
Synovial Fluid Derived from Human Knee Osteoarthritis Increases the Viability of Human Adipose-Derived Stem Cells through Upregulation of FOSL1
 , , , ,
, , , ,  , , , , ,
, , , , ,
Abstract
:1. Introduction
1.1. Widespread Use of Adipose-Derived Stem Cells (ADSCs)
1.2. ADSC Treatment for Osteoarthritis of the Knee
1.3. Availability of ADSCs in the Joint Cavity
1.4. Properties of SF
1.5. Effect of SF on ADSCs
2. Materials and Methods
2.1. Collection and Evaluation of SF
2.2. Effect of SF on ADSC Viability
2.3. Morphological Cell Evaluation
2.4. Transcriptome Analysis Using DNA Microarrays
2.5. RT-PCR (Reverse Transcription Polymerase Chain Reaction)
2.6. Western Blot Analysis
2.7. Knockdown of FOSL1 Gene by Small Interfering RNA (siRNA)
2.8. Comparison of the Viability of FOSL1-Knockdown ADSCs and the siControl Group in SF
2.9. Statistical Analysis
3. Results
3.1. SF Properties
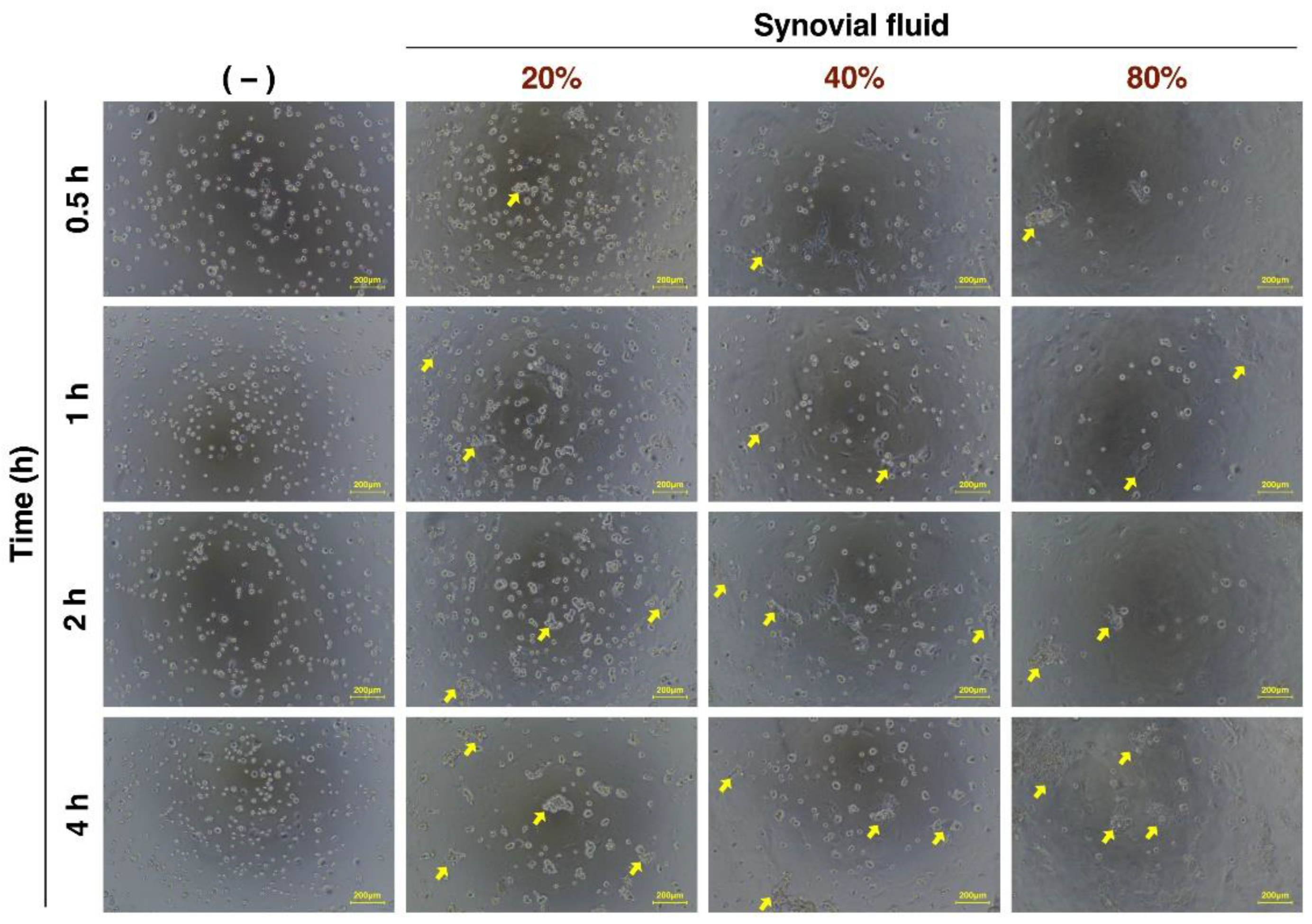
3.2. Evaluation of ADSC Morphology Incubated with Various Concentrations of SF over Time
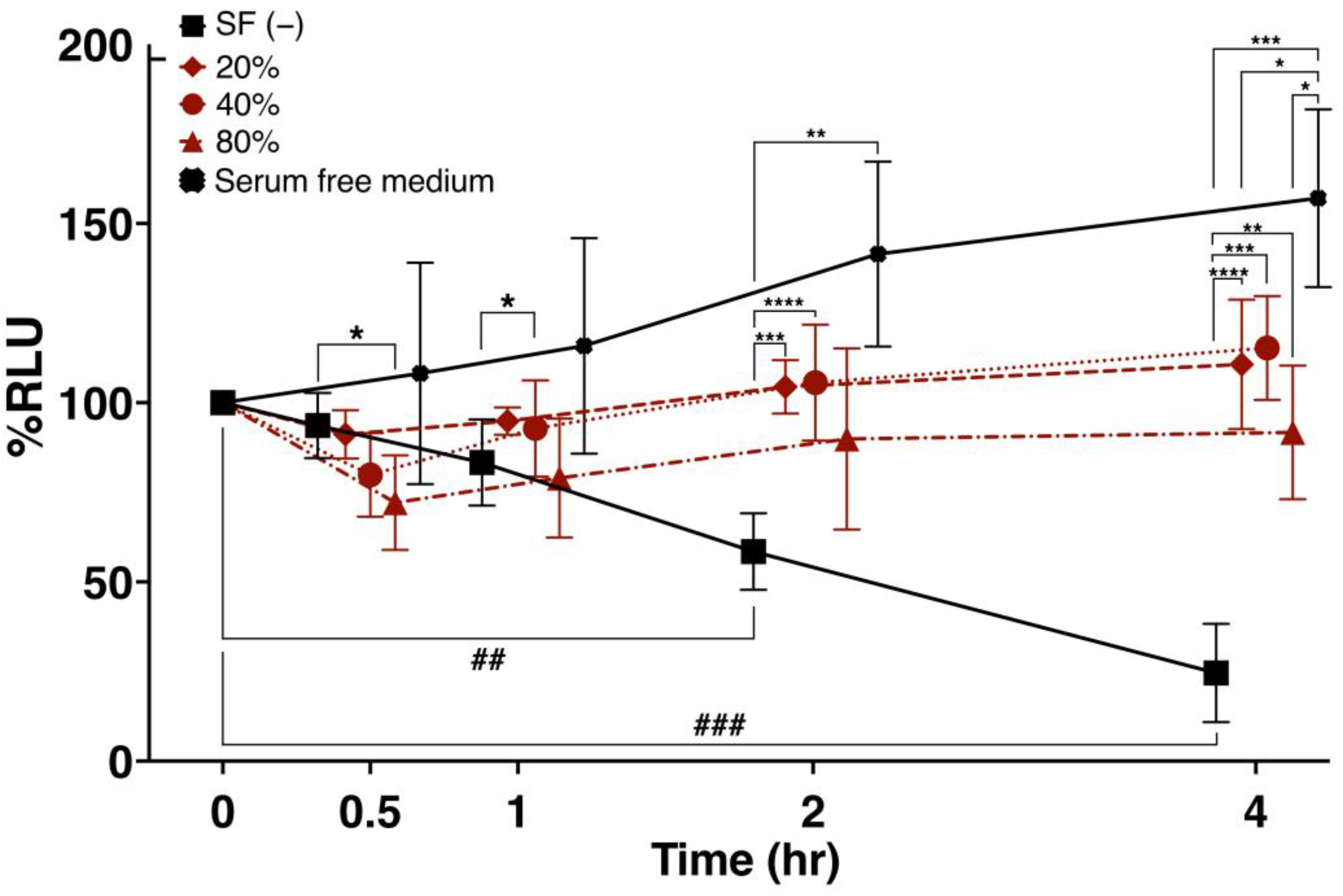
3.3. Viability of Cultured ADSCs in Response to SF Addition
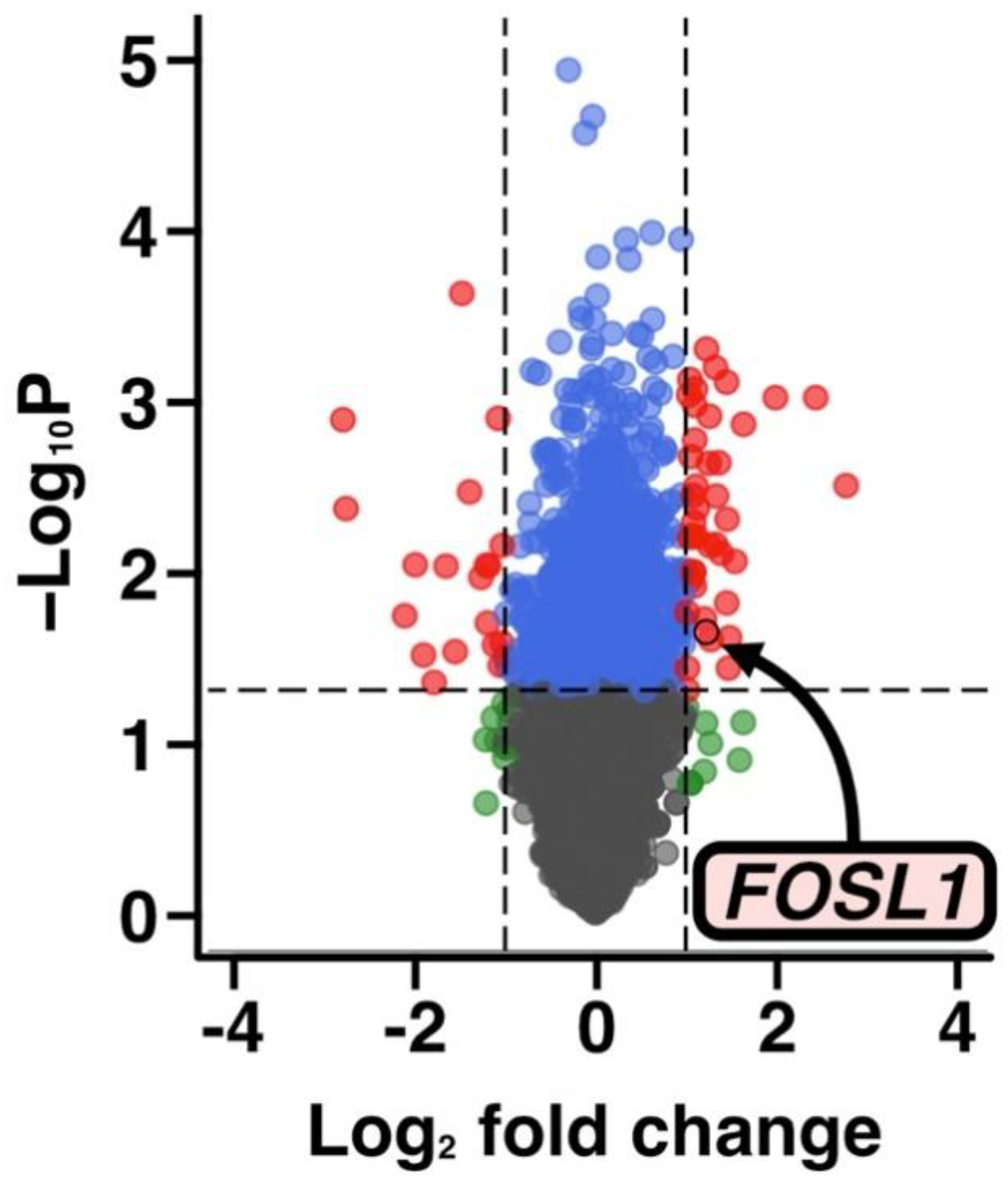
3.4. Transcriptome Analysis of Changes in Gene Expression following SF Treatment
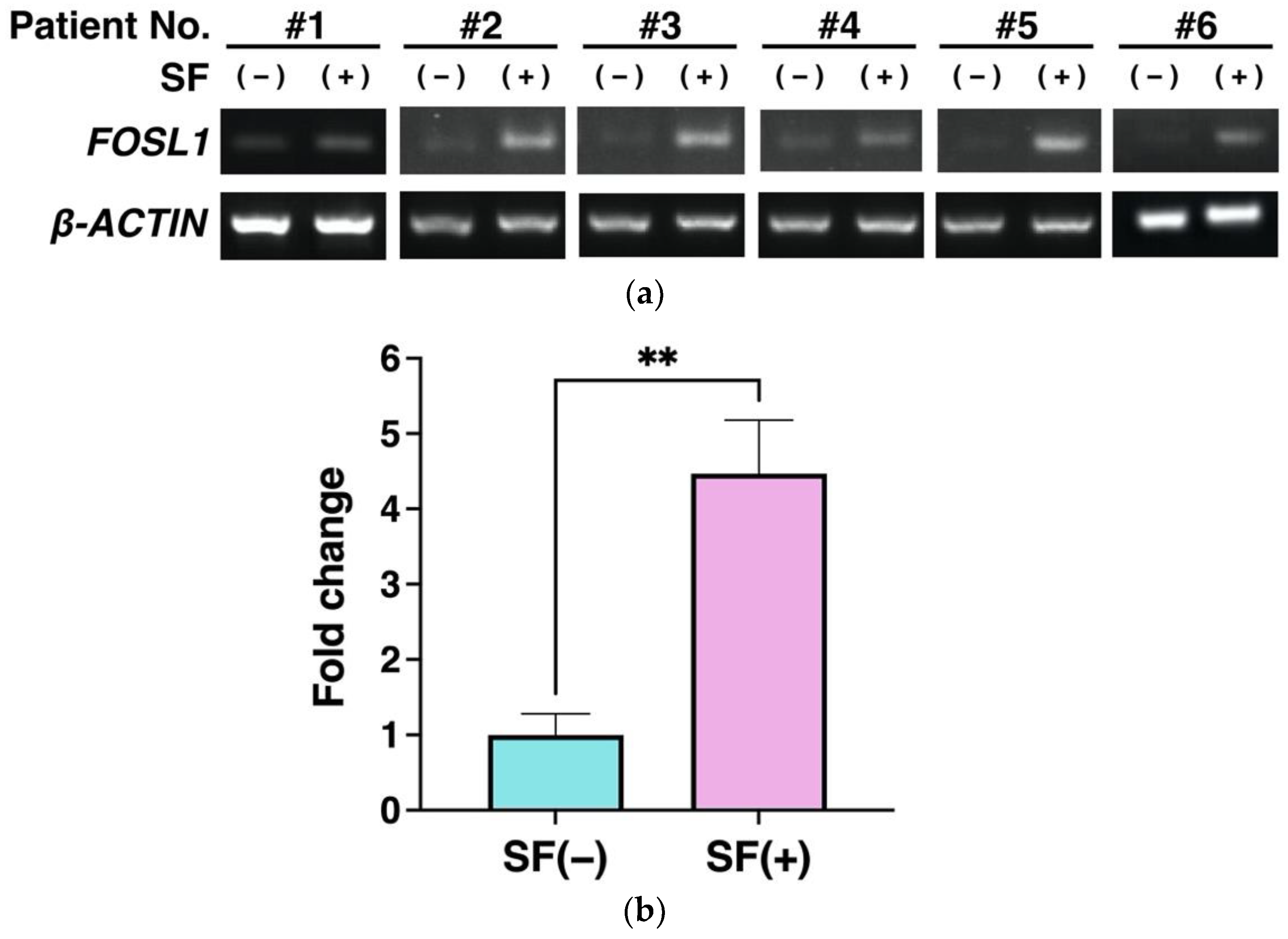
3.5. Gene Expression Analysis Using RT-PCR
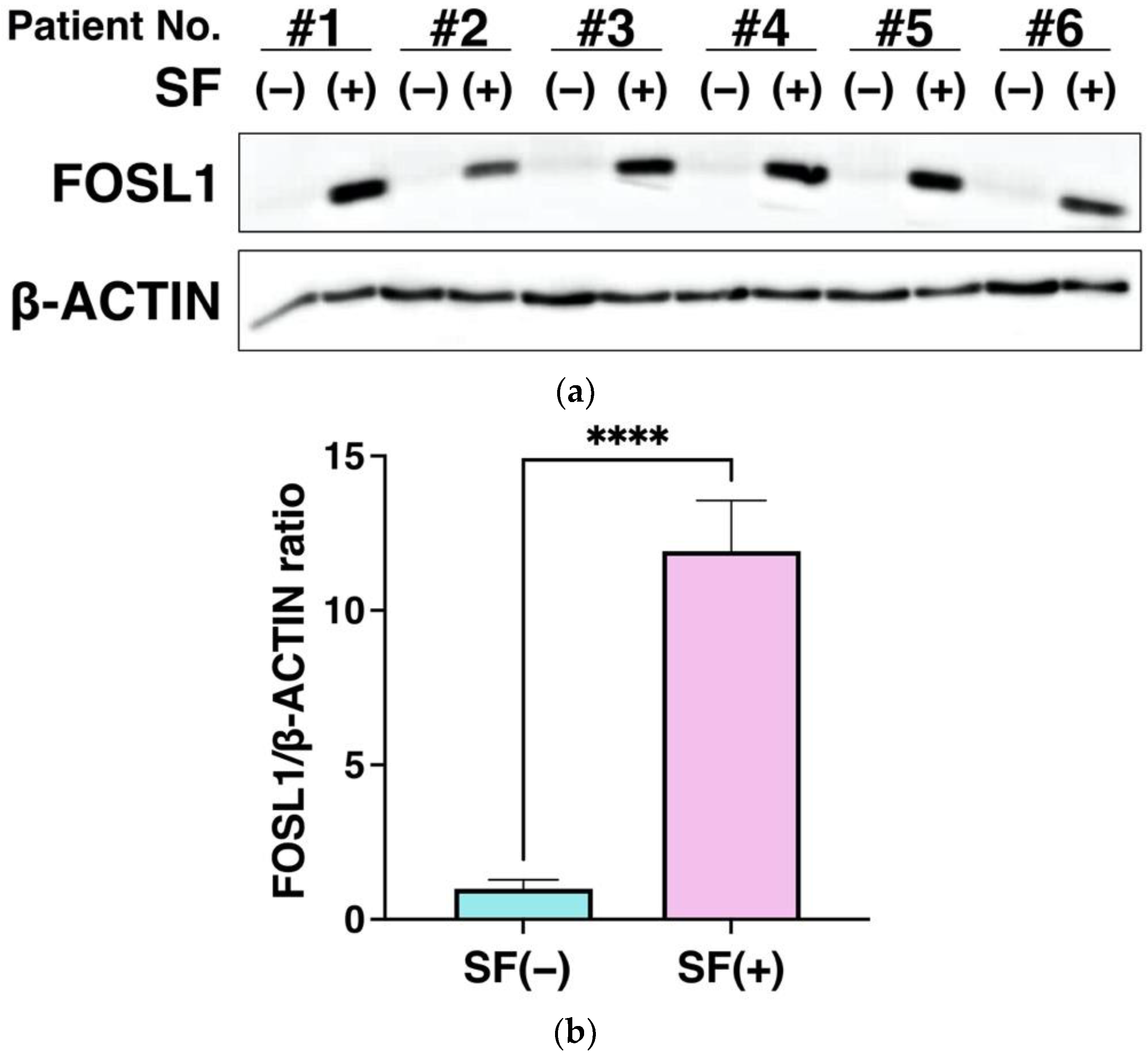
3.6. FOSL1 Protein Expression Using Western Blot Analysis
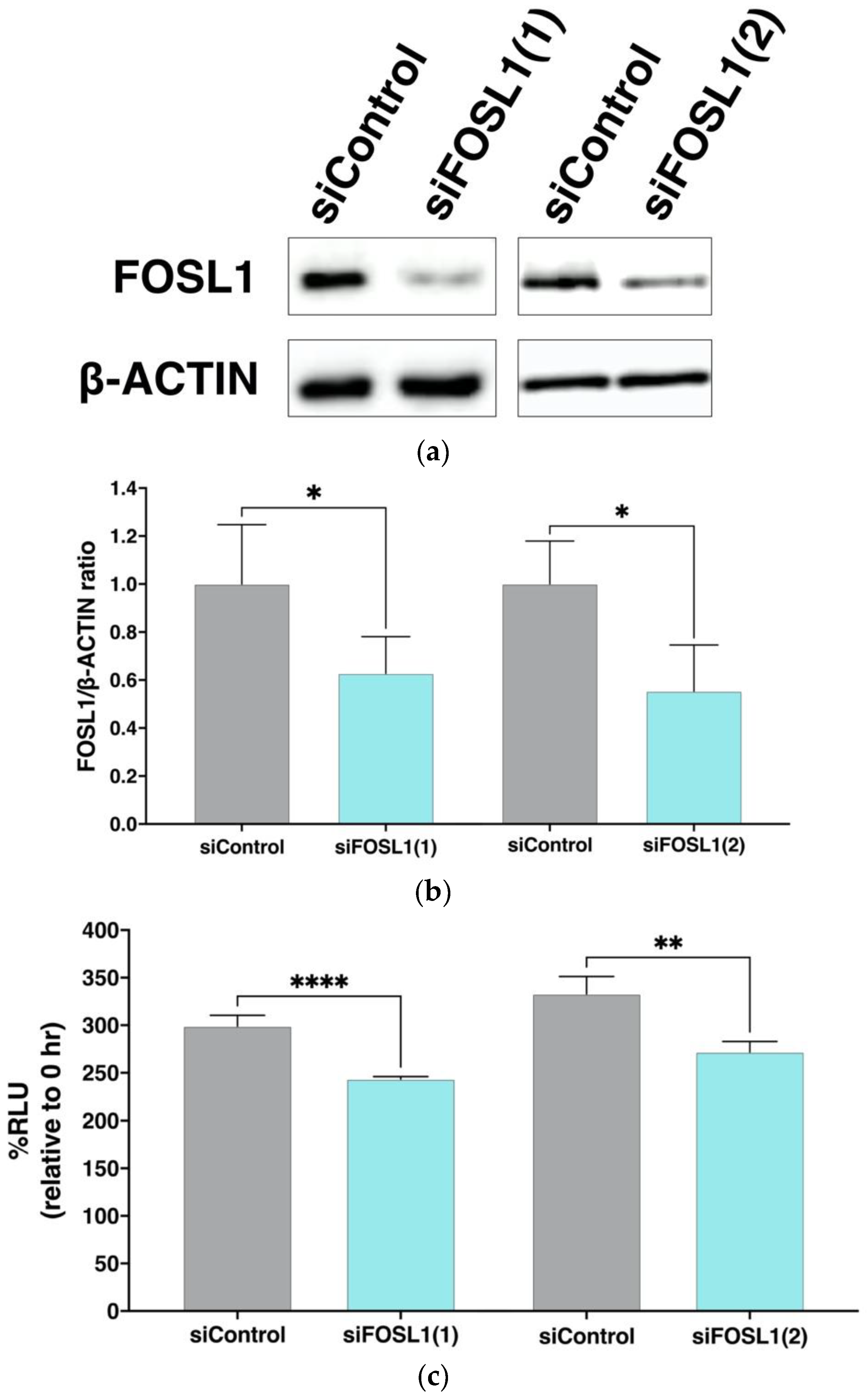
3.7. Effect of FOSL1 Knockdown on Viable Cell Rate of ADSCs
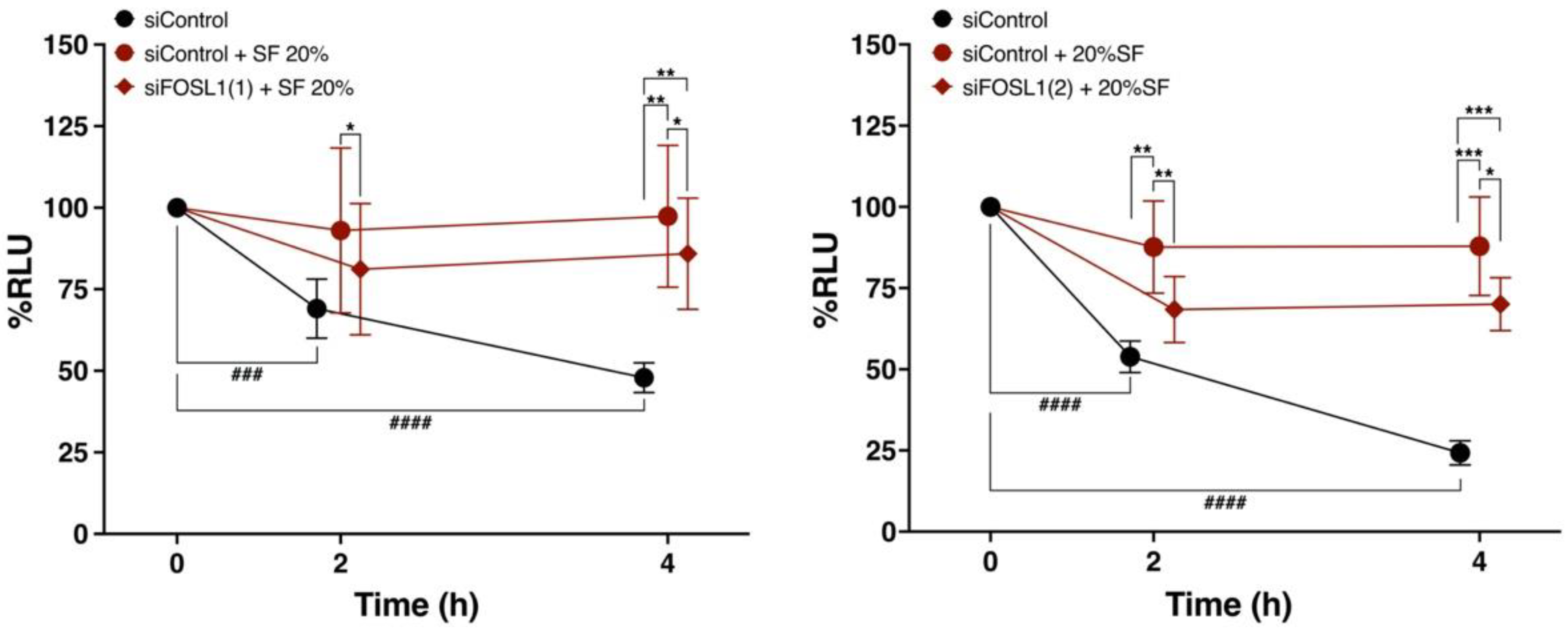
3.8. FOSL1 Is Involved in the Viability of ADSCs Treated with SF
4. Discussion
4.1. SF Is Involved in the Maintenance of ADSC Viability
4.2. FOSL1 Is Involved in SF-Induced Increase in ADSC Viability
4.3. Involvement of FOSL1 in the Immunosuppressive Effects of ADSCs
4.4. Relationship between SF Components and Elevated FOSL1 Expression in ADSCs
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zuk, P.A.; Zhu, M.; Ashjian, P.; De Ugarte, D.A.; Huang, J.I.; Mizuno, H.; Alfonso, Z.C.; Fraser, J.K.; Benhaim, P.; Hedrick, M.H. Human Adipose Tissue Is a Source of Multipotent Stem Cells. Mol. Biol. Cell 2002, 13, 4279–4295. [Google Scholar] [CrossRef]
- Zuk, P.A.; Zhu, M.; Mizuno, H.; Huang, J.; Futrell, J.W.; Katz, A.J.; Benhaim, P.; Lorenz, H.P.; Hedrick, M.H. Multilineage Cells from Human Adipose Tissue: Implications for Cell-Based Therapies. Tissue Eng. 2001, 7, 211–228. [Google Scholar] [CrossRef] [Green Version]
- Furuhashi, K.; Tsuboi, N.; Shimizu, A.; Katsuno, T.; Kim, H.; Saka, Y.; Ozaki, T.; Sado, Y.; Imai, E.; Matsuo, S.; et al. Serum-Starved Adipose-Derived Stromal Cells Ameliorate Crescentic GN by Promoting Immunoregulatory Macrophages. J. Am. Soc. Nephrol. 2013, 24, 587–603. [Google Scholar] [CrossRef] [Green Version]
- Caplan, A.I.; Correa, D. The MSC: An Injury Drugstore. Cell Stem Cell 2011, 9, 11–15. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Bai, X.; Shen, K.; Luo, L.; Zhao, M.; Xu, C.; Jia, Y.; Xiao, D.; Li, Y.; Gao, X.; et al. Exosomes Derived from Adipose Mesenchymal Stem Cells Promote Diabetic Chronic Wound Healing through SIRT3/SOD2. Cells 2022, 11, 2568. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, N.; Mak, C.; Bojanic, C.; To, K.; Khan, W. Meta-analysis of adipose tissue derived cell-based therapy for the treatment of knee osteoarthritis. Cells 2021, 10, 1365. [Google Scholar] [CrossRef] [PubMed]
- Delanois, R.E.; Sax, O.C.; Chen, Z.; Cohen, J.M.; Callahan, D.M.; Mont, M.A. Biologic Therapies for the Treatment of Knee Osteoarthritis: An Updated Systematic Review. J. Arthroplast. 2022, 37, 2480–2506. [Google Scholar] [CrossRef]
- Delanois, R.E.; Etcheson, J.I.; Sodhi, N.; Henn, R.F.; Gwam, C.U.; George, N.E.; Mont, M.A. Biologic Therapies for the Treatment of Knee Osteoarthritis. J. Arthroplast. 2019, 34, 801–813. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Chae, D.-S.; Song, B.-W.; Lim, S.; Kim, S.W.; Kim, I.-K.; Hwang, K.-C. ADSC-Based Cell Therapies for Musculoskeletal Disorders: A Review of Recent Clinical Trials. Int. J. Mol. Sci. 2021, 22, 10586. [Google Scholar] [CrossRef]
- Chang, T.-H.; Wu, C.-S.; Chiou, S.-H.; Chang, C.-H.; Liao, H.-J. Adipose-Derived Stem Cell Exosomes as a Novel Anti-Inflammatory Agent and the Current Therapeutic Targets for Rheumatoid Arthritis. Biomedicines 2022, 10, 1725. [Google Scholar] [CrossRef]
- Barakat, B.; Franke, K.; Schakaki, S.; Hijazi, S.; Hasselhof, V.; Vögeli, T.-A. Stem cell applications in regenerative medicine for stress urinary incontinence: A review of effectiveness based on clinical trials. Arab J. Urol. 2020, 18, 194–205. [Google Scholar] [CrossRef] [Green Version]
- Rosa, I.; Romano, E.; Fioretto, B.S.; Matucci-Cerinic, M.; Manetti, M. Adipose-derived stem cells: Pathophysiologic implications vs therapeutic potential in systemic sclerosis. World J. Stem Cells 2021, 13, 30–48. [Google Scholar] [CrossRef] [PubMed]
- Holm, J.S.; Toyserkani, N.M.; Sorensen, J.A. Adipose-derived stem cells for treatment of chronic ulcers: Current status. Stem Cell Res. Ther. 2018, 9, 142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spasovski, D.; Spasovski, V.; Baščarević, Z.; Stojiljković, M.; Vreća, M.; Anđelković, M.; Pavlović, S. Intra-articular injection of autologous adipose-derived mesenchymal stem cells in the treatment of knee osteoarthritis. J. Gene Med. 2018, 20, e3002. [Google Scholar] [CrossRef] [PubMed]
- Jo, C.H.; Lee, Y.G.; Shin, W.H.; Kim, H.; Chai, J.W.; Jeong, E.C.; Kim, J.E.; Shim, H.; Shin, J.S.; Shin, I.S.; et al. Intra-articular injection of mesenchymal stem cells for the treatment of osteoarthritis of the knee: A proof-of-concept clinical trial. Stem Cells 2014, 32, 1254–1266. [Google Scholar] [CrossRef]
- Higuchi, J.; Yamagami, R.; Matsumoto, T.; Terao, T.; Inoue, K.; Tsuji, S.; Maenohara, Y.; Matsuzaki, T.; Chijimatsu, R.; Omata, Y.; et al. Associations of clinical outcomes and MRI findings in intra-articular administration of autologous adipose-derived stem cells for knee osteoarthritis. Regen. Ther. 2020, 14, 332–340. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Jorgensen, C. Mesenchymal Stromal Cells in Osteoarthritis: Evidence for Structural Benefit and Cartilage Repair. Biomedicines 2022, 10, 1278. [Google Scholar] [CrossRef]
- Ścieżyńska, A.; Soszyńska, M.; Szpak, P.; Krześniak, N.; Malejczyk, J.; Kalaszczyńska, I. Influence of hypothermic storage fluids on mesenchymal stem cell stability: A comprehensive review and personal experience. Cells 2021, 10, 1043. [Google Scholar] [CrossRef]
- Moll, G.; Alm, J.J.; Davies, L.C.; von Bahr, L.; Heldring, N.; Stenbeck-Funke, L.; Hamad, O.A.; Hinsch, R.; Ignatowicz, L.; Locke, M.; et al. Do Cryopreserved Mesenchymal Stromal Cells Display Impaired Immunomodulatory and Therapeutic Properties? Stem Cells 2014, 32, 2430–2442. [Google Scholar] [CrossRef] [Green Version]
- Freitag, J.; Bates, D.; Wickham, J.; Shah, K.; Huguenin, L.; Tenen, A.; Paterson, K.; Boyd, R. Adipose-derived mesenchymal stem cell therapy in the treatment of knee osteoarthritis: A randomized controlled trial. Regen. Med. 2019, 14, 213–230. [Google Scholar] [CrossRef]
- Lee, W.S.; Kim, H.J.; Kim, K.I.; Kim, G.B.; Jin, W. Intra-Articular Injection of Autologous Adipose Tissue-Derived Mesenchymal Stem Cells for the Treatment of Knee Osteoarthritis: A Phase IIb, Randomized, Placebo-Controlled Clinical Trial. Stem Cells Transl. Med. 2019, 8, 504–511. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.-H.; Kim, K.-I.; Yoon, W.K.; Song, S.-J.; Jin, W. Intra-articular Injection of Mesenchymal Stem Cells After High Tibial Osteotomy in Osteoarthritic Knee: Two-Year Follow-up of Randomized Control Trial. Stem Cells Transl. Med. 2022, 11, 572–585. [Google Scholar] [CrossRef]
- Kim, K.-I.; Lee, W.-S.; Kim, J.-H.; Bae, J.-K.; Jin, W. Safety and Efficacy of the Intra-articular Injection of Mesenchymal Stem Cells for the Treatment of Osteoarthritic Knee: A 5-Year Follow-up Study. Stem Cells Transl. Med. 2022, 11, 586–596. [Google Scholar] [CrossRef]
- Nasb, M.; Liangjiang, H.; Gong, C.; Hong, C. Human adipose-derived Mesenchymal stem cells, low-intensity pulsed ultrasound, or their combination for the treatment of knee osteoarthritis: Study protocol for a first-in-man randomized controlled trial. BMC Musculoskelet. Disord. 2020, 21, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jo, C.H.; Chai, J.W.; Jeong, E.C.; Oh, S.; Shin, J.S.; Shim, H.; Yoon, K.S. Intra-articular Injection of Mesenchymal Stem Cells for the Treatment of Osteoarthritis of the Knee: A 2-Year Follow-up Study. Am. J. Sports Med. 2017, 45, 2774–2783. [Google Scholar] [CrossRef]
- Freitag, J.; Shah, K.; Wickham, J.; Boyd, R.; Tenen, A. The effect of autologous adipose derived mesenchymal stem cell therapy in the treatment of a large osteochondral defect of the knee following unsuccessful surgical intervention of osteochondritis dissecans—A case study. BMC Musculoskelet. Disord. 2017, 18, 298. [Google Scholar] [CrossRef] [PubMed]
- Kuwasawa, A.; Okazaki, K.; Noda, K.; Fukushima, T.; Nihei, K. Intra-articular injection of culture-expanded adipose tissue-derived stem cells for knee osteoarthritis: Assessments with clinical symptoms and quantitative measurements of articular cartilage volume. J. Orthop. Sci. 2022. [Google Scholar] [CrossRef] [PubMed]
- Carvalho Schweich-Adami, L.; da Silva, R.A.; Menezes, J.N.d.S.; Baranoski, A.; Kassuya, C.A.L.; Bernardi, L.; Juliano Oliveira, R.; Conceição Milan Brochado Antoniolli-Silva, A. The intra-articular injection of adipose-derived stem cells decreases pain and reduces inflammation in knee osteoarthritis, with or without the addition of platelet-rich plasma also improves functionality. J. Tissue Eng. Regen. Med. 2022, 16, 900–912. [Google Scholar] [CrossRef]
- Sadri, B.; Tamimi, A.; Nouraein, S.; Bagheri Fard, A.; Mohammadi, J.; Mohammadpour, M.; Hassanzadeh, M.; Bajouri, A.; Madani, H.; Barekat, M.; et al. Clinical and laboratory findings following transplantation of allogeneic adipose-derived mesenchymal stromal cells in knee osteoarthritis, a brief report. Connect Tissue Res. 2022, 63, 663–674. [Google Scholar] [CrossRef] [PubMed]
- Pers, Y.-M.; Rackwitz, L.; Ferreira, R.; Pullig, O.; Delfour, C.; Barry, F.; Sensebe, L.; Casteilla, L.; Fleury, S.; Bourin, P.; et al. Adipose Mesenchymal Stromal Cell-Based Therapy for Severe Osteoarthritis of the Knee: A Phase I Dose-Escalation Trial. Stem Cells Transl. Med. 2016, 5, 847–856. [Google Scholar] [CrossRef]
- Yokota, N.; Lyman, S.; Hanai, H.; Shimomura, K.; Ando, W.; Nakamura, N. Clinical Safety and Effectiveness of Adipose-Derived Stromal Cell vs Stromal Vascular Fraction Injection for Treatment of Knee Osteoarthritis: 2-Year Results of Parallel Single-Arm Trials. Am. J. Sports Med. 2022, 50, 2659–2668. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Du, H.; Dai, C.; Zhang, L.; Li, S.; Hunter, D.J.; Lu, L.; Bao, C. Human adipose-derived mesenchymal stem cells for osteoarthritis: A pilot study with long-term follow-up and repeated injections. Regen. Med. 2018, 13, 295–307. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Ruan, J.; Tang, H.; Li, J.; Shi, Y.; Li, M.; Li, S.; Xu, C.; Lu, Q.; Dai, C. Multi-compositional MRI evaluation of repair cartilage in knee osteoarthritis with treatment of allogeneic human adipose-derived mesenchymal progenitor cells. Stem Cell Res. Ther. 2019, 10, 308. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.-H.; Chen, Y.-C.; Yu, S.-N.; Lai, W.-L.; Shen, Y.-S.; Shen, P.-C.; Lin, S.-H.; Chang, C.-H.; Lee, S.-M. Infrapatellar fat pad-derived mesenchymal stromal cell product for treatment of knee osteoarthritis: A first-in-human study with evaluation of the potency marker. Cytotherapy 2022, 24, 72–85. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Dai, C.; Zhang, Z.; Du, H.; Li, S.; Ye, P.; Fu, Q.; Zhang, L.; Wu, X.; Dong, Y.; et al. Treatment of knee osteoarthritis with intra-articular injection of autologous adipose-derived mesenchymal progenitor cells: A prospective, randomized, double-blind, active-controlled, phase IIb clinical trial. Stem Cell Res. Ther. 2019, 10, 143. [Google Scholar] [CrossRef] [Green Version]
- Qiao, Z.; Tang, J.; Yue, B.; Wang, J.; Zhang, J.; Xuan, L.; Dai, C.; Li, S.; Li, M.; Xu, C.; et al. Human adipose-derived mesenchymal progenitor cells plus microfracture and hyaluronic acid for cartilage repair: A Phase IIa trial. Regen. Med. 2020, 15, 1193–1214. [Google Scholar] [CrossRef] [Green Version]
- Lu, L.; Dai, C.; Du, H.; Li, S.; Ye, P.; Zhang, L.; Wang, X.; Song, Y.; Togashi, R.; Vangsness, C.T.; et al. Intra-articular injections of allogeneic human adipose-derived mesenchymal progenitor cells in patients with symptomatic bilateral knee osteoarthritis: A Phase I pilot study. Regen. Med. 2020, 15, 1625–1636. [Google Scholar] [CrossRef] [PubMed]
- Freitag, J.; Norsworthy, C.; Wickham, J.; Shah, K.; Tenen, A. High tibial osteotomy in combination with arthroscopic abrasion arthroplasty and autologous adipose-derived mesenchymal stem cell therapy in the treatment of advanced knee osteoarthritis. BMJ Case Rep. 2019, 12, bcr-2018-228003. [Google Scholar] [CrossRef] [Green Version]
- Freitag, J.; Wickham, J.; Shah, K.; Li, D.; Norsworthy, C.; Tenen, A. Mesenchymal stem cell therapy combined with arthroscopic abrasion arthroplasty regenerates cartilage in patients with severe knee osteoarthritis: A case series. Regen. Med. 2020, 15, 1957–1977. [Google Scholar] [CrossRef]
- Freitag, J.; Wickham, J.; Shah, K.; Tenen, A. Real-world evidence of mesenchymal stem cell therapy in knee osteoarthritis: A large prospective two-year case series. Regen. Med. 2022, 17, 355–373. [Google Scholar] [CrossRef]
- Chen, C.-F.; Hu, C.-C.; Wu, C.-T.; Wu, H.-T.H.; Chang, C.-S.; Hung, Y.-P.; Tsai, C.-C.; Chang, Y. Treatment of knee osteoarthritis with intra-articular injection of allogeneic adipose-derived stem cells (ADSCs) ELIXCYTE®: A phase I/II, randomized, active-control, single-blind, multiple-center clinical trial. Stem Cell Res. Ther. 2021, 12, 562. [Google Scholar] [CrossRef] [PubMed]
- Kuah, D.; Sivell, S.; Longworth, T.; James, K.; Guermazi, A.; Cicuttini, F.; Wang, Y.; Craig, S.; Comin, G.; Robinson, D.; et al. Safety, tolerability and efficacy of intra-articular Progenza in knee osteoarthritis: A randomized double-blind placebo-controlled single ascending dose study. J. Transl. Med. 2018, 16, 49. [Google Scholar] [CrossRef] [Green Version]
- Yokota, N.; Hattori, M.; Ohtsuru, T.; Otsuji, M.; Lyman, S.; Shimomura, K.; Nakamura, N. Comparative Clinical Outcomes After Intra-articular Injection With Adipose-Derived Cultured Stem Cells or Noncultured Stromal Vascular Fraction for the Treatment of Knee Osteoarthritis. Am. J. Sports Med. 2019, 47, 2577–2583. [Google Scholar] [CrossRef] [PubMed]
- Alp, E.; Gonen, Z.B.; Gundogan, K.; Esmaoglu, A.; Kaynar, L.; Cetin, A.; Karakukcu, M.; Cetin, M.; Kalin, G.; Doganay, M. The Effect of Mesenchymal Stromal Cells on the Mortality of Patients with Sepsis and Septic Shock: A Promising Therapy. Emerg. Med. Int. 2022, 2022, 1–9. [Google Scholar] [CrossRef]
- Vij, R.; Stebbings, K.A.; Kim, H.; Park, H.; Chang, D. Safety and efficacy of autologous, adipose-derived mesenchymal stem cells in patients with rheumatoid arthritis: A phase I/IIa, open-label, non-randomized pilot trial. Stem Cell Res. Ther. 2022, 13, 88. [Google Scholar] [CrossRef] [PubMed]
- Alió del Barrio, J.L.; El Zarif, M.; de Miguel, M.P.; Azaar, A.; Makdissy, N.; Harb, W.; El Achkar, I.; Arnalich-Montiel, F.; Alió, J.L. Cellular Therapy With Human Autologous Adipose-Derived Adult Stem Cells for Advanced Keratoconus. Cornea 2017, 36, 952–960. [Google Scholar] [CrossRef]
- Oner, A.; Gonen, Z.B.; Sinim, N.; Cetin, M.; Ozkul, Y. Subretinal adipose tissue-derived mesenchymal stem cell implantation in advanced stage retinitis pigmentosa: A phase I clinical safety study. Stem Cell Res. Ther. 2016, 7, 178. [Google Scholar] [CrossRef] [Green Version]
- Zheng, G.; Huang, L.; Tong, H.; Shu, Q.; Hu, Y.; Ge, M.; Deng, K.; Zhang, L.; Zou, B.; Cheng, B.; et al. Treatment of acute respiratory distress syndrome with allogeneic adipose-derived mesenchymal stem cells: A randomized, placebo-controlled pilot study. Respir. Res. 2014, 15, 39. [Google Scholar] [CrossRef] [Green Version]
- Tzouvelekis, A.; Paspaliaris, V.; Koliakos, G.; Ntolios, P.; Bouros, E.; Oikonomou, A.; Zissimopoulos, A.; Boussios, N.; Dardzinski, B.; Gritzalis, D.; et al. A prospective, non-randomized, no placebo-controlled, phase Ib clinical trial to study the safety of the adipose derived stromal cells-stromal vascular fraction in idiopathic pulmonary fibrosis. J. Transl. Med. 2013, 11, 171. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Long, X.; Si, L.; Chen, B.; Zhang, Y.; Sun, T.; Zhang, X.; Zhao, R.C.; Wang, X. A Pilot Study On Ex Vivo Expanded Autologous Adipose-Derived Stem Cells of Improving Fat Retention in Localized Scleroderma Patients. Stem Cells Transl. Med. 2021, 10, 1148–1156. [Google Scholar] [CrossRef]
- Huang, K.-C.; Chuang, M.-H.; Lin, Z.-S.; Lin, Y.-C.; Chen, C.-H.; Chang, C.-L.; Huang, P.-C.; Syu, W.-S.; Chiou, T.-W.; Hong, Z.-H.; et al. Transplantation with GXHPC1 for Liver Cirrhosis: Phase 1 Trial. Cell Transpl. 2019, 28, 100S–111S. [Google Scholar] [CrossRef] [PubMed]
- Freitag, J.; Ford, J.; Bates, D.; Boyd, R.; Hahne, A.; Wang, Y.; Cicuttini, F.; Huguenin, L.; Norsworthy, C.; Shah, K. Adipose derived mesenchymal stem cell therapy in the treatment of isolated knee chondral lesions: Design of a randomised controlled pilot study comparing arthroscopic microfracture versus arthroscopic microfracture combined with postoperative mesenchymal. BMJ Open 2015, 5, e009332. [Google Scholar] [CrossRef] [Green Version]
- Toma, C.; Wagner, W.R.; Bowry, S.; Schwartz, A.; Villanueva, F. Fate Of Culture-Expanded Mesenchymal Stem Cells in The Microvasculature. Circ. Res. 2009, 104, 398–402. [Google Scholar] [CrossRef] [Green Version]
- Ko, J.-Y.; Lee, J.; Lee, J.; Ryu, Y.H.; Im, G.-I. SOX-6, 9-Transfected Adipose Stem Cells to Treat Surgically-Induced Osteoarthritis in Goats. Tissue Eng. Part A 2019, 25, 990–1000. [Google Scholar] [CrossRef]
- Toupet, K.; Maumus, M.; Luz-Crawford, P.; Lombardo, E.; Lopez-Belmonte, J.; van Lent, P.; Garin, M.I.; van den Berg, W.; Dalemans, W.; Jorgensen, C.; et al. Survival and Biodistribution of Xenogenic Adipose Mesenchymal Stem Cells Is Not Affected by the Degree of Inflammation in Arthritis. PLoS ONE 2015, 10, e0114962. [Google Scholar] [CrossRef] [Green Version]
- Toupet, K.; Maumus, M.; Peyrafitte, J.A.; Bourin, P.; Van Lent, P.L.E.M.; Ferreira, R.; Orsetti, B.; Pirot, N.; Casteilla, L.; Jorgensen, C.; et al. Long-term detection of human adipose-derived mesenchymal stem cells after intraarticular injection in SCID mice. Arthritis Rheum. 2013, 65, 1786–1794. [Google Scholar] [CrossRef] [PubMed]
- Yamada, Y.; Okano, T.; Orita, K.; Makino, T.; Shima, F.; Nakamura, H. 3D-cultured small size adipose-derived stem cell spheroids promote bone regeneration in the critical-sized bone defect rat model. Biochem. Biophys. Res. Commun. 2022, 603, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Brindo da Cruz, I.C.; Velosa, A.P.P.; Carrasco, S.; dos Santos Filho, A.; Tomaz de Miranda, J.; Pompeu, E.; Fernandes, T.L.; Bueno, D.F.; Fanelli, C.; Goldenstein-Schainberg, C.; et al. Post-Adipose-Derived Stem Cells (ADSC) Stimulated by Collagen Type V (Col V) Mitigate the Progression of Osteoarthritic Rabbit Articular Cartilage. Front. Cell Dev. Biol. 2021, 9, 606890. [Google Scholar] [CrossRef]
- Koellensperger, E.; Bollinger, N.; Dexheimer, V.; Gramley, F.; Germann, G.; Leimer, U. Choosing the right type of serum for different applications of human adipose tissue–derived stem cells: Influence on proliferation and differentiation abilities. Cytotherapy 2014, 16, 789–799. [Google Scholar] [CrossRef]
- Horcharoensuk, P.; Yang-en, S.; Narkwichean, A.; Rungsiwiwut, R. Proline-based solution maintains cell viability and stemness of canine adipose-derived mesenchymal stem cells after hypothermic storage. PLoS ONE 2022, 17, e0264773. [Google Scholar] [CrossRef]
- Ko, J.; Park, J.; Kim, J.; Im, G. Characterization of adipose-derived stromal/stem cell spheroids versus single-cell suspension in cell survival and arrest of osteoarthritis progression. J. Biomed. Mater. Res. Part A 2021, 109, 869–878. [Google Scholar] [CrossRef]
- Mancuso, P.; Raman, S.; Glynn, A.; Barry, F.; Murphy, J.M. Mesenchymal Stem Cell Therapy for Osteoarthritis: The Critical Role of the Cell Secretome. Front. Bioeng. Biotechnol. 2019, 7, 9. [Google Scholar] [CrossRef] [Green Version]
- Manferdini, C.; Paolella, F.; Gabusi, E.; Cattini, L.; Rojewski, M.; Schrezenmeier, H.; Addimanda, O.; Meliconi, R.; Lisignoli, G. Osteoarthritic Milieu Affects Adipose-Derived Mesenchymal Stromal Cells. J. Orthop. Res. 2020, 38, 336–347. [Google Scholar] [CrossRef] [Green Version]
- Martin-Alarcon, L.; Schmidt, T.A. Rheological effects of macromolecular interactions in synovial fluid. Biorheology 2016, 53, 49–67. [Google Scholar] [CrossRef]
- Belcher, C.; Yaqub, R.; Fawthrop, F.; Bayliss, M.; Doherty, M. Synovial fluid chondroitin and keratan sulphate epitopes, glycosaminoglycans, and hyaluronan in arthritic and normal knees. Ann. Rheum. Dis. 1997, 56, 299–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heilmann, H.-H.; Lindenhayn, K.; Walther, H.-U. Das Synovia-Volumen gesunder und arthrotischer menschlicher Kniegelenke*. Z. Orthop. Ihre Grenzgeb. 1996, 134, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Nees, T.A.; Rosshirt, N.; Zhang, J.A.; Reiner, T.; Sorbi, R.; Tripel, E.; Walker, T.; Schiltenwolf, M.; Hagmann, S.; Moradi, B. Synovial Cytokines Significantly Correlate with Osteoarthritis-Related Knee Pain and Disability: Inflammatory mediators of potential clinical relevance. J. Clin. Med. 2019, 8, 1343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sayegh, S.; El Atat, O.; Diallo, K.; Rauwel, B.; Degboé, Y.; Cavaignac, E.; Constantin, A.; Cantagrel, A.; Trak-Smayra, V.; Alaaeddine, N.; et al. Rheumatoid Synovial Fluids Regulate the Immunomodulatory Potential of Adipose-Derived Mesenchymal Stem Cells Through a TNF/NF-κB-Dependent Mechanism. Front. Immunol. 2019, 10, 1482. [Google Scholar] [CrossRef]
- Kiefer, K.M.; O’Brien, T.D.; Pluhar, E.G.; Conzemius, M. Canine adipose-derived stromal cell viability following exposure to synovial fluid from osteoarthritic joints. Vet. Rec. Open 2015, 2, e000063. [Google Scholar] [CrossRef] [Green Version]
- Fuku, A.; Taki, Y.; Nakamura, Y.; Kitajima, H.; Takaki, T.; Koya, T.; Tanida, I.; Nozaki, K.; Sunami, H.; Hirata, H.; et al. Evaluation of the Usefulness of Human Adipose-Derived Stem Cell Spheroids Formed Using SphereRing® and the Lethal Damage Sensitivity to Synovial Fluid In Vitro. Cells 2022, 11, 337. [Google Scholar] [CrossRef]
- Pecce, V.; Verrienti, A.; Fiscon, G.; Sponziello, M.; Conte, F.; Abballe, L.; Durante, C.; Farina, L.; Filetti, S.; Paci, P. The role of FOSL1 in stem-like cell reprogramming processes. Sci. Rep. 2021, 11, 14677. [Google Scholar] [CrossRef]
- Schwabe, K.; Garcia, M.; Ubieta, K.; Hannemann, N.; Herbort, B.; Luther, J.; Noël, D.; Jorgensen, C.; Casteilla, L.; David, J.P.; et al. Inhibition of Osteoarthritis by Adipose-Derived Stromal Cells Overexpressing Fra-1 in Mice. Arthritis Rheumatol. 2016, 68, 138–151. [Google Scholar] [CrossRef] [Green Version]
- Kellgren, J.H.; Lawrence, J.S. Radiological Assessment of Osteo-Arthrosis. Ann. Rheum. Dis. 1957, 16, 494–502. [Google Scholar] [CrossRef] [Green Version]
- Leijs, M.J.C.; van Buul, G.M.; Lubberts, E.; Bos, P.K.; Verhaar, J.A.N.; Hoogduijn, M.J.; van Osch, G.J.V.M. Effect of Arthritic Synovial Fluids on the Expression of Immunomodulatory Factors by Mesenchymal Stem Cells: An Explorative in vitro Study. Front. Immunol. 2012, 3, 231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.S.; Deans, R.J.; Keating, A.; Prockop, D.J.; Horwitz, E.M. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Taki, Y.; Fuku, A.; Nakamura, Y.; Koya, T.; Kitajima, H.; Tanida, I.; Takaki, T.; Nozaki, K.; Sunami, H.; Hirata, H.; et al. A morphological study of adipose-derived stem cell sheets created with temperature-responsive culture dishes using scanning electron microscopy. Med. Mol. Morphol. 2022, 55, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Pearson, M.J.; Herndler-Brandstetter, D.; Tariq, M.A.; Nicholson, T.A.; Philp, A.M.; Smith, H.L.; Davis, E.T.; Jones, S.W.; Lord, J.M. IL-6 secretion in osteoarthritis patients is mediated by chondrocyte-synovial fibroblast cross-talk and is enhanced by obesity. Sci. Rep. 2017, 7, 3451. [Google Scholar] [CrossRef] [Green Version]
- Nanus, D.E.; Wijesinghe, S.N.; Pearson, M.J.; Hadjicharalambous, M.R.; Rosser, A.; Davis, E.T.; Lindsay, M.A.; Jones, S.W. Regulation of the Inflammatory Synovial Fibroblast Phenotype by Metastasis-Associated Lung Adenocarcinoma Transcript 1 Long Noncoding RNA in Obese Patients With Osteoarthritis. Arthritis Rheumatol. 2020, 72, 609–619. [Google Scholar] [CrossRef]
- Ryu, J.-H.; Yang, S.; Shin, Y.; Rhee, J.; Chun, C.-H.; Chun, J.-S. Interleukin-6 plays an essential role in hypoxia-inducible factor 2α-induced experimental osteoarthritic cartilage destruction in mice. Arthritis Rheum. 2011, 63, 2732–2743. [Google Scholar] [CrossRef]
- Wu, Y.; Li, M.; Liao, X.; Li, S.; Yan, J.; Fan, L.; She, W.; Song, J.; Liu, H. Effects of storage culture media, temperature and duration on human adipose derived stem cell viability for clinical use. Mol. Med. Rep. 2019, 19, 2189–2201. [Google Scholar] [CrossRef] [Green Version]
- Tran, T.D.X.; Wu, C.-M.; Dubey, N.K.; Deng, Y.-H.; Su, C.-W.; Pham, T.T.; Thi Le, P.B.; Sestili, P.; Deng, W.-P. Time- and Kellgren–Lawrence Grade-Dependent Changes in Intra-Articularly Transplanted Stromal Vascular Fraction in Osteoarthritic Patients. Cells 2019, 8, 308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.P.C.; Cheng, C.-H.; Hsu, C.-C.; Lin, H.-C.; Tsai, Y.-R.; Chen, J.-L. The influence of platelet rich plasma on synovial fluid volumes, protein concentrations, and severity of pain in patients with knee osteoarthritis. Exp. Gerontol. 2017, 93, 68–72. [Google Scholar] [CrossRef]
- Luther, J.; Driessler, F.; Megges, M.; Hess, A.; Herbort, B.; Mandic, V.; Zaiss, M.M.; Reichardt, A.; Zech, C.; Tuckermann, J.P.; et al. Elevated Fra-1 expression causes severe lipodystrophy. J. Cell Sci. 2011, 124, 1465–1476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jochum, W.; David, J.-P.; Elliott, C.; Wutz, A.; Plenk, H.; Matsuo, K.; Wagner, E.F. Increased bone formation and osteosclerosis in mice overexpressing the transcription factor Fra-1. Nat. Med. 2000, 6, 980–984. [Google Scholar] [CrossRef]
- Sobolev, V.V.; Khashukoeva, A.Z.; Evina, O.E.; Geppe, N.A.; Chebysheva, S.N.; Korsunskaya, I.M.; Tchepourina, E.; Mezentsev, A. Role of the Transcription Factor FOSL1 in Organ Development and Tumorigenesis. Int. J. Mol. Sci. 2022, 23, 1521. [Google Scholar] [CrossRef]
- Casalino, L.; Talotta, F.; Cimmino, A.; Verde, P. The Fra-1/AP-1 Oncoprotein: From the “Undruggable” Transcription Factor to Therapeutic Targeting. Cancers 2022, 14, 1480. [Google Scholar] [CrossRef]
- Zhou, Z.; Liu, Q.; Zhang, G.; Mohammed, D.; Amadou, S.; Tan, G.; Zhang, X. HOXA11-AS1 Promotes PD-L1-Mediated Immune Escape and Metastasis of Hypopharyngeal Carcinoma by Facilitating PTBP1 and FOSL1 Association. Cancers 2022, 14, 3694. [Google Scholar] [CrossRef]
- Diego-González, L.; Fernández-Carrera, A.; Igea, A.; Martínez-Pérez, A.; Real Oliveira, M.E.C.D.; Gomes, A.C.; Guerra, C.; Barbacid, M.; González-Fernández, Á.; Simón-Vázquez, R. Combined Inhibition of FOSL-1 and YAP Using siRNA-Lipoplexes Reduces the Growth of Pancreatic Tumor. Cancers 2022, 14, 3102. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; King, P.; Liang, E.; Guo, A.A.; Liu, M. LncRNA HOTAIR sponges miR-301a-3p to promote glioblastoma proliferation and invasion through upregulating FOSL1. Cell. Signal. 2022, 94, 110306. [Google Scholar] [CrossRef]
- Liu, H.; Ren, G.; Wang, T.; Chen, Y.; Gong, C.; Bai, Y.; Wang, B.; Qi, H.; Shen, J.; Zhu, L.; et al. Aberrantly expressed Fra-1 by IL-6/STAT3 transactivation promotes colorectal cancer aggressiveness through epithelial–mesenchymal transition. Carcinogenesis 2015, 36, 459–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taha, S.; Volkmer, E.; Haas, E.; Alberton, P.; Straub, T.; David-Rus, D.; Aszodi, A.; Giunta, R.; Saller, M.M. Differences in the Inflammatory Response of White Adipose Tissue and Adipose-Derived Stem Cells. Int. J. Mol. Sci. 2020, 21, 1086. [Google Scholar] [CrossRef] [Green Version]
- Beekhuizen, M.; Gierman, L.M.; van Spil, W.E.; Van Osch, G.J.V.M.; Huizinga, T.W.J.; Saris, D.B.F.; Creemers, L.B.; Zuurmond, A.-M. An explorative study comparing levels of soluble mediators in control and osteoarthritic synovial fluid. Osteoarthr. Cartil. 2013, 21, 918–922. [Google Scholar] [CrossRef] [Green Version]
- Kawaguchi, H.; Sakamoto, T.; Koya, T.; Togi, M.; Date, I.; Watanabe, A.; Yoshida, K.; Kato, T.; Nakamura, Y.; Ishigaki, Y.; et al. Quality Verification with a Cluster−Controlled Manufacturing System to Generate Monocyte−Derived Dendritic Cells. Vaccines 2021, 9, 533. [Google Scholar] [CrossRef] [PubMed]
- Rengel, Y.; Ospelt, C.; Gay, S. Proteinases in the joint: Clinical relevance of proteinases in joint destruction. Arthritis Res. Ther. 2007, 9, 221. [Google Scholar] [CrossRef] [Green Version]
- Tchetverikov, I. MMP protein and activity levels in synovial fluid from patients with joint injury, inflammatory arthritis, and osteoarthritis. Ann. Rheum. Dis. 2005, 64, 694–698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almalki, S.G.; Agrawal, D.K. Effects of matrix metalloproteinases on the fate of mesenchymal stem cells. Stem Cell Res. Ther. 2016, 7, 129. [Google Scholar] [CrossRef] [Green Version]
- Barreto, G.; Soliymani, R.; Baumann, M.; Waris, E.; Eklund, K.K.; Zenobi-Wong, M.; Lalowski, M. Functional analysis of synovial fluid from osteoarthritic knee and carpometacarpal joints unravels different molecular profiles. Rheumatology 2019, 58, 897–907. [Google Scholar] [CrossRef]
- Moss, S.G.; Schweitzer, M.E.; Jacobson, J.A.; Brossmann, J.; Lombardi, J.V.; Dellose, S.M.; Coralnick, J.R.; Standiford, K.N.; Resnick, D. Hip joint fluid: Detection and distribution at MR imaging and US with cadaveric correlation. Radiology 1998, 208, 43–48. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient No. | #1 | #2 | #3 | #4 | #5 | #6 |
|---|---|---|---|---|---|---|
| Age | 80 | 81 | 59 | 83 | 73 | 59 |
| Sex | M | M | F | F | F | M |
| Weight (kg) | 81 | 60 | 75 | 60 | 46 | 75 |
| Height (cm) | 168 | 167 | 161 | 159 | 158 | 160 |
| K–L grading scale | 2 | 2 | 3 | 2 | 3 | 2 |
| Hyaluronic acid (μg/mL) | 74.9 | 82.3 | 97.3 | 82.4 | 103.5 | 92.2 |
| BMI (kg/m2) | 28.7 | 21.5 | 28.9 | 23.7 | 18.4 | 29.3 |
| IL-6 (pg/mL) | 1462.6 | 395.3 | 13,111.7 | 3866.9 | 798.9 | 2808.1 |
| Molecular and Cellular Functions | p-Value Range | #Molecules |
|---|---|---|
| Cellular Movement | 3.00 × 10−3–1.15 × 10−9 | ABL2, ARRDC3, CCN2, CXCL2, DKK1, DUSP1, ETS1, ETS2, FOS, FOSL1, HBEGF, LINC00312, mir-154, mir-221, mir-29, NFATC2, NFKBIA, SPRY2, DDIT4, USP38, CXCL2 |
| Cellular Development | 3.86 × 10−3–1.73 × 10−9 | ABL2, AMOTL2, ARRDC3, CCN2, CXCL2, DAGLB, DDIT4, DKK1, DUSP1, DUSP10, DUSP14, ETS1, ETS2, FOS, FOSL1, HBEGF, HEY1, LINC00312, mir-154, mir-221, mir-29, NFATC2, NOCT, NFKBIA, RGS2, SERTAD1, SPRY2, USP38, VGLL3, ZSWIM6 |
| Cellular Growth and Proliferation | 3.86 × 10−3–1.73 × 10−9 | ABL2, AMOTL2, ARRDC3, BNC1, CCN2, CXCL2, DAGLB, DKK1, DDIT4, DUSP1, DUSP10, DUSP14, ETS1, ETS2, FOS, FOSL1, HBEGF, HEY1, LINC00312, mir-154, mir-221, mir-29, NFATC2, NFKBIA, RGS2, SERTAD1, SPRY2, USP38, VGLL3, ZSWIM6 |
| Cell Death and Survival | 3.77 × 10−3–1.18 × 10−7 | ABL2, CCN2, CXCL2, DDIT4, DKK1, DUSP1, DUSP10, DUSP14, ETS1, ETS2, FOS, FOSL1, HBEGF, HEY1, mir-154, mir-221, mir-29, NFATC2, NFKBIA, SPRY2, TJP2, USP38 |
| Cell Cycle | 2.55 × 10−3–7.48 × 10−7 | CCN2, DDIT4, DKK, DUSP1, ETS1, ETS2, FOS, FOSL1, HBEGF, HEY1, mir-154, mir-221, mir-29, NFATC2, NFKBIA, NOCT, SERTAD1, SPRY2, UBC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kitajima, H.; Sakamoto, T.; Horie, T.; Kuwano, A.; Fuku, A.; Taki, Y.; Nakamura, Y.; Tanida, I.; Sunami, H.; Hirata, H.; et al. Synovial Fluid Derived from Human Knee Osteoarthritis Increases the Viability of Human Adipose-Derived Stem Cells through Upregulation of FOSL1. Cells 2023, 12, 330. https://doi.org/10.3390/cells12020330
Kitajima H, Sakamoto T, Horie T, Kuwano A, Fuku A, Taki Y, Nakamura Y, Tanida I, Sunami H, Hirata H, et al. Synovial Fluid Derived from Human Knee Osteoarthritis Increases the Viability of Human Adipose-Derived Stem Cells through Upregulation of FOSL1. Cells. 2023; 12(2):330. https://doi.org/10.3390/cells12020330
Chicago/Turabian StyleKitajima, Hironori, Takuya Sakamoto, Tetsuhiro Horie, Ayane Kuwano, Atsushi Fuku, Yasuhiko Taki, Yuka Nakamura, Ikuhiro Tanida, Hiroshi Sunami, Hiroaki Hirata, and et al. 2023. "Synovial Fluid Derived from Human Knee Osteoarthritis Increases the Viability of Human Adipose-Derived Stem Cells through Upregulation of FOSL1" Cells 12, no. 2: 330. https://doi.org/10.3390/cells12020330