
The Effects of Aging on Rod Bipolar Cell Ribbon Synapses

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Zebrafish Rearing
2.2. Retinal Bipolar Cell Isolation
2.3. Bipolar Cell Voltage Clamp Recordings
2.4. Laser-Scanning Confocal Microscopy
2.5. Immunohistochemistry (IHC)
2.6. Data Analysis
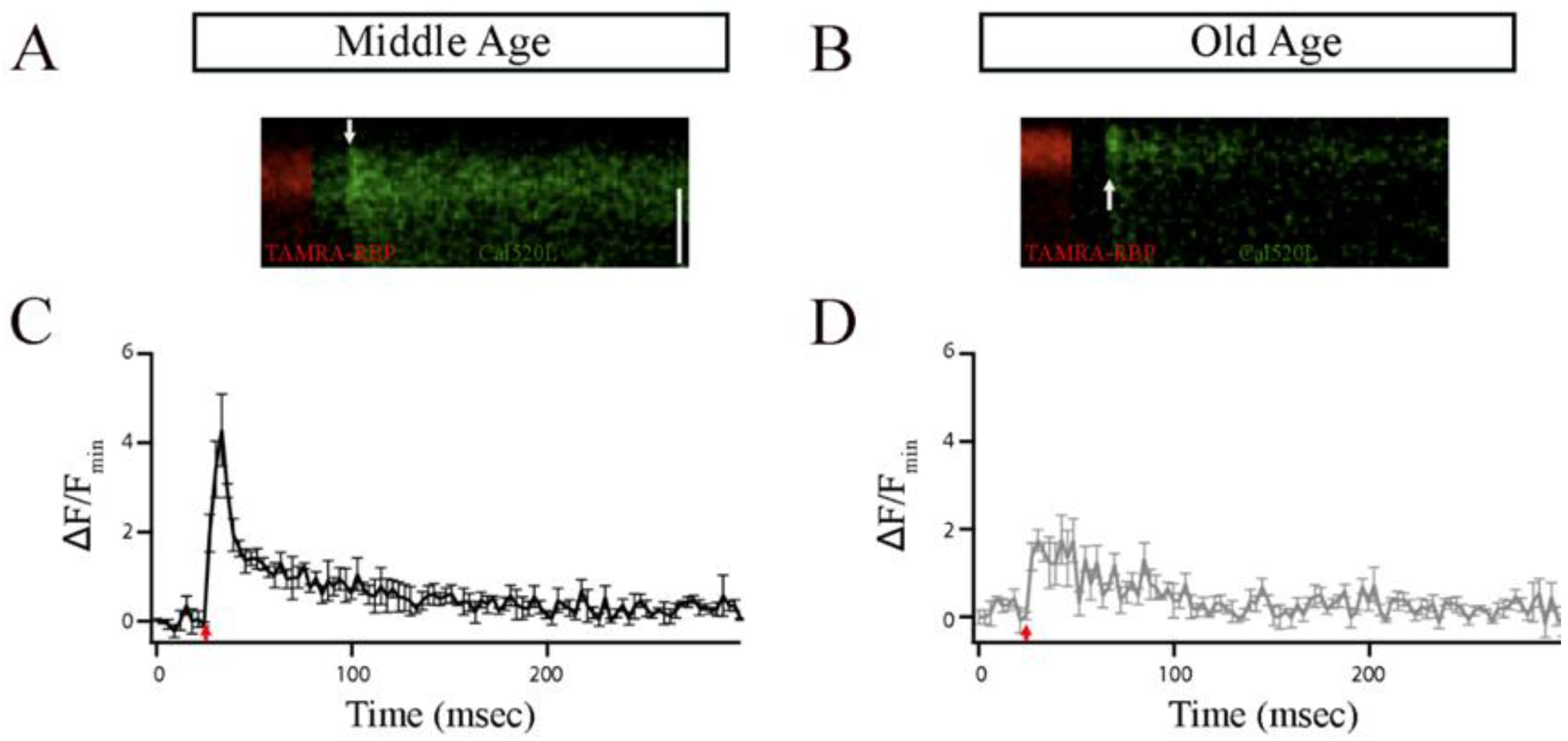
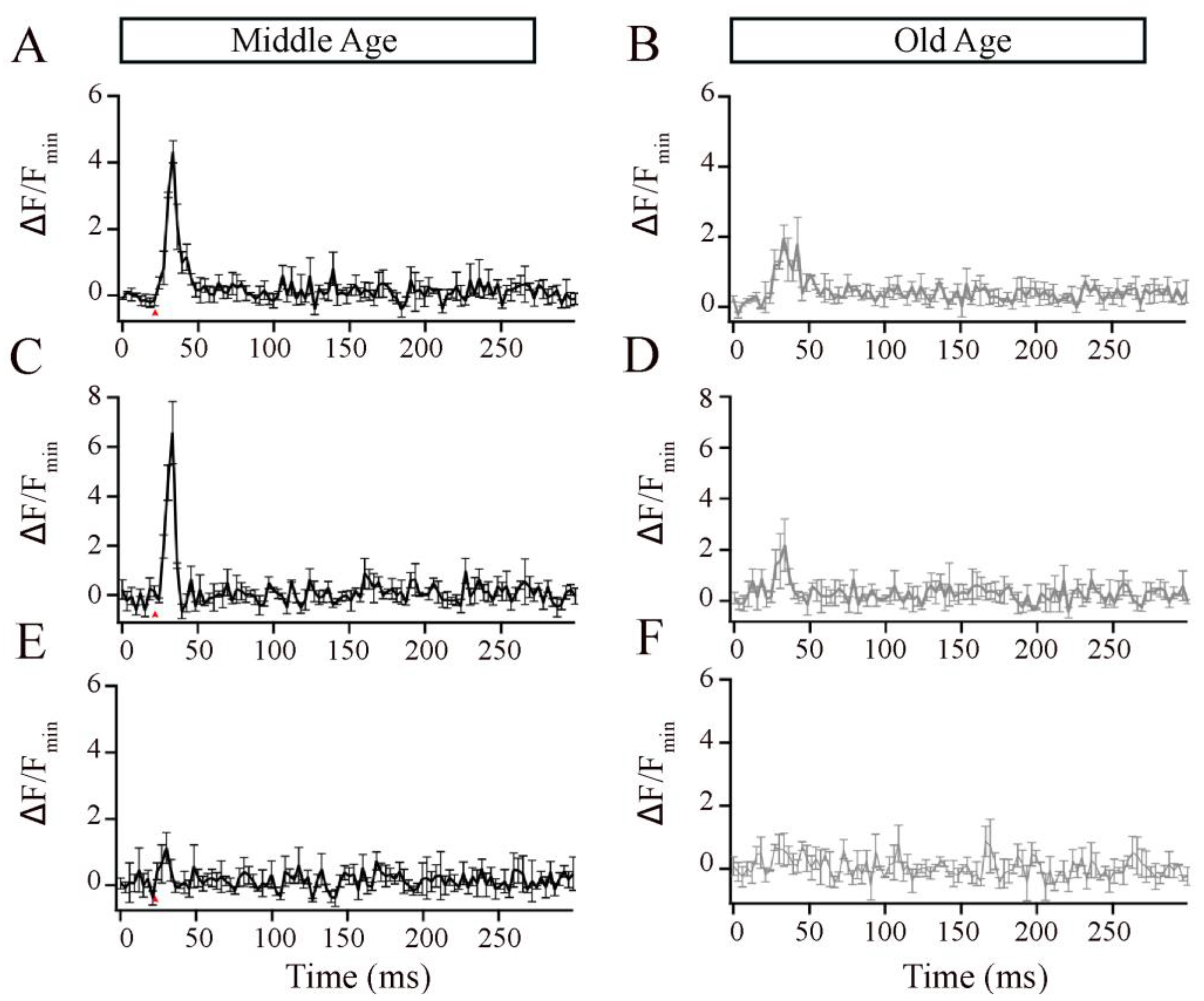
2.6.1. Analysis of x-t Scans
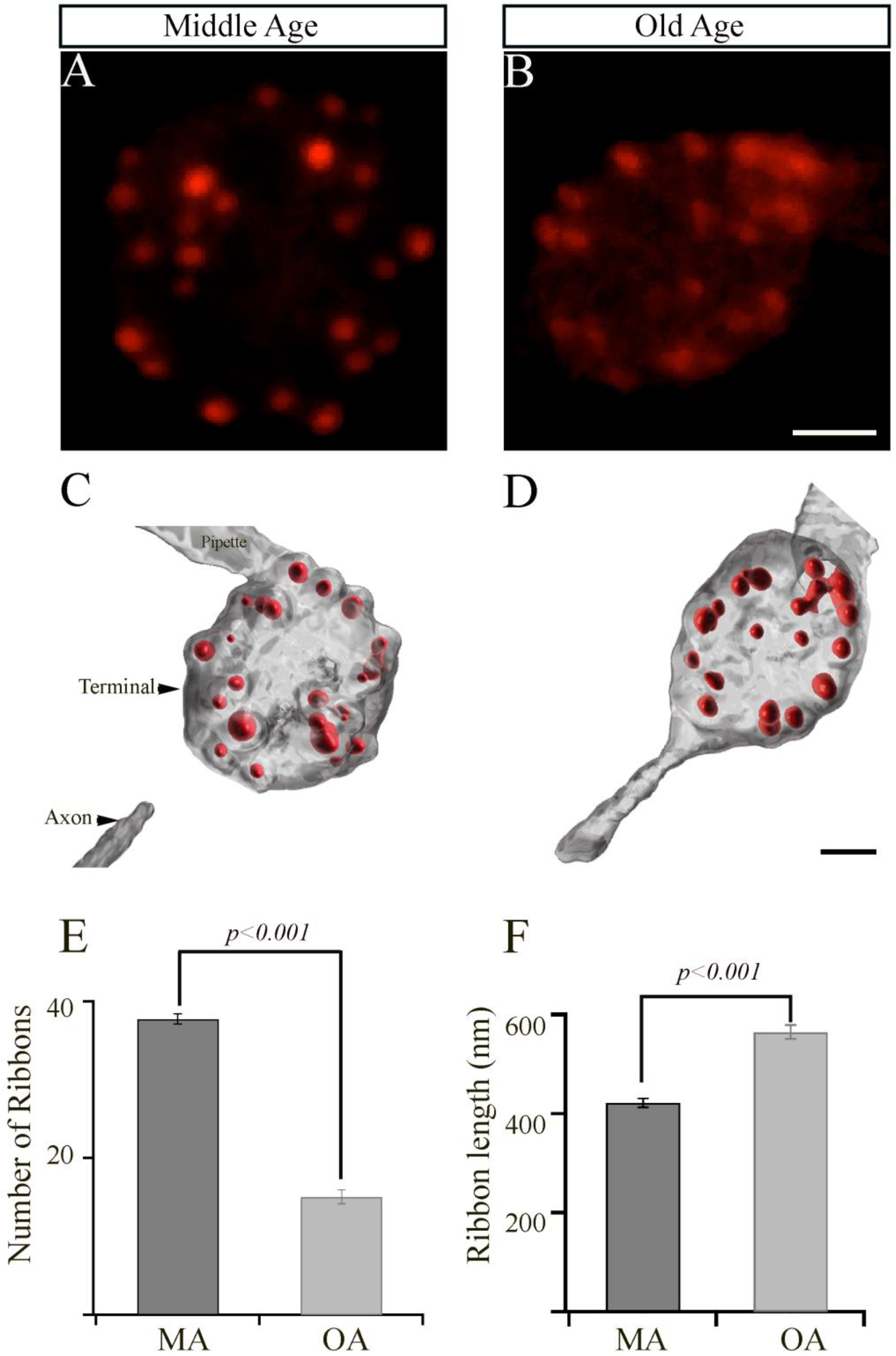
2.6.2. Analysis of Ribbon Synapses
2.6.3. Statistical Analyses
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Saftari, L.N.; Kwon, O.-S. Ageing vision and falls: A review. J. Physiol. Anthropol. 2018, 37, 11–25. [Google Scholar] [CrossRef]
- Owsley, C. Aging and vision. Vision Res. 2011, 51, 1610–1622. [Google Scholar] [CrossRef] [PubMed]
- Salvi, S.M.; Akhtar, S.; Currie, Z. Ageing changes in the eye. Postgrad. Med. J. 2006, 82, 581–587. [Google Scholar] [CrossRef] [PubMed]
- Cavazzana, A.; Röhrborn, A.; Garthus-Niegel, S.; Larsson, M.; Hummel, T.; Croy, I. Sensory-specific impairment among older people. An investigation using both sensory thresholds and subjective measures across the five senses. PLoS ONE 2018, 13, 202969–202984. [Google Scholar] [CrossRef] [PubMed]
- Jeng, J.-Y.; Ceriani, F.; Olt, J.; Brown, S.D.M.; Holley, M.C.; Bowl, M.R.; Johnson, S.L.; Marcotti, W. Pathophysiological changes in inner hair cell ribbon synapses in the ageing mammalian cochlea. J. Physiol. 2020, 598, 4339–4355. [Google Scholar] [CrossRef] [PubMed]
- Samuel, M.A.; Zhang, Y.; Meister, M.; Sanes, J.R. Age-related alterations in neurons of the mouse retina. J. Neurosci. 2011, 31, 16033–16044. [Google Scholar] [CrossRef]
- Marshall, J. The ageing retina: Physiology or pathology. Eye 1987, 1 Pt 2, 282–295. [Google Scholar] [CrossRef]
- Martins, R.R.; Zamzam, M.; Tracey-White, D.; Moosajee, M.; Thummel, R.; Henriques, C.M.; MacDonald, R.B. Müller Glia maintain their regenerative potential despite degeneration in the aged zebrafish retina. Aging Cell 2022, 21, 13597–13617. [Google Scholar] [CrossRef]
- Jackson, G.R.; Owsley, C.; Curcio, C.A. Photoreceptor degeneration and dysfunction in aging and age-related maculopathy. Ageing Res. Rev. 2002, 1, 381–396. [Google Scholar] [CrossRef]
- Szabadfi, K.; Estrada, C.; Fernandez-Villalba, E.; Tarragon, E.; Setalo, G., Jr.; Izura, V.; Reglodi, D.; Tamas, A.; Gabriel, R.; Herrero, M.T. Retinal aging in the diurnal Chilean rodent (Octodon degus): Histological, ultrastructural and neurochemical alterations of the vertical information processing pathway. Front. Cell Neurosci. 2015, 9, 126–140. [Google Scholar] [CrossRef]
- Chhetri, J.; Jacobson, G.; Gueven, N. Zebrafish-on the move towards ophthalmological research. Eye 2014, 28, 367–380. [Google Scholar] [CrossRef] [PubMed]
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The zebrafish reference genome sequence and its relationship to the human genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Tucci, V.; Kishi, S.; Zhdanova, I.V. Cognitive Aging in Zebrafish. PLoS ONE 2006, 1, 14–27. [Google Scholar] [CrossRef]
- Khor, E.-S.; Noor, S.M.; Wong, P.-F. Understanding the role of ztor in aging-related diseases using the zebrafish model. In Vivo 2019, 33, 1713–1720. [Google Scholar] [CrossRef]
- Gilbert, M.J.H.; Zerulla, T.C.; Tierney, K.B. Zebrafish (Danio rerio) as a model for the study of aging and exercise: Physical ability and trainability decrease with age. Exp. Gerontol. 2014, 50, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Kishi, S.; Uchiyama, J.; Baughman, A.M.; Goto, T.; Lin, M.C.; Tsai, S.B. The zebrafish as a vertebrate model of functional aging and very gradual senescence. Exp. Gerontol. 2003, 38, 777–786. [Google Scholar] [CrossRef] [PubMed]
- Murtha, J.M.; Keller, E.T. Characterization of the heat shock response in mature zebrafish (Danio rerio). Exp. Gerontol. 2003, 38, 683–691. [Google Scholar] [CrossRef]
- Rosa, C.E.D.; Kuradomi, R.Y.; Almeida, D.V.; Lannes, C.F.C.; Figueiredo, M.D.A.; Dytz, A.G.; Fonseca, D.B.; Marins, L.F. GH overexpression modifies muscle expression of anti-oxidant enzymes and increases spinal curvature of old zebrafish. Exp. Gerontol. 2010, 45, 449–456. [Google Scholar] [CrossRef]
- Yusuke, K.; Wang, W.; Yoji, I.; Kazutoshi, Y.; Shuichi, A.; Yutaka, S.; Shugo, W.; Shigeharu, K. Age-associated different transcriptome profiling in zebrafish and rat: Insight into diversity of vertebrate aging. Mar. Biotechnol. 2022, 24, 895–910. [Google Scholar]
- Sengupta, P. The Laboratory Rat: Relating Its Age with Human’s. Int. J. Prev. Med. 2013, 4, 624–630. [Google Scholar]
- Snellman, J.; Mehta, B.; Babai, N.; Bartoletti, T.M.; Akmentin, W.; Francis, A.; Matthews, G.; Thoreson, W.; Zenisek, D. Acute destruction of the synaptic ribbon reveals a role for the ribbon in vesicle priming. Nat. Neurosci. 2011, 14, 1135–1141. [Google Scholar] [CrossRef] [PubMed]
- Snellman, J.; Zenisek, D.; Nawy, S. Switching between transient and sustained signalling at the rod bipolar-AII amacrine cell synapse of the mouse retina. J. Physiol. 2009, 587 Pt 11, 2443–2455. [Google Scholar] [CrossRef]
- Singer, J.H.; Diamond, J.S. Sustained Ca2+ entry elicits transient postsynaptic currents at a retinal ribbon synapse. J. Neurosci. 2003, 23, 10923–10933. [Google Scholar] [CrossRef]
- Singer, J.H.; Diamond, J.S. Vesicle depletion and synaptic depression at a mammalian ribbon synapse. J. Neurophysiol. 2006, 95, 3191–3198. [Google Scholar] [CrossRef] [PubMed]
- Burrone, J.; Lagnado, L. Synaptic depression and the kinetics of exocytosis in retinal bipolar cells. J. Neurosci. 2000, 20, 568–578. [Google Scholar] [CrossRef] [PubMed]
- Burrone, J.; Neves, G.; Gomis, A.; Cooke, A.; Lagnado, L. Endogenous calcium buffers regulate fast exocytosis in the synaptic terminal of retinal bipolar cells. Neuron 2002, 33, 101–112. [Google Scholar] [CrossRef]
- von Gersdorff, H.; Matthews, G. Dynamics of synaptic vesicle fusion and membrane retrieval in synaptic terminals. Nature 1994, 367, 735–739. [Google Scholar] [CrossRef]
- von Gersdorff, H.; Matthews, G. Depletion and replenishment of vesicle pools at a ribbon-type synaptic terminal. J. Neurosci. 1997, 17, 1919–1927. [Google Scholar] [CrossRef]
- von Gersdorff, H.; Sakaba, T.; Berglund, K.; Tachibana, M. Submillisecond kinetics of glutamate release from a sensory synapse. Neuron 1998, 21, 1177–1188. [Google Scholar] [CrossRef]
- Mennerick, S.; Matthews, G. Ultrafast exocytosis elicited by calcium current in synaptic terminals of retinal bipolar neurons. Neuron 1996, 17, 1241–1249. [Google Scholar] [CrossRef]
- Matthews, G.; Fuchs, P. The diverse roles of ribbon synapses in sensory neurotransmission. Nat. Rev. Neurosci. 2010, 11, 812–822. [Google Scholar] [CrossRef]
- Shrestha, A.P.; Vaithianathan, T. Tracking the dynamics of single fused synaptic vesicle proteins from a single ribbon active zone in zebrafish retinal bipolar cells. STAR Protoc. 2022, 3, 101107–101128. [Google Scholar] [CrossRef] [PubMed]
- Vaithianathan, T.; Matthews, G. Visualizing synaptic vesicle turnover and pool refilling driven by calcium nanodomains at presynaptic active zones of ribbon synapses. Proc. Natl. Acad. Sci. USA 2014, 111, 8655–8660. [Google Scholar] [CrossRef] [PubMed]
- Vaithianathan, T.; Wollmuth, L.P.; Henry, D.; Zenisek, D.; Matthews, G. Tracking Newly Released Synaptic Vesicle Proteins at Ribbon Active Zones. iScience 2019, 17, 10–23. [Google Scholar] [CrossRef] [PubMed]
- Oesch, N.W.; Diamond, J.S. Ribbon synapses compute temporal contrast and encode luminance in retinal rod bipolar cells. Nat. Neurosci. 2011, 14, 1555–1561. [Google Scholar] [CrossRef]
- Peineau, T.; Belleudy, S.; Pietropaolo, S.; Bouleau, Y.; Dulon, D. Synaptic Release Potentiation at Aging Auditory Ribbon Synapses. Front. Aging Neurosci. 2021, 13, 756449–756469. [Google Scholar] [CrossRef] [PubMed]
- Mesnard, C.S.; Barta, C.L.; Sladek, A.L.; Zenisek, D.; Thoreson, W.B. Eliminating Synaptic Ribbons from Rods and Cones Halves the Releasable Vesicle Pool and Slows Down Replenishment. Int. J. Mol. Sci. 2022, 23, 6429–6452. [Google Scholar] [CrossRef] [PubMed]
- Maxeiner, S.; Luo, F.; Tan, A.; Schmitz, F.; Südhof, T.C. How to make a synaptic ribbon: RIBEYE deletion abolishes ribbons in retinal synapses and disrupts neurotransmitter release. Embo J. 2016, 35, 1098–1114. [Google Scholar] [CrossRef] [PubMed]
- Okawa, H.; Yu, W.Q.; Matti, U.; Schwarz, K.; Odermatt, B.; Zhong, H.; Tsukamoto, Y.; Lagnado, L.; Rieke, F.; Schmitz, F.; et al. Dynamic assembly of ribbon synapses and circuit maintenance in a vertebrate sensory system. Nat. Commun. 2019, 10, 2167–2180. [Google Scholar] [CrossRef] [PubMed]
- Zenisek, D.; Horst, N.K.; Merrifield, C.; Sterling, P.; Matthews, G. Visualizing synaptic ribbons in the living cell. J. Neurosci. 2004, 24, 9752–9759. [Google Scholar] [CrossRef]
- Connaughton, V.P.; Graham, D.; Nelson, R. Identification and morphological classification of horizontal, bipolar, and amacrine cells within the zebrafish retina. J. Comp. Neurol. 2004, 477, 371–385. [Google Scholar] [CrossRef] [PubMed]
- Negishi, K.; Kato, S.; Teranishi, T. Dopamine cells and rod bipolar cells contain protein kinase C-like immunoreactivity in some vertebrate retinas. Neurosci. Lett. 1988, 94, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, A.P.; Saravanakumar, A.; Konadu, B.; Madireddy, S.; Gibert, Y.; Vaithianathan, T. Embryonic Hyperglycemia Delays the Development of Retinal Synapses in a Zebrafish Model. Int. J. Mol. Sci. 2022, 23, 9693–9712. [Google Scholar] [CrossRef]
- Li, Y.N.; Tsujimura, T.; Kawamura, S.; Dowling, J.E. Bipolar cell-photoreceptor connectivity in the zebrafish (Danio rerio) retina. J. Comp. Neurol. 2012, 520, 3786–3802. [Google Scholar] [CrossRef] [PubMed]
- Haug, M.F.; Berger, M.; Gesemann, M.; Neuhauss, S.C.F. Differential expression of PKCα and -β in the zebrafish retina. Histochem. Cell Biol. 2019, 151, 521–530. [Google Scholar] [CrossRef]
- Heidelberger, R.; Matthews, G. Calcium influx and calcium current in single synaptic terminals of goldfish retinal bipolar neurons. J. Physiol. 1992, 447, 235–256. [Google Scholar] [CrossRef]
- Zenisek, D.; Davila, V.; Wan, L.; Almers, W. Imaging calcium entry sites and ribbon structures in two presynaptic cells. J. Neurosci. 2003, 23, 2538–2548. [Google Scholar] [CrossRef]
- Beaumont, V.; Llobet, A.; Lagnado, L. Expansion of calcium microdomains regulates fast exocytosis at a ribbon synapse. Proc. Natl. Acad. Sci. USA 2005, 102, 10700–10705. [Google Scholar] [CrossRef]
- Singer, J.H.; Lassová, L.; Vardi, N.; Diamond, J.S. Coordinated multivesicular release at a mammalian ribbon synapse. Nat. Neurosci. 2004, 7, 826–833. [Google Scholar] [CrossRef]
- Graydon, C.W.; Cho, S.; Li, G.L.; Kachar, B.; von Gersdorff, H. Sharp Ca2⁺ nanodomains beneath the ribbon promote highly synchronous multivesicular release at hair cell synapses. J. Neurosci. 2011, 31, 16637–16650. [Google Scholar] [CrossRef]
- Jarsky, T.; Tian, M.; Singer, J.H. Nanodomain control of exocytosis is responsible for the signaling capability of a retinal ribbon synapse. J. Neurosci. 2010, 30, 11885–11895. [Google Scholar] [CrossRef]
- Thoreson, W.B.; Mercer, A.J.; Cork, K.M.; Szalewski, R.J. Lateral mobility of L-type calcium channels in synaptic terminals of retinal bipolar cells. Mol. Vis. 2013, 19, 16–24. [Google Scholar] [PubMed]
- Fuchs, M.; Scholz, M.; Sendelbeck, A.; Atorf, J.; Schlegel, C.; Enz, R.; Brandstätter, J.H. Rod photoreceptor ribbon synapses in DBA/2J mice show progressive age-related structural changes. PLoS ONE 2012, 7, 44645–44655. [Google Scholar] [CrossRef] [PubMed]
- Nag, T.C.; Wadhwa, S. Ultrastructure of the human retina in aging and various pathological states. Micron 2012, 43, 759–781. [Google Scholar] [CrossRef] [PubMed]
- Terzibasi, E.; Calamusa, M.; Novelli, E.; Domenici, L.; Strettoi, E.; Cellerino, A. Age-dependent remodelling of retinal circuitry. Neurobiol. Aging 2009, 30, 819–828. [Google Scholar] [CrossRef]
- Haverkamp, S.; Wässle, H. Immunocytochemical analysis of the mouse retina. J. Comp. Neurol. 2000, 424, 1–23. [Google Scholar] [CrossRef]
- Caminos, E.; Velasco, A.; Jarrín, M.; Aijón, J.; Lara, J.M. Protein kinase C-like immunoreactive cells in embryo and adult chicken retinas. Brain Res. Dev. Brain Res. 1999, 118, 227–230. [Google Scholar] [CrossRef]
- Vaquero, C.F.; Velasco, A.; de la Villa, P. Protein kinase C localization in the synaptic terminal of rod bipolar cells. Neuroreport 1996, 7, 2176–2180. [Google Scholar] [CrossRef]
- Grünert, U.; Martin, P.R.; Wässle, H. Immunocytochemical analysis of bipolar cells in the macaque monkey retina. J. Comp. Neurol. 1994, 348, 607–627. [Google Scholar] [CrossRef]
- Zhang, D.R.; Yeh, H.H. Protein kinase C-like immunoreactivity in rod bipolar cells of the rat retina: A developmental study. Vis. Neurosci. 1991, 6, 429–437. [Google Scholar] [CrossRef]
- Osborne, N.N.; Broyden, N.J.; Barnett, N.L.; Morris, N.J. Protein kinase C (alpha and beta) immunoreactivity in rabbit and rat retina: Effect of phorbol esters and transmitter agonists on immunoreactivity and the translocation of the enzyme from cytosolic to membrane compartments. J. Neurochem. 1991, 57, 594–604. [Google Scholar] [CrossRef]
- Greferath, U.; Grünert, U.; Wässle, H. Rod bipolar cells in the mammalian retina show protein kinase C-like immunoreactivity. J. Comp. Neurol. 1990, 301, 433–442. [Google Scholar] [CrossRef] [PubMed]
- Mack, A.F. Evidence for a columnar organization of cones, Müller cells, and neurons in the retina of a cichlid fish. Neuroscience 2007, 144, 1004–1014. [Google Scholar] [CrossRef] [PubMed]
- Yazulla, S.; Studholme, K.M. Light-dependent plasticity of the synaptic terminals of Mb bipolar cells in goldfish retina. J. Comp. Neurol. 1992, 320, 521–530. [Google Scholar] [CrossRef] [PubMed]
- Chua, J.; Tham, Y.C.; Tan, B.; Devarajan, K.; Schwarzhans, F.; Gan, A.; Wong, D.; Cheung, C.Y.; Majithia, S.; Thakur, S.; et al. Age-related changes of individual macular retinal layers among Asians. Sci. Rep. 2019, 9, 20352–20363. [Google Scholar] [CrossRef]
- Liets, L.C.; Eliasieh, K.; van der List, D.A.; Chalupa, L.M. Dendrites of rod bipolar cells sprout in normal aging retina. Proc. Natl. Acad. Sci. USA 2006, 103, 12156–12160. [Google Scholar] [CrossRef]
- Dick, O.; tom Dieck, S.; Altrock, W.D.; Altrock, A.D.; Ammermüller, J.; Weiler, R.; Garner, C.; Gundelfinger, E.D.; Brandstätter, J.H. The presynaptic active zone protein bassoon is essential for photoreceptor ribbon synapse formation in the retina. Neuron 2003, 37, 775–786. [Google Scholar] [CrossRef]
- Midorikawa, M.; Tsukamoto, Y.; Berglund, K.; Ishii, M.; Tachibana, M. Different roles of ribbon-associated and ribbon-free active zones in retinal bipolar cells. Nat. Neurosci. 2007, 10, 1268–1276. [Google Scholar] [CrossRef]
- Sheets, L.; Trapani, J.G.; Mo, W.; Obholzer, N.; Nicolson, T. Ribeye is required for presynaptic Ca(V)1.3a channel localization and afferent innervation of sensory hair cells. Development 2011, 138, 1309–1319. [Google Scholar] [CrossRef]
- Jung, S.; Oshima-Takago, T.; Chakrabarti, R.; Wong, A.B.; Jing, Z.; Yamanbaeva, G.; Picher, M.M.; Wojcik, S.M.; Göttfert, F.; Predoehl, F.; et al. Rab3-interacting molecules 2α and 2β promote the abundance of voltage-gated CaV1.3 Ca2+ channels at hair cell active zones. Proc. Natl. Acad. Sci. USA 2015, 112, 3141–3149. [Google Scholar] [CrossRef]
- Grabner, C.P.; Moser, T. The mammalian rod synaptic ribbon is essential for Ca(v) channel facilitation and ultrafast synaptic vesicle fusion. Elife 2021, 10, e63844. [Google Scholar] [CrossRef] [PubMed]
- Frank, T.; Rutherford, M.A.; Strenzke, N.; Neef, A.; Pangršič, T.; Khimich, D.; Fejtova, A.; Gundelfinger, E.D.; Liberman, M.C.; Harke, B.; et al. Bassoon and the synaptic ribbon organize Ca2+ channels and vesicles to add release sites and promote refilling. Neuron 2010, 68, 724–738. [Google Scholar] [CrossRef]
- Wong, H.C.; Zhang, Q.; Beirl, A.J.; Petralia, R.S.; Wang, Y.X.; Kindt, K. Synaptic mitochondria regulate hair-cell synapse size and function. Elife 2019, 8, e48914. [Google Scholar] [CrossRef] [PubMed]
- Krinner, S.; Butola, T.; Jung, S.; Wichmann, C.; Moser, T. RIM-Binding Protein 2 Promotes a Large Number of Ca(V)1.3 Ca(2+)-Channels and Contributes to Fast Synaptic Vesicle Replenishment at Hair Cell Active Zones. Front. Cell Neurosci. 2017, 11, 334–354. [Google Scholar] [CrossRef] [PubMed]
- Heidelberger, R.; Thoreson, W.B.; Witkovsky, P. Synaptic transmission at retinal ribbon synapses. Prog. Retin. Eye Res. 2005, 24, 682–720. [Google Scholar] [CrossRef] [PubMed]
- Nachman-Clewner, M.; St Jules, R.; Townes-Anderson, E. L-type calcium channels in the photoreceptor ribbon synapse: Localization and role in plasticity. J. Comp. Neurol. 1999, 415, 1–16. [Google Scholar] [CrossRef]
- Zanazzi, G.; Matthews, G. The molecular architecture of ribbon presynaptic terminals. Mol. Neurobiol. 2009, 39, 130–148. [Google Scholar] [CrossRef] [PubMed]
- Dembla, E.; Dembla, M.; Maxeiner, S.; Schmitz, F. Synaptic ribbons foster active zone stability and illumination-dependent active zone enrichment of RIM2 and Cav1.4 in photoreceptor synapses. Sci. Rep. 2020, 10, 5957–5977. [Google Scholar] [CrossRef]
- Naraghi, M.; Neher, E. Linearized buffered Ca2+ diffusion in microdomains and its implications for calculation of [Ca2+] at the mouth of a calcium channel. J. Neurosci. 1997, 17, 6961–6973. [Google Scholar] [CrossRef]
- Moser, T.; Predoehl, F.; Starr, A. Review of hair cell synapse defects in sensorineural hearing impairment. Otol. Neurotol. 2013, 34, 995–1004. [Google Scholar] [CrossRef]
- Xiong, W.; Yu, S.; Liu, K.; Gong, S. Loss of cochlear ribbon synapses in the early stage of aging causes initial hearing impairment. Am. J. Transl. Res. 2020, 12, 7354–7366. [Google Scholar]
- Mehta, B.; Ke, J.B.; Zhang, L.; Baden, A.D.; Markowitz, A.L.; Nayak, S.; Briggman, K.L.; Zenisek, D.; Singer, J.H. Global Ca2+ signaling drives ribbon-independent synaptic transmission at rod bipolar cell synapses. J. Neurosci. 2014, 34, 6233–6244. [Google Scholar] [CrossRef] [PubMed]
- Joselevitch, C.; Zenisek, D. Direct Observation of Vesicle Transport on the Synaptic Ribbon Provides Evidence That Vesicles Are Mobilized and Prepared Rapidly for Release. J. Neurosci. 2020, 40, 7390–7404. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Van Hook, M.J.; Zenisek, D.; Thoreson, W.B. Properties of ribbon and non-ribbon release from rod photoreceptors revealed by visualizing individual synaptic vesicles. J. Neurosci. 2013, 33, 2071–2086. [Google Scholar] [CrossRef] [PubMed]
- Luo, F.; Bacaj, T.; Südhof, T.C. Synaptotagmin-7 Is Essential for Ca2+-Triggered Delayed Asynchronous Release But Not for Ca2+-Dependent Vesicle Priming in Retinal Ribbon Synapses. J. Neurosci. 2015, 35, 11024–11033. [Google Scholar] [CrossRef]
- Courtney, N.A.; Bao, H.; Briguglio, J.S.; Chapman, E.R. Synaptotagmin 1 clamps synaptic vesicle fusion in mammalian neurons independent of complexin. Nat. Commun. 2019, 10, 4076–4090. [Google Scholar] [CrossRef]
- Kishi, S. Functional aging and gradual senescence in zebrafish. Ann. N. Y. Acad. Sci. 2004, 1019, 521–526. [Google Scholar] [CrossRef]
- Hull, C.; Studholme, K.; Yazulla, S.; von Gersdorff, H. Diurnal changes in exocytosis and the number of synaptic ribbons at active zones of an ON-type bipolar cell terminal. J. Neurophysiol. 2006, 96, 2025–2033. [Google Scholar] [CrossRef]
- Živković, M.; Dayanir, V.; Stamenović, J.; Ljubisavljević, S.; Pražić, A.; Zlatanović, M.; Zlatanović, G.; Jakšić, V.; Radenković, M.; Jovanović, S. Retinal ganglion cell/inner plexiform layer thickness in patients with Parkinson’s disease. Folia Neuropathol. 2017, 55, 168–173. [Google Scholar] [CrossRef]
- Kim, M.H.; von Gersdorff, H. Postsynaptic Plasticity Triggered by Ca2⁺-Permeable AMPA Receptor Activation in Retinal Amacrine Cells. Neuron 2016, 89, 507–520. [Google Scholar] [CrossRef]
- Sebe, J.Y.; Cho, S.; Sheets, L.; Rutherford, M.A.; von Gersdorff, H.; Raible, D.W. Ca(2+)-Permeable AMPARs Mediate Glutamatergic Transmission and Excitotoxic Damage at the Hair Cell Ribbon Synapse. J. Neurosci. 2017, 37, 6162–6175. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Shi, Y.; Wei, H. Calcium Dysregulation in Alzheimer’s Disease: A Target for New Drug Development. J. Alzheimers. Dis. Park. 2017, 7, 374. [Google Scholar] [CrossRef] [PubMed]
- Kiani, L. Calcium dysregulation could underlie lysosomal impairment in AD. Nat. Rev. Neurol. 2023, 19, 65. [Google Scholar] [CrossRef] [PubMed]
- Small, D.H. Dysregulation of calcium homeostasis in Alzheimer’s disease. Neurochem. Res. 2009, 34, 1824–1829. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antigen | Antiserum | Host | Dilution | Source (Number) | Marker for |
|---|---|---|---|---|---|
| PKC | Monoclonal anti-PKC | Mouse | 1:250 | Santa Cruz (sc-17769) | Rod-type bipolar cells |
| Ribeye-A | Polyclonal anti-ribeye | Rabbit | 1:1000 | Zenisek lab (s4561-2) | Synaptic ribbons (IPL) |
| Antibody | Conjugation | Source (Number) |
|---|---|---|
| Donkey anti-guinea pig | Cy3 | Jackson Immunoresearch, #706-165-148 |
| Donkey anti-mouse | Alexa Fluor 647 | Southern Biotech, #6440-31 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shrestha, A.P.; Rameshkumar, N.; Boff, J.M.; Rajmanna, R.; Chandrasegaran, T.; Frederick, C.E.; Zenisek, D.; Vaithianathan, T. The Effects of Aging on Rod Bipolar Cell Ribbon Synapses. Cells 2023, 12, 2385. https://doi.org/10.3390/cells12192385
Shrestha AP, Rameshkumar N, Boff JM, Rajmanna R, Chandrasegaran T, Frederick CE, Zenisek D, Vaithianathan T. The Effects of Aging on Rod Bipolar Cell Ribbon Synapses. Cells. 2023; 12(19):2385. https://doi.org/10.3390/cells12192385
Chicago/Turabian StyleShrestha, Abhishek P., Nirujan Rameshkumar, Johane M. Boff, Rhea Rajmanna, Thadshayini Chandrasegaran, Courtney E. Frederick, David Zenisek, and Thirumalini Vaithianathan. 2023. "The Effects of Aging on Rod Bipolar Cell Ribbon Synapses" Cells 12, no. 19: 2385. https://doi.org/10.3390/cells12192385