Reductions of Circulating Nitric Oxide are Followed by Hypertension during Pregnancy and Increased Activity of Matrix Metalloproteinases-2 and -9 in Rats


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Design
- Normotensive virgin rats (Virgin group) received daily intraperitoneal (i.p.) injections of saline for 6 consecutive days;
- Hypertensive virgin rats (Virgin+L-NAME group) received daily i.p. injections of L-NAME for 5 consecutive days;
- Pregnant rats were studied according to the different pregnancy stages, which correspond to the three gestational trimesters:
- Normotensive early pregnancy (Early-Preg group): pregnant rats received daily i.p. injections of saline from pregnancy day 3 to 8;
- Hypertensive early pregnancy (Early-Preg+L-NAME group): pregnant rats received daily i.p. injections of L-NAME from pregnancy day 3 to 8;
- Normotensive middle pregnancy (Mid-Preg group): pregnant rats received daily i.p. injections of saline from pregnancy day 9 to 15;
- Hypertensive middle pregnancy (Mid-Preg+L-NAME group): pregnant rats received daily i.p. injections of L-NAME from pregnancy day 9 to 15;
- Normotensive late pregnancy (Late-Preg group): pregnant rats received daily i.p. injections of saline from pregnancy day 13 to 20;
- Hypertensive late pregnancy (Late-Preg+L-NAME group): pregnant rats received daily i.p. injections of L-NAME from pregnancy day 13 to 20.
2.3. Blood Pressure Measurements
2.4. Animal Euthanasia and Tissue Collection
2.5. Fetal and Placental Parameters
2.6. Determination of Plasma Levels of Nitrite/Nitrate (Total NOx)
2.7. Determination of Lipid Peroxidation
2.8. Zymography for MMP-2 and MMP-9 Activity
2.9. Statistical Analysis
3. Results
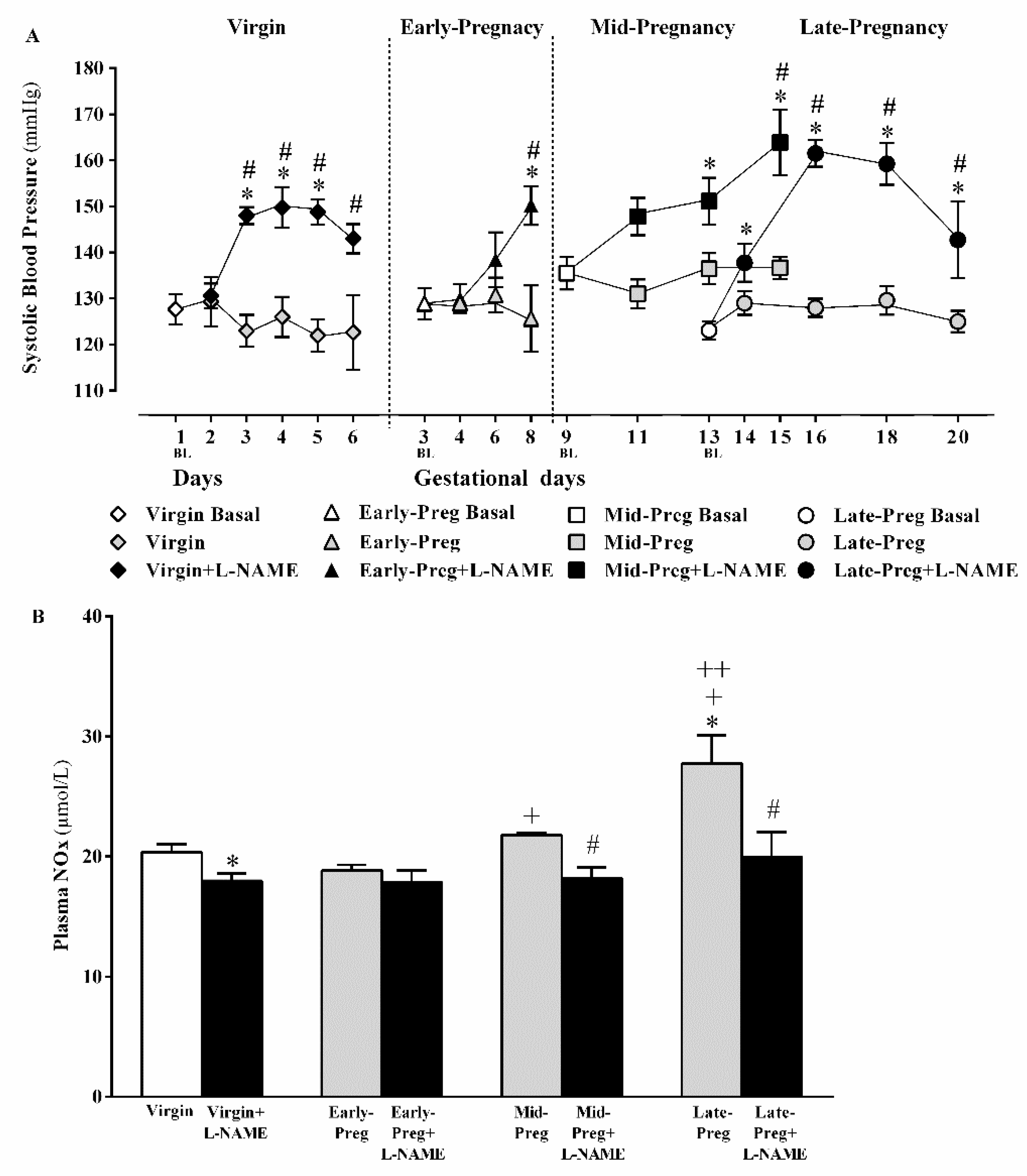
3.1. L-NAME Treatment Presented Hypertnsion in Virgin and Early, Middle and Lat Pregnancy Stages
3.2. Circulating NO Increases in Middle and Late but Not in Early Pregnancy Stage, While Decreases in Circulating NO Are Observed in Virgin Rats and in Middle and Late Pregnant Rtas Treated with L-NAME. But, Early Pregnant Rats Treated (Or Not) with L-Name Presented Similar Circulating NO
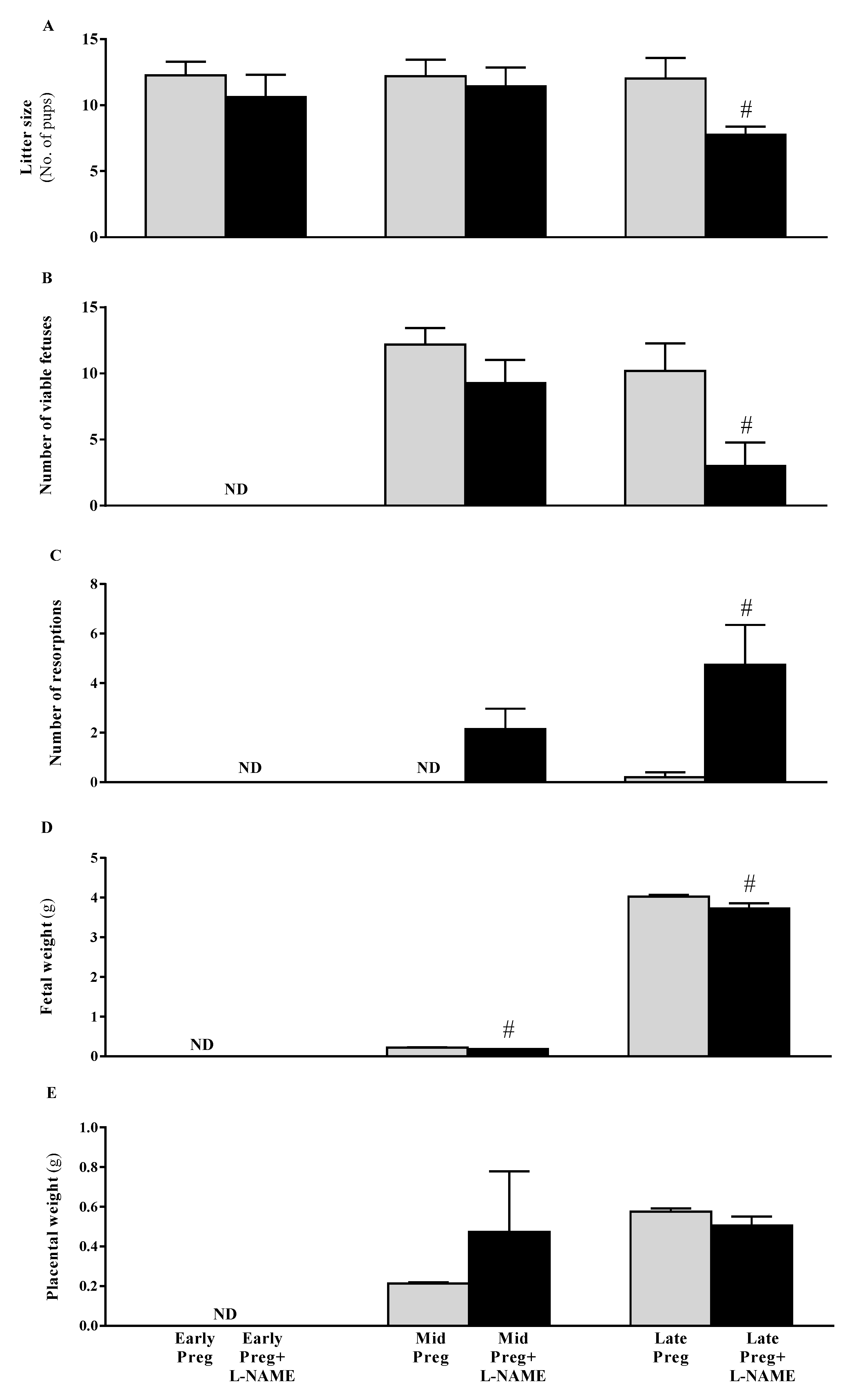
3.3. Intrauterine Growth Restrictions Are Found in Middle and Late Pregnant Rats Treated with L-NAME
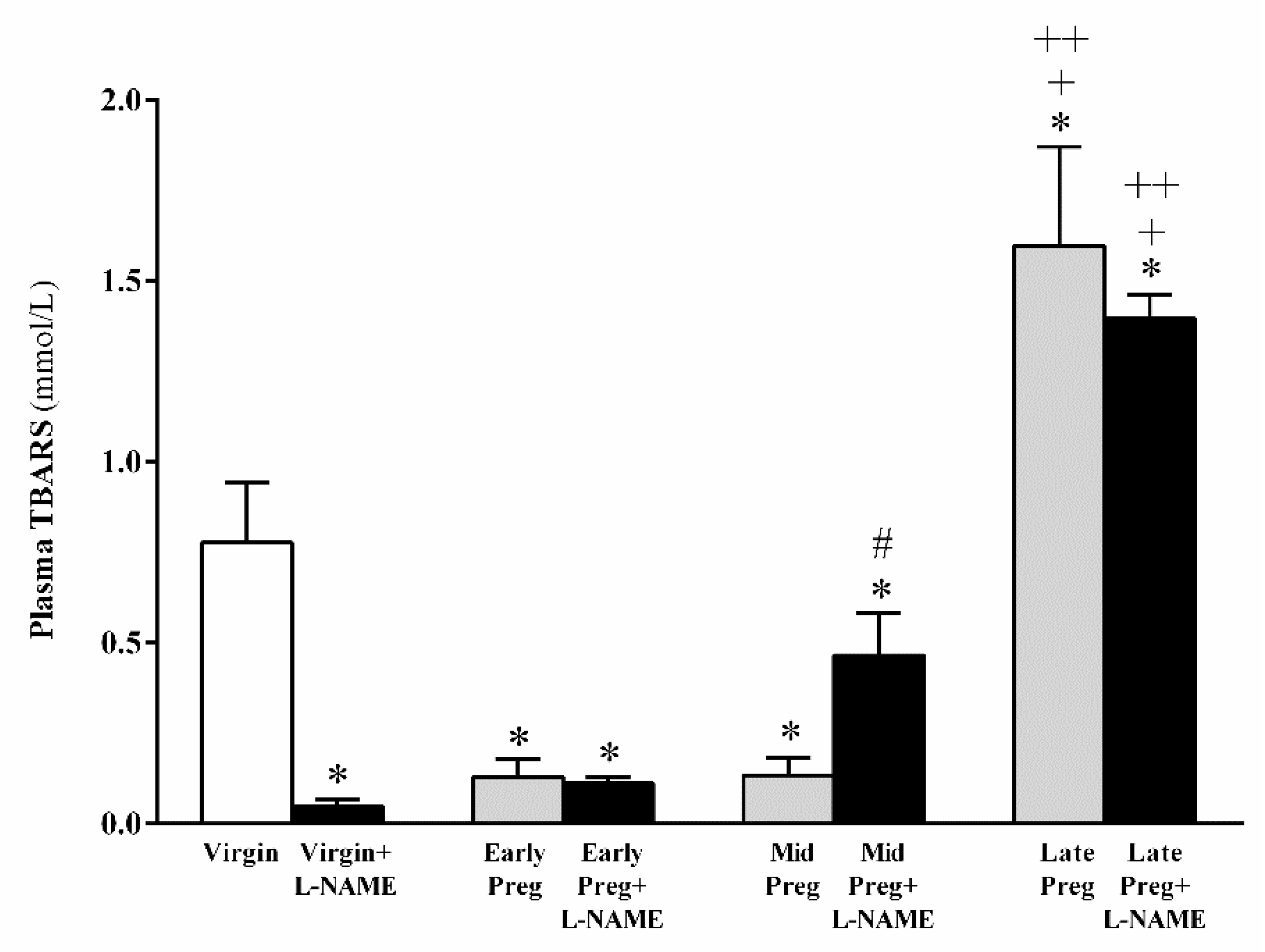
3.4. Oxidative Stress Is Increased in Middle and Late Pregnant Rats Treated with L-NAME as Well as in Late Pregnant Rats Treated with Saline. However, Oxidative Stress Is Reduced in L-NAME-Treated Virgin Rats, While No Significant Difference Is Found in Early Pregnancy Stage Treated (Or Not) with L-NAME
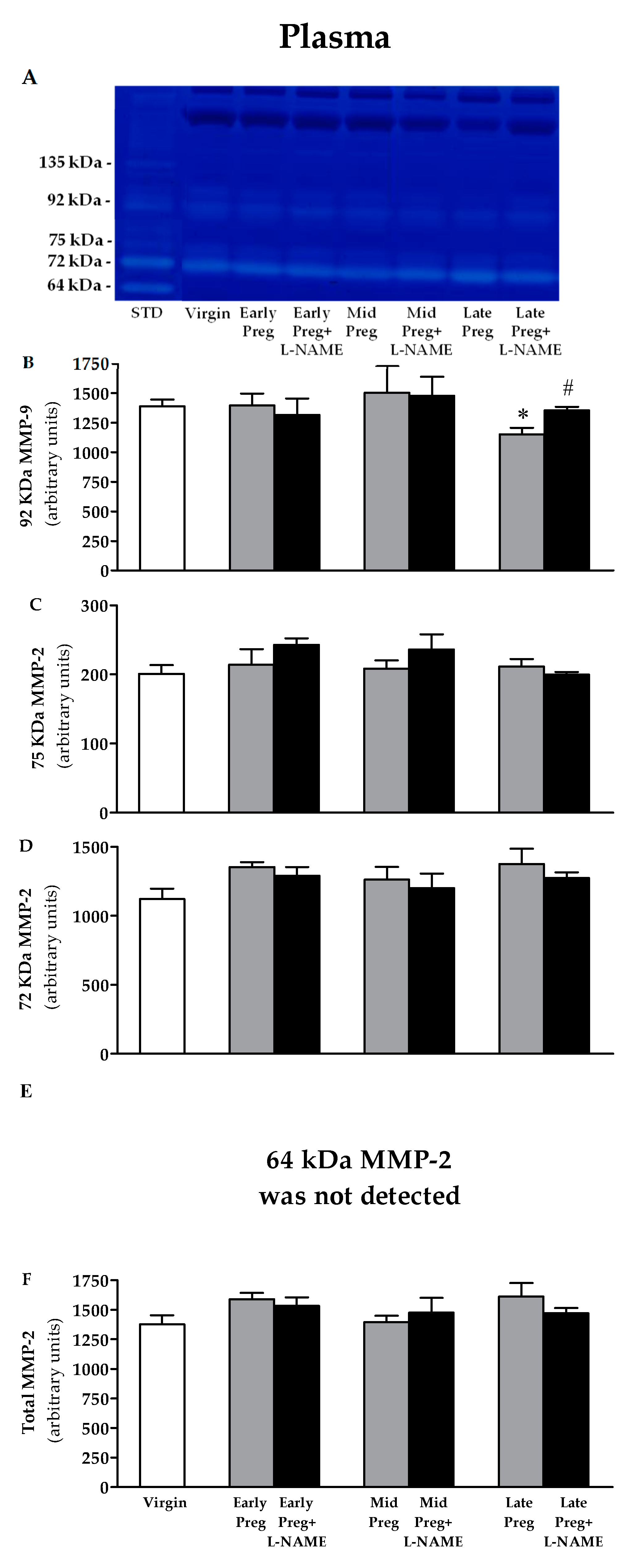
3.5. Changes in Activity of MMP-9 in Plasma Are Found Only at the Late Pregnancy Stage, in Which a Decrease Is Observed in Saline-Treated Pregnant Rats, While Increases Are Observed in L-NAME-Treated Pregnant Rats Compared to the Virgin Group. However, No Differences Are Found in the Activity of MMP-2 in Plasma
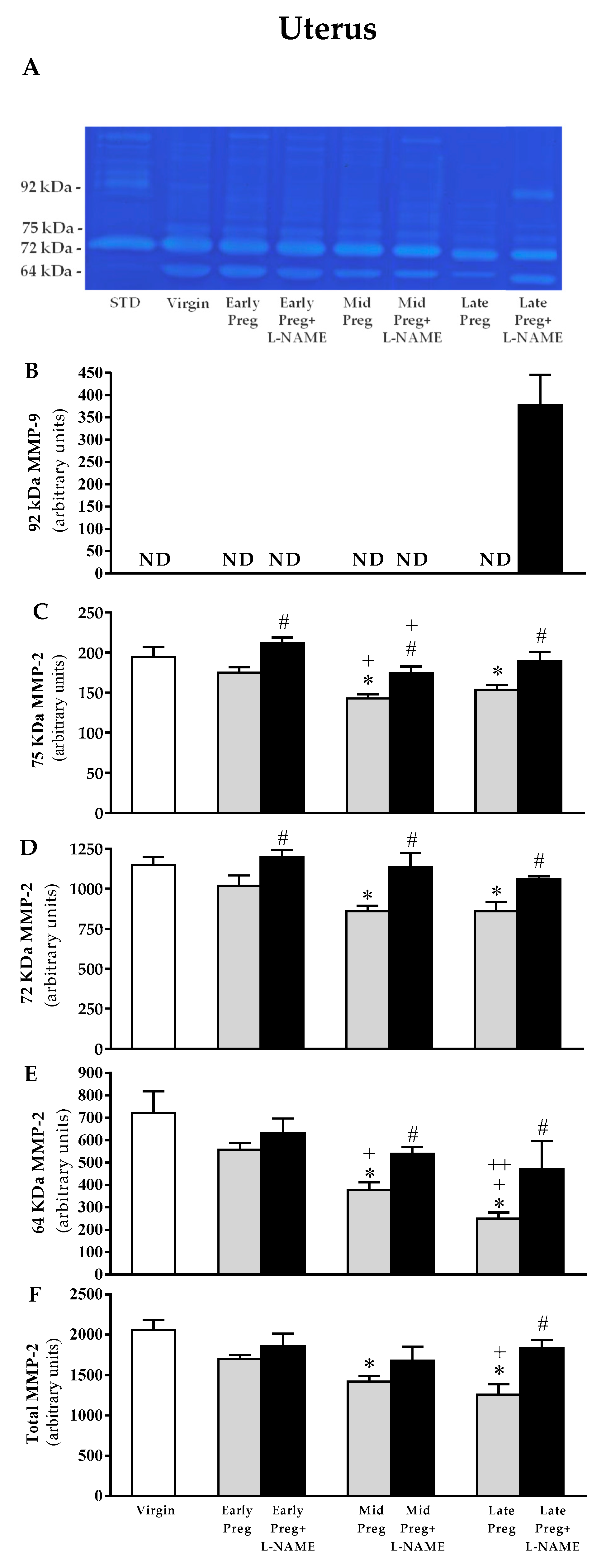
3.6. Uterine MMP-9 Activity Is Only Detected in L-NAME-Treated Pregnant Rats, While Uterine MMP-2 Is Reduced in Middle and Late, but Not in Early Pregnant Rats Treated with Saline. However, L-NAME-Treated Pregnant Rats Presented Increases in MMP-2 Activity in Early, Middle, and Late Pregnancy Stages
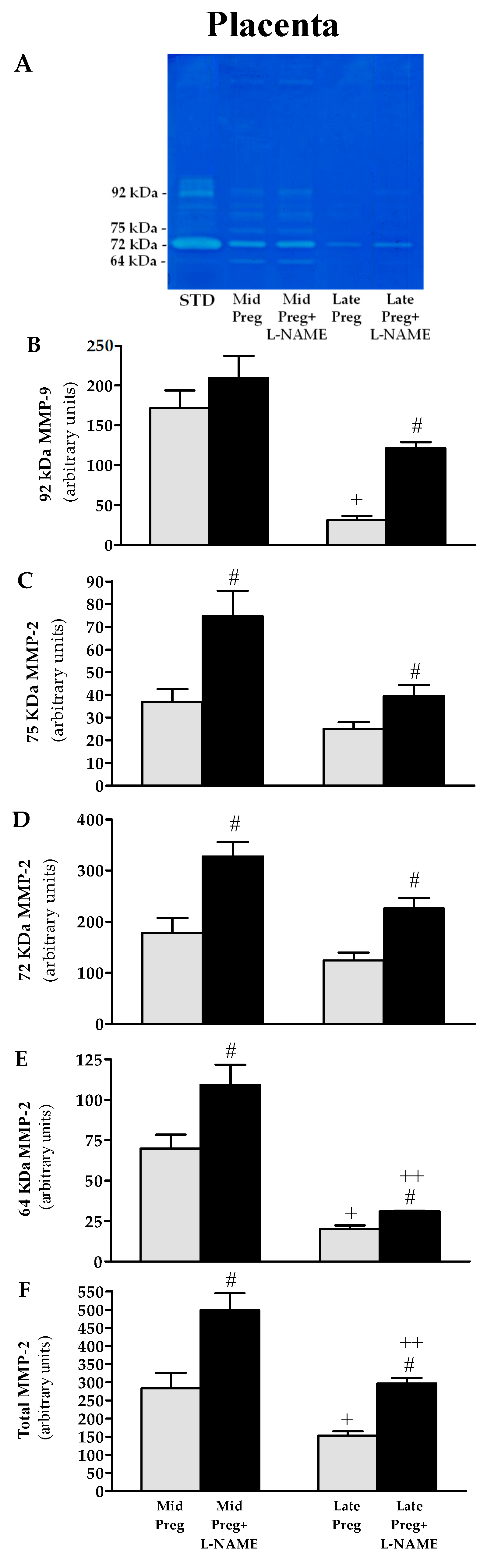
3.7. Placental MMP-9 and MMP-2 Activities Are Decreased in Saline-Treated Pregnant Rats, While Increases Are Found in L-NAME-Treated Pregnant Rats
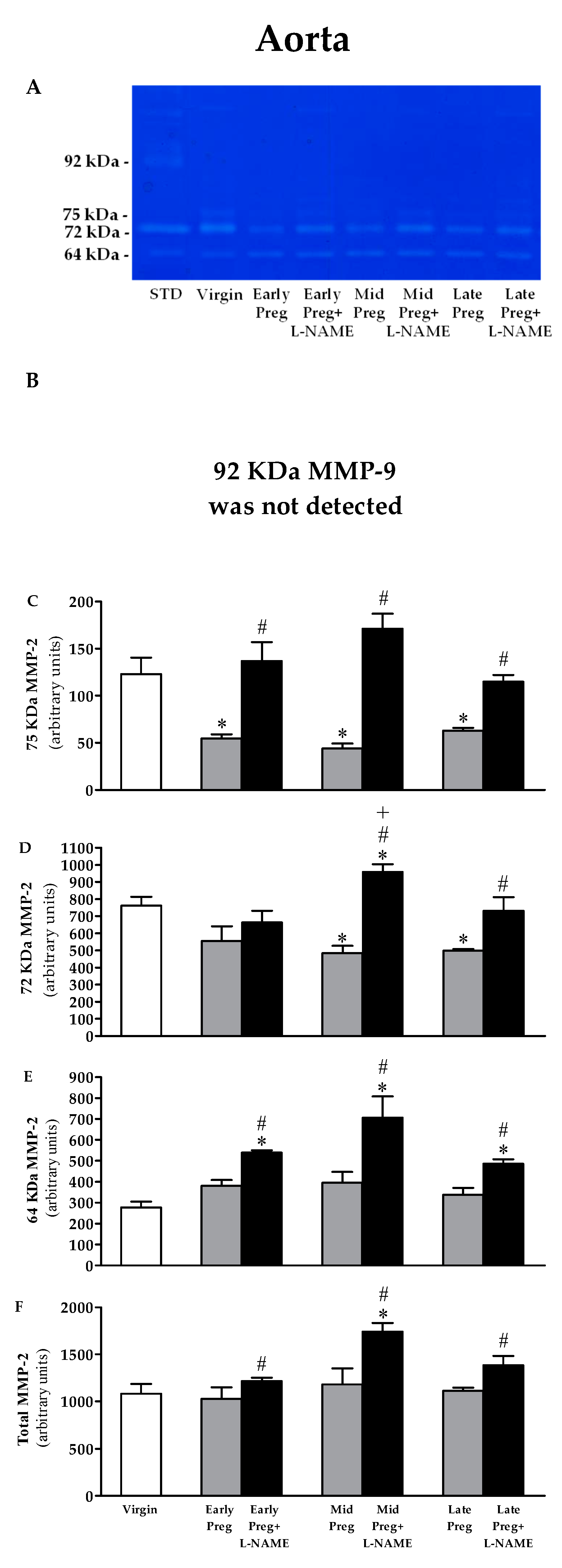
3.8. MMP-9 Is Not Detected in the Aorta. Moreover, MMP-2 Activity Is Reduced in Saline-Treated Pregnant Rats, While Increased MMP-2 Activity Is Observed in L-NAME-Treated Pregnant Rats
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Compliance with Ethical Standards
References
- James, J.L.; Saghian, R.; Perwick, R.; Clark, A.R. Trophoblast plugs: Impact on utero-placental haemodynamics and spiral artery remodelling. Hum. Reprod. 2018, 33, 1430–1441. [Google Scholar] [CrossRef] [PubMed]
- Ouzounian, J.G.; Elkayam, U. Physiologic changes during normal pregnancy and delivery. Cardiol. Clin. 2012, 30, 317–329. [Google Scholar] [CrossRef] [PubMed]
- Lopes van Balen, V.A.; Spaan, J.J.; Ghossein, C.; van Kuijk, S.M.; Spaanderman, M.E.; Peeters, L.L. Early pregnancy circulatory adaptation and recurrent hypertensive disease: An explorative study. Reprod. Sci. 2013, 20, 1069–1074. [Google Scholar] [CrossRef] [PubMed]
- Poston, L.; McCarthy, A.L.; Ritter, J.M. Control of vascular resistance in the maternal and feto-placental arterial beds. Pharmacol. Ther. 1995, 65, 215–239. [Google Scholar] [CrossRef]
- Hodzic, J.; Izetbegovic, S.; Muracevic, B.; Iriskic, R.; Stimjanin Jovic, H. Nitric oxide biosynthesis during normal pregnancy and pregnancy complicated by preeclampsia. Med. Glas. (Zenica) 2017, 14, 211–217. [Google Scholar] [CrossRef]
- Khalil, A.; Hardman, L.O.; Brien, P. The role of arginine, homoarginine and nitric oxide in pregnancy. Amino Acids 2015, 47, 1715–1727. [Google Scholar] [CrossRef]
- Chen, J.; Khalil, R.A. Matrix Metalloproteinases in Normal Pregnancy and Preeclampsia. Prog. Mol. Biol. Transl. Sci. 2017, 148, 87–165. [Google Scholar] [CrossRef] [Green Version]
- Shimonovitz, S.; Hurwitz, A.; Dushnik, M.; Anteby, E.; Geva-Eldar, T.; Yagel, S. Developmental regulation of the expression of 72 and 92 kd type IV collagenases in human trophoblasts: A possible mechanism for control of trophoblast invasion. Am. J. Obstet. Gynecol. 1994, 171, 832–838. [Google Scholar] [CrossRef]
- Isaka, K.; Usuda, S.; Ito, H.; Sagawa, Y.; Nakamura, H.; Nishi, H.; Suzuki, Y.; Li, Y.F.; Takayama, M. Expression and activity of matrix metalloproteinase 2 and 9 in human trophoblasts. Placenta 2003, 24, 53–64. [Google Scholar] [CrossRef]
- Alexander, B.T.; Miller, M.T.; Kassab, S.; Novak, J.; Reckelhoff, J.F.; Kruckeberg, W.C.; Granger, J.P. Differential expression of renal nitric oxide synthase isoforms during pregnancy in rats. Hypertension 1999, 33, 435–439. [Google Scholar] [CrossRef]
- Morris, N.H.; Eaton, B.M.; Dekker, G. Nitric oxide, the endothelium, pregnancy and pre-eclampsia. Br. J. Obstet. Gynaecol. 1996, 103, 4–15. [Google Scholar] [CrossRef] [PubMed]
- Sandrim, V.C.; Montenegro, M.F.; Palei, A.C.; Metzger, I.F.; Sertorio, J.T.; Cavalli, R.C.; Tanus-Santos, J.E. Increased circulating cell-free hemoglobin levels reduce nitric oxide bioavailability in preeclampsia. Free Radic. Biol. Med. 2010, 49, 493–500. [Google Scholar] [CrossRef] [PubMed]
- Ehsanipoor, R.M.; Fortson, W.; Fitzmaurice, L.E.; Liao, W.X.; Wing, D.A.; Chen, D.B.; Chan, K. Nitric oxide and carbon monoxide production and metabolism in preeclampsia. Reprod. Sci. 2013, 20, 542–548. [Google Scholar] [CrossRef]
- Matsubara, K.; Matsubara, Y.; Hyodo, S.; Katayama, T.; Ito, M. Role of nitric oxide and reactive oxygen species in the pathogenesis of preeclampsia. J. Obstet. Gynaecol. Res. 2010, 36, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Pimentel, A.M.; Pereira, N.R.; Costa, C.A.; Mann, G.E.; Cordeiro, V.S.; de Moura, R.S.; Brunini, T.M.; Mendes-Ribeiro, A.C.; Resende, A.C. L-arginine-nitric oxide pathway and oxidative stress in plasma and platelets of patients with pre-eclampsia. Hypertens Res. 2013, 36, 783–788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiessl, B.; Strasburger, C.; Bidlingmaier, M.; Mylonas, I.; Jeschke, U.; Kainer, F.; Friese, K. Plasma- and urine concentrations of nitrite/nitrate and cyclic Guanosinemonophosphate in intrauterine growth restricted and preeclamptic pregnancies. Arch. Gynecol. Obstet. 2006, 274, 150–154. [Google Scholar] [CrossRef] [PubMed]
- Amaral, L.M.; Cornelius, D.C.; Harmon, A.; Moseley, J.; Martin, J.N., Jr.; LaMarca, B. 17-hydroxyprogesterone caproate significantly improves clinical characteristics of preeclampsia in the reduced uterine perfusion pressure rat model. Hypertension 2015, 65, 225–231. [Google Scholar] [CrossRef]
- Alexander, B.T.; Llinas, M.T.; Kruckeberg, W.C.; Granger, J.P. L-arginine attenuates hypertension in pregnant rats with reduced uterine perfusion pressure. Hypertension 2004, 43, 832–836. [Google Scholar] [CrossRef]
- Mazzuca, M.Q.; Li, W.; Reslan, O.M.; Yu, P.; Mata, K.M.; Khalil, R.A. Downregulation of microvascular endothelial type B endothelin receptor is a central vascular mechanism in hypertensive pregnancy. Hypertension 2014, 64, 632–643. [Google Scholar] [CrossRef]
- Montagnana, M.; Lippi, G.; Albiero, A.; Scevarolli, S.; Salvagno, G.L.; Franchi, M.; Guidi, G.C. Evaluation of metalloproteinases 2 and 9 and their inhibitors in physiologic and pre-eclamptic pregnancy. J. Clin. Lab. Anal. 2009, 23, 88–92. [Google Scholar] [CrossRef] [PubMed]
- Visse, R.; Nagase, H. Matrix metalloproteinases and tissue inhibitors of metalloproteinases: Structure, function, and biochemistry. Circ. Res. 2003, 92, 827–839. [Google Scholar] [CrossRef] [PubMed]
- Novaro, V.; Pustovrh, C.; Colman-Lerner, A.; Radisky, D.; Lo Nostro, F.; Paz, D.; Jawerbaum, A.; González, E. Nitric oxide induces gelatinase A (matrix metalloproteinase 2) during rat embryo implantation. Fertil. Steril. 2002, 78, 1278–1287. [Google Scholar] [CrossRef]
- Diessler, M.; Ventureira, M.; Hernandez, R.; Sobarzo, C.; Casas, L.; Barbeito, C.; Cebral, E. Differential expression and activity of matrix metalloproteinases 2 and 9 in canine early placenta. Reprod. Domest. Anim. 2017, 52, 35–43. [Google Scholar] [CrossRef]
- Dang, Y.; Li, W.; Tran, V.; Khalil, R.A. EMMPRIN-mediated induction of uterine and vascular matrix metalloproteinases during pregnancy and in response to estrogen and progesterone. Biochem. Pharmacol. 2013, 86, 734–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kizaki, K.; Ushizawa, K.; Takahashi, T.; Yamada, O.; Todoroki, J.; Sato, T.; Ito, A.; Hashizume, K. Gelatinase (MMP-2 and -9) expression profiles during gestation in the bovine endometrium. Reprod. Biol. Endocrinol. 2008, 6, 66. [Google Scholar] [CrossRef]
- Myers, J.E.; Merchant, S.J.; Macleod, M.; Mires, G.J.; Baker, P.N.; Davidge, S.T. MMP-2 levels are elevated in the plasma of women who subsequently develop preeclampsia. Hypertens. Pregnancy 2005, 24, 103–115. [Google Scholar] [CrossRef]
- Eleuterio, N.M.; Palei, A.C.; Rangel Machado, J.S.; Tanus-Santos, J.E.; Cavalli, R.C.; Sandrim, V.C. Positive correlations between circulating adiponectin and MMP2 in preeclampsia pregnant. Pregnancy Hypertens. 2015, 5, 205–208. [Google Scholar] [CrossRef]
- Martinez-Fierro, M.L.; Perez-Favila, A.; Garza-Veloz, I.; Espinoza-Juarez, M.A.; Avila-Carrasco, L.; Delgado-Enciso, I.; Ortiz-Castro, Y.; Cardenas-Vargas, E.; Cid-Baez, M.A.; Ramirez-Santoyo, R.M.; et al. Matrix metalloproteinase multiplex screening identifies increased MMP-2 urine concentrations in women predicted to develop preeclampsia. Biomarkers 2018, 23, 18–24. [Google Scholar] [CrossRef]
- Meschiari, C.A.; Izidoro-Toledo, T.; Gerlach, R.F.; Tanus-Santos, J.E. Nitric oxide attenuates matrix metalloproteinase-9 production by endothelial cells independent of cGMP- or NFkappaB-mediated mechanisms. Mol. Cell Biochem. 2013, 378, 127–135. [Google Scholar] [CrossRef]
- Demacq, C.; Metzger, I.F.; Gerlach, R.F.; Tanus-Santos, J.E. Inverse relationship between markers of nitric oxide formation and plasma matrix metalloproteinase-9 levels in healthy volunteers. Clin. Chim. Acta 2008, 394, 72–76. [Google Scholar] [CrossRef]
- Metzger, I.F.; Sandrim, V.C.; Tanus-Santos, J.E. Endogenous nitric oxide formation correlates negatively with circulating matrix metalloproteinase (MMP)-2 and MMP-9 levels in black subjects. Mol. Cell Biochem. 2012, 360, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Eberhardt, W.; Beeg, T.; Beck, K.F.; Walpen, S.; Gauer, S.; Böhles, H.; Pfeilschifter, J. Nitric oxide modulates expression of matrix metalloproteinase-9 in rat mesangial cells. Kidney Int. 2000, 57, 59–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinha, I.; Hannawa, K.K.; Ailawadi, G.; Woodrum, D.T.; Ford, J.W.; Henke, P.K.; Stanley, J.C.; Eagleton, M.J.; Upchurch, G.R., Jr. The nitric oxide donor DETA-NONOate decreases matrix metalloproteinase-9 expression and activity in rat aortic smooth muscle and abdominal aortic explants. Ann. Vasc. Surg. 2006, 20, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Dey, N.B.; Lincoln, T.M. Possible involvement of Cyclic-GMP-dependent protein kinase on matrix metalloproteinase-2 expression in rat aortic smooth muscle cells. Mol. Cell Biochem. 2012, 368, 27–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baijnath, S.; Murugesan, S.; Mackraj, I.; Gathiram, P.; Moodley, J. The effects of sildenafil citrate on urinary podocin and nephrin mRNA expression in an L-NAME model of pre-eclampsia. Mol. Cell Biochem. 2017, 427, 59–67. [Google Scholar] [CrossRef]
- Baijnath, S.; Soobryan, N.; Mackraj, I.; Gathiram, P.; Moodley, J. The optimization of a chronic nitric oxide synthase (NOS) inhibition model of pre-eclampsia by evaluating physiological changes. Eur. J. Obstet. Gynecol. Reprod. Biol. 2014, 182, 71–75. [Google Scholar] [CrossRef]
- Osol, G.; Barron, C.; Gokina, N.; Mandala, M. Inhibition of nitric oxide synthases abrogates pregnancy-induced uterine vascular expansive remodeling. J. Vasc. Res. 2009, 46, 478–486. [Google Scholar] [CrossRef]
- Yamaleyeva, L.M.; Sun, Y.; Bledsoe, T.; Hoke, A.; Gurley, S.B.; Brosnihan, K.B. Photoacoustic imaging for in vivo quantification of placental oxygenation in mice. FASEB J. 2017, 31, 5520–5529. [Google Scholar] [CrossRef]
- Marcondes, F.K.; Bianchi, F.J.; Tanno, A.P. Determination of the estrous cycle phases of rats: Some helpful considerations. Braz. J. Biol. 2002, 62, 609–614. [Google Scholar] [CrossRef]
- Fernández Celadilla, L.; Carbajo Rueda, M.; Muñoz Rodríguez, M. Prolonged inhibition of nitric oxide synthesis in pregnant rats: Effects on blood pressure, fetal growth and litter size. Arch. Gynecol. Obstet. 2005, 271, 243–248. [Google Scholar] [CrossRef]
- Nascimento, R.A.; Mendes, G.; Possomato-Vieira, J.S.; Gonçalves-Rizzi, V.H.; Kushima, H.; Delella, F.K.; Dias-Junior, C.A. Metalloproteinase Inhibition Protects against Reductions in Circulating Adrenomedullin during Lead-induced Acute Hypertension. Basic Clin. Pharmacol. Toxicol. 2015, 116, 508–515. [Google Scholar] [CrossRef] [PubMed]
- Goncalves-Rizzi, V.H.; Possomato-Vieira, J.S.; Nascimento, R.A.; Caldeira-Dias, M.; Dias-Junior, C.A. Maternal hypertension and feto-placental growth restriction is reversed by sildenafil: Evidence of independent effects of circulating nitric oxide levels. Eur. J. Pharmacol. 2018, 822, 119–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, R.Q.; Sun, M.N.; Yang, Z. Effects of preeclampsia-like symptoms at early gestational stage on feto-placental outcomes in a mouse model. Chin. Med. J. (Engl.) 2010, 123, 707–712. [Google Scholar] [PubMed]
- Miranda, K.M.; Espey, M.G.; Wink, D.A. A rapid, simple spectrophotometric method for simultaneous detection of nitrate and nitrite. Nitric Oxide 2001, 5, 62–71. [Google Scholar] [CrossRef]
- Perico, L.L.; Heredia-Vieira, S.C.; Beserra, F.P.; de Cássia Dos Santos, R.; Weiss, M.B.; Resende, F.A.; Dos Santos Ramos, M.A.; Bonifácio, B.V.; Bauab, T.M.; Varanda, E.A.; et al. Does the gastroprotective action of a medicinal plant ensure healing effects? An integrative study of the biological effects of Serjania marginata Casar (Sapindaceae) in rats. J. Ethnopharmacol. 2015, 172, 312–324. [Google Scholar] [CrossRef]
- Rizzi, E.; Castro, M.M.; Prado, C.M.; Silva, C.A.; Fazan, R., Jr.; Rossi, M.A.; Tanus-Santos, J.E.; Gerlach, R.F. Matrix metalloproteinase inhibition improves cardiac dysfunction and remodeling in 2-kidney, 1-clip hypertension. J Card Fail. 2010, 16, 599–608. [Google Scholar] [CrossRef]
- Rizzi, R.; Ceron, C.S.; Guimaraes, D.A.; Prado, C.M.; Rossi, M.A.; Gerlach, R.F.; Tanus-Santos, J.E. Temporal changes in cardiac matrix metalloproteinase activity, oxidative stress, and TGF-beta in renovascular hypertension-induced cardiac hypertrophy. Exp. Mol. Pathol. 2013, 94, 1–9. [Google Scholar] [CrossRef]
- Li, W.; Mata, K.M.; Mazzuca, M.Q.; Khalil, R.A. Altered matrix metalloproteinase-2 and -9 expression/activity links placental ischemia and anti-angiogenic sFlt-1 to uteroplacental and vascular remodeling and collagen deposition in hypertensive pregnancy. Biochem. Pharmacol. 2014, 89, 370–385. [Google Scholar] [CrossRef] [Green Version]
- Guimaraes, D.A.; Rizzi, E.; Ceron, C.S.; Martins-Oliveira, A.; Gerlach, R.F.; Shiva, S.; Tanus-Santos, J.E. Atorvastatin and sildenafil decrease vascular TGF-β levels and MMP-2 activity and ameliorate arterial remodeling in a model of renovascular hypertension. Redox. Biol. 2015, 6, 386–395. [Google Scholar] [CrossRef]
- Goncalves-Rizzi, V.H.; Possomato-Vieira, J.S.; Sales Graca, T.U.; Nascimento, R.A.; Dias-Junior, C.A. Sodium nitrite attenuates hypertension-in-pregnancy and blunts increases in soluble fms-like tyrosine kinase-1 and in vascular endothelial growth factor. Nitric Oxide 2016, 57, 71–78. [Google Scholar] [CrossRef] [Green Version]
- Del Mauro, J.S.; Prince, P.D.; Donato, M.; Fernandez Machulsky, N.; Morettón, M.A.; González, G.E.; Bertera, F.M.; Carranza, A.; Gorzalczany, S.B.; Chiappetta, D.A.; et al. Effects of carvedilol or amlodipine on target organ damage in L-NAME hypertensive rats: Their relationship with blood pressure variability. J. Am. Soc. Hypertens. 2017, 11, 227–240. [Google Scholar] [CrossRef] [PubMed]
- Amaral, L.M.; Pinheiro, L.C.; Guimaraes, D.A.; Palei, A.C.; Sertório, J.T.; Portella, R.L.; Tanus-Santos, J.E. Antihypertensive effects of inducible nitric oxide synthase inhibition in experimental pre-eclampsia. J. Cell Mol. Med. 2013, 17, 1300–1307. [Google Scholar] [CrossRef] [PubMed]
- Kusinski, L.C.; Stanley, J.L.; Dilworth, M.R.; Hirt, C.J.; Andersson, I.J.; Renshall, L.J.; Baker, B.C.; Baker, P.N.; Sibley, C.P.; Wareing, M.; et al. eNOS knockout mouse as a model of fetal growth restriction with an impaired uterine artery function and placental transport phenotype. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2012, 303, R86–R93. [Google Scholar] [CrossRef] [PubMed]
- VanWijk, M.J.; Kublickiene, K.; Boer, K.; VanBavel, E. Vascular function in preeclampsia. Cardiovasc Res. 2000, 47, 38–48. [Google Scholar] [CrossRef]
- Myers, J.; Mires, G.; Macleod, M.; Baker, P. In preeclampsia, the circulating factors capable of altering in vitro endothelial function precede clinical disease. Hypertension 2005, 45, 258–263. [Google Scholar] [CrossRef]
- Roberts, J.M.; Taylor, R.N.; Goldfien, A. Clinical and biochemical evidence of endothelial cell dysfunction in the pregnancy syndrome preeclampsia. Am. J. Hypertens. 1991, 4, 700–708. [Google Scholar] [CrossRef]
- Roberts, J.M.; Taylor, R.N.; Musci, T.J.; Rodgers, G.M.; Hubel, C.A.; McLaughlin, M.K. Pre-eclampsia: A disorders of the endothelial cells? Gynakologe 1992, 25, 2–6. [Google Scholar]
- Fernandez-Patron, C.; Stewart, K.G.; Zhang, Y.; Koivunen, E.; Radomski, M.W.; Davidge, S.T. Vascular matrix metalloproteinase-2-dependent cleavage of calcitonin gene-related peptide promotes vasoconstriction. Circ. Res. 2000, 87, 670–676. [Google Scholar] [CrossRef]
- Martinez, A.; Oh, H.R.; Unsworth, E.J.; Bregonzio, C.; Saavedra, J.M.; Stetler-Stevenson, W.G.; Cuttitta, F. Matrix metalloproteinase-2 cleavage of adrenomedullin produces a vasoconstrictor out of a vasodilator. Biochem. J. 2004, 383, 413–418. [Google Scholar] [CrossRef]
- Fernandez-Patron, C.; Radomski, M.W.; Davidge, S.T. Vascular matrix metalloproteinase-2 cleaves big endothelin-1 yielding a novel vasoconstrictor. Circ. Res. 1999, 85, 906–911. [Google Scholar] [CrossRef]
- Fernandez-Patron, C.; Zouki, C.; Whittal, R.; Chan, J.S.; Davidge, S.T.; Filep, J.G. Matrix metalloproteinases regulate neutrophil-endothelial cell adhesion through generation of endothelin-1[1-32]. FASEB J. 2001, 15, 2230–2240. [Google Scholar] [CrossRef] [PubMed]
- Roberts, J.M.; Bell, M.J. If we know so much about preeclampsia, why haven’t we cured the disease? J. Reprod. Immunol. 2013, 99, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Damsky, C.H.; Fitzgerald, M.L.; Fisher, S.J. Distribution patterns of extracellular matrix components and adhesion receptors are intricately modulated during first trimester cytotrophoblast differentiation along the invasive pathway, in vivo. J. Clin. Invest. 1992, 89, 210–222. [Google Scholar] [CrossRef] [PubMed]
- Salas, S.P.; Marshall, G.; Gutierrez, B.L.; Rosso, P. Time course of maternal plasma volume and hormonal changes in women with preeclampsia or fetal growth restriction. Hypertension 2006, 47, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Brosens, I.A.; Robertson, W.B.; Dixon, H.G. The role of the spiral arteries in the pathogenesis of preeclampsia. Obstet. Gynecol. Annu. 1972, 1, 177–191. [Google Scholar] [CrossRef]
- Meekins, J.W.; Pijnenborg, R.; Hanssens, M.; McFadyen, I.R.; van Asshe, A. A study of placental bed spiral arteries and trophoblast invasion in normal and severe pre-eclamptic pregnancies. Br. J. Obstet. Gynaecol. 1994, 101, 669–674. [Google Scholar] [CrossRef]
- Lavee, M.; Goldman, S.; Daniel-Spiegel, E.; Shalev, E. Matrix metalloproteinase-2 is elevated in midtrimester amniotic fluid prior to the development of preeclampsia. Reprod. Biol. Endocrinol. 2009, 7, 85. [Google Scholar] [CrossRef]
- Palei, A.C.; Sandrim, V.C.; Amaral, L.M.; Machado, J.S.; Cavalli, R.C.; Lacchini, R.; Duarte, G.; Tanus-Santos, J.E. Matrix metalloproteinase-9 polymorphisms affect plasma MMP-9 levels and antihypertensive therapy responsiveness in hypertensive disorders of pregnancy. Pharm. J. 2012, 12, 489–498. [Google Scholar] [CrossRef]
- Poon, L.C.; Nekrasova, E.; Anastassopoulos, P.; Livanos, P.; Nicolaides, K.H. First-trimester maternal serum matrix metalloproteinase-9 (MMP-9) and adverse pregnancy outcome. Prenat. Diagn. 2009, 29, 553–559. [Google Scholar] [CrossRef]
- Palei, A.C.; Sandrim, V.C.; Amaral, L.M.; Machado, J.S.; Cavalli, R.C.; Duarte, G.; Tanus-Santos, J.E. Association between matrix metalloproteinase (MMP)-2 polymorphisms and MMP-2 levels in hypertensive disorders of pregnancy. Exp. Mol. Pathol. 2012, 92, 217–221. [Google Scholar] [CrossRef]
- Belo, B.A.; Guimaraes, D.A.; Castro, M.M. Matrix Metalloproteinase 2 as a Potential Mediator of Vascular Smooth Muscle Cell Migration and Chronic Vascular Remodeling in Hypertension. Vasc. Res. 2015, 52, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Matthews, J.R.; Botting, C.H.; Panico, M.; Morris, H.R.; Hay, R.T. Inhibition of NF-kappaB DNA binding by nitric oxide. Nucleic Acids Res. 1996, 24, 2236–2242. [Google Scholar] [CrossRef] [PubMed]
- Van der Heiden, K.; Cuhlmann, S.; le Luong, A.; Zakkar, M.; Evans, P.C. Role of nuclear factor kappaB in cardiovascular health and disease. Clin. Sci. (Lond.) 2010, 118, 593–605. [Google Scholar] [CrossRef] [PubMed]
- Clark, I.M.; Swingler, T.E.; Sampieri, C.L.; Edwards, D.R. The regulation of matrix metalloproteinases and their inhibitors. Int. J. Biochem. Cell Biol. 2007, 40, 1362–1378. [Google Scholar] [CrossRef]
- Pfeilschifter, J.; Eberhardt, W.; Beck, K.F. Regulation of gene expression by nitric oxide. Pflugers Arch. 2001, 442, 479–486. [Google Scholar] [CrossRef]
- Chen, J.; Ren, Z.; Zhu, M.; Khalil, R.A. Decreased homodimerization and increased TIMP-1 complexation of uteroplacental and uterine arterial matrix metalloproteinase-9 during hypertension-in-pregnancy. Biochem. Pharmacol. 2017, 138, 81–95. [Google Scholar] [CrossRef]
- Nascimento, R.A.; Possomato-Vieira, J.S.; Gonçalves-Rizzi, V.H.; Bonacio, G.F.; Rizzi, E.; Dias-Junior, C.A. Hypertension, augmented activity of matrix metalloproteinases-2 and -9 and angiogenic imbalance in hypertensive pregnancy are attenuated by doxycycline. Eur. J. Pharmacol. 2018, 840, 60–69. [Google Scholar] [CrossRef]
- Jeyabalan, A.; Novak, J.; Danielson, L.A.; Kerchner, L.J.; Opett, S.L.; Conrad, K.P. Essential role for vascular gelatinase activity in relaxin-induced renal vasodilation, hyperfiltration, and reduced myogenic reactivity of small arteries. Circ. Res. 2003, 93, 1249–1257. [Google Scholar] [CrossRef]
- Lalu, M.M.; Xu, H.; Davidge, S.T. Matrix metalloproteinases: Control of vascular function and their potential role in preeclampsia. Front. Biosci. 2007, 12, 2484–2493. [Google Scholar] [CrossRef]
- Sankaralingam, S.; Arenas, I.A.; Lalu, M.M.; Davidge, S.T. Preeclampsia: Current understanding of the molecular basis of vascular dysfunction. Expert. Rev. Mol. Med. 2006, 8, 1–20. [Google Scholar] [CrossRef]
- Karthikeyan, V.J.; Lane, D.A.; Beevers, D.G.; Lip, G.Y.; Blann, A.D. Matrix metalloproteinases and their tissue inhibitors in hypertension-related pregnancy complications. J. Hum. Hypertens. 2013, 27, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Lambert, E.; Dasse, E.; Haye, B.; Petitfrere, E. TIMPs as multifacial proteins. Crit. Ver. Oncol. Hematol. 2004, 49, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Yuan, W.; Xu, N.; Li, J.; Chang, W. Icariin improves acute kidney injury and proteinuria in a rat model of pregnancy-induced hypertension. Mol. Med. Rep. 2017, 16, 7398–7404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdalvand, A.; Morton, J.S.; Bourque, S.L.; Quon, A.L.; Davidge, S.T. Matrix metalloproteinase enhances big-endothelin-1 constriction in mesenteric vessels of pregnant rats with reduced uterine blood flow. Hypertension 2013, 61, 488–493. [Google Scholar] [CrossRef]
- Amaral, T.A.S.; Ognibene, D.T.; Carvalho, L.C.R.M.; Rocha, A.P.M.; Costa, C.A.; Moura, R.S.; Resende, A.C. Differential responses of mesenteric arterial bed to vasoactive substances in L-NAME-induced preeclampsia: Role of oxidative stress and endothelial dysfunction. Clin. Exp. Hypertens. 2018, 40, 126–135. [Google Scholar] [CrossRef]
- Possomato-Vieira, J.S.; Chimini, J.S.; da Silva, M.L.S.; Dias-Junior, C.A. Increases in placental nitric oxide, but not nitric oxide-mediated relaxation, underlie the improvement in placental efficiency and antihypertensive effects of hydrogen sulphide donor in hypertensive pregnancy. Clin. Exp. Pharmacol. Physiol. 2018, 45, 1118–1127. [Google Scholar] [CrossRef]
- Myatt, L.; Webster, R.P. Vascular biology of preeclampsia. J. Thromb. Haemost. 2009, 7, 375–384. [Google Scholar] [CrossRef]
- Gilbert, J.S.; Ryan, M.J.; LaMarca, B.B.; Sedeek, M.; Murphy, S.R.; Granger, J.P. Pathophysiology of hypertension during preeclampsia: Linking placental ischemia with endothelial dysfunction. Am. J. Physiol. Heart Circ. Physiol. 2008, 294, H541–550. [Google Scholar] [CrossRef]
- Burton, G.J.; Jauniaux, E. Placental oxidative stress: From miscarriage to preeclampsia. J. Soc. Gynecol. Investig. 2004, 11, 342–352. [Google Scholar] [CrossRef]
- Roggensack, A.M.; Zhang, Y.; Davidge, S.T. Evidence for peroxynitrite formation in the vasculature of women with preeclampsia. Hypertension 1999, 33, 83–89. [Google Scholar] [CrossRef]
- Walsh, S.W. Maternal-placental interactions of oxidative stress and antioxidants in preeclampsia. Semin Reprod. Endocrinol. 1998, 16, 93–104. [Google Scholar] [CrossRef] [PubMed]
- Chow, A.K.; Cena, J.; Schulz, R. Acute actions and novel targets of matrix metalloproteinases in the heart and vasculature. Br. J. Pharmacol. 2007, 152, 189–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arenas, I.A.; Xu, Y.; Lopez-Jaramillo, P.; Davidge, S.T. Angiotensin II-induced MMP-2 release from endothelial cells is mediated by TNF-alpha. Am. J. Physiol. Cell Physiol. 2004, 286, C779–784. [Google Scholar] [CrossRef]
- Mori, K.; Uchida, T.; Yoshie, T.; Mizote, Y.; Ishikawa, F.; Katsuyama, M.; Shibanuma, M. A mitochondrial ROS pathway controls matrix metalloproteinase 9 levels and invasive properties in RAS-activated cancer cells. FEBS J. 2019, 286, 459–478. [Google Scholar] [CrossRef]
- Stanhewicz, A.E.; Jandu, S.; Santhanam, L.; Alexander, L.M. Increased angiotensin II sensitivity contributes to microvascular dysfunction in women who have had preeclampsia. Hypertension 2017, 70, 382–389. [Google Scholar] [CrossRef]
- Dinh, Q.N.; Drummond, G.R.; Kemp-Harper, B.K.; Diep, H.; De Silva, T.M.; Kim, H.A.; Vinh, A.; Robertson, A.A.B.; Cooper, M.A.; Mansell, A.; et al. Pressor response to angiotensin II is enhanced in aged mice and associated with inflammation, vasoconstriction and oxidative stress. Aging (Albany NY) 2017, 9, 1595–1606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen Dinh Cat, A.; Montezano, A.C.; Burger, D.; Touyz, R.M. Angiotensin II, NADPH oxidase, and redox signaling in the vasculature. Antioxid. Redox Signal. 2013, 19, 1110–1120. [Google Scholar] [CrossRef]
- Romao, M.; Weel, I.C.; Lifshitz, S.J.; Peraçoli, M.T. Elevated hyaluronan and extracellular matrix metalloproteinase inducer levels in women with preeclampsia. Arch. Gynecol. Obstet. 2014, 289, 575–579. [Google Scholar] [CrossRef]
- Sanchez-Aranguren, L.C.; Prada, C.E.; Riano-Medina, C.E.; Lopez, M. Endothelial dysfunction and preeclampsia: Role of oxidative stress. Front. Physiol. 2014, 5, 372. [Google Scholar] [CrossRef]
- Ali, S.M.; Khalil, R.A. Genetic, immune and vasoactive factors in the vascular dysfunction associated with hypertension in pregnancy. Expert Opin. Ther. Targets 2015, 19, 1495–1515. [Google Scholar] [CrossRef] [Green Version]
- Vanderlelie, J.J.; Perkins, A.V. Chronic nitric oxide synthase inhibition in pregnant rats does not result in placental oxidative stress. Hypertens. Pregnancy 2006, 25, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Gao, W.; Rong, D.; Wu, Z.; Khalil, R.A. Molecular determinants of microvascular dysfunction in hypertensive pregnancy and preeclampsia. Microcirculation 2018. [Google Scholar] [CrossRef] [PubMed]
- Castro, M.M.; Rizzi, E.; Prado, C.M.; Rossi, M.A.; Tanus-Santos, J.E.; Gerlach, R.F. Imbalance between matrix metalloproteinases and tissue inhibitor of metalloproteinases in hypertensive vascular remodeling. Matrix Biol. 2010, 29, 194–201. [Google Scholar] [CrossRef]
- Palei, A.C.; Granger, J.P.; Tanus-Santos, J.E. Matrix metalloproteinases as drug targets in preeclampsia. Curr. Drug. Targets. 2013, 14, 325–334. [Google Scholar]
- Redman, C.W.; Sacks, G.P.; Sargent, I.L. Preeclampsia: An excessive maternal inflammatory response to pregnancy. Am. J. Obstet. Gynecol. 1999, 180, 499–506. [Google Scholar] [CrossRef]
- Greer, I.A.; Lyall, F.; Perera, T.; Boswell, F.; Macara, L.M. Increased concentrations of cytokines interleukin-6 and interleukin-1 receptor antagonist in plasma of women with preeclampsia: A mechanism for endothelial dysfunction? Obstet. Gynecol. 1994, 84, 937–940. [Google Scholar]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nascimento, R.A.; Possomato-Vieira, J.S.; Bonacio, G.F.; Rizzi, E.; Dias-Junior, C.A. Reductions of Circulating Nitric Oxide are Followed by Hypertension during Pregnancy and Increased Activity of Matrix Metalloproteinases-2 and -9 in Rats. Cells 2019, 8, 1402. https://doi.org/10.3390/cells8111402
Nascimento RA, Possomato-Vieira JS, Bonacio GF, Rizzi E, Dias-Junior CA. Reductions of Circulating Nitric Oxide are Followed by Hypertension during Pregnancy and Increased Activity of Matrix Metalloproteinases-2 and -9 in Rats. Cells. 2019; 8(11):1402. https://doi.org/10.3390/cells8111402
Chicago/Turabian StyleNascimento, Regina A., Jose S. Possomato-Vieira, Giselle F. Bonacio, Elen Rizzi, and Carlos A. Dias-Junior. 2019. "Reductions of Circulating Nitric Oxide are Followed by Hypertension during Pregnancy and Increased Activity of Matrix Metalloproteinases-2 and -9 in Rats" Cells 8, no. 11: 1402. https://doi.org/10.3390/cells8111402