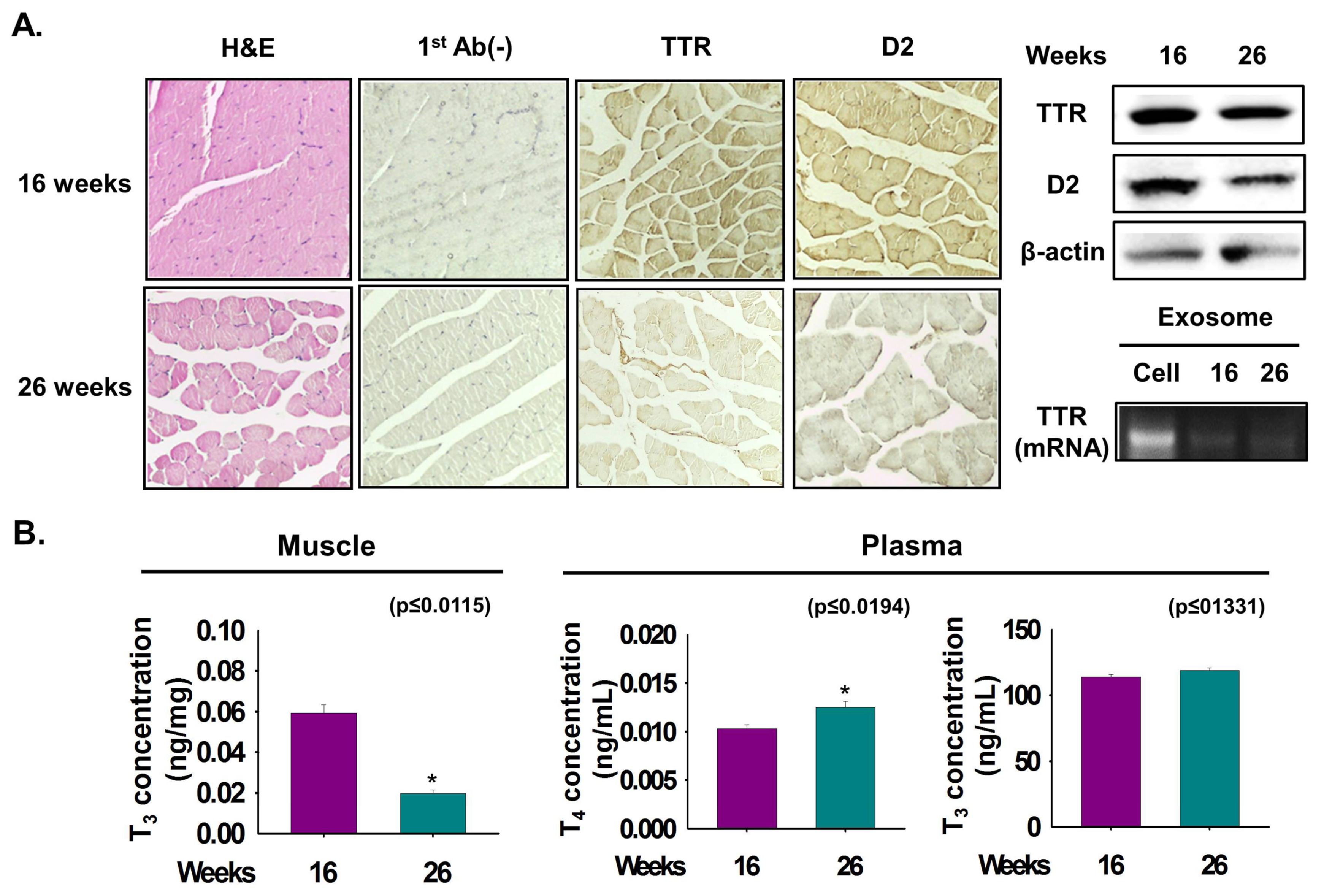
3.6. Relationship between TTR and D2 According to Muscle Age
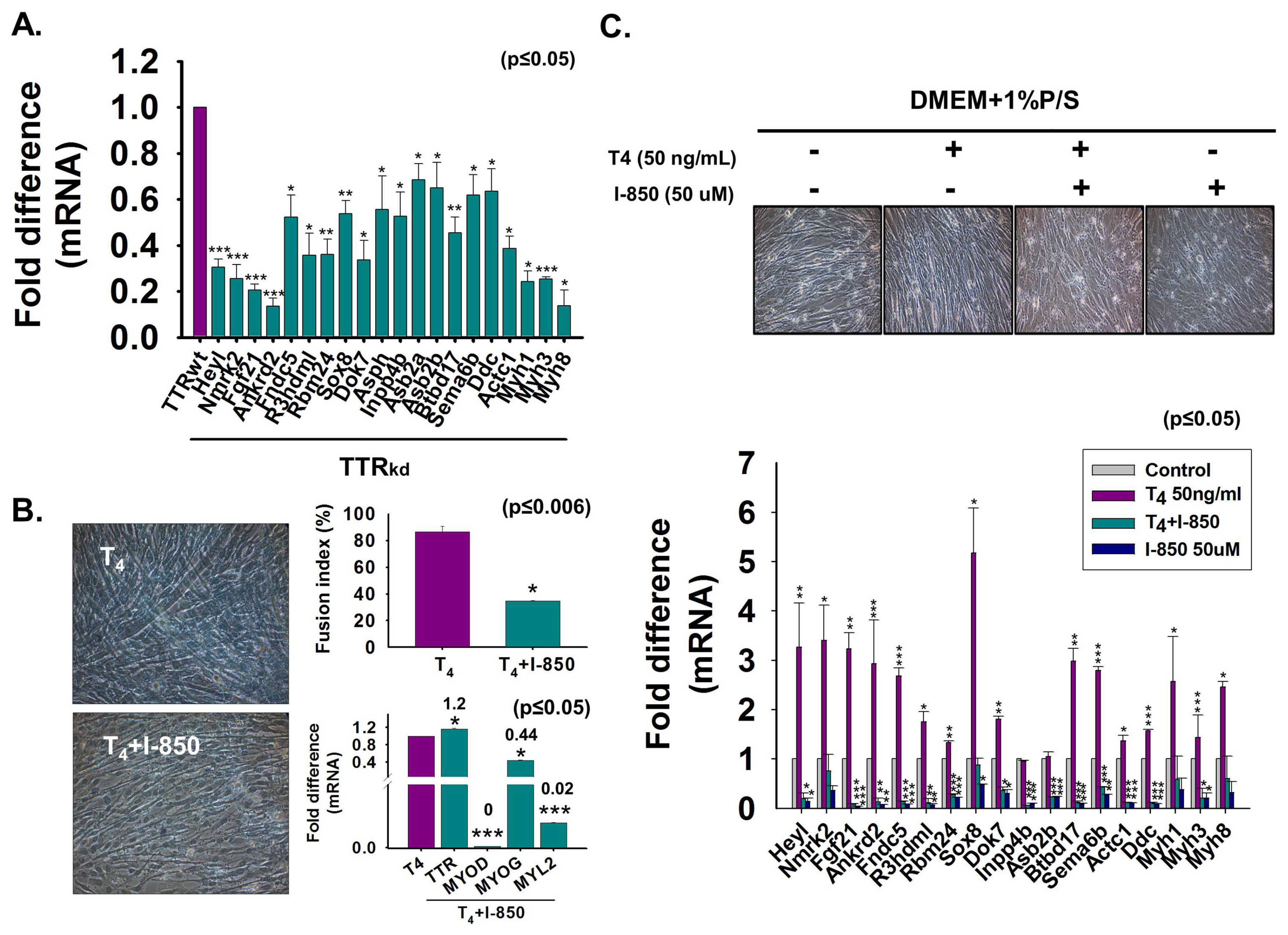
3.7. Microarray Assessment of Gene Expression in TTRkd Cells and Effect of T4 on Gene Expression
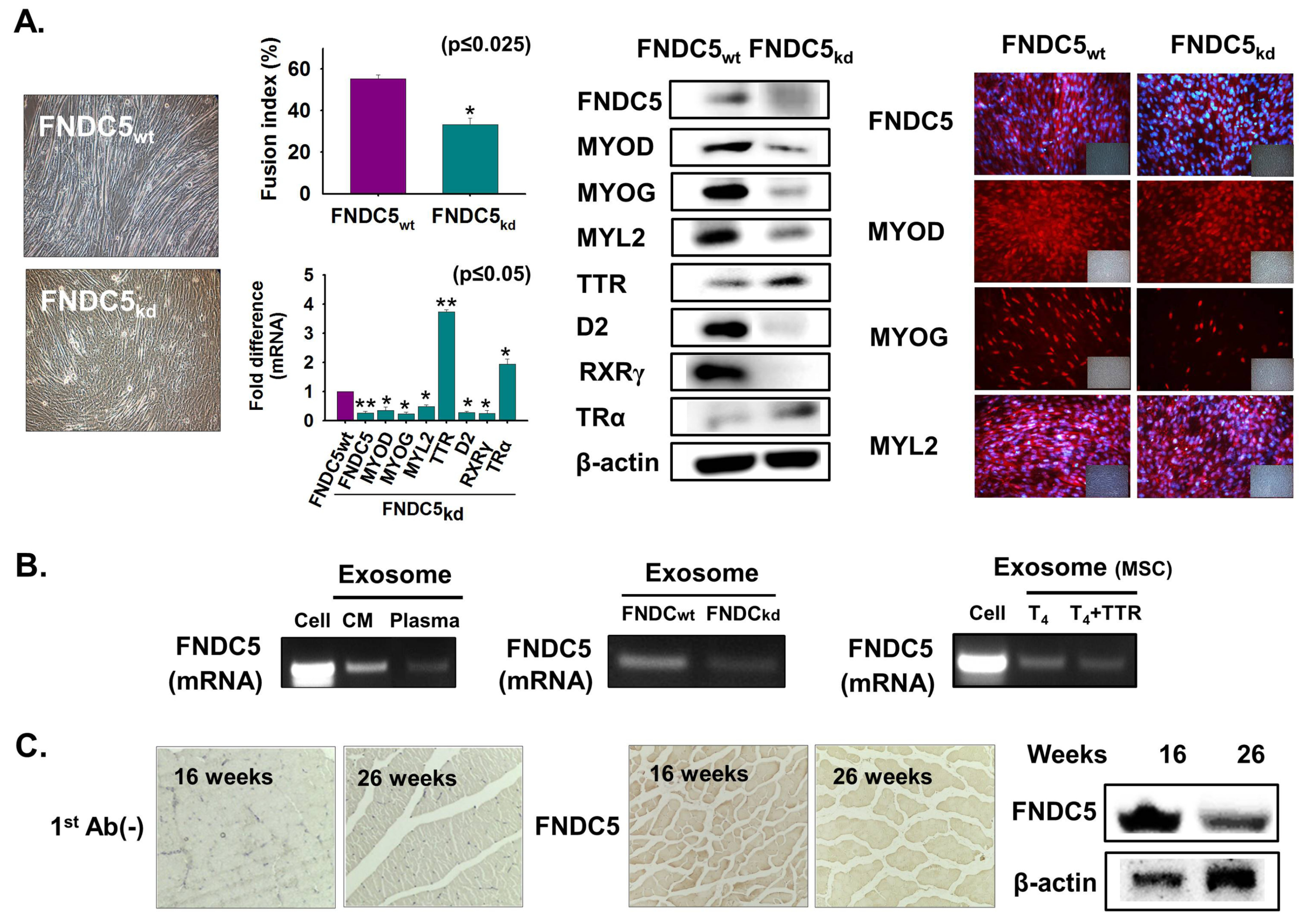
3.8. FNDC5 Expression During Myoblast Differentiation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| TTR | Transthyretin |
| RXR | Retinoid X receptor |
| MSCs | Muscle satellite cells |
| MYOG | Myogenin |
| T4 | Thyroxin |
| T3 | Triiodothyronine |
| THs | Thyroid hormones |
| TR | Thyroid hormone receptors |
| MYOD | Myoblast determination protein |
| TBG | Thyroxine binding globulin |
| FBS | Fetal bovine serum |
| P/S | Penicillin/Streptomycin |
| D2 | Iodothyronine deiodinase type 2 |
| TRE | Thyroid hormone response elements |
| Mct8 | Monocarboxylate transporter 8 |
| TSS | Transcription start site |
| CSF | Cerebrospinal fluid |
| BSA | Bovine serum albumin |
| FNDC5 | Fibronectin type III domain containing 5 |
| CM | Culture media MYL2 (Myosin light chain 2) |
References
- Blau, H.M.; Cosgrove, B.D.; Ho, A.T. The central role of muscle stem cells in regenerative failure with aging. Nat. Med. 2015, 21, 854–862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, K.; Lee, E.J.; Moon, J.S.; Park, S.Y.; Choi, I. Multifaceted Interweaving between Extracellular Matrix, Insulin Resistance, and Skeletal Muscle. Cells 2018, 8, 332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baig, M.H.; Jan, A.T.; Rabbani, G.; Ahmad, K.; Ashraf, J.M.; Kim, T.; Min, H.S.; Lee, Y.H.; Cho, W.K.; Ma, J.Y.; et al. Methylglyoxal and Advanced Glycation End products: Insight of the regulatory machinery affecting the myogenic program and of its modulation by natural compounds. Sci. Rep. 2017, 7, 5916. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Zhang, Y.; Gu, L.; Lan, M.; Liu, C.; Wang, M.; Su, Y.; Ge, M.; Wang, T.; Yu, Y.; et al. Islr regulates canonical Wnt signaling-mediated skeletal muscle regeneration by stabilizing Dishevelled-2 and preventing autophagy. Nat. Commun. 2018, 9, 5129. [Google Scholar] [CrossRef] [Green Version]
- Lee, E.J.; Jan, A.T.; Baig, M.H.; Ashraf, J.M.; Nahm, S.S.; Kim, Y.W.; Park, S.Y.; Choi, I. Fibromodulin: A master regulator of myostatin controlling progression of satellite cells through a myogenic program. FASEB J. 2016, 30, 2708–2719. [Google Scholar] [CrossRef]
- Lee, E.J.; Bhat, A.R.; Kamli, M.R.; Pokharel, S.; Chun, T.; Lee, Y.H.; Nahm, S.S.; Nam, J.H.; Hong, S.K.; Yang, B.; et al. Transthyretin is a key regulator of myoblast differentiation. PLoS ONE 2013, 8, e63627. [Google Scholar] [CrossRef]
- Mishra, A.; Zhu, X.G.; Ge, K.; Cheng, S.Y. Adipogenesis is differentially impaired by thyroid hormone receptor mutant isoforms. J. Mol. Endocrinol. 2010, 44, 247–255. [Google Scholar] [CrossRef] [Green Version]
- Ambrosio, R.; De Stefano, M.A.; Di Girolamo, D.; Salvatore, D. Thyroid hormone signaling and deiodinase actions in muscle stem/progenitor cells. Mol. Cell Endocrinol. 2017, 459, 79–83. [Google Scholar] [CrossRef]
- Milanesi, A.; Lee, J.W.; Yang, A.; Liu, Y.Y.; Sedrakyan, S.; Cheng, S.Y.; Perin, L.; Brent, G.A. Thyroid Hormone Receptor Alpha is Essential to Maintain the Satellite Cell Niche During Skeletal Muscle Injury and Sarcopenia of Aging. Thyroid 2017, 27, 1316–1322. [Google Scholar] [CrossRef]
- Soukup, T.; Smerdu, V. Effect of altered innervation and thyroid hormones on myosin heavy chain expression and fiber type transitions: A mini-review. Histochem. Cell Biol. 2015, 143, 123–130. [Google Scholar] [CrossRef]
- Salvatore, D.; Simonides, W.S.; Dentice, M.; Zavacki, A.M.; Larsen, P.R. Thyroid hormones and skeletal muscle--new insights and potential implications. Nat. Rev. Endocrinol. 2014, 10, 206–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simonides, W.S.; Brent, G.A.; Thelen, M.H.; van der Linden, C.G.; Larsen, P.R.; van Hardeveld, C. Characterization of the promoter of the rat sarcoplasmic endoplasmic reticulum Ca2+-ATPase 1 gene and analysis of thyroid hormone responsiveness. J. Biol. Chem. 1996, 271, 32048–32056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brent, G.A. Mechanisms of thyroid hormone action. J. Clin. Investig. 2012, 122, 3035–3043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazar, M.A. Thyroid hormone receptors: Multiple forms, multiple possibilities. Endocr. Rev. 1993, 14, 184–193. [Google Scholar] [PubMed]
- Weiss, R.E.; Murata, Y.; Cua, K.; Hayashi, Y.; Seo, H.; Refetoff, S. Thyroid hormone action on liver, heart, and energy expenditure in thyroid hormone receptor beta-deficient mice. Endocrinology 1998, 139, 4945–4952. [Google Scholar] [CrossRef] [PubMed]
- Muscat, G.E.; Mynett-Johnson, L.; Dowhan, D.; Downes, M.; Griggs, R. Activation of myoD gene transcription by 3,5,3’-triiodo-L-thyronine: A direct role for the thyroid hormone and retinoid X receptors. Nucleic Acids Res. 1994, 22, 583–591. [Google Scholar] [CrossRef] [Green Version]
- Leid, M.; Kastner, P.; Lyons, R.; Nakshatri, H.; Saunders, M.; Zacharewski, T.; Chen, J.Y.; Staub, A.; Garnier, J.A.; Mader, S.; et al. Purification, cloning, and RXR identity of the HeLa cell factor with which RAR or TR heterodimerizes to bind target sequences efficiently. Cell 1992, 68, 377–395. [Google Scholar] [CrossRef]
- Mangelsdorf, D.J.; Borgmeyer, U.; Heyman, R.A.; Zhou, J.Y.; Ong, E.S.; Oro, A.E.; Kakizuka, A.; Evans, R.M. Characterization of three RXR genes that mediate the action of 9-cis retinoic acid. Genes Dev. 1992, 6, 329–344. [Google Scholar] [CrossRef] [Green Version]
- Simpson, R.J.; Jensen, S.S.; Lim, J.W. Proteomic profiling of exosomes: Current perspectives. Proteomics 2008, 8, 4083–4099. [Google Scholar] [CrossRef]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [Green Version]
- Jan, A.T.; Malik, M.A.; Rahman, S.; Yeo, H.R.; Lee, E.J.; Abdullah, T.S.; Choi, I. Perspective Insights of Exosomes in Neurodegenerative Diseases: A Critical Appraisal. Front. Aging Neurosci. 2017, 9, 317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, S.M.; Connelly, S.; Fearns, C.; Powers, E.T.; Kelly, J.W. The transthyretin amyloidoses: From delineating the molecular mechanism of aggregation linked to pathology to a regulatory-agency-approved drug. J. Mol. Biol. 2012, 421, 185–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richardson, S.J. Cell and molecular biology of transthyretin and thyroid hormones. Int. Rev. Cytol. 2007, 258, 137–193. [Google Scholar] [PubMed]
- Monk, J.A.; Sims, N.A.; Dziegielewska, K.M.; Weiss, R.E.; Ramsay, R.G.; Richardson, S.J. Delayed development of specific thyroid hormone-regulated events in transthyretin null mice. Am. J. Physiol. Endocrinol. Metab. 2013, 304, E23–E31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alshehri, B.; D’Souza, D.G.; Lee, J.Y.; Petratos, S.; Richardson, S.J. The diversity of mechanisms influenced by transthyretin in neurobiology: Development, disease and endocrine disruption. J. Neuroendocrinol. 2015, 27, 303–323. [Google Scholar] [CrossRef] [PubMed]
- Blaner, W.S.; Bonifacio, M.J.; Feldman, H.D.; Piantedosi, R.; Saraiva, M.J. Studies on the synthesis and secretion of transthyretin by the human hepatoma cell line Hep G2. FEBS Lett. 1991, 287, 193–196. [Google Scholar] [CrossRef] [Green Version]
- Landers, K.A.; McKinnon, B.D.; Li, H.; Subramaniam, V.N.; Mortimer, R.H.; Richard, K. Carrier-mediated thyroid hormone transport into placenta by placental transthyretin. J. Clin. Endocrinol. Metab. 2009, 94, 2610–2616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKinnon, B.; Li, H.; Richard, K.; Mortimer, R. Synthesis of thyroid hormone binding proteins transthyretin and albumin by human trophoblast. J. Clin. Endocrinol. Metab. 2005, 90, 6714–6720. [Google Scholar] [CrossRef] [Green Version]
- Lee, E.J.; Pokharel, S.; Jan, A.T.; Huh, S.; Galope, R.; Lim, J.H.; Lee, D.M.; Choi, S.W.; Nahm, S.S.; Kim, Y.W.; et al. Transthyretin: A Transporter Protein Essential for Proliferation of Myoblast in the Myogenic Program. Int. J. Mol. Sci. 2017, 18, 115. [Google Scholar] [CrossRef] [Green Version]
- Lee, E.J.; Jan, A.T.; Baig, M.H.; Ahmad, K.; Malik, A.; Rabbani, G.; Kim, T.; Lee, I.K.; Lee, Y.H.; Park, S.Y.; et al. Fibromodulin and regulation of the intricate balance between myoblast differentiation to myocytes or adipocyte-like cells. FASEB J. 2018, 32, 768–781. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Freire, M.; Semba, R.D.; Ubaida-Mohien, C.; Fabbri, E.; Scalzo, P.; Hojlund, K.; Dufresne, C.; Lyashkov, A.; Ferrucci, L. The Human Skeletal Muscle Proteome Project: A reappraisal of the current literature. J. Cachexia Sarcopenia Muscle 2017, 8, 5–18. [Google Scholar] [CrossRef] [PubMed]
- Di Cosmo, C.; Liao, X.H.; Ye, H.; Ferrara, A.M.; Weiss, R.E.; Refetoff, S.; Dumitrescu, A.M. Mct8-deficient mice have increased energy expenditure and reduced fat mass that is abrogated by normalization of serum T3 levels. Endocrinology 2013, 154, 4885–4895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kassem, N.A.; Deane, R.; Segal, M.B.; Preston, J.E. Role of transthyretin in thyroxine transfer from cerebrospinal fluid to brain and choroid plexus. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 291, R1310–R1315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuchler-Bopp, S.; Dietrich, J.B.; Zaepfel, M.; Delaunoy, J.P. Receptor-mediated endocytosis of transthyretin by ependymoma cells. Brain Res. 2000, 870, 185–194. [Google Scholar] [CrossRef]
- Chen, R.L.; Kassem, N.A.; Preston, J.E. Dose-dependent transthyretin inhibition of T4 uptake from cerebrospinal fluid in sheep. Neurosci. Lett. 2006, 396, 7–11. [Google Scholar] [CrossRef]
- Palha, J.A. Transthyretin as a thyroid hormone carrier: Function revisited. Clin. Chem. Lab. Med. 2002, 40, 1292–1300. [Google Scholar] [CrossRef]
- Bentzinger, C.F.; Wang, Y.X.; Dumont, N.A.; Rudnicki, M.A. Cellular dynamics in the muscle satellite cell niche. EMBO Rep. 2013, 14, 1062–1072. [Google Scholar] [CrossRef] [Green Version]
- Allen, D.L.; Sartorius, C.A.; Sycuro, L.K.; Leinwand, L.A. Different pathways regulate expression of the skeletal myosin heavy chain genes. J. Biol. Chem. 2001, 276, 43524–43533. [Google Scholar] [CrossRef] [Green Version]
- Bianco, A.C.; Salvatore, D.; Gereben, B.; Berry, M.J.; Larsen, P.R. Biochemistry, cellular and molecular biology, and physiological roles of the iodothyronine selenodeiodinases. Endocr. Rev. 2002, 23, 38–89. [Google Scholar] [CrossRef]
- Dekki, N.; Refai, E.; Holmberg, R.; Kohler, M.; Jornvall, H.; Berggren, P.O.; Juntti-Berggren, L. Transthyretin binds to glucose-regulated proteins and is subjected to endocytosis by the pancreatic beta-cell. Cell Mol. Life Sci. 2012, 69, 1733–1743. [Google Scholar] [CrossRef]
- Sousa, M.M.; Norden, A.G.; Jacobsen, C.; Willnow, T.E.; Christensen, E.I.; Thakker, R.V.; Verroust, P.J.; Moestrup, S.K.; Saraiva, M.J. Evidence for the role of megalin in renal uptake of transthyretin. J. Biol. Chem. 2000, 275, 38176–38181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Divino, C.M.; Schussler, G.C. Transthyretin receptors on human astrocytoma cells. J. Clin. Endocrinol. Metab. 1990, 71, 1265–1268. [Google Scholar] [CrossRef] [PubMed]
- Mariotti, S.; Franceschi, C.; Cossarizza, A.; Pinchera, A. The aging thyroid. Endocr. Rev. 1995, 16, 686–715. [Google Scholar] [CrossRef] [PubMed]
- Silvestri, E.; Lombardi, A.; de Lange, P.; Schiavo, L.; Lanni, A.; Goglia, F.; Visser, T.J.; Moreno, M. Age-related changes in renal and hepatic cellular mechanisms associated with variations in rat serum thyroid hormone levels. Am. J. Physiol. Endocrinol. Metab. 2008, 294, E1160–E1168. [Google Scholar] [CrossRef] [Green Version]
- Savu, L.; Vranckx, R.; Rouaze-Romet, M.; Maya, M.; Nunez, E.A.; Treton, J.; Flink, I.L. A senescence up-regulated protein: The rat thyroxine-binding globulin (TBG). Biochim. Biophys. Acta 1991, 1097, 19–22. [Google Scholar] [CrossRef]
- Jang, M.; DiStefano, J.J., 3rd. Some quantitative changes in iodothyronine distribution and metabolism in mild obesity and aging. Endocrinology 1985, 116, 457–468. [Google Scholar] [CrossRef]
- Van Mullem, A.; van Heerebeek, R.; Chrysis, D.; Visser, E.; Medici, M.; Andrikoula, M.; Tsatsoulis, A.; Peeters, R.; Visser, T.J. Clinical phenotype and mutant TRalpha1. N. Engl. J. Med. 2012, 366, 1451–1453. [Google Scholar] [CrossRef]
- Haugen, B.R.; Jensen, D.R.; Sharma, V.; Pulawa, L.K.; Hays, W.R.; Krezel, W.; Chambon, P.; Eckel, R.H. Retinoid X receptor gamma-deficient mice have increased skeletal muscle lipoprotein lipase activity and less weight gain when fed a high-fat diet. Endocrinology 2004, 145, 3679–3685. [Google Scholar] [CrossRef] [Green Version]
- Hartong, R.; Wang, N.; Kurokawa, R.; Lazar, M.A.; Glass, C.K.; Apriletti, J.W.; Dillmann, W.H. Delineation of three different thyroid hormone-response elements in promoter of rat sarcoplasmic reticulum Ca2+ATPase gene. Demonstration that retinoid X receptor binds 5′ to thyroid hormone receptor in response element 1. J. Biol. Chem. 1994, 269, 13021–13029. [Google Scholar]
- Solanes, G.; Pedraza, N.; Calvo, V.; Vidal-Puig, A.; Lowell, B.B.; Villarroya, F. Thyroid hormones directly activate the expression of the human and mouse uncoupling protein-3 genes through a thyroid response element in the proximal promoter region. Biochem. J. 2005, 386, 505–513. [Google Scholar] [CrossRef] [Green Version]
- Zorzano, A.; Palacin, M.; Guma, A. Mechanisms regulating GLUT4 glucose transporter expression and glucose transport in skeletal muscle. Acta Physiol. Scand. 2005, 183, 43–58. [Google Scholar] [CrossRef] [PubMed]
- Desvergne, B.; Petty, K.J.; Nikodem, V.M. Functional characterization and receptor binding studies of the malic enzyme thyroid hormone response element. J. Biol. Chem. 1991, 266, 1008–1013. [Google Scholar] [PubMed]
- Dummler, K.; Muller, S.; Seitz, H.J. Regulation of adenine nucleotide translocase and glycerol 3-phosphate dehydrogenase expression by thyroid hormones in different rat tissues. Biochem. J. 1996, 317, 913–918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morkin, E. Control of cardiac myosin heavy chain gene expression. Microsc. Res. Tech. 2000, 50, 522–531. [Google Scholar] [CrossRef]
- Gereben, B.; Zavacki, A.M.; Ribich, S.; Kim, B.W.; Huang, S.A.; Simonides, W.S.; Zeold, A.; Bianco, A.C. Cellular and molecular basis of deiodinase-regulated thyroid hormone signaling. Endocr. Rev. 2008, 29, 898–938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Visser, W.E.; Friesema, E.C.; Visser, T.J. Minireview: Thyroid hormone transporters: The knowns and the unknowns. Mol. Endocrinol. 2011, 25, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friesema, E.C.; Jansen, J.; Jachtenberg, J.W.; Visser, W.E.; Kester, M.H.; Visser, T.J. Effective cellular uptake and efflux of thyroid hormone by human monocarboxylate transporter 10. Mol. Endocrinol. 2008, 22, 1357–1369. [Google Scholar] [CrossRef] [Green Version]
- Marsili, A.; Ramadan, W.; Harney, J.W.; Mulcahey, M.; Castroneves, L.A.; Goemann, I.M.; Wajner, S.M.; Huang, S.A.; Zavacki, A.M.; Maia, A.L.; et al. Type 2 iodothyronine deiodinase levels are higher in slow-twitch than fast-twitch mouse skeletal muscle and are increased in hypothyroidism. Endocrinology 2010, 151, 5952–5960. [Google Scholar] [CrossRef]
- Jin, D.; Hidaka, K.; Shirai, M.; Morisaki, T. RNA-binding motif protein 24 regulates myogenin expression and promotes myogenic differentiation. Genes Cells 2010, 15, 1158–1167. [Google Scholar] [CrossRef]
- Cardinali, B.; Cappella, M.; Provenzano, C.; Garcia-Manteiga, J.M.; Lazarevic, D.; Cittaro, D.; Martelli, F.; Falcone, G. MicroRNA-222 regulates muscle alternative splicing through Rbm24 during differentiation of skeletal muscle cells. Cell Death Dis. 2016, 7, e2086. [Google Scholar] [CrossRef]
- Schmidt, K.; Glaser, G.; Wernig, A.; Wegner, M.; Rosorius, O. Sox8 is a specific marker for muscle satellite cells and inhibits myogenesis. J. Biol. Chem. 2003, 278, 29769–29775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, M.; Murakami, S.; Yoneda, T.; Nakamura, M.; Zhang, L.; Uezumi, A.; Fukuda, S.; Kokubo, H.; Tsujikawa, K.; Fukada, S. Evidence of Notch-Hesr-Nrf2 Axis in Muscle Stem Cells, but Absence of Nrf2 Has No Effect on Their Quiescent and Undifferentiated State. PLoS ONE 2015, 10, e0138517. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Mayne, R.; Wu, C. A novel muscle-specific beta 1 integrin binding protein (MIBP) that modulates myogenic differentiation. J. Cell. Biol. 1999, 147, 1391–1398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, J.L.; Patil, P.B.; Minteer, S.D.; Lipsitz, J.R.; Fisher, J.S. Possibility of autocrine beta-adrenergic signaling in C2C12 myotubes. Exp. Biol. Med. 2005, 230, 845–852. [Google Scholar] [CrossRef] [PubMed]
- Harbers, M.; Wahlstrom, G.M.; Vennstrom, B. Transactivation by the thyroid hormone receptor is dependent on the spacer sequence in hormone response elements containing directly repeated half-sites. Nucleic Acids Res. 1996, 24, 2252–2259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weth, O.; Weth, C.; Bartkuhn, M.; Leers, J.; Uhle, F.; Renkawitz, R. Modular insulators: Genome wide search for composite CTCF/thyroid hormone receptor binding sites. PLoS ONE 2010, 5, e10119. [Google Scholar] [CrossRef] [Green Version]
- Ali, T.; Renkawitz, R.; Bartkuhn, M. Insulators and domains of gene expression. Curr. Opin. Genet. Dev. 2016, 37, 17–26. [Google Scholar] [CrossRef] [Green Version]
- Lutz, M.; Burke, L.J.; LeFevre, P.; Myers, F.A.; Thorne, A.W.; Crane-Robinson, C.; Bonifer, C.; Filippova, G.N.; Lobanenkov, V.; Renkawitz, R. Thyroid hormone-regulated enhancer blocking: Cooperation of CTCF and thyroid hormone receptor. EMBO J. 2003, 22, 1579–1587. [Google Scholar] [CrossRef] [Green Version]
- Hou, C.; Zhao, H.; Tanimoto, K.; Dean, A. CTCF-dependent enhancer-blocking by alternative chromatin loop formation. Proc. Natl. Acad. Sci. USA 2008, 105, 20398–20403. [Google Scholar] [CrossRef] [Green Version]
- Bostrom, P.; Wu, J.; Jedrychowski, M.P.; Korde, A.; Ye, L.; Lo, J.C.; Rasbach, K.A.; Bostrom, E.A.; Choi, J.H.; Long, J.Z.; et al. A PGC1-alpha-dependent myokine that drives brown-fat-like development of white fat and thermogenesis. Nature 2012, 481, 463–468. [Google Scholar] [CrossRef]
- Roca-Rivada, A.; Castelao, C.; Senin, L.L.; Landrove, M.O.; Baltar, J.; Belen Crujeiras, A.; Seoane, L.M.; Casanueva, F.F.; Pardo, M. FNDC5/irisin is not only a myokine but also an adipokine. PLoS ONE 2013, 8, e60563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reza, M.M.; Subramaniyam, N.; Sim, C.M.; Ge, X.; Sathiakumar, D.; McFarlane, C.; Sharma, M.; Kambadur, R. Irisin is a pro-myogenic factor that induces skeletal muscle hypertrophy and rescues denervation-induced atrophy. Nat. Commun. 2017, 8, 1104. [Google Scholar] [CrossRef] [PubMed]
- Huh, J.Y.; Dincer, F.; Mesfum, E.; Mantzoros, C.S. Irisin stimulates muscle growth-related genes and regulates adipocyte differentiation and metabolism in humans. Int. J. Obes. 2014, 38, 1538–1544. [Google Scholar] [CrossRef] [PubMed]
- Jedrychowski, M.P.; Wrann, C.D.; Paulo, J.A.; Gerber, K.K.; Szpyt, J.; Robinson, M.M.; Nair, K.S.; Gygi, S.P.; Spiegelman, B.M. Detection and Quantitation of Circulating Human Irisin by Tandem Mass Spectrometry. Cell Metab. 2015, 22, 734–740. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A. | |||||||||
| Gene | Set1 | Set2 | Set3 | Set4 | Average | p Value | Description | ||
| Myh1 | 0.16 | 0.31 | 0.13 | 0.06 | 0.16 | 0.0001 | Mus musculus myosin, heavy polypeptide 1, skeletal muscle, adult (Myh1) | ||
| Heyl | 0.25 | 0.19 | 0.15 | 0.1 | 0.17 | 0.0001 | Mus musculus hairy/enhancer-of-split related with YRPW motif-like (Heyl) | ||
| Myo18b | 0.17 | 0.4 | 0.13 | 0.05 | 0.19 | 0.0001 | Mus musculus myosin XVIIIb (Myo18b) | ||
| Myh8 | 0.2 | 0.39 | 0.12 | 0.08 | 0.2 | 0.0001 | Mus musculus myosin, heavy polypeptide 8, skeletal muscle, perinatal (Myh8) | ||
| Nmrk2 | 0.16 | 0.37 | 0.06 | 0.22 | 0.2 | 0.0001 | Mus musculus nicotinamide riboside kinase 2 (Nmrk2) | ||
| Fgf21 | 0.14 | 0.35 | 0.06 | 0.32 | 0.22 | 0.0001 | Mus musculus fibroblast growth factor 21 (Fgf21) | ||
| Ankrd2 | 0.17 | 0.36 | 0.06 | 0.28 | 0.22 | 0.0001 | Mus musculusankyrin repeat domain 2 (stretch responsive muscle) (Ankrd2) | ||
| Myom3 | 0.18 | 0.49 | 0.11 | 0.11 | 0.22 | 0.0001 | Mus musculusmyomesin family, member 3 (Myom3) | ||
| Myh3 | 0.18 | 0.39 | 0.14 | 0.21 | 0.23 | 0.0001 | Mus musculus myosin, heavy polypeptide 3, skeletal muscle, embryonic (Myh3) | ||
| Myh7 | 0.3 | 0.33 | 0.12 | 0.2 | 0.24 | 0.0001 | Mus musculus myosin, heavy polypeptide 7, cardiac muscle, beta (Myh7) | ||
| Myh3 | 0.21 | 0.4 | 0.15 | 0.21 | 0.24 | 0.0001 | Mus musculus myosin, heavy polypeptide 3, skeletal muscle, embryonic (Myh3) | ||
| Fndc5 | 0.44 | 0.26 | 0.18 | 0.1 | 0.25 | 0.0001 | Mus musculus fibronectin type III domain containing 5 (Fndc5) | ||
| R3hdml | 0.35 | 0.4 | 0.09 | 0.15 | 0.25 | 0.0001 | Mus musculus R3H domain containing-like (R3hdml) | ||
| Myh7b | 0.31 | 0.39 | 0.07 | 0.22 | 0.25 | 0.0001 | Mus musculus myosin, heavy chain 7B, cardiac muscle, beta (Myh7b) | ||
| Rbm24 | 0.2 | 0.41 | 0.15 | 0.23 | 0.25 | 0.0001 | Mus musculus RNA binding motif protein 24 (Rbm24) | ||
| Sox8 | 0.45 | 0.21 | 0.13 | 0.22 | 0.25 | 0.0001 | Mus musculus SRY (sex determining region Y)-box 8 (Sox8) | ||
| Dok7 | 0.21 | 0.5 | 0.08 | 0.29 | 0.27 | 0.0002 | Mus musculus docking protein 7 (Dok7) | ||
| Tnnt1 | 0.31 | 0.37 | 0.12 | 0.29 | 0.27 | 0.0001 | Mus musculus troponin T1, skeletal, slow (Tnnt1), transcript variant 1 | ||
| Ttn | 0.17 | 0.48 | 0.24 | 0.2 | 0.27 | 0.0001 | Mus musculus titin (Ttn), transcript variant N2-B | ||
| Asph | 0.28 | 0.46 | 0.18 | 0.16 | 0.27 | 0.0001 | Mus musculus aspartate-beta-hydroxylase (Asph), transcript variant 8 | ||
| Inpp4b | 0.2 | 0.41 | 0.33 | 0.16 | 0.27 | 0.0001 | Mus musculus inositol polyphosphate-4-phosphatase, type II (Inpp4b) | ||
| Igf2os | 0.37 | 0.45 | 0.15 | 0.16 | 0.28 | 0.0001 | Mus musculus insulin-like growth factor 2, opposite strand (Igf2os), antisense RNA | ||
| Myh6 | 0.42 | 0.23 | 0.18 | 0.32 | 0.28 | 0.0001 | Mus musculus myosin, heavy polypeptide 6, cardiac muscle, alpha (Myh6) | ||
| Asb2 | 0.48 | 0.39 | 0.23 | 0.17 | 0.32 | 0.0001 | Mus musculusankyrin repeat and SOCS box-containing 2 (Asb2) | ||
| Mybpc1 | 0.45 | 0.44 | 0.25 | 0.13 | 0.32 | 0.0001 | Mus musculus myosin binding protein C, slow-type (Mybpc1) | ||
| Btbd17 | 0.47 | 0.45 | 0.1 | 0.29 | 0.33 | 0.0002 | Mus musculus BTB (POZ) domain containing 17 (Btbd17) | ||
| Sema6b | 0.46 | 0.39 | 0.09 | 0.38 | 0.33 | 0.0002 | Mus musculussema domain, transmembrane domain (TM), and cytoplasmic domain | ||
| Actc1 | 0.38 | 0.39 | 0.33 | 0.25 | 0.34 | 0.0001 | Mus musculus actin, alpha, cardiac muscle 1 (Actc1) | ||
| Ddc | 0.48 | 0.48 | 0.22 | 0.39 | 0.39 | 0.0001 | Mus musculusdopa decarboxylase (Ddc), transcript variant 1 | ||
| B. | |||||||||
| Gene | Set1 | Set2 | Set3 | Set4 | Average | pValue | Description | ||
| Gm10536 | 4.95 | 7.99 | 11.97 | 13.93 | 9.71 | 0.0049 | Mus musculus predicted gene 10536 (Gm10536), long non-coding RNA | ||
| Iws1 | 3.92 | 4.7 | 2.15 | 10.28 | 5.26 | 0.5130 | Mus musculus IWS1 homolog (S. cerevisiae) (Iws1) | ||
| Dkk2 | 4.02 | 3.54 | 4.95 | 2.98 | 3.87 | 0.0005 | Mus musculusdickkopf homolog 2 (Xenopuslaevis) (Dkk2) | ||
| Cdc45 | 3.44 | 2.22 | 4.92 | 3.16 | 3.43 | 0.0048 | Mus musculus cell division cycle 45 (Cdc45), transcript variant 1 | ||
| Suv420h1 | 3.25 | 2.86 | 2.71 | 3.45 | 3.07 | 0.0001 | Mus musculus suppressor of variegation 4–20 homolog 1 (Drosophila) (Suv420h1) | ||
| Cdc42bpa | 2.28 | 2.11 | 3.11 | 4.32 | 2.95 | 0.0082 | Mus musculus CDC42 binding protein kinase alpha (Cdc42bpa) | ||
| Zfp318 | 2.02 | 2.89 | 2.23 | 4.18 | 2.83 | 0.0094 | Mus musculus zinc finger protein 318 (Zfp318), transcript variant 2 | ||
| C. | |||||||||
| Term | Count | % | p Value | ||||||
| Transcription regulation | 3 | 42.9 | 7.5 × 10−2 | ||||||
| Nucleus | 4 | 57.1 | 9.8 × 10−2 | ||||||
| D. | |||||||||
| Term | Count | % | p Value | ||||||
| Muscle protein | 8 | 28.6 | 1.40 × 10−13 | ||||||
| Thick filament | 6 | 21.4 | 1.00 × 10−12 | ||||||
| Myosin | 7 | 25 | 1.40 × 10−11 | ||||||
| Motor protein | 7 | 25 | 5.90 × 10−9 | ||||||
| Actin-binding | 6 | 21.4 | 8.50 × 10−6 | ||||||
| ATP-binding | 10 | 35.7 | 1.20 × 10−5 | ||||||
| Calmodulin-binding | 5 | 17.9 | 1.80 × 10−5 | ||||||
| Methylation | 6 | 21.4 | 6.60 × 10−5 | ||||||
| Nucleotide-binding | 10 | 35.7 | 8.90 × 10−5 | ||||||
| Coiled coil | 11 | 39.3 | 1.40 × 10−3 | ||||||
| Cytoplasm | 10 | 35.7 | 4.80 × 10−2 | ||||||
| Isopeptide bond | 4 | 14.3 | 9.40 × 10−2 | ||||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, E.J.; Shaikh, S.; Choi, D.; Ahmad, K.; Baig, M.H.; Lim, J.H.; Lee, Y.-H.; Park, S.J.; Kim, Y.-W.; Park, S.-Y.; et al. Transthyretin Maintains Muscle Homeostasis through the Novel Shuttle Pathway of Thyroid Hormones during Myoblast Differentiation. Cells 2019, 8, 1565. https://doi.org/10.3390/cells8121565
Lee EJ, Shaikh S, Choi D, Ahmad K, Baig MH, Lim JH, Lee Y-H, Park SJ, Kim Y-W, Park S-Y, et al. Transthyretin Maintains Muscle Homeostasis through the Novel Shuttle Pathway of Thyroid Hormones during Myoblast Differentiation. Cells. 2019; 8(12):1565. https://doi.org/10.3390/cells8121565
Chicago/Turabian StyleLee, Eun Ju, Sibhghatulla Shaikh, Dukhwan Choi, Khurshid Ahmad, Mohammad Hassan Baig, Jeong Ho Lim, Yong-Ho Lee, Sang Joon Park, Yong-Woon Kim, So-Young Park, and et al. 2019. "Transthyretin Maintains Muscle Homeostasis through the Novel Shuttle Pathway of Thyroid Hormones during Myoblast Differentiation" Cells 8, no. 12: 1565. https://doi.org/10.3390/cells8121565