Impact of Q141K on the Transport of Epidermal Growth Factor Receptor Tyrosine Kinase Inhibitors by ABCG2

 and
and 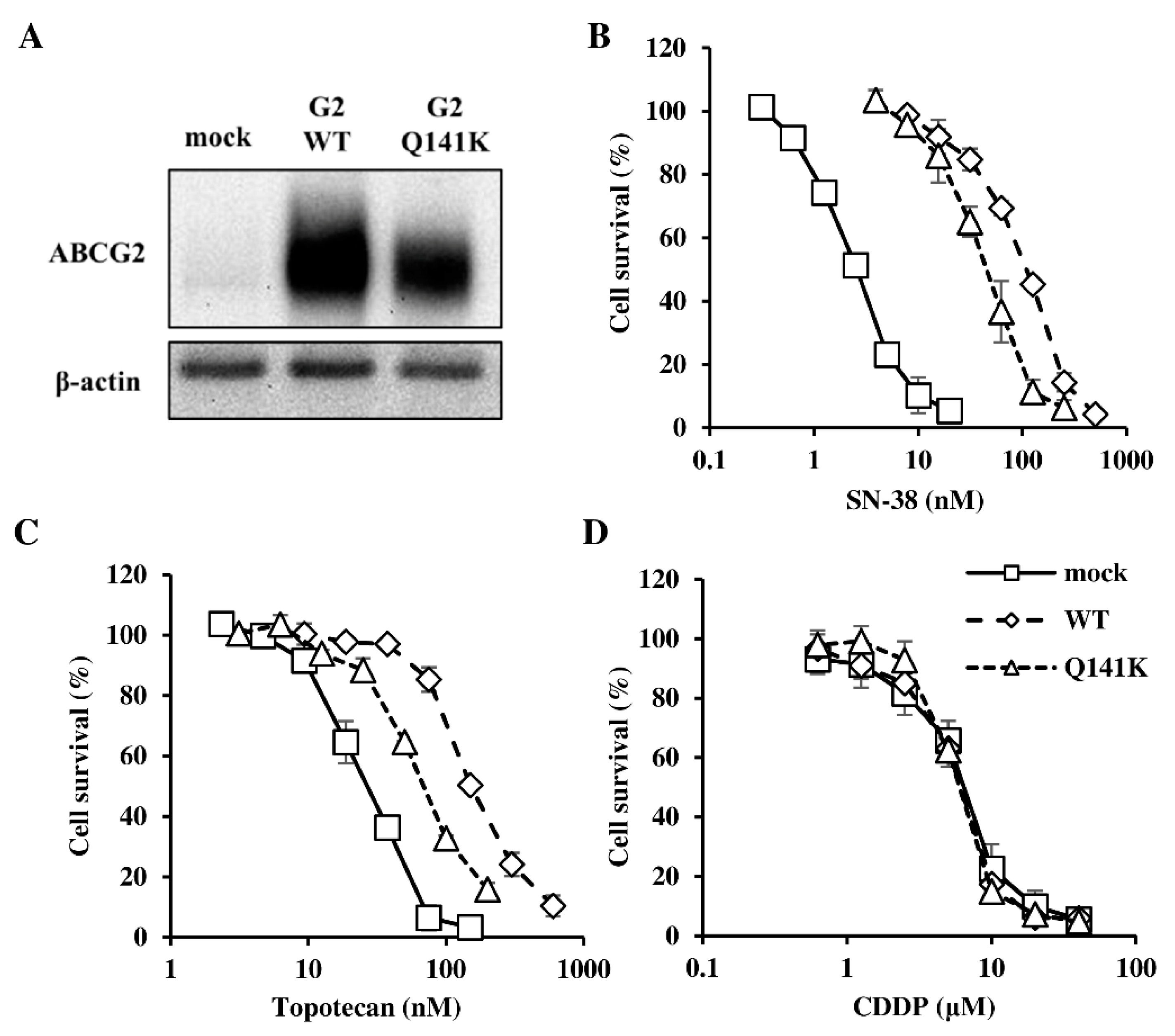{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines
2.2. Preparation of Cell Lysates
2.3. Immunoblotting Analysis
2.4. MTT Assay
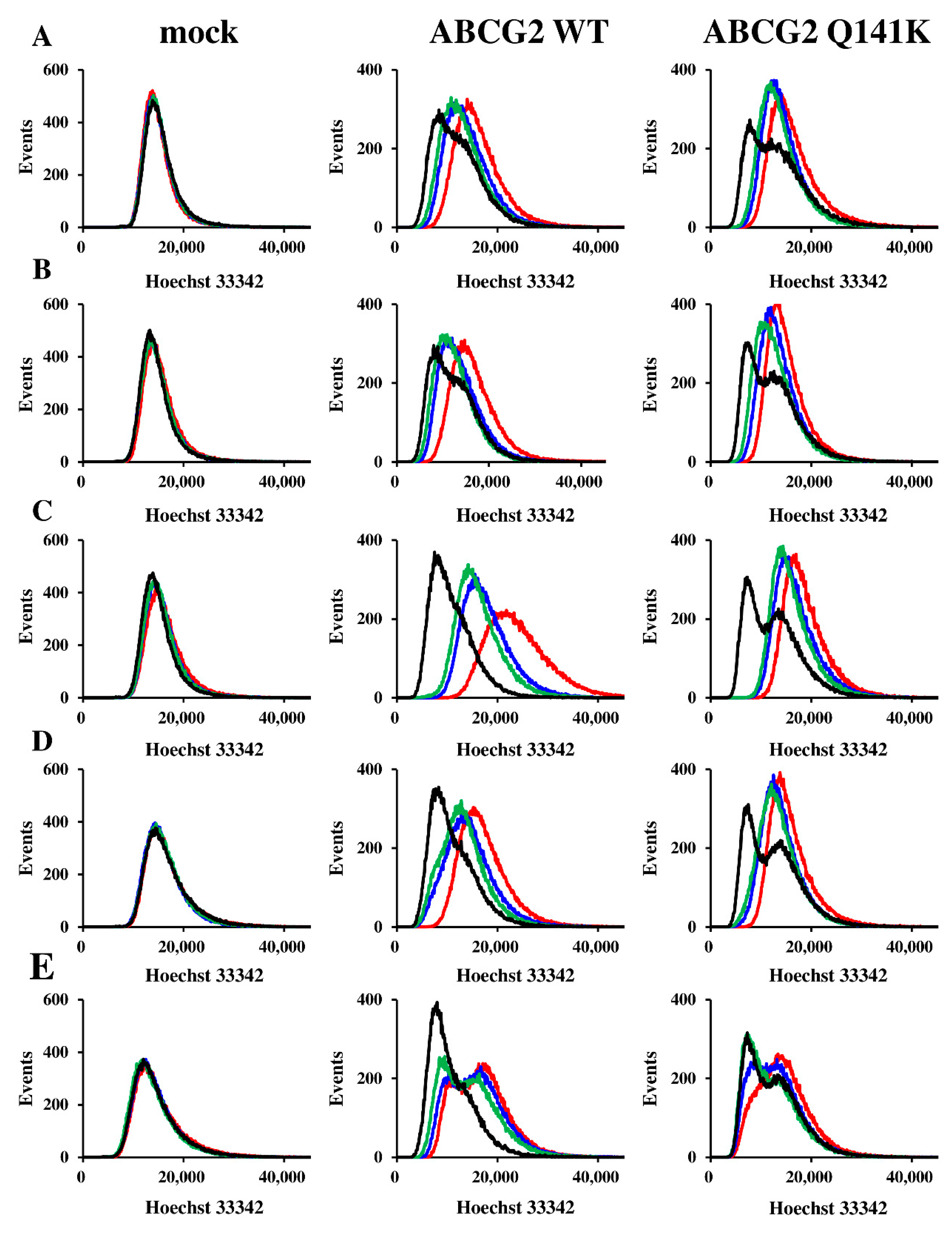
2.5. Imaging Cytometry
2.6. HPLC
2.7. Statistical Analysis
3. Results
3.1. Expression of ABCG2 and Resistance of Flp-In-293 Cells
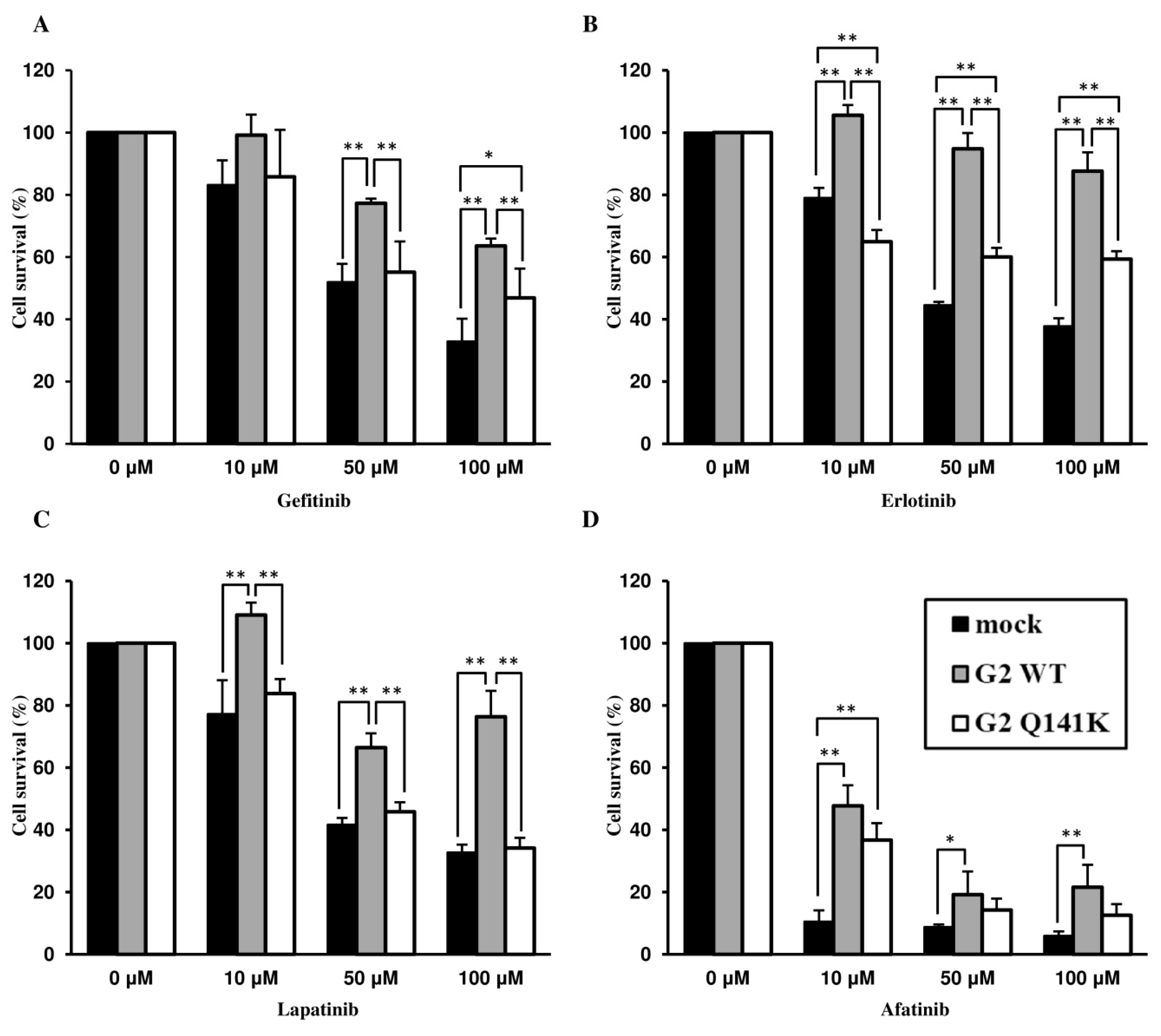
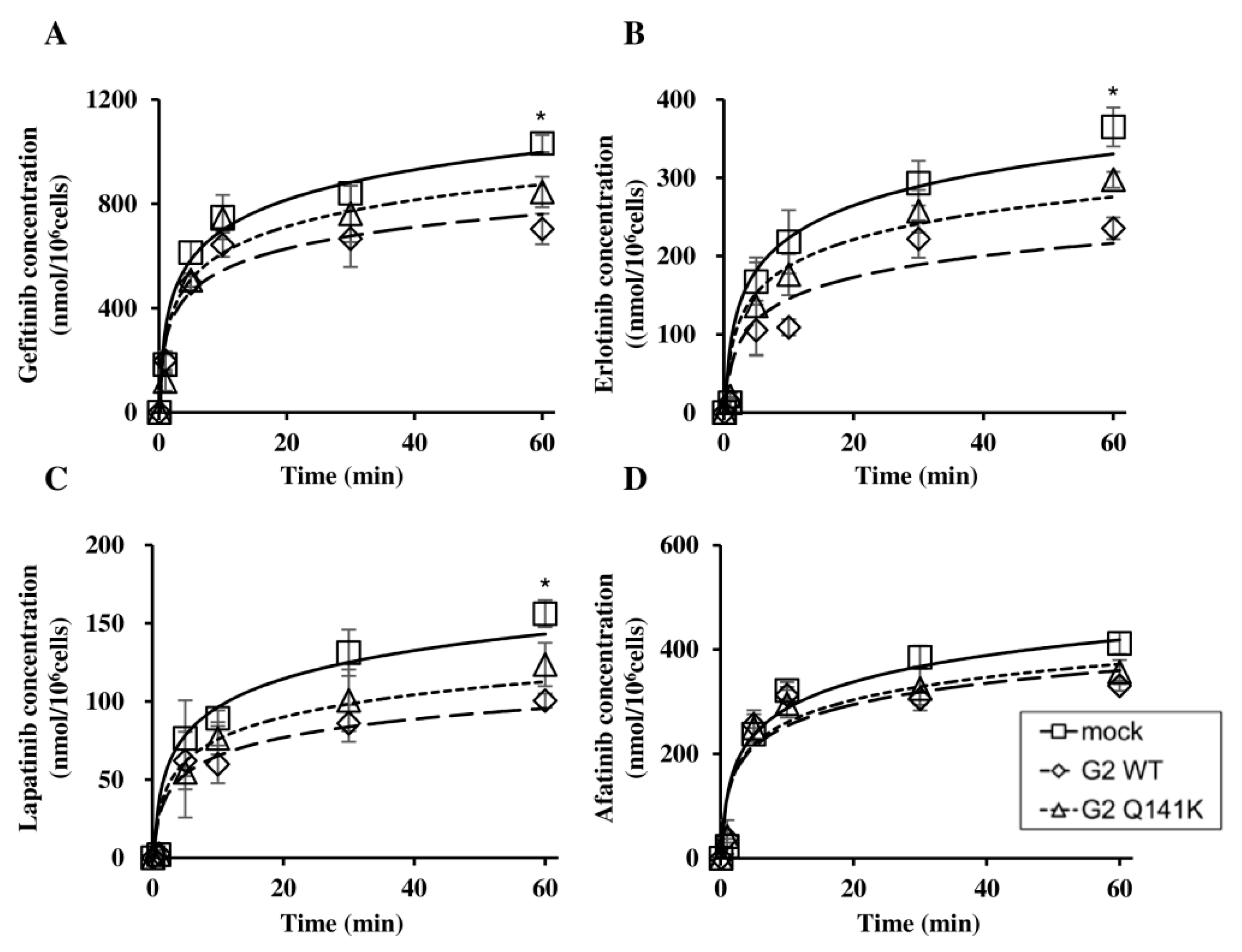
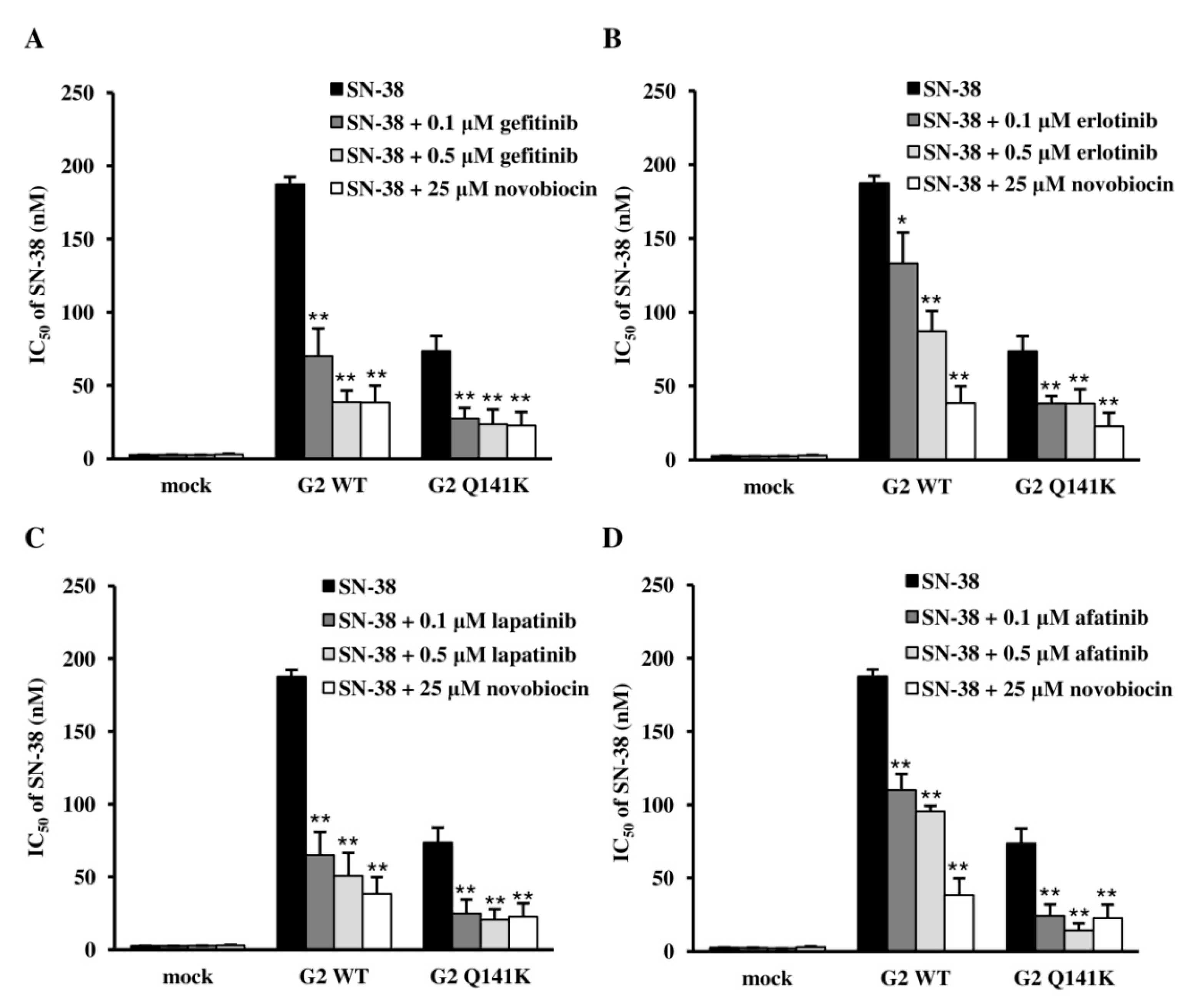
3.2. Effect of Q141K on Transport of EGFR TKIs
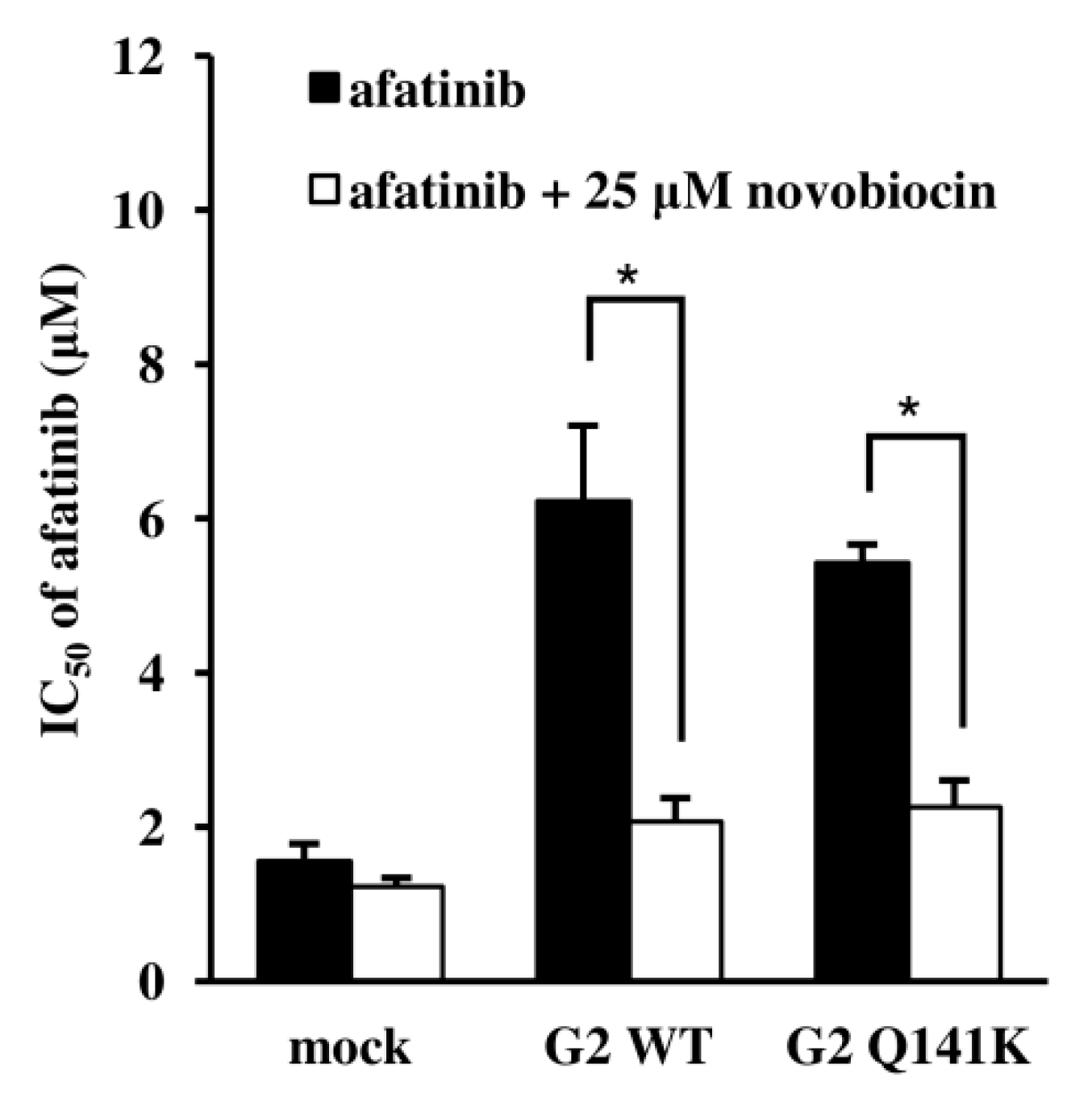
3.3. Effects of EGFR TKIs on ABCG2 Transport Activity
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Nakatomi, K.; Yoshikawa, M.; Oka, M.; Ikegami, Y.; Hayasaka, S.; Sano, K.; Shiozawa, K.; Kawabata, S.; Soda, H.; Ishikawa, T.; et al. Transport of 7-Ethyl-10-Hydroxycamptothecin (SN-38) by Breast Cancer Resistance Protein ABCG2 in Human Lung Cancer Cells. Biochem. Biophys. Res. Commun. 2001, 288, 827–832. [Google Scholar] [CrossRef]
- Brendel, C.; Scharenberg, C.; Dohse, M.; Robey, R.W.; Bates, S.E.; Shukla, S.; Ambudkar, S.V.; Wang, Y.; Wennemuth, G.; Burchert, A.; et al. Imatinib Mesylate and Nilotinib (AMN107) Exhibit High-Affinity Interaction with ABCG2 on Primitive Hematopoietic Stem Cells. Leukemia 2007, 21, 1267–1275. [Google Scholar] [CrossRef]
- Chen, Z.S.; Robey, R.W.; Belinsky, M.G.; Shchaveleva, I.; Ren, X.Q.; Sugimoto, Y.; Ross, D.D.; Bates, S.E.; Kruh, G.D. Transport of Methotrexate, Methotrexate Polyglutamates, and 17β-Estradiol 17-(β-D-Glucuronide) by ABCG2: Effects of Acquired Mutations at R482 on Methotrexate Transport. Cancer Res. 2003, 63, 4048–4054. [Google Scholar]
- Houghton, P.J.; Germain, G.S.; Harwood, F.C.; Schuetz, J.D.; Stewart, C.F.; Buchdunger, E.; Traxler, P. Imatinib Mesylate is a Potent Inhibitor of the ABCG2 (BCRP) Transporter and Reverses Resistance to Topotecan and SN-38 in Vitro. Cancer Res. 2004, 64, 2333–2337. [Google Scholar] [CrossRef]
- Tamura, A.; Wakabayashi, K.; Onishi, Y.; Takeda, M.; Ikegami, Y.; Sawada, S.; Tsuji, M.; Matsuda, Y.; Ishikawa, T. Re-Evaluation and Functional Classification of Non-Synonymous Single Nucleotide Polymorphisms of the Human ATP-Binding Cassette Transporter ABCG2. Cancer Sci. 2007, 98, 231–239. [Google Scholar] [CrossRef]
- Durmus, S.; Sparidans, R.W.; Esch, A.; Wagenaar, E.; Beijnen, J.H.; Schinkel, A.H. Breast Cancer Resistance Protein (BCRP/ABCG2) and P-Glycoprotein (P-GP/ABCB1) Restrict Oral Availability and Brain Accumulation of the PARP Inhibitor Rucaparib (AG-014699). Pharm. Res. 2015, 32, 37–46. [Google Scholar] [CrossRef]
- Birmingham, B.K.; Bujac, S.R.; Elsby, R.; Azumaya, C.T.; Wei, C.; Chen, Y.; Mosqueda-Garcia, R.; Ambrose, H.J. Impact of ABCG2 and SLCO1B1 Polymorphisms on Pharmacokinetics of Rosuvastatin, Atorvastatin and Simvastatin Acid in Caucasian and Asian Subjects: A Class Effect? Eur. J. Clin. Pharmacol. 2015, 71, 341–355. [Google Scholar] [CrossRef]
- Hegedus, C.; Özvegy-Laczka, C.; Apáti, A.Á.; Magócsi, M.; Német, K.; Orfi, L.; Kéri, G.; Katona, M.; Takáts, Z.; Váradi, A.; et al. Interaction of Nilotinib, Dasatinib and Bosutinib with ABCB1 and ABCG2: Implications for Altered Anti-Cancer Effects and Pharmacological Properties. Br. J. Pharmacol. 2009, 158, 1153–1164. [Google Scholar] [CrossRef]
- Inoue, Y.; Ikegami, Y.; Sano, K.; Suzuki, T.; Yoshida, H.; Nakamura, Y.; Nakagawa, H.; Ishikawa, T. Gefitinib Enhances the Antitumor Activity of CPT-11 in Vitro and in Vivo by Inhibiting ABCG2 but not ABCB1: A New Clue to Circumvent Gastrointestinal Toxicity Risk. Chemotherapy 2014, 59, 260–272. [Google Scholar] [CrossRef]
- Chen, Y.J.; Huang, W.C.; Wei, Y.L.; Hsu, S.C.; Yuan, P.; Lin, H.Y.; Wistuba, I.I.; Lee, J.J.; Yen, C.J.; Su, W.C.; et al. Elevated BCRP/ABCG2 Expression Confers Acquired Resistance to Gefitinib in Wild-Type EGFR-Expressing Cells. PLoS ONE 2011, 6, e21428. [Google Scholar] [CrossRef]
- Lemos, C.; Kathmann, I.; Giovannetti, E.; Calhau, C.; Jansen, G.; Peters, G.J. Impact of Cellular Folate Status and Epidermal Growth Factor Receptor Expression on BCRP/ABCG2-Mediated Resistance to Gefitinib and Erlotinib. Br. J. Cancer 2009, 100, 1120–1127. [Google Scholar] [CrossRef]
- Liang, S.C.; Yang, C.Y.; Tseng, J.Y.; Wang, H.L.; Tung, C.Y.; Liu, H.W.; Chen, C.Y.; Yeh, Y.C.; Chou, T.Y.; Yang, M.H.; et al. ABCG2 Localizes to the Nucleus and Modulates CDH1 Expression in Lung Cancer Cells. Neoplasia 2015, 17, 265–278. [Google Scholar] [CrossRef] [Green Version]
- Kawabata, S.; Oka, M.; Soda, H.; Shiozawa, K.; Nakatomi, K.; Tsurutani, J.; Nakamura, Y.; Doi, S.; Kitazaki, T.; Sugahara, K.; et al. Expression and Functional Analyses of Breast Cancer Resistance Protein in Lung Cancer. Clin. Cancer Res. 2003, 9, 3052–3057. [Google Scholar]
- Xiang, L.; Su, P.; Xia, S.; Liu, Z.; Wang, Y.; Gao, P.; Zhou, G. ABCG2 Is Associated with HER-2 Expression, Lymph Node Metastasis and Clinical Stage in Breast Invasive Ductal Carcinoma. Diagn. Pathol. 2011, 6, 90. [Google Scholar] [CrossRef]
- Low, S.K.; Fukunaga, K.; Takahashi, A.; Matsuda, K.; Hongo, F.; Nakanishi, H.; Kitamura, H.; Inoue, T.; Kato, Y.; Tomita, Y.; et al. Association Study of a Functional Variant on ABCG2 Gene with Sunitinib-Induced Severe Adverse Drug Reaction. PLoS ONE 2016, 11, e0148177. [Google Scholar] [CrossRef]
- Keskitalo, J.E.; Pasanen, M.K.; Neuvonen, P.J.; Niemi, M. Different Effects of the ABCG2 c.421C>A SNP on the Pharmacokinetics of Fluvastatin, Pravastatin and Simvastatin. Pharmacogenomics 2009, 10, 1617–1624. [Google Scholar] [CrossRef]
- Sakiyama, M.; Matsuo, H.; Takada, Y.; Nakamura, T.; Nakayama, A.; Takada, T.; Kitajiri, S.; Wakai, K.; Suzuki, H.; Shinomiya, N. Ethnic Differences in ATP-Binding Cassette Transporter, Sub-Family G, Member 2 (ABCG2/BCRP): Genotype Combinations and Estimated Functions. Drug Metab. Pharmacokinet. 2014, 29, 490–492. [Google Scholar] [CrossRef]
- Zhou, D.; Liu, Y.; Zhang, X.; Gu, X.; Wang, H.; Luo, X.; Zhang, J.; Zou, H.; Guan, M. Functional Polymorphisms of the ABCG2 Gene Are Associated with Gout Disease in the Chinese Han Male Population. Int. J. Mol. Sci. 2014, 15, 9149–9159. [Google Scholar] [CrossRef] [Green Version]
- Furukawa, T.; Wakabayashi, K.; Tamura, A.; Nakagawa, H.; Morishima, Y.; Osawa, Y.; Ishikawa, T. Major SNP (Q141K) Variant of Human ABC Transporter ABCG2 Undergoes Lysosomal and Proteasomal Degradations. Pharm. Res. 2009, 26, 469–479. [Google Scholar] [CrossRef]
- Cusatis, G.; Gregorc, V.; Li, J.; Spreafico, A.; Ingersoll, R.G.; Verweij, J.; Ludovini, V.; Villa, E.; Hidalgo, M.; Sparreboom, A.; et al. Pharmacogenetics of ABCG2 and Adverse Reactions to Gefitinib. J. Natl. Cancer Inst. 2006, 98, 1739–1742. [Google Scholar] [CrossRef]
- Wakabayashi-Nakao, K.; Tamura, A.; Koshiba, S.; Toyoda, Y.; Nakagawa, H.; Ishikawa, T. Production of Cells with Targeted Integration of Gene Variants of Human ABC Transporter for Stable and Regulated Expression Using the Flp Recombinase System. Methods Mol. Biol. 2010, 648, 139–159. [Google Scholar]
- Shi, Z.; Parmar, S.; Peng, X.X.; Shen, T.; Robey, R.W.; Bates, S.E.; Fu, L.W.; Shao, Y.; Chen, Y.M.; Zang, F.; et al. The Epidermal Growth Factor Tyrosine Kinase Inhibitor AG1478 and Erlotinib Reverse ABCG2-Mediated Drug Resistance. Oncol. Rep. 2009, 21, 483–489. [Google Scholar] [CrossRef]
- Gao, F.; Zhang, Y.; Wang, S.; Liu, Y.; Zheng, L.; Yang, J.; Huang, W.; Ye, Y.; Luo, W.; Xiao, D. Hes1 is Involved in the Self-Renewal and Tumourigenicity of Stem-like Cancer Cells in Colon Cancer. Sci. Rep. 2014, 4. [Google Scholar] [CrossRef]
- Kosztyu, P.; Bukvova, R.; Dolezel, P.; Mlejnek, P. Resistance to Daunorubicin, Imatinib, or Nilotinib Depends on Expression Levels of ABCB1 and ABCG2 in Human Leukemia Cells. Chem. Biol. Interact. 2014, 219, 203–210. [Google Scholar] [CrossRef]
- Awasthi, S.; Nair, M.; Awasthi, Y.; Lacko, A.; Singhal, J.; Singhal, S. Doxorubicin Transport by RALBP1 and ABCG2 in Lung and Breast Cancer. Int. J. Oncol. 2014, 1, 717–725. [Google Scholar] [CrossRef]
- Pavek, P. Human Breast Cancer Resistance Protein: Interactions with Steroid Drugs, Hormones, the Dietary Carcinogen 2-Amino-1-Methyl-6-Phenylimidazo(4,5-b)Pyridine, and Transport of Cimetidine. J. Pharmacol. Exp. Ther. 2004, 312, 144–152. [Google Scholar] [CrossRef]
- Herwaarden, A.E.; Wagenaar, E.; Karnekamp, B.; Merino, G.; Jonker, J.W.; Schinkel, A.H. Breast Cancer Resistance Protein (Bcrp1/Abcg2) Reduces Systemic Exposure of the Dietary Carcinogens Aflatoxin B1, IQ and Trp-P-1 but Also Mediates Their Secretion into Breast Milk. Carcinogenesis 2006, 27, 123–130. [Google Scholar] [CrossRef]
- Blazquez, A.G.; Briz, O.; Romero, M.R.; Rosales, R.; Monte, M.J.; Vaquero, J.; Macias, R.I.R.; Cassio, D.; Marin, J.J.G. Characterization of the Role of ABCG2 as a Bile Acid Transporter in Liver and Placenta. Mol. Pharmacol. 2012, 81, 273–283. [Google Scholar] [CrossRef]
- Yamagishi, K.; Tanigawa, T.; Kitamura, A.; Köttgen, A.; Folsom, A.R.; Iso, H. The Rs2231142 Variant of the ABCG2 Gene is Associated with Uric Acid Levels and Gout among Japanese People. Rheumatology 2010, 49, 1461–1465. [Google Scholar] [CrossRef]
- Ishikawa, T.; Tamura, A.; Saito, H.; Wakabayashi, K.; Nakagawa, H. Pharmacogenomics of the Human ABC Transporter ABCG2: From Functional Evaluation to Drug Molecular Design. Naturwissenschaften 2005, 451–463. [Google Scholar] [CrossRef]
- Sparreboom, A.; Gelderblom, H.; Marsh, S.; Ahluwalia, R.; Obach, R.; Principe, P.; Twelves, C.; Verweij, J.; McLeod, H.L. Diflomotecan Pharmacokinetics in Relation to ABCG2 421C>A Genotype. Clin. Pharmacol. Ther. 2004, 76, 38–44. [Google Scholar] [CrossRef]
- Li, J.; Cusatis, G.; Brahmer, J.; Sparreboom, A.; Robey, R.W.; Bates, S.E.; Hidalgo, M.; Baker, S.D. Association of Variant ABCG2 and the Pharmacokinetics of Epidermal Growth Factor Receptor Tyrosine Kinase Inhibitors in Cancer Patients. Cancer Biol. Ther. 2007, 6, 432–438. [Google Scholar] [CrossRef]
- Takahashi, N.; Miura, M.; Scott, S.A.; Kagaya, H.; Kameoka, Y.; Tagawa, H.; Saitoh, H.; Fujishima, N.; Yoshioka, T.; Hirokawa, M.; et al. Influence of CYP3A5 and Drug Transporter Polymorphisms on Imatinib Trough Concentration and Clinical Response among Patients with Chronic Phase Chronic Myeloid Leukemia. J. Hum. Genet. 2010, 55, 731–737. [Google Scholar] [CrossRef]
- Miura, Y.; Imamura, C.K.; Fukunaga, K.; Katsuyama, Y.; Suyama, K.; Okaneya, T.; Mushiroda, T.; Ando, Y.; Takano, T.; Tanigawara, Y. Sunitinib-Induced Severe Toxicities in a Japanese Patient with the ABCG2 421 AA Genotype. BMC Cancer 2014, 14. [Google Scholar] [CrossRef]
- Rudin, C.M.; Liu, W.; Desai, A.; Karrison, T.; Jiang, X.; Janisch, L.; Das, S.; Ramirez, J.; Poonkuzhali, B.; Schuetz, E.; et al. Pharmacogenomic and Pharmacokinetic Determinants of Erlotinib Toxicity. J. Clin. Oncol. 2008, 26, 1119–1127. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.D.; Lee, V.H.F.; Zhu, G.; Zou, B.; Ma, L.; Yan, H. Selectivity Profile of Afatinib for EGFR-Mutated Non-Small-Cell Lung Cancer. Mol. Biosyst. 2016, 12, 1552–1563. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Inoue, Y.; Morita, T.; Onozuka, M.; Saito, K.-i.; Sano, K.; Hanada, K.; Kondo, M.; Nakamura, Y.; Kishino, T.; Nakagawa, H.; et al. Impact of Q141K on the Transport of Epidermal Growth Factor Receptor Tyrosine Kinase Inhibitors by ABCG2. Cells 2019, 8, 763. https://doi.org/10.3390/cells8070763
Inoue Y, Morita T, Onozuka M, Saito K-i, Sano K, Hanada K, Kondo M, Nakamura Y, Kishino T, Nakagawa H, et al. Impact of Q141K on the Transport of Epidermal Growth Factor Receptor Tyrosine Kinase Inhibitors by ABCG2. Cells. 2019; 8(7):763. https://doi.org/10.3390/cells8070763
Chicago/Turabian StyleInoue, Yutaka, Takashi Morita, Mari Onozuka, Ken-ichi Saito, Kazumi Sano, Kazuhiko Hanada, Masami Kondo, Yoichi Nakamura, Tohru Kishino, Hiroshi Nakagawa, and et al. 2019. "Impact of Q141K on the Transport of Epidermal Growth Factor Receptor Tyrosine Kinase Inhibitors by ABCG2" Cells 8, no. 7: 763. https://doi.org/10.3390/cells8070763