Thymosin Beta-4 and Ciprofloxacin Adjunctive Therapy Improves Pseudomonas aeruginosa-Induced Keratitis

 , and
, and
Abstract
:1. Background
2. Methods
2.1. Experimental Animal Protocol
2.2. Ocular Response to Bacterial Infection
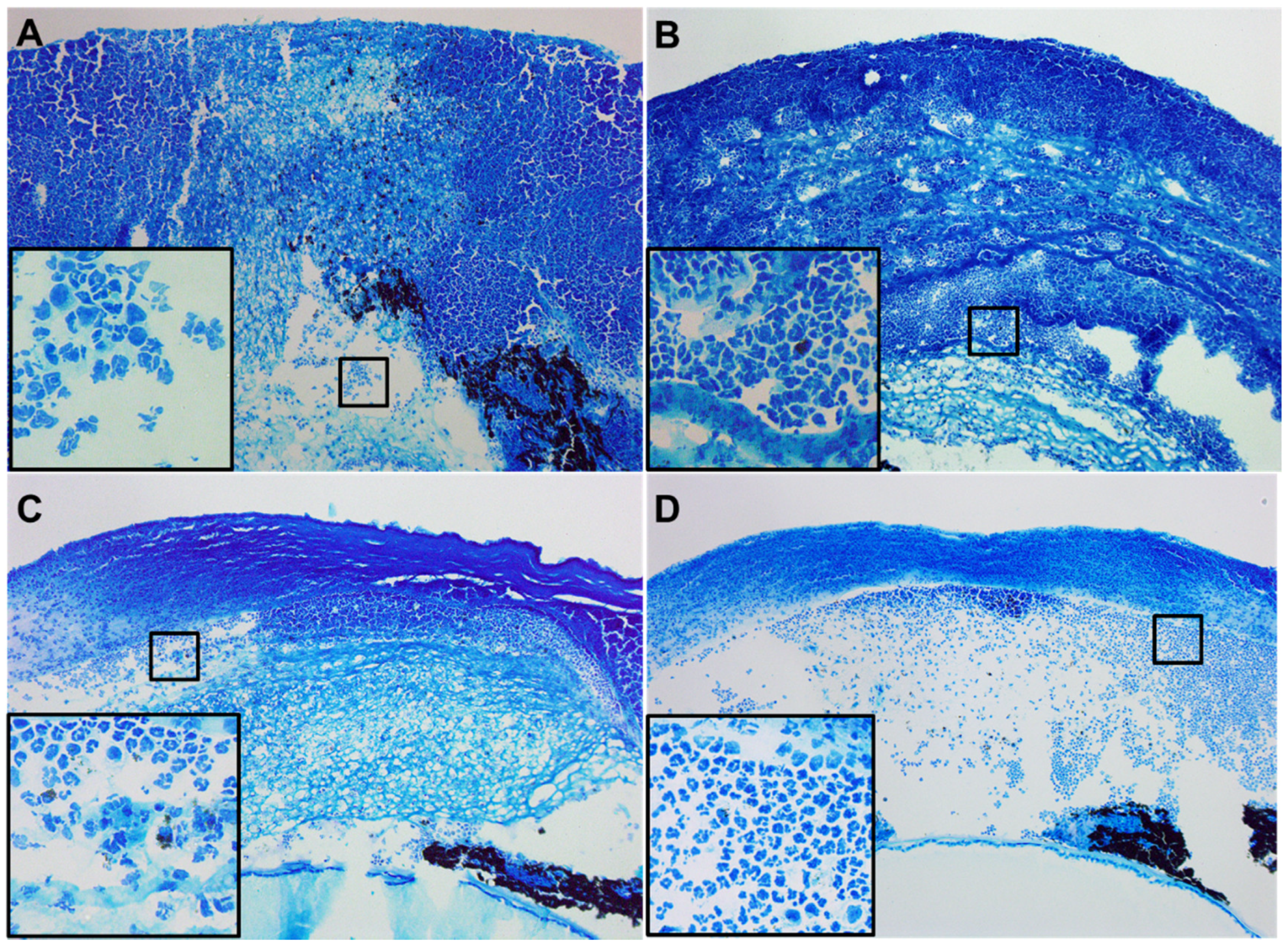
2.3. Histopathology
2.4. Bacterial Load
2.5. Measurement of ROS Levels
2.6. Myeloperoxidase (MPO) Assay
2.7. Greiss Reaction
2.8. Real-Time RT-PCR
2.9. ELISA Analysis
2.10. Statistical Analysis
3. Results
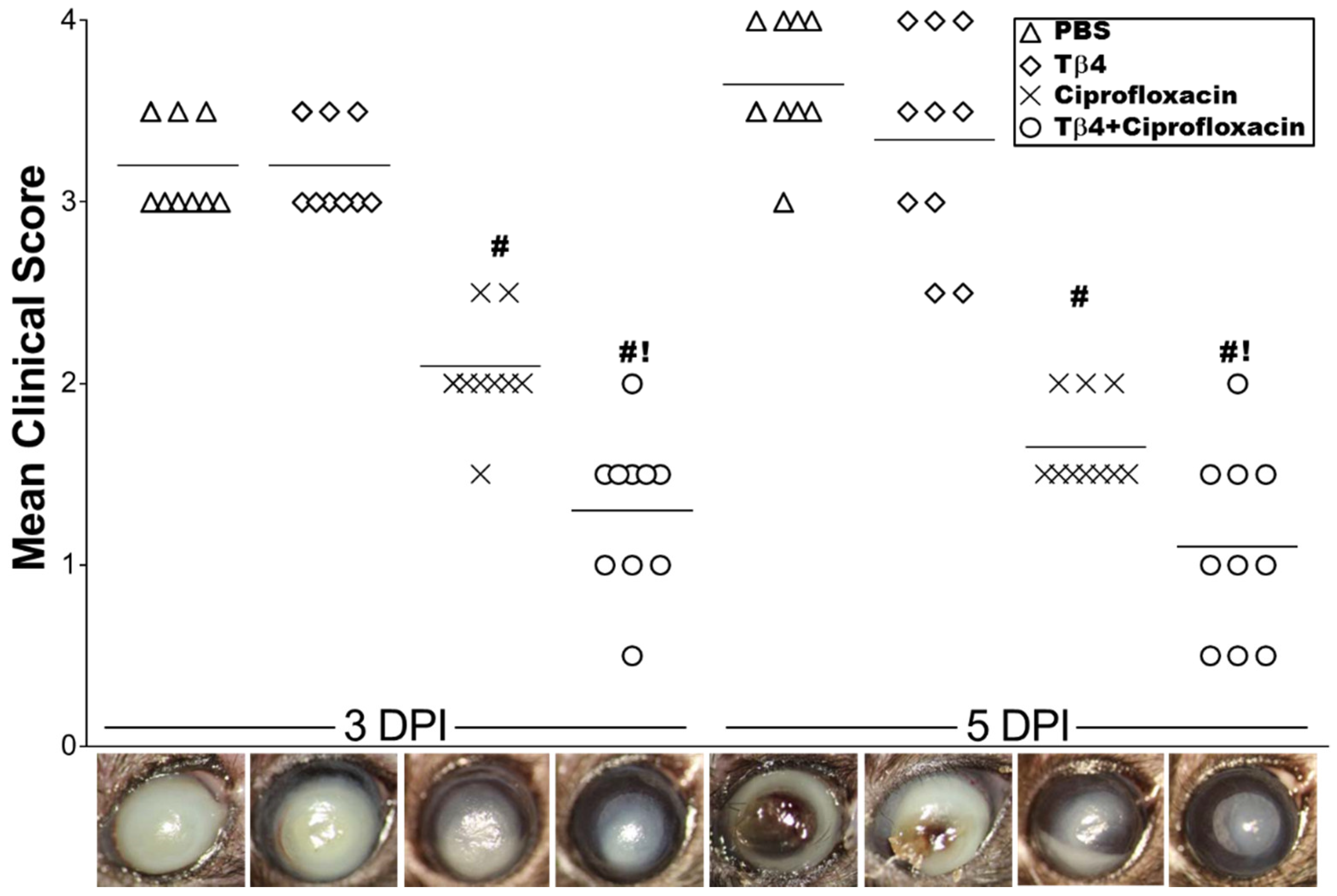
3.1. Tβ4 Adjunct Therapy Significantly Improves Disease Response
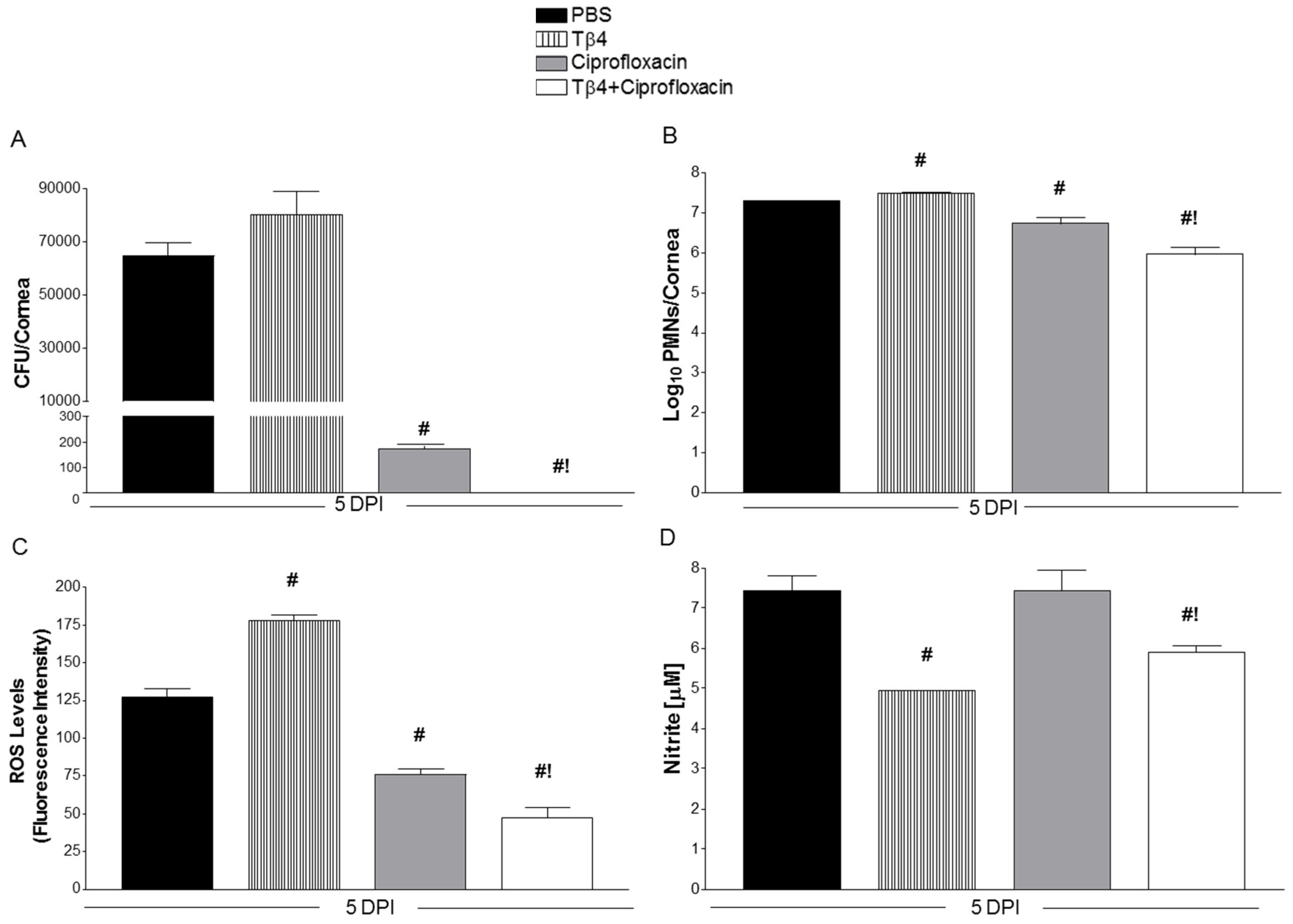
3.2. Tβ4 Adjunct Treatment Significantly Reduces Bacterial Load, ROS, MPO, and Nitrite Levels
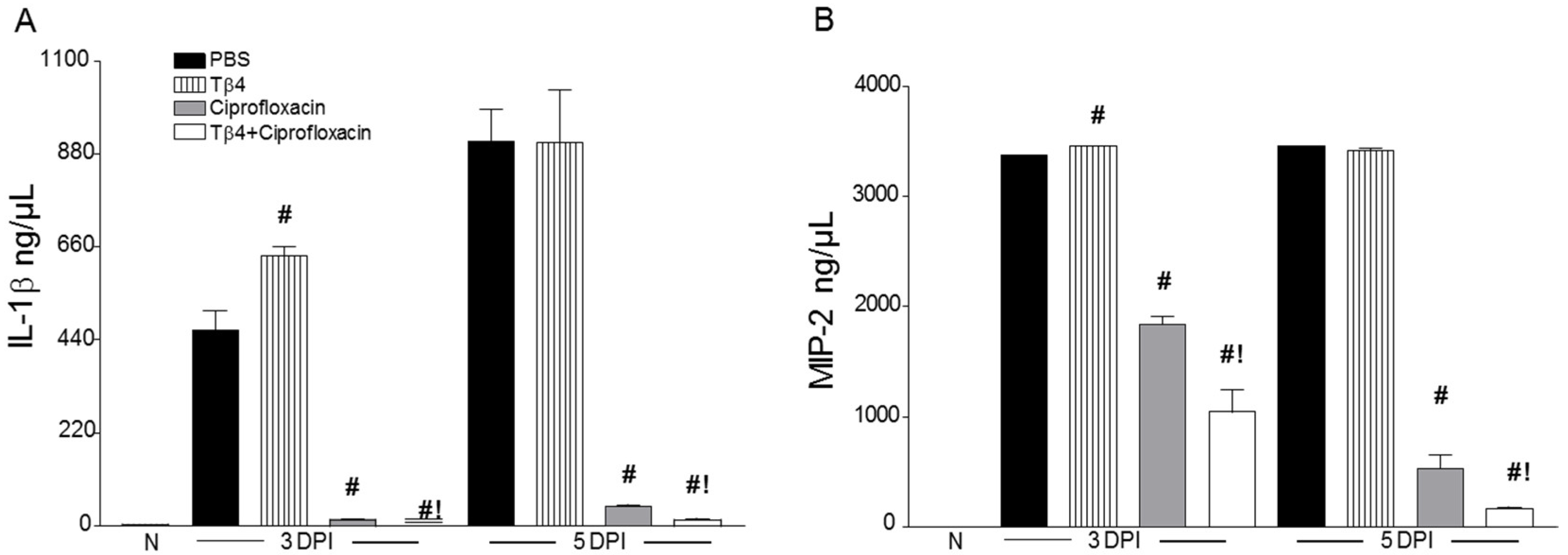
3.3. Tβ4 Adjunct Treatment Modulates Key Proinflammatory Mediators
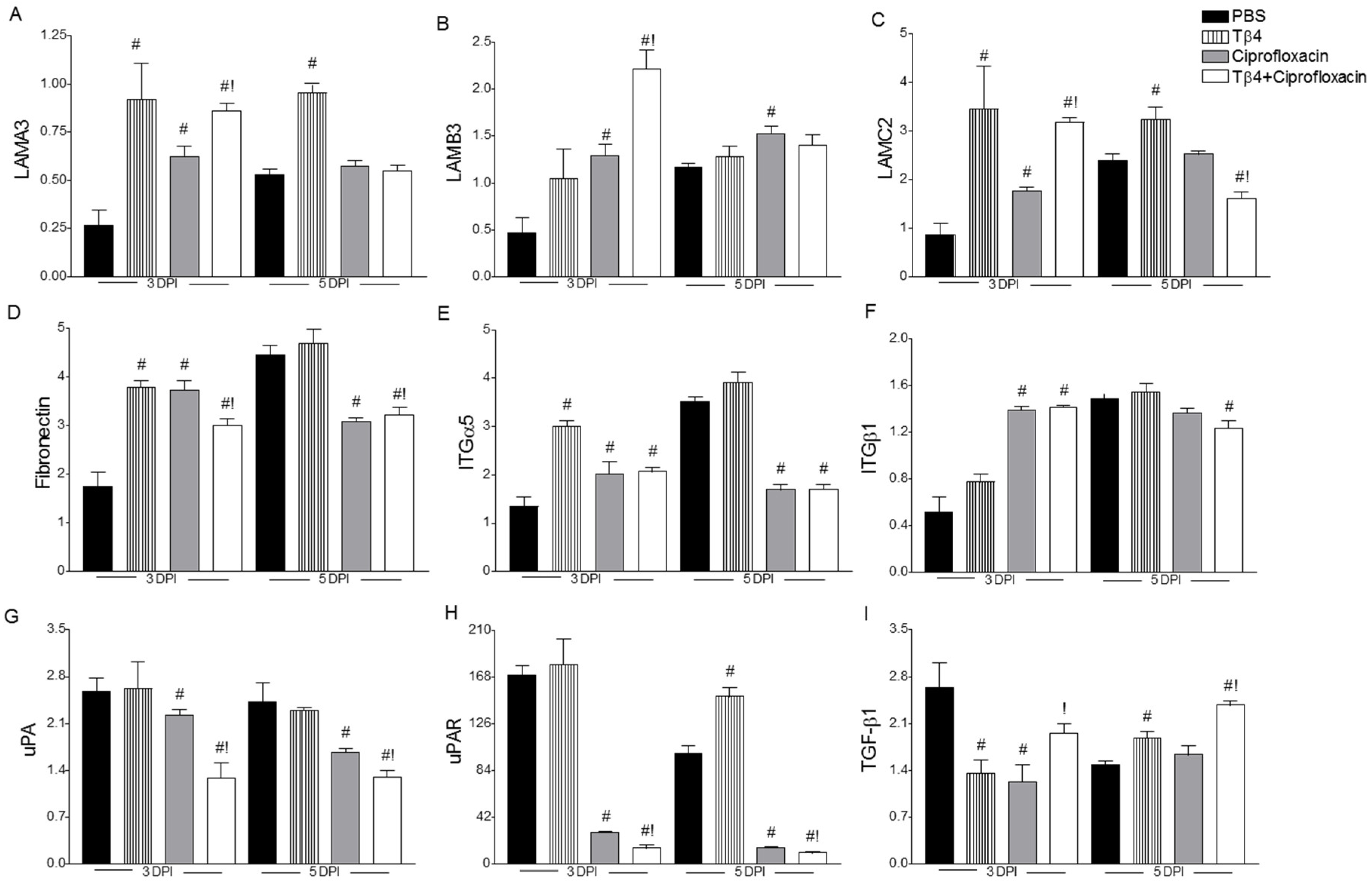
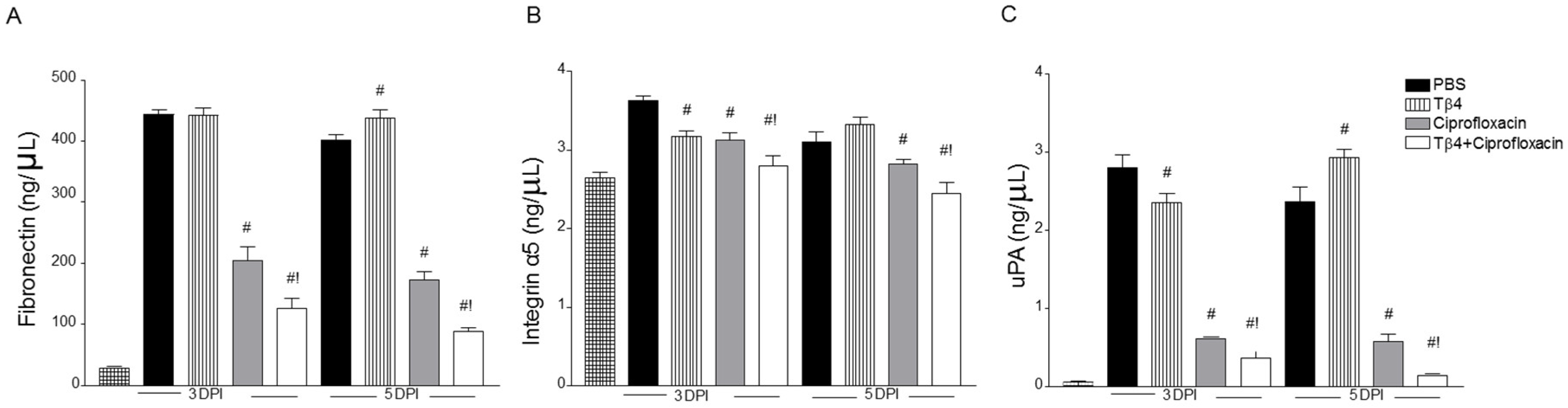
3.4. Treatment Activates Corneal Wound Healing Pathways after Infection
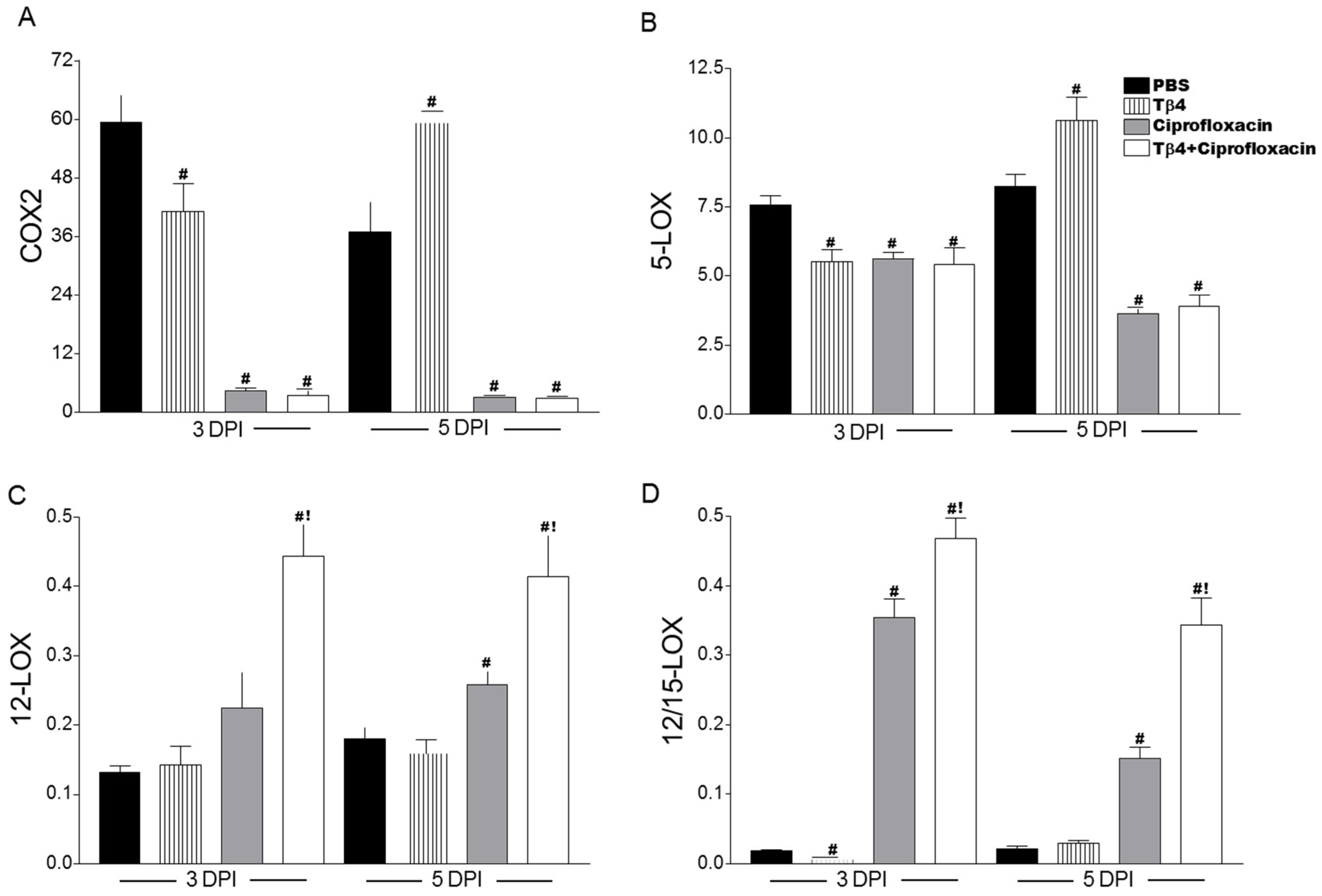
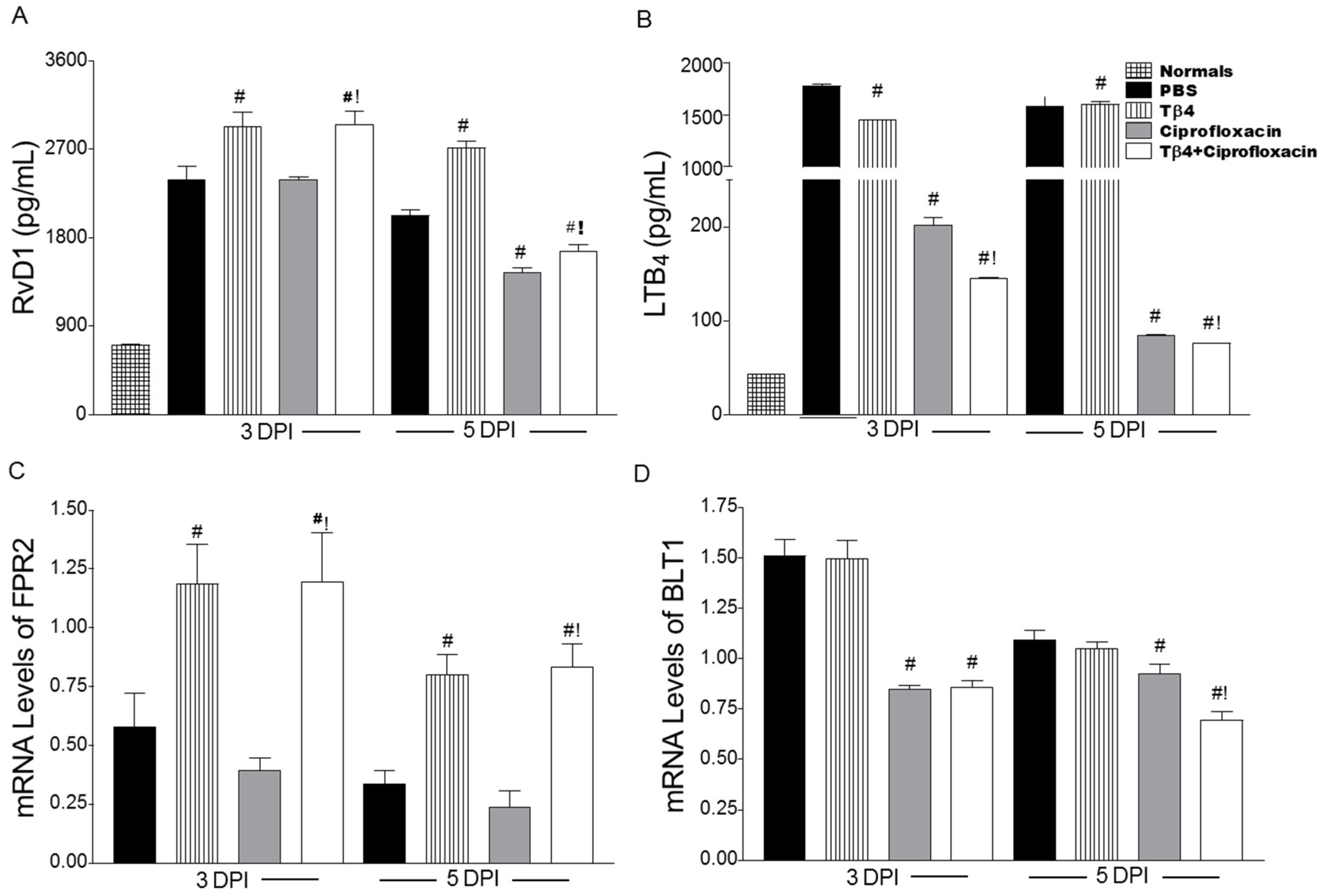
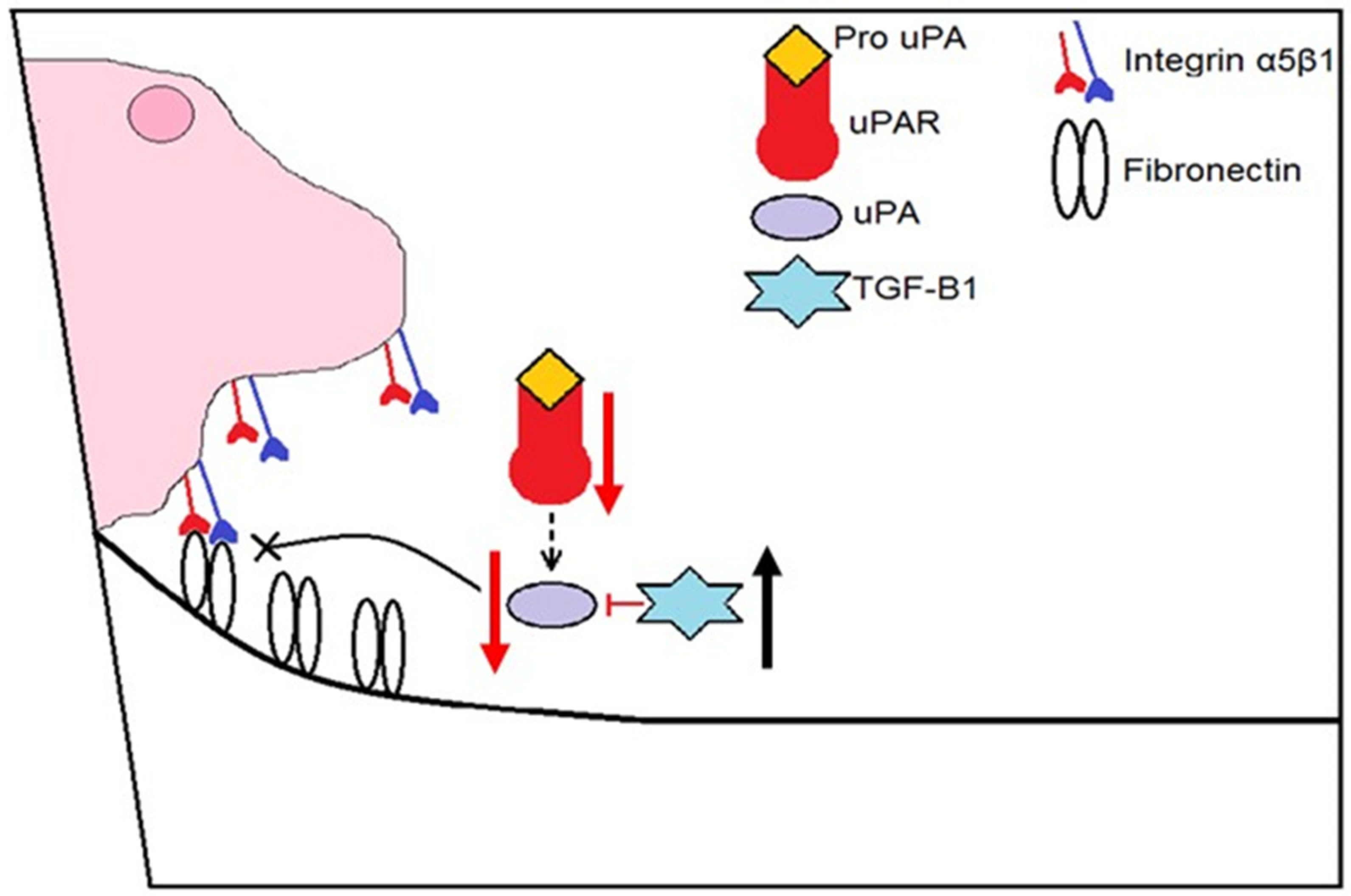
3.5. Tβ4 Influences a Lipid Mediator Circuit Known to Play a Role in Epithelial Wound Healing
4. Discussion
Supplementary Materials
Supplementary File 1Author Contributions
Funding
Conflicts of Interest
References
- Lichtinger, A.; Yeung, S.N.; Kim, P.; Amiran, M.D.; Iovieno, A.; Elbaz, U.; Ku, J.Y.; Wolff, R.; Rootman, D.S.; Slomovic, A.R. Shifting trends in bacterial keratitis in Toronto: An 11-year review. Ophthalmology 2012, 119, 1785–1790. [Google Scholar] [CrossRef] [PubMed]
- Chang, V.S.; Dhaliwal, D.K.; Raju, L.; Kowalski, R.P. Antibiotic Resistance in the Treatment of Staphylococcus aureus Keratitis: A 20-Year Review. Cornea 2015, 34, 698–703. [Google Scholar] [CrossRef] [PubMed]
- Wong, R.L.; Gangwani, R.A.; Yu, L.W.; Lai, J.S. New treatments for bacterial keratitis. J. Ophthalmol. 2012, 2012, 831502. [Google Scholar] [CrossRef] [PubMed]
- Lakhundi, S.; Siddiqui, R.; Khan, N.A. Pathogenesis of microbial keratitis. Microb. Pathog. 2017, 104, 97–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, J.W.; Fernandez, M.M.; Rebong, R.A.; Carlson, A.N.; Kim, T.; Afshari, N.A. Microbiological profiles of fungal keratitis: A 10-year study at a tertiary referral center. J. Ophthalmic Inflamm. Infect. 2016, 6, 5. [Google Scholar] [CrossRef] [PubMed]
- Rattanatam, T.; Heng, W.J.; Rapuano, C.J.; Laibson, P.R.; Cohen, E.J. Trends in contact lens-related corneal ulcers. Cornea 2001, 20, 290–294. [Google Scholar] [CrossRef] [PubMed]
- Sosa, A.B.; Epstein, S.P.; Asbell, P.A. Evaluation of toxicity of commercial ophthalmic fluoroquinolone antibiotics as assessed on immortalized corneal and conjunctival epithelial cells. Cornea 2008, 27, 930–934. [Google Scholar] [CrossRef] [PubMed]
- Oum, B.S.; Kim, N.M.; Lee, J.S.; Park, Y.M. Effects of fluoroquinolone eye solutions without preservatives on human corneal epithelial cells in vitro. Ophthalmic Res. 2014, 51, 216–223. [Google Scholar] [CrossRef] [PubMed]
- Wilhelmus, K.R. Indecision about corticosteroids for bacterial keratitis: An evidence-based update. Ophthalmology 2002, 109, 835–842; quiz 843. [Google Scholar] [CrossRef]
- Tallab, R.T.; Stone, D.U. Corticosteroids as a therapy for bacterial keratitis: An evidence-based review of ‘who, when and why’. Br. J. Ophthalmol. 2016, 100, 731–735. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, A.L.; Hannappel, E.; Sosne, G.; Kleinman, H.K. Thymosin beta4: A multi-functional regenerative peptide. Basic properties and clinical applications. Expert Opin. Boil. Ther. 2012, 12, 37–51. [Google Scholar] [CrossRef] [PubMed]
- Sosne, G.; Kleinman, H.K. Primary Mechanisms of Thymosin beta4 Repair Activity in Dry Eye Disorders and Other Tissue Injuries. Investig. Ophthalmol. Vis. Sci. 2015, 56, 5110–5117. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Kwon, J. Thymosin beta 4 improves dermal burn wound healing via downregulation of receptor of advanced glycation end products in db/db mice. Biochim. Biophys. Acta 2014, 1840, 3452–3459. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.J.; Wang, Q.; Ma, X.W.; Zhang, Z.; Zhang, W.; Xue, X.C.; Zhang, C.; Hao, Q.; Li, W.N.; Zhang, Y.Q.; et al. A novel dimeric thymosin beta 4 with enhanced activities accelerates the rate of wound healing. Drug Des. Dev. Ther. 2013, 7, 1075–1088. [Google Scholar] [CrossRef] [Green Version]
- Qiu, P.; Wheater, M.K.; Qiu, Y.; Sosne, G. Thymosin beta4 inhibits TNF-alpha-induced NF-kappaB activation, IL-8 expression, and the sensitizing effects by its partners PINCH-1 and ILK. FASEB J. 2011, 25, 1815–1826. [Google Scholar] [CrossRef] [PubMed]
- Sosne, G.; Albeiruti, A.R.; Hollis, B.; Siddiqi, A.; Ellenberg, D.; Kurpakus-Wheater, M. Thymosin beta4 inhibits benzalkonium chloride-mediated apoptosis in corneal and conjunctival epithelial cells in vitro. Exp. Eye Res. 2006, 83, 502–507. [Google Scholar] [CrossRef] [PubMed]
- Ho, J.H.; Chuang, C.H.; Ho, C.Y.; Shih, Y.R.; Lee, O.K.; Su, Y. Internalization is essential for the antiapoptotic effects of exogenous thymosin beta-4 on human corneal epithelial cells. Investig. Ophthalmol. Vis. Sci. 2007, 48, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.C.; Jean, D.; Proske, R.J.; Reins, R.Y.; McDermott, A.M. Ocular surface expression and in vitro activity of antimicrobial peptides. Curr. Eye Res. 2007, 32, 595–609. [Google Scholar] [CrossRef] [PubMed]
- Dunn, S.P.; Heidemann, D.G.; Chow, C.Y.; Crockford, D.; Turjman, N.; Angel, J.; Allan, C.B.; Sosne, G. Treatment of chronic nonhealing neurotrophic corneal epithelial defects with thymosin beta 4. Arch. Ophthalmol. 2010, 128, 636–638. [Google Scholar] [CrossRef] [PubMed]
- Sosne, G.; Dunn, S.P.; Kim, C. Thymosin beta4 significantly improves signs and symptoms of severe dry eye in a phase 2 randomized trial. Cornea 2015, 34, 491–496. [Google Scholar] [CrossRef] [PubMed]
- Sosne, G.; Ousler, G.W. Thymosin beta 4 ophthalmic solution for dry eye: A randomized, placebo-controlled, Phase II clinical trial conducted using the controlled adverse environment (CAE) model. Clin. Ophthalmol. 2015, 9, 877–884. [Google Scholar] [CrossRef] [PubMed]
- Rudner, X.L.; Kernacki, K.A.; Barrett, R.P.; Hazlett, L.D. Prolonged elevation of IL-1 in Pseudomonas aeruginosa ocular infection regulates macrophage-inflammatory protein-2 production, polymorphonuclear neutrophil persistence, and corneal perforation. J. Immunol. 2000, 164, 6576–6582. [Google Scholar] [CrossRef] [PubMed]
- Hazlett, L.D.; Moon, M.M.; Strejc, M.; Berk, R.S. Evidence for N-acetylmannosamine as an ocular receptor for P. aeruginosa adherence to scarified cornea. Investig. Ophthalmol. Vis. Sci. 1987, 28, 1978–1985. [Google Scholar]
- Keiichi, I.; Kenji, A.; Eiichi, T.; Shukkur, E.A.; Atsushi, S.; Noriko, S.; Ikuyo, I.; Mayuko, T.; Yuto, U.; Haruhiko, S.; et al. Increased lipid peroxidation in Down’s syndrome mouse models. J. Neurochem. 2009, 110, 1965–1976. [Google Scholar] [CrossRef] [Green Version]
- Kalyanaraman, B.; Darley-Usmar, V.; Davies, K.J.; Dennery, P.A.; Forman, H.J.; Grisham, M.B.; Mann, G.E.; Moore, K.; Roberts, L.J., 2nd; Ischiropoulos, H. Measuring reactive oxygen and nitrogen species with fluorescent probes: Challenges and limitations. Free Radic. Biol. Med. 2012, 52, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Szliter, E.A.; Lighvani, S.; Barrett, R.P.; Hazlett, L.D. Vasoactive intestinal peptide balances pro- and anti-inflammatory cytokines in the Pseudomonas aeruginosa-infected cornea and protects against corneal perforation. J. Immunol. 2007, 178, 1105–1114. [Google Scholar] [CrossRef] [PubMed]
- Williams, R.N.; Paterson, C.A.; Eakins, K.E.; Bhattacherjee, P. Quantification of ocular inflammation: Evaluation of polymorphonuclear leucocyte infiltration by measuring myeloperoxidase activity. Curr. Eye Res. 1982, 2, 465–470. [Google Scholar] [CrossRef] [PubMed]
- Berger, E.A.; McClellan, S.A.; Vistisen, K.S.; Hazlett, L.D. HIF-1alpha is essential for effective PMN bacterial killing, antimicrobial peptide production and apoptosis in Pseudomonas aeruginosa keratitis. PLoS Pathog. 2013, 9, e1003457. [Google Scholar] [CrossRef] [PubMed]
- Carion, T.W.; Greenwood, M.; Ebrahim, A.S.; Jerome, A.; Suvas, S.; Gronert, K.; Berger, E.A. Immunoregulatory role of 15-lipoxygenase in the pathogenesis of bacterial keratitis. FASEB J. 2018, 32, 5026–5038. [Google Scholar] [CrossRef] [PubMed]
- Roux, K.H. Optimization and troubleshooting in PCR. PCR Methods Appl. 1995, 4, S185–S194. [Google Scholar] [CrossRef] [PubMed]
- Carion, T.W.; McWhirter, C.R.; Grewal, D.K.; Berger, E.A. Efficacy of VIP as Treatment for Bacteria-Induced Keratitis Against Multiple Pseudomonas aeruginosa Strains. Investig. Ophthalmol. Vis. Sci. 2015, 56, 6932–6940. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, A.M.; Twining, S.S.; Warejcka, D.J.; Tall, E.; Masur, S.K. Urokinase Receptor Cleavage: A Crucial Step in Fibroblast-to-Myofibroblast Differentiation. Mol. Boil. Cell 2007, 18, 2716–2727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sosne, G.; Qiu, P.; Ousler, G.W., 3rd; Dunn, S.P.; Crockford, D. Thymosin beta4: A potential novel dry eye therapy. Ann. N. Y. Acad. Sci. 2012, 1270, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Sosne, G.; Szliter, E.A.; Barrett, R.; Kernacki, K.A.; Kleinman, H.; Hazlett, L.D. Thymosin beta 4 promotes corneal wound healing and decreases inflammation in vivo following alkali injury. Exp. Eye Res. 2002, 74, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Tantos, A.; Szabo, B.; Lang, A.; Varga, Z.; Tsylonok, M.; Bokor, M.; Verebelyi, T.; Kamasa, P.; Tompa, K.; Perczel, A.; et al. Multiple fuzzy interactions in the moonlighting function of thymosin-beta4. Intrinsically Disord. Proteins 2013, 1, e26204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gronert, K.; Maheshwari, N.; Khan, N.; Hassan, I.R.; Dunn, M.; Laniado Schwartzman, M. A role for the mouse 12/15-lipoxygenase pathway in promoting epithelial wound healing and host defense. J. Biol. Chem. 2005, 280, 15267–15278. [Google Scholar] [CrossRef] [PubMed]
- Ock, M.S.; Song, K.S.; Kleinman, H.; Cha, H.J. Thymosin beta4 stabilizes hypoxia-inducible factor-1alpha protein in an oxygen-independent manner. Ann. N. Y. Acad. Sci. 2012, 1269, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Fitsialos, G.; Bourget, I.; Augier, S.; Ginouves, A.; Rezzonico, R.; Odorisio, T.; Cianfarani, F.; Virolle, T.; Pouyssegur, J.; Meneguzzi, G.; et al. HIF1 transcription factor regulates laminin-332 expression and keratinocyte migration. J. Cell Sci. 2008, 121, 2992–3001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, M.; Yano, W.; Kondo, S.; Hattori, Y.; Yamada, N.; Yanai, R.; Nishida, T. Up-regulation of urokinase-type plasminogen activator in corneal epithelial cells induced by wounding. Investig. Ophthalmol. Vis. Sci. 2003, 44, 3332–3338. [Google Scholar] [CrossRef]
- Sugioka, K.; Mishima, H.; Kodama, A.; Itahashi, M.; Fukuda, M.; Shimomura, Y. Regulatory Mechanism of Collagen Degradation by Keratocytes and Corneal Inflammation: The Role of Urokinase-Type Plasminogen Activator. Cornea 2016, 35 (Suppl. 1), S59–S64. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Nucleotide Sequence | Primer |
|---|---|---|
| b-actin | 5′- GAT TAC TGC TCT GGC TCC TAG C -3′ | F |
| 5′- GAC TCA TCG TAC TCC TGC TTG C -3′ | R | |
| TNF-a | 5′- ACC CTC ACA CTC AGA TCA TCT T -3′ | F |
| 5′- GGT TGT CTT TGA GAT CCA TGC -3′ | R | |
| IL-1b | 5′- CGC AGC AGC ACA TCA ACA AGA GC -3′ | F |
| 5′- TGT CCT CAT CCT GGA AGG TCC ACG -3′ | R | |
| iNOS | 5′- ACA GGA GAA GGG GAC GAA CT -3′ | F |
| 5′- TGT TGC ATT GGA AGT GAA GC -3′ | R | |
| CXCL2/MIP-2 | 5′- TGT CAA TGC CTG AAG ACC CTG CC -3′ | F |
| 5′- AAC TTT TTG ACC GCC CTT GAG AGT GG -3′ | R | |
| LAMA3 | 5′- GAC CTA CGT TCC ATC CTC CA -3′ | F |
| 5′- CTG GCT TTT GTC CAT CTG CT -3′ | R | |
| LAMB3 | 5′- TTG ATT GAG CGC TCT TCT GA -3′ | F |
| 5′- ATG CAG GGA TAG CTG ATG CT -3′ | R | |
| LAMC2 | 5′- TCC CCA GCT GAG TTA TTT CG -3′ | F |
| 5′- CTG GCA GAA TTG TCC CTT GT -3′ | R | |
| Fibronectin | 5′- GAA GGT TTG CAA CCC ACT GT -3′ | F |
| 5′- CAT CCT CAG GGC TCG AGT AG -3′ | R | |
| ITG α5 | 5′- TGG ACC AAG ACG GCT ACA AT -3′ | F |
| 5′- ATT GCC ATC CAG ATC TCG TC -3′ | R | |
| ITG β1 | 5′- GGG CAC ACT GTC TGG AAA CT -3′ | F |
| 5′- TCG TCC ATT TTC TCC TGT CC -3′ | R | |
| uPA | 5′- GCC TGC TGT CCT TCA GAA AC -3′ | F |
| 5′- AAG AGA GCA GTC ATG CAC CA -3′ | R | |
| uPAR | 5′- TTT GGA CCA GAG CTG TGA GA -3′ | F |
| 5′- CAC CAT TGC AGT GGG TGT AG -3′ | R | |
| TGF- β1 | 5′- TCT CTG CTC TCT GCT GCT GAT ATG C -3′ | F |
| 5′- AGG ACA AAT GGC TCT GAC ACA GTA CC -3′ | R | |
| COX-2 | 5′- TGA GCA ACT ATT CCA AAC CAG C -3′ | F |
| 5′- GCA CGT AGT CTT CGA TCA CTA TC -3′ | R | |
| 5-LOX | 5′- ACT ACA TCT ACC TCA GCC TCA TT -3′ | F |
| 5′- GGT GAC ATC GTA GGA GTC CAC -3′ | R | |
| 12-LOX | 5′- TAC CCT CCT GAG AAG CTG GA -3′ | F |
| 5′- TCA TCT TCC TGC CAA CAC TG -3′ | R | |
| 12/15-LOX | 5′- GCG ACG CTG CCC AAT CCT AAT C -3′ | F |
| 5′- ATA TGG CCA CGC TGT TTT CTA CC -3′ | R | |
| FPR2 | 5′- CCT TGG ACC GCT GTA TTT GT -3′ | F |
| 5′- CCC CAG GAT ACA AAG CTC AA -3′ | R | |
| BLT1 | 5′- GCA TGT ATG CCA GTG TCC TG -3′ | F |
| 5′- AAA AGA CAC CAC CCA GAT GC -3′ | R |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carion, T.W.; Ebrahim, A.S.; Kracht, D.; Agrawal, A.; Strand, E.; Kaddurah, O.; McWhirter, C.R.; Sosne, G.; Berger, E.A. Thymosin Beta-4 and Ciprofloxacin Adjunctive Therapy Improves Pseudomonas aeruginosa-Induced Keratitis. Cells 2018, 7, 145. https://doi.org/10.3390/cells7100145
Carion TW, Ebrahim AS, Kracht D, Agrawal A, Strand E, Kaddurah O, McWhirter CR, Sosne G, Berger EA. Thymosin Beta-4 and Ciprofloxacin Adjunctive Therapy Improves Pseudomonas aeruginosa-Induced Keratitis. Cells. 2018; 7(10):145. https://doi.org/10.3390/cells7100145
Chicago/Turabian StyleCarion, Thomas W, Abdul Shukkur Ebrahim, David Kracht, Aditya Agrawal, Eliisa Strand, Omar Kaddurah, Cody R. McWhirter, Gabriel Sosne, and Elizabeth A. Berger. 2018. "Thymosin Beta-4 and Ciprofloxacin Adjunctive Therapy Improves Pseudomonas aeruginosa-Induced Keratitis" Cells 7, no. 10: 145. https://doi.org/10.3390/cells7100145