
Microglia and Other Cellular Mediators of Immunological Dysfunction in Schizophrenia: A Narrative Synthesis of Clinical Findings

 ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Cellular Constituents of CNS Immunological Aberrations in SCZ
2.1. Microglia
2.2. Other Immune Cell Types
3. Alterations in Circulating Immune Cells in SCZ
3.1. Monocytes
3.2. Granulocytes
3.3. Natural Killer Cells
3.4. B Lymphocytes
3.5. T Lymphocytes

{kind=link}
{kind=link}
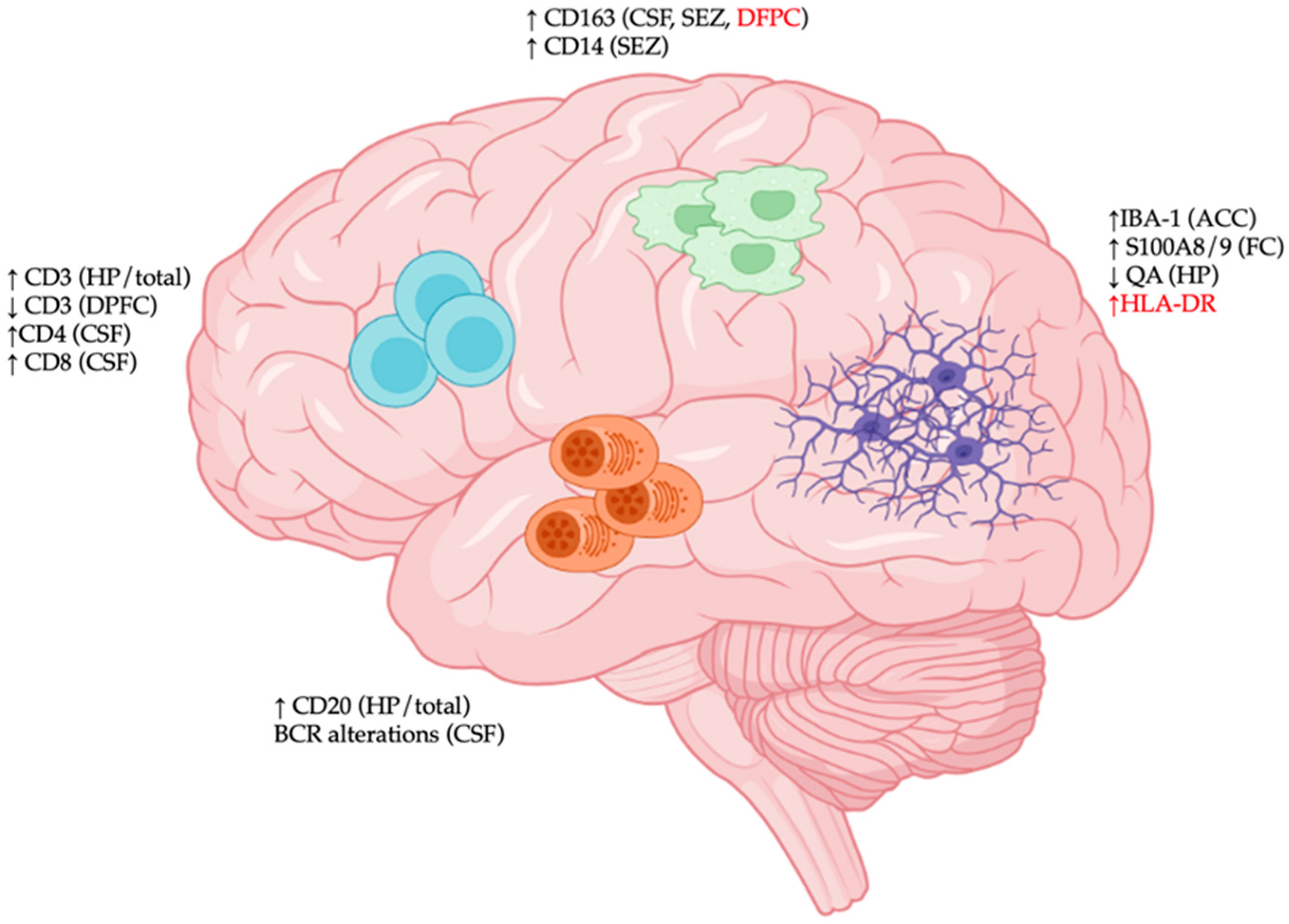
| Central Nervous System | Features | Anatomical Location |
|---|---|---|
| Microglia | Increased phagocytosis [13] | Whole brain |
| No change in phagocytosis [31] | PFC | |
| Increased IBA-1 [30] | ACC | |
| No change in IBA-1 [31] | PFC | |
| Increased HLA-DR [19,20] | Frontal/temporal cortex, hippocampus | |
| No change in HLA-DR [25,26] | PFC, ACC, hippocampus | |
| Increased S100A8/9 [28] | Frontal cortex | |
| Activated morphology [14,15,16] | PFC, visual cortex | |
| Microgliosis [32] | Frontal/temporal cortex | |
| Decreased quinolinic acid [29] | Hippocampus | |
| T cells | Increased CD3+ frequency [19,48] | Whole brain, hippocampus |
| Decreased CD3+ frequency [19] | Dorsal PFC | |
| Increased CD4+ frequency [55] | CSF | |
| Increased CD8+ frequency [55] | CSF | |
| B cells | Increased CD20+ frequency [19,52] | Whole brain, hippocampus |
| BCR alterations [60] | CSF | |
| Macrophages | Increased CD163 [53,55,56] | Dorsal PFC, CSF, SEZ |
| No change in CD163 [33] | Dorsal PFC | |
| Increased CD14 [53] | SEZ | |
| Increased frequency [57] | CSF | |
| Peripheral Blood | Features | |
| Monocytes | Increased total monocyte counts [62,63,64] | |
| Increased classical monocyte counts [66] | ||
| Increased pro-inflammatory monocyte counts [67] | ||
| Increased monocyte to lymphocyte ratio [68] | ||
| Increased monocyte to HDL ratio [72,73] | ||
| Increased TREM1/2 [79,80] | ||
| Increased ATF3/EGR3 [78] | ||
| Increased HLA-DR [82] | ||
| Increased phagocytosis [81] | ||
| Increased IL-1, IL-6, TNF-α [83,85,86] | ||
| Increased CD36 [93] | ||
| Increased reactive oxygen species [92] | ||
| Alterations in TLR4 signaling [84,87,88] | ||
| Unique interferon gene signature [77] | ||
| Reduced GLUT1 [89] | ||
| Granulocytes | Increased neutrophil counts [68,94,95,96,97,98,99] | |
| Increased neutrophil to lymphocyte ratio [100,101] | ||
| Increased oxidative stress [108] | ||
| Increased malonaldehyde [122] | ||
| Increased superoxide anion [123,123] | ||
| Increased phagocytosis [81,124] | ||
| Natural killer cells | Increased total counts [68] | |
| Decreased total counts [125,126,127] | ||
| No change in total counts [128] | ||
| Increased cytotoxicity [132] | ||
| Decreased cytotoxicity [129,130] | ||
| No change in cytotoxicity [121] | ||
| Increased NKG2C [133] | ||
| Increased S100B [134] | ||
| T Cells | Increased CD3+ and CD4+ T cell counts [153] | |
| Reduced CD3+ T cell counts [150] | ||
| Reduced CD4+ T cell counts [146,152] | ||
| Increased activation [63,154,155] | ||
| Increased CD25 [156] | ||
| Reduced IL2 production [156,157,158] | ||
| No change in IL2 production [92] | ||
| Increased Treg [63,155,159,160,161,162] | ||
| Increased Th17 [63,154,155,156,157,158,159,160,161,162] | ||
| Increased mucosal-associated invariant T cells [162] | ||
| Increased oxidative stress [166] | ||
| Altered methylation profile [167] | ||
| Altered TCR repertoire [167] | ||
| B Cells | Increased CD19+ B cell frequency [156] | |
| Increased IgD+CD27-CD19+ naïve B cell frequency [67] | ||
| Increased CD5+ B cell frequency [157,158] | ||
| No change in CD5+ B cell frequency [159] | ||
| Increased GAD, GABAR1, A7ChR, NMDAR autoantibodies [136,137,138,139] | ||
| No changes in GAD, GABAR1, A7ChR, NMDAR autoantibodies [140,141,142] | ||
| Increased DNA/RNA autoantibodies [143,144] | ||
| Increased heat-shock protein autoantibodies [145,146] | ||
| Increased neuronal marker autoantibodies [147,148,149,150] | ||
| Increased IL-1/IL-6/IL-8/gliadin autoantibodies [151,152,153,154,155] | ||
4. Implications for Mechanistic Studies and Therapeutic Development
5. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
References
- Faden, J.; Citrome, L. Schizophrenia: One name, many different manifestations. Med. Clin. N. Am. 2023, 107, 61–72. [Google Scholar] [CrossRef]
- Costanza, A.; Baertschi, M.; Weber, K.; Canuto, A. Maladies neurologiques et suicide: De la neurobiology au manque d’éspoir [neurological diseases and suicide: From neurobiology to hopelessness]. Rev. Med. Suisse 2015, 11, 402–405. [Google Scholar] [PubMed]
- Lewine, R.; Hart, M. Schizophrenia spectrum and other psychotic disorders. In Handbook of Clinical Neurology; Elsevier: Amsterdam, The Netherlands, 2020; pp. 315–333. [Google Scholar]
- Janoutová, J.; Ambroz, P.; Kovalová, M.; Machaczka, O.; Němeček, K.; Zatloukalová, A.; Mrázková, E.; Košta, O.; Hálová, A.; Hosák, L.; et al. Epidemiology of mild cognitive impairment. Česká A Slov. Neurol. A Neurochir. 2018, 81, 284–289. [Google Scholar] [CrossRef]
- Sabe, M.; Chen, C.; Perez, N.; Solmi, M.; Mucci, A.; Galderisi, S.; Strauss, G.P.; Kaiser, S. Thirty years of research on negative symptoms of schizophrenia: A scientometric analysis of hotspots, bursts, and research trends. Neurosci. Biobehav. Rev. 2023, 144, 104979. [Google Scholar] [CrossRef] [PubMed]
- Bakhshi, K.; Chance, S.A. The neuropathology of schizophrenia: A selective review of past studies and emerging themes in brain structure and cytoarchitecture. Neuroscience 2015, 303, 82–102. [Google Scholar] [CrossRef] [PubMed]
- Costanza, A.; Amerio, A.; Aguglia, A.; Serafini, G.; Amore, M.; Hasler, R.; Ambrosetti, J.; Bondolfi, G.; Sampogna, G.; Berardelli, I.; et al. Hyper/neuroinflammation in COVID-19 and suicide etiopathogenesis: Hypothesis for a nefarious collision? Neurosci. Biobehav. Rev. 2022, 136, 104606. [Google Scholar] [CrossRef]
- Serafini, G.; Costanza, A.; Aguglia, A.; Amerio, A.; Trabucco, A.; Escelsior, A.; Sher, L.; Amore, M. The role of inflammation in the pathophysiology of depression and suicidal behavior: Implications for treatment. Med. Clin. N. Am. 2023, 107, 1–29. [Google Scholar] [CrossRef]
- Fišar, Z. Biological hypotheses, risk factors, and biomarkers of schizophrenia. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2023, 120, 110626. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, Y.; Yuan, X.; Kang, Y.; Song, X. New insight in the cross-talk between microglia and schizophrenia: From the perspective of neurodevelopment. Front. Psychiatr. 2023, 14, 1126632. [Google Scholar] [CrossRef]
- Cao, Y.; Ji, S.; Chen, Y.; Zhang, X.; Ding, G.; Tang, F. Association between autoimmune diseases of the nervous system and schizophrenia: A systematic review and meta-analysis of cohort studies. Compr. Psychiatry 2023, 122, 152370. [Google Scholar] [CrossRef]
- Kierdorf, K.; Masuda, T.; Jordão, M.J.C.; Prinz, M. Macrophages at cns interfaces: Ontogeny and function in health and disease. Nat. Rev. Neurosci. 2019, 20, 547–562. [Google Scholar] [CrossRef] [PubMed]
- Solov’eva, Z.; Orlovskaia, D.D. Microglia-type cells in normal and pathologic human embryonic brains. Zhurnal Nevropatol. I Psikhiatrii Im. SS Korsakova 1979, 79, 852–857. [Google Scholar]
- Uranova, N.A.; Vikhreva, O.V.; Rakhmanova, V.I.; Orlovskaya, D.D. Ultrastructural pathology of oligodendrocytes in the white matter in continuous paranoid schizophrenia: A role for microglia. Zhurnal Nevropatol. I Psikhiatrii Im. SS Korsakova 2017, 117, 76. [Google Scholar] [CrossRef] [PubMed]
- Uranova, N.A.; Zimina, I.S.; Vikhreva, O.V.; Krukov, N.O.; Rachmanova, V.I.; Orlovskaya, D.D. Ultrastructural damage of capillaries in the neocortex in schizophrenia. World J. Biol. Psychiatry 2010, 11, 567–578. [Google Scholar] [CrossRef]
- Vikhreva, O.V.; Rakhmanova, V.I.; Orlovskaya, D.D.; Uranova, N.A. Ultrastructural pathology of oligodendrocytes in white matter in continuous attack-like schizophrenia and a role for microglia. Zhurnal Nevropatol. I Psikhiatrii Im. SS Korsakova 2018, 118, 69. [Google Scholar] [CrossRef]
- Uranova, N.A.; Vikhreva, O.V.; Rakhmanova, V.I.; Orlovskaya, D.D. Ultrastructural pathology of oligodendrocytes adjacent to microglia in prefrontal white matter in schizophrenia. NPJ Schizophr. 2018, 4, 26. [Google Scholar] [CrossRef] [PubMed]
- Uranova, N.A.; Vikhreva, O.V.; Rakhmanova, V.I.; Orlovskaya, D.D. Dystrophy of oligodendrocytes and adjacent microglia in prefrontal gray matter in schizophrenia. Front. Psychiatr. 2020, 11, 204. [Google Scholar] [CrossRef] [PubMed]
- Busse, S.; Busse, M.; Schiltz, K.; Bielau, H.; Gos, T.; Brisch, R.; Mawrin, C.; Schmitt, A.; Jordan, W.; Müller, U.J.; et al. Different distribution patterns of lymphocytes and microglia in the hippocampus of patients with residual versus paranoid schizophrenia: Further evidence for disease course-related immune alterations? Brain Behav. Immun. 2012, 26, 1273–1279. [Google Scholar] [CrossRef] [PubMed]
- Radewicz, K.; Garey, L.J.; Gentleman, S.M.; Reynolds, R. Increase in hla-dr immunoreactive microglia in frontal and temporal cortex of chronic schizophrenics. J. Neuropathol. Exp. Neurol. 2000, 59, 137–150. [Google Scholar] [CrossRef] [PubMed]
- Wierzba-Bobrowicz, T.; Lewandowska, E.; Kosno-Kruszewska, E.; Lechowicz, W.; Pasennik, E.; Schmidt-Sidor, B. Degeneration of microglial cells in frontal and temporal lobes of chronic schizophrenics. Folia Neuropathol. 2004, 42, 157–165. [Google Scholar] [PubMed]
- Fillman, S.G.; Cloonan, N.; Catts, V.S.; Miller, L.C.; Wong, J.; McCrossin, T.; Cairns, M.; Weickert, C.S. Increased inflammatory markers identified in the dorsolateral prefrontal cortex of individuals with schizophrenia. Mol. Psychiatry 2012, 18, 206–214. [Google Scholar] [CrossRef]
- Bayer, T.A.; Buslei, R.; Havas, L.; Falkai, P. Evidence for activation of microglia in patients with psychiatric illnesses. Neurosci. Lett. 1999, 271, 126–128. [Google Scholar] [CrossRef]
- Costanza, A.; Xekardaki, A.; Kovari, E.; Gold, G.; Bouras, C.; Giannakopoulos, P. Microvascular burden and alzheimer-type lesions across the age spectrum. J. Alzheimers Dis. 2012, 32, 643–652. [Google Scholar] [CrossRef]
- Steiner, J.; Bielau, H.; Brisch, R.; Danos, P.; Ullrich, O.; Mawrin, C.; Bernstein, H.-G.; Bogerts, B. Immunological aspects in the neurobiology of suicide: Elevated microglial density in schizophrenia and depression is associated with suicide. J. Psychiatr. Res. 2008, 42, 151–157. [Google Scholar] [CrossRef]
- Steiner, J.; Mawrin, C.; Ziegeler, A.; Bielau, H.; Ullrich, O.; Bernstein, H.-G.; Bogerts, B. Distribution of hla-dr-positive microglia in schizophrenia reflects impaired cerebral lateralization. Acta Neuropathol. 2006, 112, 305–316. [Google Scholar] [CrossRef] [PubMed]
- Foster, R.; Kandanearatchi, A.; Beasley, C.; Williams, B.; Khan, N.; Fagerhol, M.K.; Everall, I.P. Calprotectin in microglia from frontal cortex is up-regulated in schizophrenia: Evidence for an inflammatory process? Eur. J. Neurosci. 2006, 24, 3561–3566. [Google Scholar] [CrossRef] [PubMed]
- Gos, T.; Myint, A.-M.; Schiltz, K.; Meyer-Lotz, G.; Dobrowolny, H.; Busse, S.; Müller, U.J.; Mawrin, C.; Bernstein, H.-G.; Bogerts, B.; et al. Reduced microglial immunoreactivity for endogenous nmda receptor agonist quinolinic acid in the hippocampus of schizophrenia patients. Brain Behav. Immun. 2014, 41, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Mildner, A.; Huang, H.; Radke, J.; Stenzel, W.; Priller, J. P2y(12) receptor is expressed on human microglia under physiological conditions throughout development and is sensitive to neuroinflammatory diseases. Glia 2017, 65, 375–387. [Google Scholar] [CrossRef] [PubMed]
- Petrasch-Parwez, E.; Schöbel, A.; Benali, A.; Moinfar, Z.; Förster, E.; Brüne, M.; Juckel, G. Lateralization of increased density of iba1-immunopositive microglial cells in the anterior midcingulate cortex of schizophrenia and bipolar disorder. Eur. Arch. Psychiatry Clin. Neurosci. 2020, 270, 819–828. [Google Scholar] [CrossRef] [PubMed]
- Tzioras, M.; Stevenson, A.J.; Boche, D.; Spires-Jones, T.L. Microglial contribution to synaptic uptake in the prefrontal cortex in schizophrenia. Neuropathol. Appl. Neurobiol. 2020, 47, 346–351. [Google Scholar] [CrossRef]
- Gober, R.; Ardalan, M.; Shiadeh, S.M.J.; Duque, L.; Garamszegi, S.P.; Ascona, M.; Barreda, A.; Sun, X.; Mallard, C.; Vontell, R.T. Microglia activation in postmortem brains with schizophrenia demonstrates distinct morphological changes between brain regions. Brain Pathol. 2022, 32, e13003. [Google Scholar] [CrossRef] [PubMed]
- De Picker, L.J.; Victoriano, G.M.; Richards, R.; Gorvett, A.J.; Lyons, S.; Buckland, G.R.; Tofani, T.; Norman, J.L.; Chatelet, D.S.; Nicoll, J.A.R.; et al. Immune environment of the brain in schizophrenia and during the psychotic episode: A human post-mortem study. Brain Behav. Immun. 2021, 97, 319–327. [Google Scholar] [CrossRef]
- Uranova, N.A.; Vikhreva, O.V.; Rakhmanova, V.I. Abnormal microglial reactivity in gray matter of the prefrontal cortex in schizophrenia. Asian J. Psychiatry 2021, 63, 102752. [Google Scholar] [CrossRef] [PubMed]
- Mokrov, G.V.; Deeva, O.A.; Gudasheva, T.A. The ligands of translocator protein: Design and biological properties. Curr. Pharm. Des. 2021, 27, 217–237. [Google Scholar] [CrossRef]
- Papadopoulos, V.; Baraldi, M.; Guilarte, T.R.; Knudsen, T.B.; Lacapère, J.J.; Lindemann, P.; Norenberg, M.D.; Nutt, D.; Weizman, A.; Zhang, M.R.; et al. Translocator protein (18kda): New nomenclature for the peripheral-type benzodiazepine receptor based on its structure and molecular function. Trends Pharmacol. Sci. 2006, 27, 402–409. [Google Scholar] [CrossRef]
- Wolf, A.; Herb, M.; Schramm, M.; Langmann, T. The tspo-nox1 axis controls phagocyte-triggered pathological angiogenesis in the eye. Nat. Commun. 2020, 11, 2709. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.; Han, R.; Chen, X.; Liu, X.; Wan, J.; Wang, L.; Yang, X.; Wang, J. Potential therapeutic targets for intracerebral hemorrhage-associated inflammation: An update. J. Cereb. Blood Flow Metab. Off. J. Int. Soc. Cereb. Blood Flow Metab. 2020, 40, 1752–1768. [Google Scholar] [CrossRef] [PubMed]
- Collste, K.; Plavén-Sigray, P.; Fatouros-Bergman, H.; Victorsson, P.; Schain, M.; Forsberg, A.; Amini, N.; Aeinehband, S.; Erhardt, S.; Halldin, C.; et al. Lower levels of the glial cell marker tspo in drug-naive first-episode psychosis patients as measured using pet and [11c]pbr28. Mol. Psychiatry 2017, 22, 850–856. [Google Scholar] [CrossRef] [PubMed]
- Conen, S.; Gregory, C.J.; Hinz, R.; Smallman, R.; Corsi-Zuelli, F.; Deakin, B.; Talbot, P.S. Neuroinflammation as measured by positron emission tomography in patients with recent onset and established schizophrenia: Implications for immune pathogenesis. Mol. Psychiatry 2021, 26, 5398–5406. [Google Scholar] [CrossRef]
- Hafizi, S.; Tseng, H.-H.; Rao, N.; Selvanathan, T.; Kenk, M.; Bazinet, R.P.; Suridjan, I.; Wilson, A.A.; Meyer, J.H.; Remington, G.; et al. Imaging microglial activation in untreated first-episode psychosis: A pet study with [(18)f]feppa. Am. J. Psychiatry 2017, 174, 118–124. [Google Scholar] [CrossRef]
- Holmes, S.E.; Hinz, R.; Drake, R.J.; Gregory, C.J.; Conen, S.; Matthews, J.C.; Anton-Rodriguez, J.M.; Gerhard, A.; Talbot, P.S. In vivo imaging of brain microglial activity in antipsychotic-free and medicated schizophrenia: A [11c](r)-pk11195 positron emission tomography study. Mol. Psychiatry 2016, 21, 1672–1679. [Google Scholar] [CrossRef] [PubMed]
- Ottoy, J.; De Picker, L.; Verhaeghe, J.; Deleye, S.; Wyffels, L.; Kosten, L.; Sabbe, B.; Coppens, V.; Timmers, M.; van Nueten, L.; et al. (18)f-pbr111 pet imaging in healthy controls and schizophrenia: Test-retest reproducibility and quantification of neuroinflammation. J. Nucl. Med. Off. Publ. Soc. Nucl. Med. 2018, 59, 1267–1274. [Google Scholar]
- Selvaraj, S.; Bloomfield, P.S.; Cao, B.; Veronese, M.; Turkheimer, F.; Howes, O.D. Brain tspo imaging and gray matter volume in schizophrenia patients and in people at ultra high risk of psychosis: An [(11)c]pbr28 study. Schizophr. Res. 2018, 195, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Van der Doef, T.F.; de Witte, L.D.; Sutterland, A.L.; Jobse, E.; Yaqub, M.; Boellaard, R.; de Haan, L.; Eriksson, J.; Lammertsma, A.A.; Kahn, R.S.; et al. In vivo (r)-[(11)c]pk11195 pet imaging of 18kda translocator protein in recent onset psychosis. NPJ Schizophr. 2016, 2, 16031. [Google Scholar] [CrossRef]
- Marques, T.R.; Ashok, A.H.; Pillinger, T.; Veronese, M.; Turkheimer, F.E.; Dazzan, P.; Sommer, I.E.C.; Howes, O.D. Neuroinflammation in schizophrenia: Meta-analysis of in vivo microglial imaging studies. Psychol. Med. 2019, 49, 2186–2196. [Google Scholar] [CrossRef]
- Plavén-Sigray, P.; Matheson, G.J.; Collste, K.; Ashok, A.H.; Coughlin, J.M.; Howes, O.D.; Mizrahi, R.; Pomper, M.G.; Rusjan, P.; Veronese, M.; et al. Positron emission tomography studies of the glial cell marker translocator protein in patients with psychosis: A meta-analysis using individual participant data. Biol. Psychiatry 2018, 84, 433–442. [Google Scholar] [CrossRef]
- Notter, T.; Coughlin, J.M.; Gschwind, T.; Weber-Stadlbauer, U.; Wang, Y.; Kassiou, M.; Vernon, A.C.; Benke, D.; Pomper, M.G.; Sawa, A.; et al. Translational evaluation of translocator protein as a marker of neuroinflammation in schizophrenia. Mol. Psychiatry 2017, 23, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Corley, E.; Holleran, L.; Fahey, L.; Corvin, A.; Morris, D.W.; Donohoe, G. Microglial-expressed genetic risk variants, cognitive function and brain volume in patients with schizophrenia and healthy controls. Transl. Psychiatry 2021, 11, 490. [Google Scholar] [CrossRef]
- Mou, T.-C.M.; Lane, M.V.; Ireland, D.D.C.; Verthelyi, D.; Tonelli, L.H.; Clark, S.M. Association of complement component 4 with neuroimmune abnormalities in the subventricular zone in schizophrenia and autism spectrum disorders. Neurobiol. Dis. 2022, 173, 105840. [Google Scholar] [CrossRef]
- Snijders, G.J.L.J.; van Zuiden, W.; Sneeboer, M.A.M.; Berdenis van Berlekom, A.; van der Geest, A.T.; Schnieder, T.; MacIntyre, D.J.; Hol, E.M.; Kahn, R.S.; de Witte, L.D. A loss of mature microglial markers without immune activation in schizophrenia. Glia 2021, 69, 1251–1267. [Google Scholar] [CrossRef]
- Schlaaff, K.; Dobrowolny, H.; Frodl, T.; Mawrin, C.; Gos, T.; Steiner, J.; Bogerts, B. Increased densities of t and b lymphocytes indicate neuroinflammation in subgroups of schizophrenia and mood disorder patients. Brain Behav. Immun. 2020, 88, 497–506. [Google Scholar] [CrossRef] [PubMed]
- North, H.F.; Weissleder, C.; Fullerton, J.M.; Webster, M.J.; Weickert, C.S. Increased immune cell and altered microglia and neurogenesis transcripts in an australian schizophrenia subgroup with elevated inflammation. Schizophr. Res. 2022, 248, 208–218. [Google Scholar] [CrossRef] [PubMed]
- North, H.F.; Weissleder, C.; Fullerton, J.M.; Sager, R.; Webster, M.J.; Weickert, C.S. A schizophrenia subgroup with elevated inflammation displays reduced microglia, increased peripheral immune cell and altered neurogenesis marker gene expression in the subependymal zone. Transl. Psychiatry 2021, 11, 635. [Google Scholar] [CrossRef] [PubMed]
- Purves-Tyson, T.D.; Robinson, K.; Brown, A.M.; Boerrigter, D.; Cai, H.Q.; Weissleder, C.; Owens, S.J.; Rothmond, D.A.; Shannon Weickert, C. Increased macrophages and c1qa, c3, c4 transcripts in the midbrain of people with schizophrenia. Front. Immunol. 2020, 11, 2002. [Google Scholar] [CrossRef]
- Zhu, Y.; Webster, M.J.; Murphy, C.E.; Middleton, F.A.; Massa, P.T.; Liu, C.; Dai, R.; Weickert, C.S. Distinct phenotypes of inflammation associated macrophages and microglia in the prefrontal cortex schizophrenia compared to controls. Front. Neurosci. 2022, 16, 858989. [Google Scholar] [CrossRef]
- Nikkilä, H.V.; Müller, K.; Ahokas, A.; Miettinen, K.; Rimón, R.; Andersson, L.C. Accumulation of macrophages in the csf of schizophrenic patients during acute psychotic episodes. Am. J. Psychiatry 1999, 156, 1725–1729. [Google Scholar] [CrossRef]
- Nikkilä, H.V.; Müller, K.; Ahokas, A.; Rimón, R.; Andersson, L.C. Increased frequency of activated lymphocytes in the cerebrospinal fluid of patients with acute schizophrenia. Schizophr. Res. 2001, 49, 99–105. [Google Scholar] [CrossRef]
- Nikkilä, H.; Müller, K.; Ahokas, A.; Miettinen, K.; Andersson, L.C.; Rimón, R. Abnormal distributions of t-lymphocyte subsets in the cerebrospinal fluid of patients with acute schizophrenia. Schizophr. Res. 1995, 14, 215–221. [Google Scholar] [CrossRef]
- Husain-Krautter, S.; Lee, J.; Vos, D.; Gallego, J.A.; Malhotra, A.K.; Rothstein, T.L. Skewing of the antibody repertoire in cerebrospinal fluid b cells from healthy controls and patients with schizophrenia. Behav. Brain Res. 2022, 422, 113743. [Google Scholar] [CrossRef]
- Ermakov, E.A.; Melamud, M.M.; Buneva, V.N.; Ivanova, S.A. Immune system abnormalities in schizophrenia: An integrative view and translational perspectives. Front. Psychiatr. 2022, 13, 880568. [Google Scholar] [CrossRef]
- Mazza, M.G.; Capellazzi, M.; Lucchi, S.; Tagliabue, I.; Rossetti, A.; Clerici, M. Monocyte count in schizophrenia and related disorders: A systematic review and meta-analysis. Acta Neuropsychiatr. 2020, 32, 229–236. [Google Scholar] [CrossRef]
- Orhan, F.; Schwieler, L.; Fatouros-Bergman, H.; Malmqvist, A.; Cervenka, S.; Collste, K.; Flyckt, L.; Farde, L.; Sellgren, C.M.; Piehl, F.; et al. Increased number of monocytes and plasma levels of mcp-1 and ykl-40 in first-episode psychosis. Acta Psychiatr. Scand. 2018, 138, 432–440. [Google Scholar] [CrossRef] [PubMed]
- Steiner, J.; Frodl, T.; Schiltz, K.; Dobrowolny, H.; Jacobs, R.; Fernandes, B.S.; Guest, P.C.; Meyer-Lotz, G.; Borucki, K.; Bahn, S.; et al. Innate immune cells and c-reactive protein in acute first-episode psychosis and schizophrenia: Relationship to psychopathology and treatment. Schizophr. Bull. 2020, 46, 363–373. [Google Scholar] [CrossRef] [PubMed]
- Miller, B.J.; Kandhal, P.; Rapaport, M.H.; Mellor, A.; Buckley, P. Total and differential white blood cell counts, high-sensitivity c-reactive protein, and cardiovascular risk in non-affective psychoses. Brain Behav. Immun. 2015, 45, 28–35. [Google Scholar] [CrossRef]
- Drexhage, R.C.; Hoogenboezem, T.A.; Cohen, D.; Versnel, M.A.; Nolen, W.A.; van Beveren, N.J.M.; Drexhage, H.A. An activated set point of t-cell and monocyte inflammatory networks in recent-onset schizophrenia patients involves both pro- and anti-inflammatory forces. Int. J. Neuropsychopharmacol. 2011, 14, 746–755. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Egea, E.; Vértes, P.E.; Flint, S.M.; Turner, L.; Mustafa, S.; Hatton, A.; Smith, K.G.C.; Lyons, P.A.; Bullmore, E.T. Peripheral immune cell populations associated with cognitive deficits and negative symptoms of treatment-resistant schizophrenia. PLoS ONE 2016, 11, e0155631. [Google Scholar] [CrossRef]
- Xu, H.; Wei, Y.; Zheng, L.; Zhang, H.; Luo, T.; Li, H.; Ma, J.; Chen, J. Relation between unconjugated bilirubin and peripheral biomarkers of inflammation derived from complete blood counts in patients with acute stage of schizophrenia. Front. Psychiatr. 2022, 13, 843985. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Weng, W.; Zhou, H.; Tang, Y.; Ding, S.; Huang, K.; Liu, Y. Elevated platelet parameter in first-episode schizophrenia patients: A cross-sectional study. J. Interferon Cytokine Res. 2020, 40, 524–529. [Google Scholar] [CrossRef]
- Özdin, S.; Sarisoy, G.; Böke, Ö. A comparison of the neutrophil-lymphocyte, platelet-lymphocyte and monocyte-lymphocyte ratios in schizophrenia and bipolar disorder patients—A retrospective file review. Nord. J. Psychiatry 2017, 71, 509–512. [Google Scholar] [CrossRef]
- Wei, Y.; Wang, T.; Li, G.; Feng, J.; Deng, L.; Xu, H.; Yin, L.; Ma, J.; Chen, D.; Chen, J. Investigation of systemic immune-inflammation index, neutrophil/high-density lipoprotein ratio, lymphocyte/high-density lipoprotein ratio, and monocyte/high-density lipoprotein ratio as indicators of inflammation in patients with schizophrenia and bipolar disorder. Front. Psychiatr. 2022, 13, 941728. [Google Scholar]
- Kılıç, N.; Tasci, G.; Yılmaz, S.; Öner, P.; Korkmaz, S. Monocyte/hdl cholesterol ratios as a new inflammatory marker in patients with schizophrenia. J. Pers. Med. 2023, 13, 276. [Google Scholar] [CrossRef] [PubMed]
- Sahpolat, M.; Ayar, D.; Ari, M.; Karaman, M.A. Elevated monocyte to high-density lipoprotein ratios as an inflammation markers for schizophrenia patients. Clin. Psychopharmacol. Neurosci. 2021, 19, 112–116. [Google Scholar] [CrossRef] [PubMed]
- Aguglia, A.; Salvi, V.; Amerio, A.; Gari, M.; Dragogna, F.; Mencacci, C.; Volpe, U.; Serafini, G.; Amore, M. Number of episodes and duration of illness associated with hypertension and 10-year cardiovascular risk in patients with bipolar disorder type i. Psychiatry Res. 2022, 308, 114344. [Google Scholar] [CrossRef] [PubMed]
- Salvi, V.; Aguglia, A.; Barone-Adesi, F.; Bianchi, D.; Donfrancesco, C.; Dragogna, F.; Palmieri, L.; Serafini, G.; Amore, M.; Mencacci, C. Cardiovascular risk in patients with severe mental illness in italy. Eur. Psychiatry J. Assoc. Eur. Psychiatr. 2020, 63, e96. [Google Scholar] [CrossRef]
- Whitcroft, K.L.; Hummel, T. Olfactory dysfunction in COVID-19. JAMA 2020, 323, 2512. [Google Scholar] [CrossRef]
- Melbourne, J.K.; Rosen, C.; Chase, K.A.; Feiner, B.; Sharma, R.P. Monocyte transcriptional profiling highlights a shift in immune signatures over the course of illness in schizophrenia. Front. Psychiatr. 2021, 12, 649494. [Google Scholar] [CrossRef] [PubMed]
- Drexhage, R.C.; van der Heul-Nieuwenhuijsen, L.; Padmos, R.C.; van Beveren, N.; Cohen, D.; Versnel, M.A.; Nolen, W.A.; Drexhage, H.A. Inflammatory gene expression in monocytes of patients with schizophrenia: Overlap and difference with bipolar disorder. A study in naturalistically treated patients. Int. J. Neuropsychopharmacol. 2010, 13, 1369–1381. [Google Scholar] [CrossRef]
- Weigelt, K.; Carvalho, L.A.; Drexhage, R.C.; Wijkhuijs, A.; Wit, H.d.; van Beveren, N.J.M.; Birkenhäger, T.K.; Bergink, V.; Drexhage, H.A. Trem-1 and dap12 expression in monocytes of patients with severe psychiatric disorders. Egr3, atf3 and pu.1 as important transcription factors. Brain Behav. Immun. 2011, 25, 1162–1169. [Google Scholar] [CrossRef]
- Yoshino, Y.; Kawabe, K.; Yamazaki, K.; Watanabe, S.; Numata, S.; Mori, Y.; Yoshida, T.; Iga, J.; Ohmori, T.; Ueno, S.-i. Elevated trem2 mrna expression in leukocytes in schizophrenia but not major depressive disorder. J. Neural Transm. 2016, 123, 637–641. [Google Scholar] [CrossRef]
- Bonartsev, P.D.; Brusov, O.S.; Morozova, M.A.; Tsutsul’kovskaia, M. Electron-microscopic study of typical lymphocytes and atypical cells of peripheral blood during the treatment of patients with schizophrenia. Zhurnal Nevropatol. I Psikhiatrii Im. SS Korsakova 2009, 109, 54–58. [Google Scholar]
- Krause, D.; Wagner, J.; Matz, J.; Weidinger, E.; Obermeier, M.; Riedel, M.; Gruber, R.; Schwarz, M.; Mueller, N. Monocytic hla dr antigens in schizophrenic patients. Neurosci. Res. 2012, 72, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Kowalski, J.; Blada, P.; Kucia, K.; Madej, A.; Herman, Z.S. Neuroleptics normalize increased release of interleukin-1β and tumor necrosis factor-α from monocytes in schizophrenia. Schizophr. Res. 2001, 50, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Krause, D.L.; Wagner, J.K.; Wildenauer, A.; Matz, J.; Weidinger, E.; Riedel, M.; Obermeier, M.; Gruber, R.; Schwarz, M.; Müller, N. Intracellular monocytic cytokine levels in schizophrenia show an alteration of il-6. Eur. Arch. Psychiatry Clin. Neurosci. 2012, 262, 393–401. [Google Scholar] [CrossRef]
- Uranova, N.A.; Bonartsev, P.D.; Androsova, L.V.; Rakhmanova, V.I.; Kaleda, V.G. Impaired monocyte activation in schizophrenia: Ultrastructural abnormalities and increased il-1β production. Eur. Arch. Psychiatry Clin. Neurosci. 2017, 267, 417–426. [Google Scholar] [CrossRef] [PubMed]
- Müller, N.; Wagner, J.K.; Krause, D.; Weidinger, E.; Wildenauer, A.; Obermeier, M.; Dehning, S.; Gruber, R.; Schwarz, M.J. Impaired monocyte activation in schizophrenia. Psychiatry Res. 2012, 198, 341–346. [Google Scholar] [CrossRef]
- Li, N.; Li, Y.; Huang, J.; Zhang, P.; Tong, J.; Chen, S.; Cui, Y.; Tan, S.; Wang, Z.; Yang, F.; et al. Dysfunctional monocytic toll-like receptor 4 signaling pathway and cognitive deficits in chronic schizophrenia patients with tardive dyskinesia. Neurosci. Lett. 2022, 777, 136581. [Google Scholar] [CrossRef]
- Chen, S.; Tian, L.; Chen, N.; Xiu, M.-H.; Wang, Z.-R.; Wang, Y.-C.; Tan, Y.-L.; Wang, C.-Y. More dampened monocytic toll-like receptor 4 response to lipopolysaccharide and its association with cognitive function in chinese han first-episode patients with schizophrenia. Schizophr. Res. 2019, 206, 300–306. [Google Scholar] [CrossRef]
- Zaki, J.K.; Lago, S.G.; Rustogi, N.; Gangadin, S.S.; Benacek, J.; van Rees, G.F.; Haenisch, F.; Broek, J.A.; Suarez-Pinilla, P.; Ruland, T.; et al. Diagnostic model development for schizophrenia based on peripheral blood mononuclear cell subtype-specific expression of metabolic markers. Transl. Psychiatry 2022, 12, 457. [Google Scholar] [CrossRef]
- Weber, N.S.; Gressitt, K.L.; Cowan, D.N.; Niebuhr, D.W.; Yolken, R.H.; Severance, E.G. Monocyte activation detected prior to a diagnosis of schizophrenia in the us military new onset psychosis project (mnopp). Schizophr. Res. 2018, 197, 465–469. [Google Scholar] [CrossRef]
- Aichholzer, M.; Gangadin, S.S.; Sommer, I.E.C.; Wijkhuis, A.; de Witte, L.D.; Kahn, R.S.; Bahn, S.; Drexhage, H.A.; Schiweck, C. Inflammatory monocyte gene signature predicts beneficial within group effect of simvastatin in patients with schizophrenia spectrum disorders in a secondary analysis of a randomized controlled trial. Brain Behav. Immun.—Health 2022, 26, 100551. [Google Scholar] [CrossRef]
- Gross, A.; Joffe, G.; Joutsiniemi, S.-L.; Nyberg, P.; Rimón, R.; Appelberg, B. Decreased production of reactive oxygen species by blood monocytes caused by clozapine correlates with eeg slowing in schizophrenic patients. Neuropsychobiology 2003, 47, 73–77. [Google Scholar] [CrossRef]
- Tomasik, J.; Schwarz, E.; Lago, S.G.; Rothermundt, M.; Leweke, F.M.; van Beveren, N.J.M.; Guest, P.C.; Rahmoune, H.; Steiner, J.; Bahn, S. Pretreatment levels of the fatty acid handling proteins h-fabp and cd36 predict response to olanzapine in recent-onset schizophrenia patients. Brain Behav. Immun. 2016, 52, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Jackson, A.J.; Miller, B.J. Meta-analysis of total and differential white blood cell counts in schizophrenia. Acta Psychiatr. Scand. 2019, 142, 18–26. [Google Scholar] [CrossRef]
- Karageorgiou, V.; Milas, G.P.; Michopoulos, I. Neutrophil-to-lymphocyte ratio in schizophrenia: A systematic review and meta-analysis. Schizophr. Res. 2019, 206, 4–12. [Google Scholar] [CrossRef]
- Yan, J.; Chen, Y.; Ju, P.; Gao, J.; Zhang, L.; Li, J.; Wang, K.; Zhang, J.; Li, C.; Xia, Q.; et al. Network association of biochemical and inflammatory abnormalities with psychiatric symptoms in first-episode schizophrenia patients. Front. Psychiatr. 2022, 13, 834539. [Google Scholar] [CrossRef] [PubMed]
- Núñez, C.; Stephan-Otto, C.; Usall, J.; Bioque, M.; Lobo, A.; González-Pinto, A.; Pina-Camacho, L.; Vieta, E.; Castro-Fornieles, J.; Rodriguez-Jimenez, R.; et al. Neutrophil count is associated with reduced gray matter and enlarged ventricles in first-episode psychosis. Schizophr. Bull. 2019, 45, 846–858. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Rizo, C.; Casanovas, M.; Fernandez-Egea, E.; Oliveira, C.; Meseguer, A.; Cabrera, B.; Mezquida, G.; Bioque, M.; Kirkpatrick, B.; Bernardo, M. Blood cell count in antipsychotic-naive patients with non-affective psychosis. Early Interv. Psychiatry 2017, 13, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Wilke, I.; Arolt, V.; Rothermundt, M.; Weitzsch, C.; Hornberg, M.; Kirchner, H. Investigations of cytokine production in whole blood cultures of paranoid and residual schizophrenic patients. Eur. Arch. Psychiatry Clin. Neurosci. 1996, 246, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Šagud, M.; Madžarac, Z.; Nedic Erjavec, G.; Šimunović Filipčić, I.; Mikulić, F.L.; Rogić, D.; Bradaš, Z.; Bajs Janović, M.; Pivac, N. The associations of neutrophil-lymphocyte, platelet-lymphocyte, monocyte-lymphocyte ratios and immune-inflammation index with negative symptoms in patients with schizophrenia. Biomolecules 2023, 13, 297. [Google Scholar] [CrossRef]
- Zhu, X.; Zhou, J.; Zhu, Y.; Yan, F.; Han, X.; Tan, Y.; Li, R. Neutrophil/lymphocyte, platelet/lymphocyte and monocyte/lymphocyte ratios in schizophrenia. Australas. Psychiatry 2021, 30, 95–99. [Google Scholar] [CrossRef] [PubMed]
- Dadouli, K.; Janho, M.B.; Hatziefthimiou, A.; Voulgaridi, I.; Piaha, K.; Anagnostopoulos, L.; Ntellas, P.; Mouchtouri, V.A.; Bonotis, K.; Christodoulou, N.; et al. Neutrophil-to-lymphocyte, monocyte-to-lymphocyte, platelet-to-lymphocyte ratio and systemic immune-inflammatory index in different states of bipolar disorder. Brain Sci. 2022, 12, 1034. [Google Scholar] [CrossRef]
- Fusar-Poli, L.; Amerio, A.; Cimpoesu, P.; Grimaldi Filioli, P.; Natale, A.; Zappa, G.; Aguglia, E.; Amore, M.; Serafini, G.; Aguglia, A. Gender differences in complete blood count and inflammatory ratios among patients with bipolar disorder. Brain Sci. 2021, 11, 363. [Google Scholar] [CrossRef]
- Fusar-Poli, L.; Natale, A.; Amerio, A.; Cimpoesu, P.; Grimaldi Filioli, P.; Aguglia, E.; Amore, M.; Serafini, G.; Aguglia, A. Neutrophil-to-lymphocyte, platelet-to-lymphocyte and monocyte-to-lymphocyte ratio in bipolar disorder. Brain Sci. 2021, 11, 58. [Google Scholar] [CrossRef] [PubMed]
- Marazziti, D.; Torrigiani, S.; Carbone, M.G.; Mucci, F.; Flamini, W.; Ivaldi, T.; Dell’Osso, L. Neutrophil/lymphocyte, platelet/lymphocyte, and monocyte/lymphocyte ratios in mood disorders. Curr. Med. Chem. 2022, 29, 5758–5781. [Google Scholar] [CrossRef] [PubMed]
- Bulut, N.S.; Yorguner, N.; Çarkaxhiu Bulut, G. The severity of inflammation in major neuropsychiatric disorders: Comparison of neutrophil–lymphocyte and platelet–lymphocyte ratios between schizophrenia, bipolar mania, bipolar depression, major depressive disorder, and obsessive compulsive disorder. Nord. J. Psychiatry 2021, 75, 624–632. [Google Scholar] [CrossRef]
- Semiz, M.; Yildirim, O.; Canan, F.; Demir, S.; Hasbek, E.; Tuman, T.C.; Kayka, N.; Tosun, M. Elevated neutrophil/lymphocyte ratio in patients with schizophrenia. Psychiatr. Danub. 2014, 26, 220–225. [Google Scholar]
- Kulaksizoglu, B.; Kulaksizoglu, S. Relationship between neutrophil/lymphocyte ratio with oxidative stress and psychopathology in patients with schizophrenia. Neuropsychiatr. Dis. Treat. 2016, 12, 1999–2005. [Google Scholar] [CrossRef] [PubMed]
- Kovács, M.Á.; Tényi, T.; Kugyelka, R.; Prenek, L.; Hau, L.; Magyar, É.E.; Herold, R.; Balogh, P.; Simon, D. Elevated osteopontin and interferon gamma serum levels and increased neutrophil-to-lymphocyte ratio are associated with the severity of symptoms in schizophrenia. Front. Psychiatr. 2020, 10, 996. [Google Scholar] [CrossRef]
- Zhou, X.; Wang, X.; Li, R.; Yan, J.; Xiao, Y.; Li, W.; Shen, H. Neutrophil-to-lymphocyte ratio is independently associated with severe psychopathology in schizophrenia and is changed by antipsychotic administration: A large-scale cross-sectional retrospective study. Front. Psychiatr. 2020, 11, 581061. [Google Scholar] [CrossRef]
- Bioque, M.; Catarina Matias-Martins, A.; Llorca-Bofí, V.; Mezquida, G.; Cuesta, M.J.; Vieta, E.; Amoretti, S.; Lobo, A.; González-Pinto, A.; Moreno, C.; et al. Neutrophil to lymphocyte ratio in patients with a first episode of psychosis: A two-year longitudinal follow-up study. Schizophr. Bull. 2022, 48, 1327–1335. [Google Scholar] [CrossRef]
- Bustan, Y.; Drapisz, A.; Ben Dor, D.H.; Avrahami, M.; Schwartz-Lifshitz, M.; Weizman, A.; Barzilay, R. Elevated neutrophil to lymphocyte ratio in non-affective psychotic adolescent inpatients: Evidence for early association between inflammation and psychosis. Psychiatry Res. 2018, 262, 149–153. [Google Scholar] [CrossRef] [PubMed]
- Özdin, S.; Böke, Ö. Neutrophil/lymphocyte, platelet/lymphocyte and monocyte/lymphocyte ratios in different stages of schizophrenia. Psychiatry Res. 2019, 271, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Balcioglu, Y.H.; Kirlioglu, S.S. C-reactive protein/albumin and neutrophil/albumin ratios as novel inflammatory markers in patients with schizophrenia. Psychiatry Investig. 2020, 17, 902–910. [Google Scholar] [CrossRef] [PubMed]
- Mijovic, A.; MacCabe, J.H. Clozapine-induced agranulocytosis. Ann. Hematol. 2020, 99, 2477–2482. [Google Scholar] [CrossRef] [PubMed]
- Labonté, C.; Zhand, N.; Park, A.; Harvey, P.D. Complete blood count inflammatory markers in treatment-resistant schizophrenia: Evidence of association between treatment responsiveness and levels of inflammation. Psychiatry Res. 2022, 308, 114382. [Google Scholar] [CrossRef] [PubMed]
- Issidorides, M.R.; Stefanis, C.N.; Varsou, E.; Katsorchis, T. Altered chromatin ultrastructure in neutrophils of schizophrenics. Nature 1975, 258, 612–614. [Google Scholar] [CrossRef] [PubMed]
- De Campos-Carli, S.M.; Araújo, M.S.; de Oliveira Silveira, A.C.; de Rezende, V.B.; Rocha, N.P.; Ferretjans, R.; Ribeiro-Santos, R.; Teixeira-Carvalho, A.; Martins-Filho, O.A.; Berk, M.; et al. Cannabinoid receptors on peripheral leukocytes from patients with schizophrenia: Evidence for defective immunomodulatory mechanisms. J. Psychiatr. Res. 2017, 87, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Rwegellera, G.G.; Fernando, K.A.; Okong’o, O. Bactericidal activity of neutrophils of schizophrenic patients. Med. J. Zamb. 1982, 16, 21–22. [Google Scholar]
- Shcherbakova, I.V.; Neshkova, E.A.; Dotsenko, V.L.; Kozlov, L.V.; Mishin, A.A.; Platonova, T.P.; Shcherbakova, E.G.; Iarovaia, G.A. Activation of kallikrein-kinin system, degranulating activity of neutrophils and blood-brain barrier in schizophrenia. Zhurnal Nevropatol. I Psikhiatrii Im. SS Korsakova 1998, 98, 38–41. [Google Scholar]
- Tsavaris, N.; Konstantopoulos, K.; Vaidakis, S.; Koumakis, K.; Pangalis, G. Cytochemical determination of monoamine oxidase activity in lymphocytes and neutrophils of schizophrenic patients. Haematologia 1995, 26, 143–146. [Google Scholar]
- Srivastava, N.; Barthwal, M.; Dalal, P.; Agarwal, A.; Nag, D.; Srimal, R.; Seth, P.; Dikshit, M. Nitrite content and antioxidant enzyme levels in the blood of schizophrenia patients. Psychopharmacology 2001, 158, 140–145. [Google Scholar] [CrossRef] [PubMed]
- Melamed, Y.; Sirota, P.; Dicker, D.R.; Fishman, P. Superoxide anion production by neutrophils derived from peripheral blood of schizophrenic patients. Psychiatry Res. 1998, 77, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Sirota, P.; Gavrieli, R.; Wolach, B. Overproduction of neutrophil radical oxygen species correlates with negative symptoms in schizophrenic patients: Parallel studies on neutrophil chemotaxis, superoxide production and bactericidal activity. Psychiatry Res. 2003, 121, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Karpiński, P.; Frydecka, D.; Sąsiadek, M.M.; Misiak, B. Reduced number of peripheral natural killer cells in schizophrenia but not in bipolar disorder. Brain Behav. Immun. 2016, 54, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Karpiński, P.; Samochowiec, J.; Frydecka, D.; Sąsiadek, M.M.; Misiak, B. Further evidence for depletion of peripheral blood natural killer cells in patients with schizophrenia: A computational deconvolution study. Schizophr. Res. 2018, 201, 243–248. [Google Scholar] [CrossRef]
- Sperner-Unterweger, B.; Whitworth, A.; Kemmler, G.; Hilbe, W.; Thaler, J.; Weiss, G.; Fleischhacker, W.W. T-cell subsets in schizophrenia: A comparison between drug-naive first episode patients and chronic schizophrenic patients. Schizophr. Res. 1999, 38, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Rudolf, S.; Schlenke, P.; Broocks, A.; Peters, M.; Rothermundt, M.; Arolt, V.; Kirchner, H. Search for atypical lymphocytes in schizophrenia. World J. Biol. Psychiatry 2004, 5, 33–37. [Google Scholar] [CrossRef]
- Abdeljaber, M.H.; Nair, M.P.N.; Schork, M.A.; Schwartz, S.A. Depressed natural killer cell activity in schizophrenic patients. Immunol. Investig. 1994, 23, 259–268. [Google Scholar] [CrossRef] [PubMed]
- DeLisi, L.E.; Ortaldo, J.R.; Maluish, A.E.; Wyatt, R.J. Deficient natural killer cell (nk) activity and macrophage functioning in schizophrenic patients. J. Neural Transm. 1983, 58, 99–106. [Google Scholar] [CrossRef]
- McDaniel, J.; Jewart, R.; Eccard, M.; Pollard, W.; Caudle, J.; Stipetic, M.; Risby, E.; Lewine, R.; Risch, S. Natural killer cell activity in schizophrenia and schizoaffective disorder: A pilot study. Schizophr. Res. 1992, 8, 125–128. [Google Scholar] [CrossRef]
- Yovel, G.; Sirota, P.; Mazeh, D.; Shakhar, G.; Rosenne, E.; Ben-Eliyahu, S. Higher natural killer cell activity in schizophrenic patients: The impact of serum factors, medication, and smoking. Brain Behav. Immun. 2000, 14, 153–169. [Google Scholar] [CrossRef] [PubMed]
- Tarantino, N.; Leboyer, M.; Bouleau, A.; Hamdani, N.; Richard, J.R.; Boukouaci, W.; Ching-Lien, W.; Godin, O.; Bengoufa, D.; Le Corvoisier, P.; et al. Natural killer cells in first-episode psychosis: An innate immune signature? Mol. Psychiatry 2021, 26, 5297–5306. [Google Scholar] [CrossRef] [PubMed]
- Steiner, J.; Westphal, S.; Schroeter, M.L.; Schiltz, K.; Jordan, W.; Müller, U.J.; Bernstein, H.-G.; Bogerts, B.; Schmidt, R.E.; Jacobs, R. Increased s100b+ nk cell counts in acutely ill schizophrenia patients are correlated with the free cortisol index, but not with s100b serum levels. Brain Behav. Immun. 2012, 26, 564–567. [Google Scholar] [CrossRef] [PubMed]
- Borovcanin, M.M.; Minic Janicijevic, S.; Jovanovic, I.P.; Gajovic, N.M.; Jurisevic, M.M.; Arsenijevic, N.N. Type 17 immune response facilitates progression of inflammation and correlates with cognition in stable schizophrenia. Diagnostics 2020, 10, 926. [Google Scholar] [CrossRef] [PubMed]
- Chandley, M.J.; Miller, M.N.; Kwasigroch, C.N.; Wilson, T.D.; Miller, B.E. Increased antibodies for the α7 subunit of the nicotinic receptor in schizophrenia. Schizophr. Res. 2009, 109, 98–101. [Google Scholar] [CrossRef]
- Keshavarz, F.; Soltani, M.; Mokhtarian, K.; Beshkar, P.; Majidi, J.; Azadegan-Dehkordi, F.; Anjomshoa, M.; Bagheri, N. Autoantibodies against central nervous system antigens and the serum levels of il-32 in patients with schizophrenia. Neuroimmunomodulation 2022, 29, 493–499. [Google Scholar] [CrossRef]
- Shiwaku, H.; Nakano, Y.; Kato, M.; Takahashi, H. Detection of autoantibodies against gabaarα1 in patients with schizophrenia. Schizophr. Res. 2020, 216, 543–546. [Google Scholar] [CrossRef]
- Soltani, M.; Beshkar, P.; Mokhtarian, K.; Anjomshoa, M.; Mohammad Rezaei, M.; Azadegan-Dehkordi, F.; Mirzaei, Y.; Majidi, J.; Bagheri, N. A study of autoantibodies against some central nervous system antigens and the il-35 serum level in schizophrenia. Iran. J. Allergy Asthma Immunol. 2022, 21, 441–448. [Google Scholar] [CrossRef]
- Hallford, P.; Clair, D.S.; Halley, L.; Mustard, C.; Wei, J. A study of type-1 diabetes associated autoantibodies in schizophrenia. Schizophr. Res. 2016, 176, 186–190. [Google Scholar] [CrossRef]
- Hoffmann, C.; Stevens, J.; Zong, S.; van Kruining, D.; Saxena, A.; Küçükali, C.İ.; Tüzün, E.; Yalçınkaya, N.; De Hert, M.; González-Vioque, E.; et al. Alpha7 acetylcholine receptor autoantibodies are rare in sera of patients diagnosed with schizophrenia or bipolar disorder. PLoS ONE 2018, 13, e0208412. [Google Scholar] [CrossRef]
- Zhou, D.; Xie, C.; Li, X.; Song, N.; Kou, Z.; Zhang, T.; Yuan, T.-F. Rare presence of autoantibodies targeting to nmda and gabaa receptors in schizophrenia patients. Schizophr. Res. 2022, 249, 93–97. [Google Scholar] [CrossRef] [PubMed]
- Ermakov, E.A.; Ivanova, S.A.; Buneva, V.N.; Nevinsky, G.A. Blood-derived rna- and microrna-hydrolyzing igg antibodies in schizophrenia patients. Biochemistry 2018, 83, 507–526. [Google Scholar] [CrossRef] [PubMed]
- Ermakov, E.A.; Smirnova, L.P.; Parkhomenko, T.A.; Dmitrenok, P.S.; Krotenko, N.M.; Fattakhov, N.S.; Bokhan, N.A.; Semke, A.V.; Ivanova, S.A.; Buneva, V.N.; et al. DNA-hydrolysing activity of igg antibodies from the sera of patients with schizophrenia. Open Biol. 2015, 5, 150064. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.J.; Lee, S.J.; Toh, K.Y.; Lee, C.U.; Lee, C.; Paik, I.H. Identification of antibodies to heat shock proteins 90 kda and 70 kda in patients with schizophrenia. Schizophr. Res. 2001, 52, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-F.; Wang, D.; Zhu, W.; Delrahim, K.K.; Dolnak, D.; Hyman Rapaport, M. Studies characterizing 60 kda autoantibodies in subjects with schizophrenia. Biol. Psychiatry 2003, 53, 361–375. [Google Scholar] [CrossRef]
- Endres, D.; von Zedtwitz, K.; Matteit, I.; Bünger, I.; Foverskov-Rasmussen, H.; Runge, K.; Feige, B.; Schlump, A.; Maier, S.; Nickel, K.; et al. Spectrum of novel anti–central nervous system autoantibodies in the cerebrospinal fluid of 119 patients with schizophreniform and affective disorders. Biol. Psychiatry 2022, 92, 261–274. [Google Scholar] [CrossRef]
- Klyushnik, T.P.; Danilovskaya, E.V.; Vatolkina, O.E.; Turkova, I.L.; Tsutsul’kovskaya, M.Y.; Orlova, V.A.; Kaleda, V.G. Changes in serum levels of autoantibody to nerve growth factor in patients with schizophrenia. Neurosci. Behav. Physiol. 1999, 29, 355–357. [Google Scholar] [CrossRef]
- Sugiura, M.; Yokoi, Y.; Maruyama, S.; Ishido, T.; Tokunaga, Y.; Sasaki, K. Detection of anti-cerebral autoantibodies in schizophrenia and alzheimer’s disease. J. Clin. Lab. Immunol. 1989, 28, 1–3. [Google Scholar]
- Tanaka, S.; Matsunaga, H.; Kimura, M.; Tatsumi, K.-i.; Hidaka, Y.; Takano, T.; Uema, T.; Takeda, M.; Amino, N. Autoantibodies against four kinds of neurotransmitter receptors in psychiatric disorders. J. Neuroimmunol. 2003, 141, 155–164, Erratum in J. Neuroimmunol. 2003, 144, 148. [Google Scholar] [CrossRef]
- Jin, S.-Z.; Wu, N.; Xu, Q.; Zhang, X.; Ju, G.-Z.; Law, M.H.; Wei, J. A study of circulating gliadin antibodies in schizophrenia among a chinese population. Schizophr. Bull. 2012, 38, 514–518. [Google Scholar] [CrossRef]
- Kelly, D.L.; Demyanovich, H.K.; Eaton, W.W.; Cascella, N.; Jackson, J.; Fasano, A.; Carpenter, W.T. Anti gliadin antibodies (aga igg) related to peripheral inflammation in schizophrenia. Brain Behav. Immun. 2018, 69, 57–59. [Google Scholar] [CrossRef]
- Li, C.; Whelan, R.; Yang, H.; Jiang, Y.; Qiu, C.; Meng, Q.; Wei, J. Anti-tsnare1 igg plasma levels differ by sex in patients with schizophrenia in a chinese population. FEBS Open Bio 2019, 9, 1705–1712. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Zhang, X.; Wang, J.; Yang, H.; Jiang, Y.; Qiu, C.; Meng, Q. Analysis of plasma autoantibodies for inflammatory cytokines in patients with first-episode schizophrenia among a chinese population. J. Neuroimmunol. 2020, 341, 577165. [Google Scholar] [CrossRef]
- Okusaga, O.; Yolken, R.H.; Langenberg, P.; Sleemi, A.; Kelly, D.L.; Vaswani, D.; Giegling, I.; Hartmann, A.M.; Konte, B.; Friedl, M.; et al. Elevated gliadin antibody levels in individuals with schizophrenia. World J. Biol. Psychiatry 2013, 14, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Steiner, J.; Jacobs, R.; Panteli, B.; Brauner, M.; Schiltz, K.; Bahn, S.; Herberth, M.; Westphal, S.; Gos, T.; Walter, M.; et al. Acute schizophrenia is accompanied by reduced t cell and increased b cell immunity. Eur. Arch. Psychiatry Clin. Neurosci. 2010, 260, 509–518. [Google Scholar] [CrossRef] [PubMed]
- McAllister, C.G. Increased numbers of cd5 b lymphocytes in schizophrenic patients. Arch. Gen. Psychiatry 1989, 46, 890. [Google Scholar] [CrossRef] [PubMed]
- Printz, D.J.; Strauss, D.H.; Goetz, R.; Sadiq, S.; Malaspina, D.; Krolewski, J.; Gorman, J.M. Elevation of cd5+ b lymphocytes in schizophrenia. Biol. Psychiatry 1999, 46, 110–118. [Google Scholar] [CrossRef]
- Ganguli, R.; Rabin, B.S. Cd5 positive b lymphocytes in schizophrenia: No alteration in numbers or percentage as compared with control subjects. Psychiatry Res. 1993, 48, 69–78. [Google Scholar] [CrossRef]
- Müller, N.; Hofschuster, E.; Ackenheil, M.; Eckstein, R. T-cells and psychopathology in schizophrenia: Relationship to the outcome of neuroleptic therapy. Acta Psychiatr. Scand. 1993, 87, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Saĭfulina, N.A.; Arkhipov, G.S.; Stepanov, A.F.; Gusev Iu, A. T- and b-lymphocytes and their clinico-pathogenetic significance in patients with paranoid schizophrenia. Zhurnal Nevropatol. I Psikhiatrii Im. SS Korsakova 1990, 90, 87–90. [Google Scholar]
- Achiron, A.; Noy, S.; Pras, E.; Lereya, J.; Hermesh, H.; Laor, N. T-cell subsets in acute psychotic schizophrenic patients. Biol. Psychiatry 1994, 35, 27–31. [Google Scholar] [CrossRef] [PubMed]
- Sahbaz, C.; Zibandey, N.; Kurtulmus, A.; Duran, Y.; Gokalp, M.; Kırpınar, I.; Sahin, F.; Guloksuz, S.; Akkoc, T. Reduced regulatory t cells with increased proinflammatory response in patients with schizophrenia. Psychopharmacology 2020, 237, 1861–1871. [Google Scholar] [CrossRef] [PubMed]
- Theodoropoulou, S.; Spanakos, G.; Baxevanis, C.N.; Economou, M.; Gritzapis, A.D.; Papamichail, M.P.; Stefanis, C.N. Cytokine serum levels, autologous mixed lymphocyte reaction and surface marker analysis in never medicated and chronically medicated schizophrenic patients. Schizophr. Res. 2001, 47, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Bessler, H.; Levental, Z.; Karp, L.; Modai, I.; Djaldetti, M.; Weizman, A. Cytokine production in drug-free and neuroleptic-treated schizophrenic patients. Biol. Psychiatry 1995, 38, 297–302. [Google Scholar] [CrossRef] [PubMed]
- Villemain, F.; Chatenoud, L.; Galinowski, A.; Homo-Delarche, F.; Ginestet, D.; Loo, H.; Zarifian, E.; Bach, J.F. Aberrant t cell-mediated immunity in untreated schizophrenic patients: Deficient interleukin-2 production. Am. J. Psychiatry 1989, 146, 609–616. [Google Scholar] [PubMed]
- Yang Zhang, X.; Feng Zhou, D.; Yuan Cao, L.; Yan Zhang, P.; Ying Wu, G. Decreased production of interleukin-2 (il-2), il-2 secreting cells and cd4+ cells in medication-free patients with schizophrenia. J. Psychiatr. Res. 2002, 36, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Ding, M.; Song, X.; Zhao, J.; Gao, J.; Li, X.; Yang, G.; Wang, X.; Harrington, A.; Fan, X.; Lv, L. Activation of th17 cells in drug naïve, first episode schizophrenia. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2014, 51, 78–82. [Google Scholar]
- Kelly, D.L.; Li, X.; Kilday, C.; Feldman, S.; Clark, S.; Liu, F.; Buchanan, R.W.; Tonelli, L.H. Increased circulating regulatory t cells in medicated people with schizophrenia. Psychiatry Res. 2018, 269, 517–523. [Google Scholar] [CrossRef]
- Varun, C.N.; Venkataswamy, M.M.; Ravikumar, R.; Nagaraju, R.; Debnath, M.; Varambally, S.; Venkatasubramanian, G.; Ravi, V. Th17 and mait cell mediated inflammation in antipsychotic free schizophrenia patients. Schizophr. Res. 2019, 212, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Vergaelen, E.; Schiweck, C.; Van Steeland, K.; Counotte, J.; Veling, W.; Swillen, A.; Drexhage, H.; Claes, S. A pilot study on immuno-psychiatry in the 22q11.2 deletion syndrome: A role for th17 cells in psychosis? Brain Behav. Immun. 2018, 70, 88–95. [Google Scholar] [CrossRef]
- Amerio, A.; Magnani, L.; Arduino, G.; Fesce, F.; de Filippis, R.; Parise, A.; Costanza, A.; Nguyen, K.D.; Saverino, D.; De Berardis, D.; et al. Immunomodulatory effects of clozapine: More than just a side effect in schizophrenia. Curr. Neuropharmacol. 2023; in press. [Google Scholar]
- Boneberg, E.-M.; von Seydlitz, E.; Pröpster, K.; Watzl, H.; Rockstroh, B.; Illges, H. D3 dopamine receptor mrna is elevated in t cells of schizophrenic patients whereas d4 dopamine receptor mrna is reduced in cd4+-t cells. J. Neuroimmunol. 2006, 173, 180–187. [Google Scholar] [CrossRef] [PubMed]
- Brito-Melo, G.E.A.; Nicolato, R.; de Oliveira, A.C.P.; Menezes, G.B.; Lélis, F.J.N.; Avelar, R.S.; Sá, J.; Bauer, M.E.; Souza, B.R.; Teixeira, A.L.; et al. Increase in dopaminergic, but not serotoninergic, receptors in t-cells as a marker for schizophrenia severity. J. Psychiatr. Res. 2012, 46, 738–742. [Google Scholar] [CrossRef]
- Hu, A.; Li, F.; Guo, L.; Zhao, X.; Xiang, X. Mitochondrial damage of lymphocytes in patients with acute relapse of schizophrenia: A correlational study with efficacy and clinical symptoms. Neuropsychiatr. Dis. Treat. 2022, 18, 2455–2466. [Google Scholar] [CrossRef]
- Luo, C.; Pi, X.; Hu, N.; Wang, X.; Xiao, Y.; Li, S.; Sweeney, J.A.; Bishop, J.R.; Gong, Q.; Xie, D.; et al. Subtypes of schizophrenia identified by multi-omic measures associated with dysregulated immune function. Mol. Psychiatry 2021, 26, 6926–6936. [Google Scholar] [CrossRef] [PubMed]
- Lynall, M.-E.; Soskic, B.; Hayhurst, J.; Schwartzentruber, J.; Levey, D.F.; Pathak, G.A.; Polimanti, R.; Gelernter, J.; Stein, M.B.; Trynka, G.; et al. Genetic variants associated with psychiatric disorders are enriched at epigenetically active sites in lymphoid cells. Nat. Commun. 2022, 13, 6102. [Google Scholar] [CrossRef]
- Farsi, Z.; Sheng, M. Molecular mechanisms of schizophrenia: Insights from human genetics. Curr. Opin. Neurobiol. 2023, 81, 102731. [Google Scholar] [CrossRef] [PubMed]
- Sheridan, S.D.; Horng, J.E.; Perlis, R.H. Patient-Derived In Vitro Models of Microglial Function and Synaptic Engulfment in Schizophrenia. Biol. Psychiatry 2022, 92, 470–479. [Google Scholar] [CrossRef]
- Comer, A.L.; Jinadasa, T.; Sriram, B.; Phadke, R.A.; Kretsge, L.N.; Nguyen, T.P.H.; Antognetti, G.; Gilbert, J.P.; Lee, J.; Newmark, E.R.; et al. Increased expression of schizophrenia-associated gene C4 leads to hypoconnectivity of prefrontal cortex and reduced social interaction. PLoS Biol. 2020, 18, e3000604. [Google Scholar] [CrossRef]
- Sellgren, C.M.; Gracias, J.; Watmuff, B.; Biag, J.D.; Thanos, J.M.; Whittredge, P.B.; Fu, T.; Worringer, K.; Brown, H.E.; Wang, J.; et al. Increased synapse elimination by microglia in schizophrenia patient-derived models of synaptic pruning. Nat. Neurosci. 2019, 22, 374–385. [Google Scholar] [CrossRef]
- Park, G.H.; Noh, H.; Shao, Z.; Ni, P.; Qin, Y.; Liu, D.; Beaudreault, C.P.; Park, J.S.; Abani, C.P.; Park, J.M.; et al. Activated microglia cause metabolic disruptions in developmental cortical interneurons that persist in interneurons from individuals with schizophrenia. Nat. Neurosci. 2020, 23, 1352–1364. [Google Scholar] [CrossRef] [PubMed]
- Breitmeyer, R.; Vogel, S.; Heider, J.; Hartmann, S.M.; Wüst, R.; Keller, A.L.; Binner, A.; Fitzgerald, J.C.; Fallgatter, A.J.; Volkmer, H. Regulation of synaptic connectivity in schizophrenia spectrum by mutual neuron-microglia interaction. Commun. Biol. 2023, 6, 472. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, P.; Kaur, G.; Prasad, R.; Singh, H. Pharmacotherapy of schizophrenia: Immunological aspects and potential role of immunotherapy. Expert Rev. Neurother. 2021, 21, 1441–1453. [Google Scholar] [CrossRef] [PubMed]
- Miller, B.J.; Goldsmith, D.R. Towards an Immunophenotype of Schizophrenia: Progress, Potential Mechanisms, and Future Directions. Neuropsychopharmacology 2017, 42, 299–317. [Google Scholar] [CrossRef] [PubMed]
- Bejerot, S.; Sigra Stein, S.; Welin, E.; Eklund, D.; Hylén, U.; Humble, M.B. Rituximab as an Adjunctive Treatment for Schizophrenia Spectrum Disorder or Obsessive-Compulsive Disorder: Two Open-Label Pilot Studies on Treatment-Resistant Patients. J. Psychiatr. Res. 2023, 158, 319–329. [Google Scholar] [CrossRef] [PubMed]
- Karbalaee, M.; Jameie, M.; Amanollahi, M.; TaghaviZanjani, F.; Parsaei, M.; Basti, F.A.; Mokhtari, S.; Moradi, K.; Ardakani, M.-R.K.; Akhondzadeh, S. Efficacy and Safety of Adjunctive Therapy with Fingolimod in Patients with Schizophrenia: A Randomized, Double-Blind, Placebo-Controlled Clinical Trial. Schizophr. Res. 2023, 254, 92–98. [Google Scholar] [CrossRef]
- Miller, B.J.; Dias, J.K.; Lemos, H.P.; Buckley, P.F. An Open-Label, Pilot Trial of Adjunctive Tocilizumab in Schizophrenia. J. Clin. Psychiatry 2016, 77, 13353. [Google Scholar] [CrossRef] [PubMed]
- Motamed, M.; Karimi, H.; Sanjari Moghaddam, H.; Taherzadeh Boroujeni, S.; Sanatian, Z.; Hasanzadeh, A.; Khodaei Ardakani, M.-R.; Akhondzadeh, S. Risperidone Combination Therapy with Adalimumab for Treatment of Chronic Schizophrenia. Int. Clin. Psychopharmacol. 2022, 37, 92–101. [Google Scholar] [CrossRef]
- Levine, J.; Gutman, J.; Feraro, R.; Levy, P.; Kimhi, R.; Leykin, I.; Deckmann, M.; Handzel, Z.T.; Shinitzky, M. Side effect profile of azathioprine in the treatment of chronic schizophrenic patients. Neuropsychobiology 1997, 36, 172–176. [Google Scholar] [CrossRef]
- Nitta, M.; Kishimoto, T.; Müller, N.; Weiser, M.; Davidson, M.; Kane, J.M.; Correll, C.U. Adjunctive Use of Nonsteroidal Anti-Inflammatory Drugs for Schizophrenia: A Meta-Analytic Investigation of Randomized Controlled Trials. Schizophr. Bull. 2013, 39, 1230–1241. [Google Scholar] [CrossRef]
- Chaudhry, I.B.; Husain, M.O.; Khoso, A.B.; Husain, M.I.; Buch, M.H.; Kiran, T.; Fu, B.; Bassett, P.; Qurashi, I.; ur Rahman, R.; et al. A Randomised Clinical Trial of Methotrexate Points to Possible Efficacy and Adaptive Immune Dysfunction in Psychosis. Transl. Psychiatry 2020, 10, 415. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Song, X.; Zhao, M.; Jarskog, L.F.; Natarajan, R.; Shukair, N.; Freudenreich, O.; Henderson, D.C.; Goff, D.C. The Effect of Adjunctive Telmisartan Treatment on Psychopathology and Cognition in Patients with Schizophrenia. Acta Psychiatr. Scand. 2017, 136, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Sagheddu, C.; Melis, M.; Muntoni, A.L.; Pistis, M. Repurposing Peroxisome Proliferator-Activated Receptor Agonists in Neurological and Psychiatric Disorders. Pharmaceuticals 2021, 14, 1025. [Google Scholar] [CrossRef] [PubMed]
- Çakici, N.; van Beveren, N.J.M.; Judge-Hundal, G.; Koola, M.M.; Sommer, I.E.C. An update on the efficacy of anti-inflammatory agents for patients with schizophrenia: A meta-analysis. Psychol. Med. 2019, 49, 2307–2319. [Google Scholar] [CrossRef] [PubMed]
- Messina, A.; Concerto, C.; Rodolico, A.; Petralia, A.; Caraci, F.; Signorelli, M.S. Is It Time for a Paradigm Shift in the Treatment of Schizophrenia? The Use of Inflammation-Reducing and Neuroprotective Drugs—A Review. Brain Sci. 2023, 13, 957. [Google Scholar] [CrossRef]
- Girgis, R.R.; Ciarleglio, A.; Choo, T.; Haynes, G.; Bathon, J.M.; Cremers, S.; Kantrowitz, J.T.; Lieberman, J.A.; Brown, A.S. A Randomized, Double-Blind, Placebo-Controlled Clinical Trial of Tocilizumab, an Interleukin-6 Receptor Antibody, for Residual Symptoms in Schizophrenia. Neuropsychopharmacology 2017, 43, 1317–1323. [Google Scholar] [CrossRef]
- Hansen, N.; Malchow, B. Monoclonal Antibody Therapy in Autoantibody-Associated Psychotic Disorders and Schizophrenia: Narrative Review of Past and Current Clinical Trials. Psychiatr. Danub. 2023, 35, 8–15. [Google Scholar] [CrossRef]
- Garner, B.; Phillips, L.J.; Bendall, S.; Hetrick, S.E. Antiglucocorticoid and related treatments for psychosis. Cochrane Database Syst. Rev. 2016, 2016, CD006995. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguyen, K.D.; Amerio, A.; Aguglia, A.; Magnani, L.; Parise, A.; Conio, B.; Serafini, G.; Amore, M.; Costanza, A. Microglia and Other Cellular Mediators of Immunological Dysfunction in Schizophrenia: A Narrative Synthesis of Clinical Findings. Cells 2023, 12, 2099. https://doi.org/10.3390/cells12162099
Nguyen KD, Amerio A, Aguglia A, Magnani L, Parise A, Conio B, Serafini G, Amore M, Costanza A. Microglia and Other Cellular Mediators of Immunological Dysfunction in Schizophrenia: A Narrative Synthesis of Clinical Findings. Cells. 2023; 12(16):2099. https://doi.org/10.3390/cells12162099
Chicago/Turabian StyleNguyen, Khoa D., Andrea Amerio, Andrea Aguglia, Luca Magnani, Alberto Parise, Benedetta Conio, Gianluca Serafini, Mario Amore, and Alessandra Costanza. 2023. "Microglia and Other Cellular Mediators of Immunological Dysfunction in Schizophrenia: A Narrative Synthesis of Clinical Findings" Cells 12, no. 16: 2099. https://doi.org/10.3390/cells12162099