
Modeling the Differentiation of Embryonic Limb Chondroprogenitors by Cell Death and Cell Senescence in High Density Micromass Cultures and Their Regulation by FGF Signaling

Abstract
:1. Introduction
2. Materials and Methods
2.1. Micromass Cultures and Treatments
2.2. Morphological and Immunohistochemical Studies
2.3. Flow Cytometry
2.4. Quantification of Culture Growth
2.5. In situ Hybridization
2.6. Epigenetic Analysis
2.7. Real-Time Quantitative PCR (qPCR) for Gene Expression Analysis
3. Results
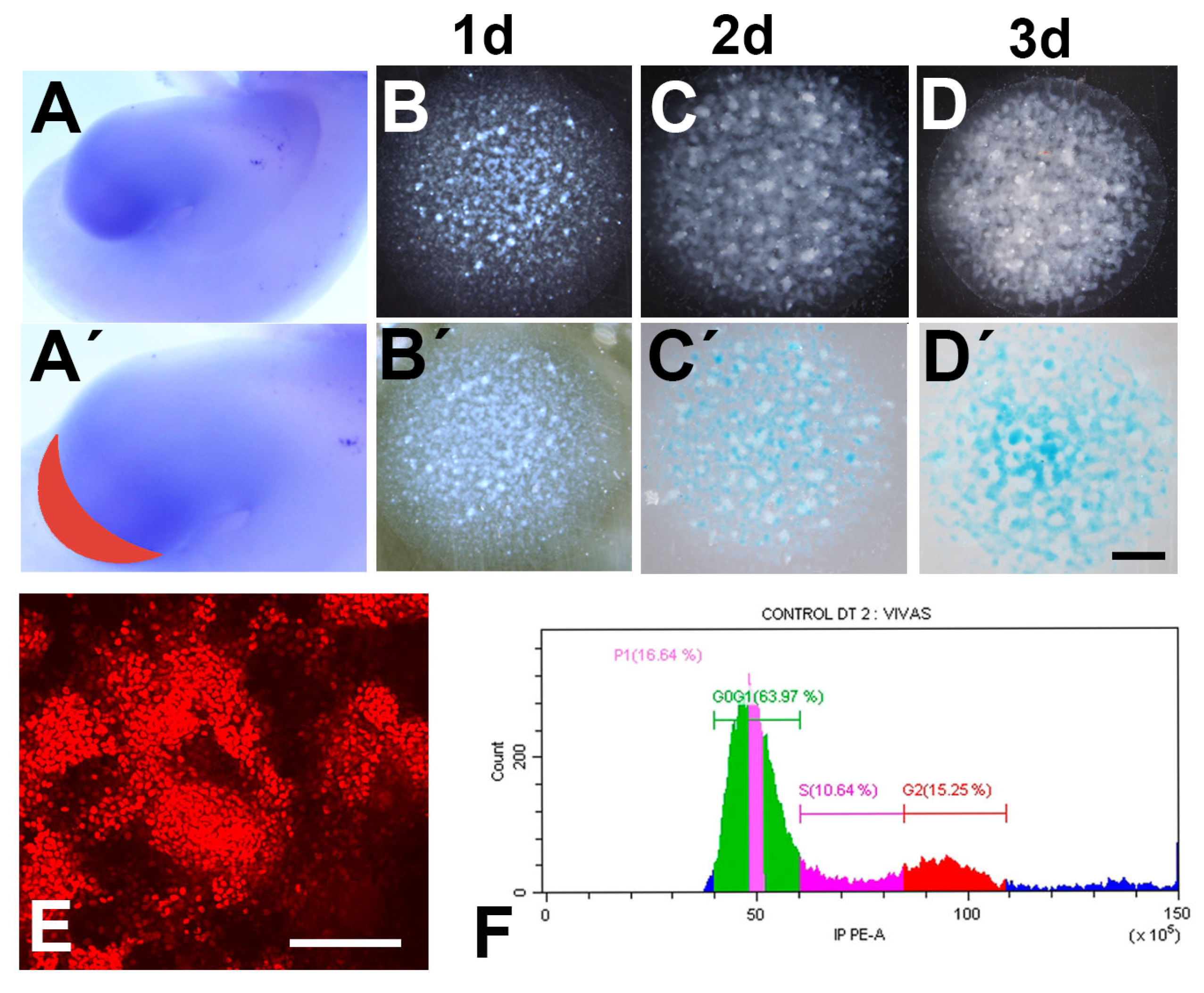
3.1. Differentiation, Proliferation, and Cell Death in Relation to the Presence or Absence of FBS in the Culture Medium
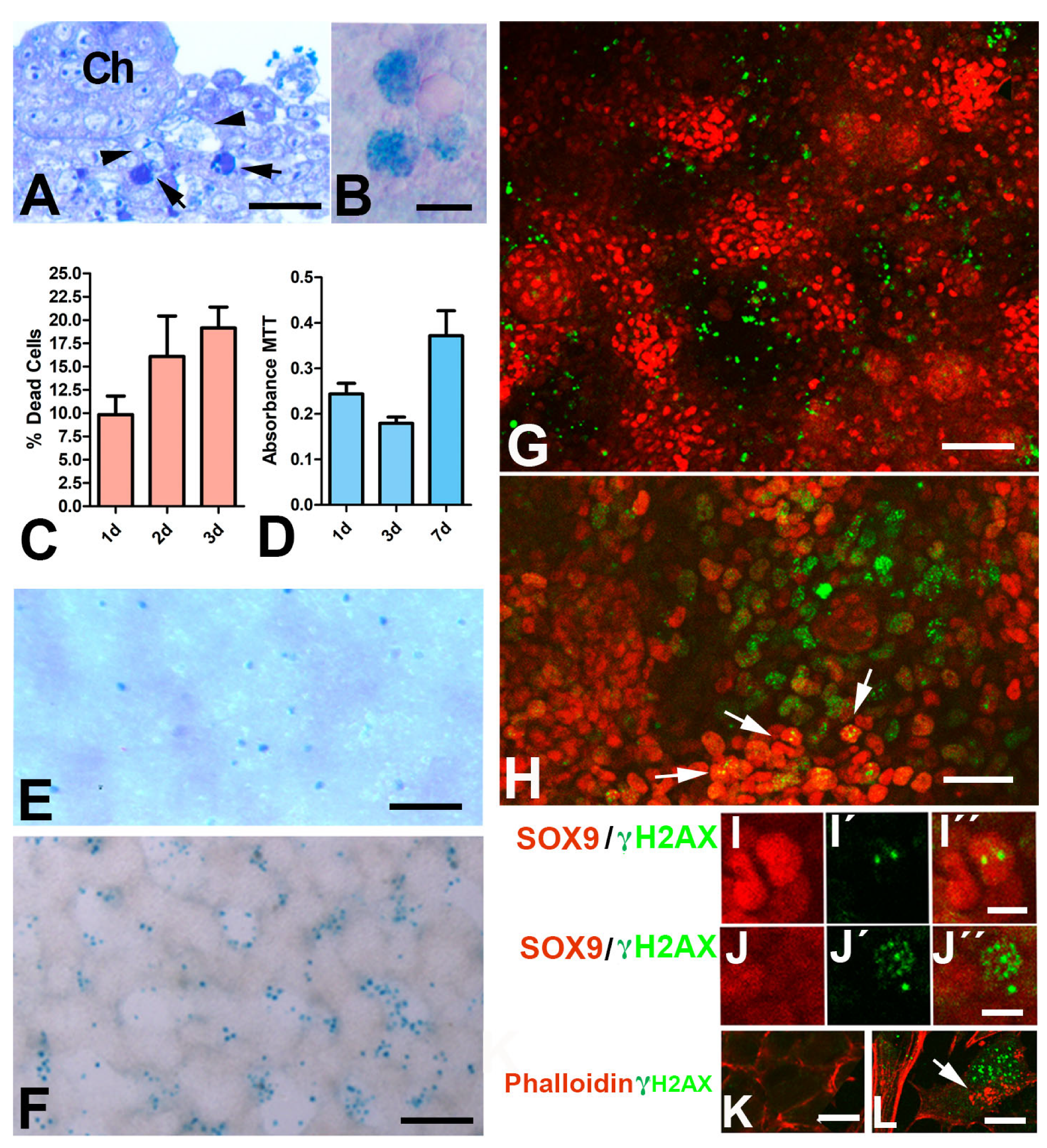
3.2. Patterned Cell Death and Cell Senescence
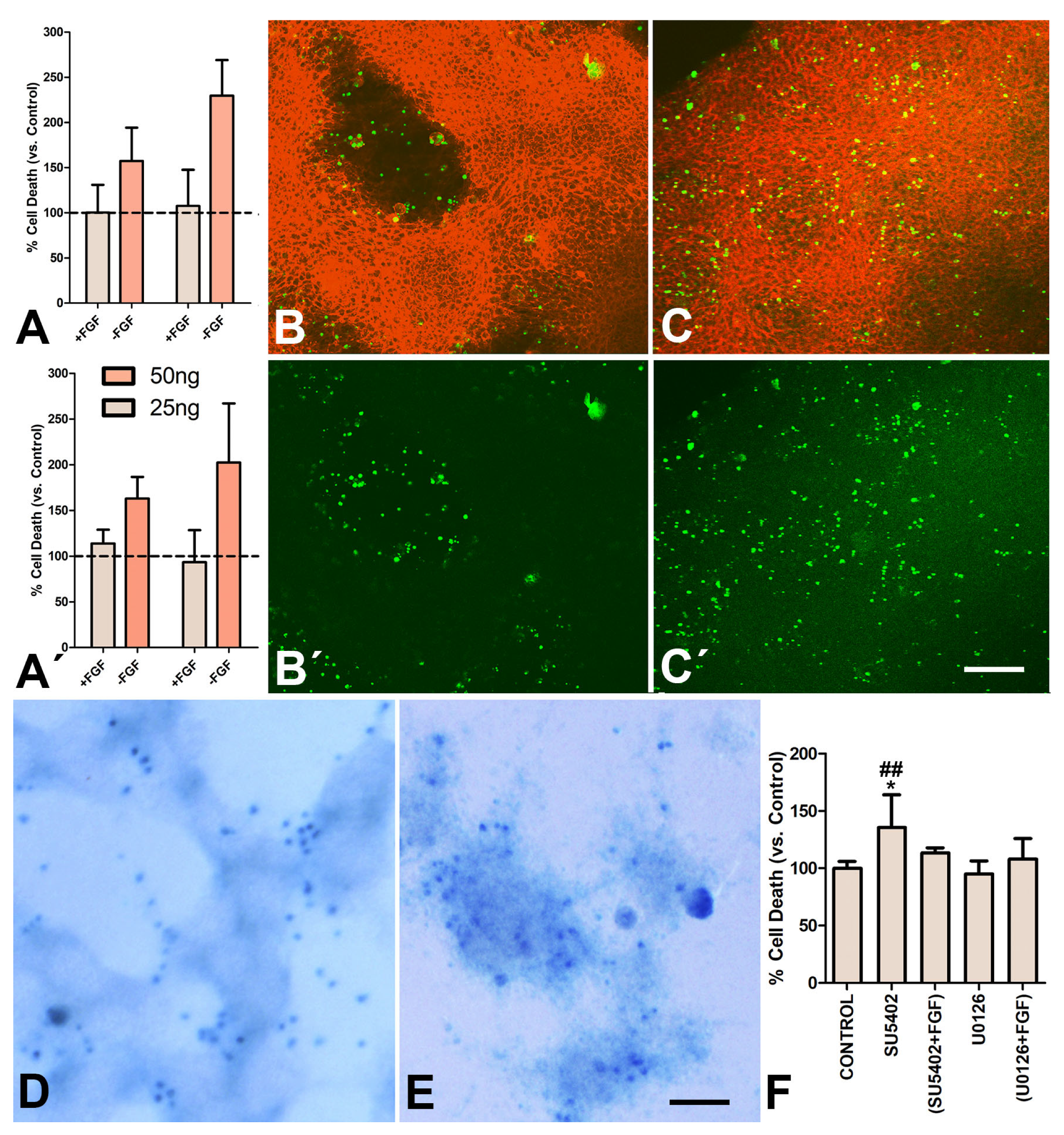
3.3. FGFs Sensitize Chondroprogenitors to Dying Signals
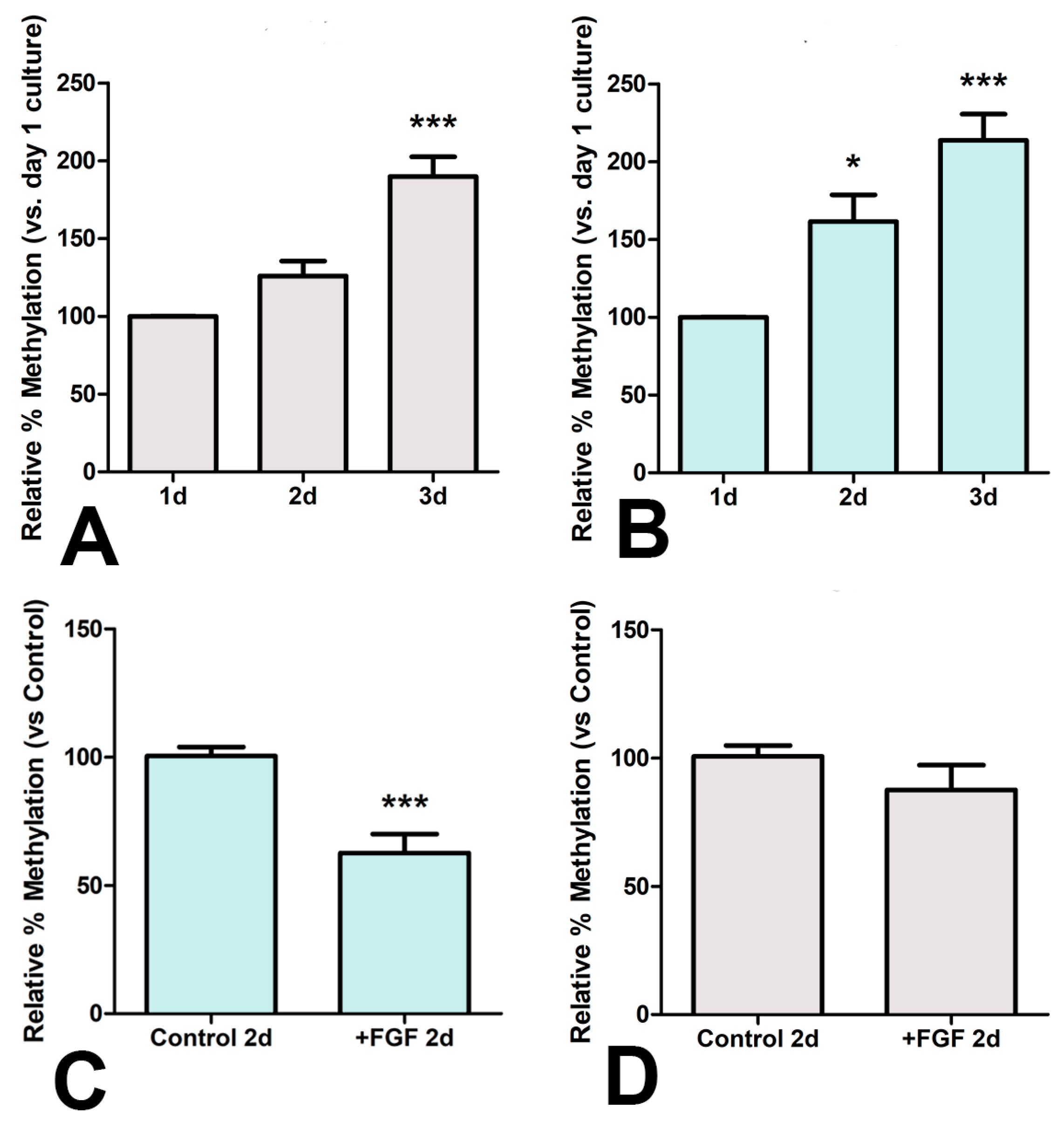
3.4. Epigenetic Influence of FGFs
3.5. Transcriptional Effects of FGFs
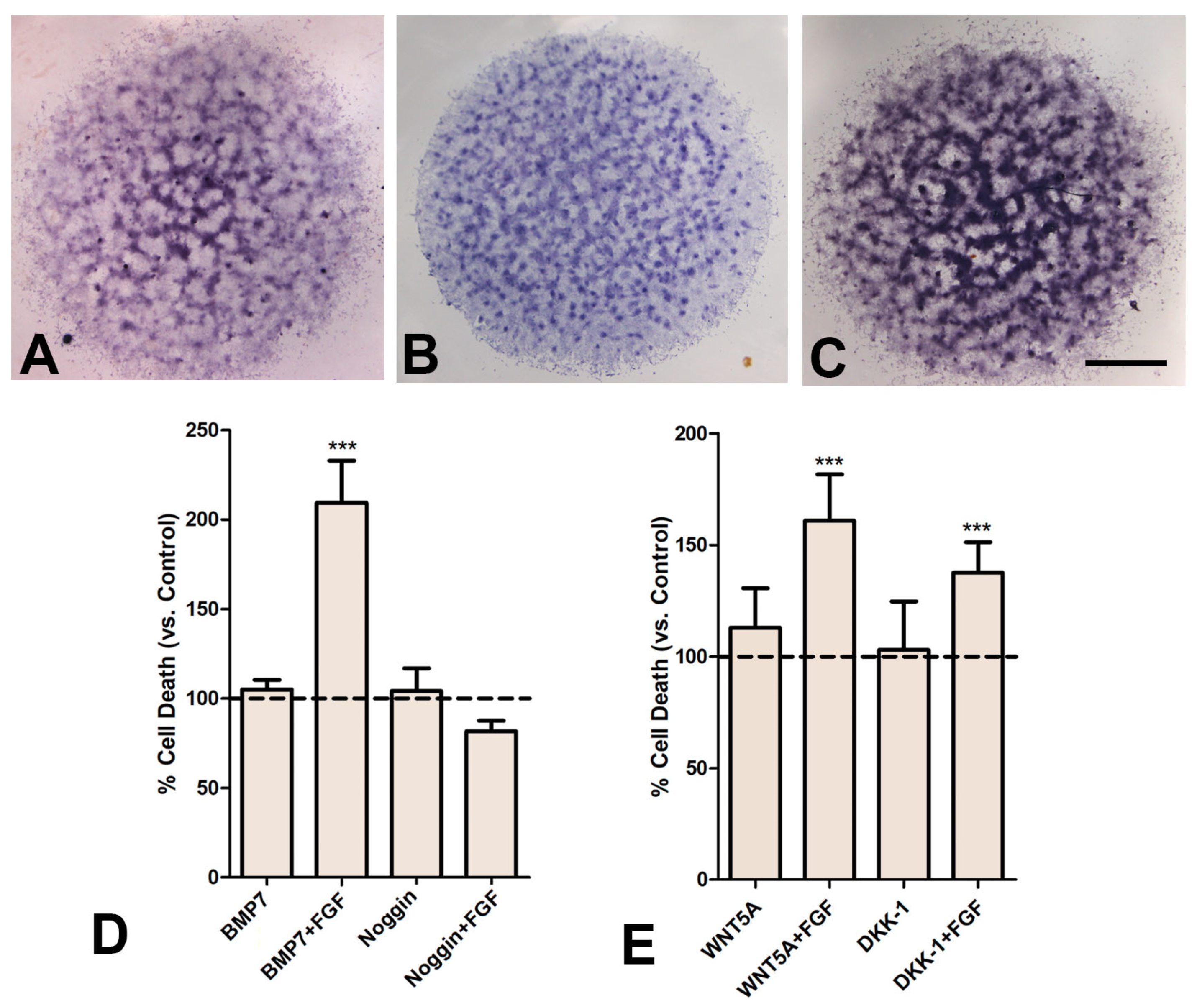
3.6. FGF2 Modifies the Response of Cultured Progenitors to BMP- and WNT- Signaling
3.6.1. The FGF/BMP Axis
3.6.2. The FGF/WNT Axis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Glücksmann, A. Cell Deaths in Normal Vertebrate Ontogeny. Biol. Rev. 1951, 26, 59–86. [Google Scholar] [CrossRef] [PubMed]
- Soteriou, D.; Fuchs, Y. A Matter of Life and Death: Stem Cell Survival in Tissue Regeneration and Tumour Formation. Nat. Rev. Cancer 2018, 18, 187–201. [Google Scholar] [CrossRef]
- Nie, X.; Luukko, K.; Fjeld, K.; Kvinnsland, I.H.; Kettunen, P. Developmental Expression of Dkk1-3 and Mmp9 and Apoptosis in Cranial Base of Mice. J. Mol. Histol. 2005, 36, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Montero, J.A.; Lorda-Diez, C.I.; Sanchez-Fernandez, C.; Hurle, J.M. Cell Death in the Developing Vertebrate Limb: A Locally Regulated Mechanism Contributing to Musculoskeletal Tissue Morphogenesis and Differentiation. Dev. Dyn. 2020, 250, 1236–1247. [Google Scholar] [CrossRef] [PubMed]
- DeLise, A.M.; Fischer, L.; Tuan, R.S. Cellular Interactions and Signaling in Cartilage Development. Osteoarthr. Cartil. 2000, 8, 309–334. [Google Scholar] [CrossRef] [Green Version]
- Yoon, B.S.; Ovchinnikov, D.A.; Yoshii, I.; Mishina, Y.; Behringer, R.R.; Lyons, K.M. Bmpr1a and Bmpr1b Have Overlapping Functions and Are Essential for Chondrogenesis in Vivo. Proc. Natl. Acad. Sci. USA 2005, 102, 5062–5067. [Google Scholar] [CrossRef] [Green Version]
- Mina, M.; Gluhak, J.; Upholt, W.B.; Kollar, E.J.; Rogers, B. Experimental Analysis of Msx-1 and Msx-2 Gene Expression during Chick Mandibular Morphogenesis. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 1995, 202, 195–214. [Google Scholar] [CrossRef]
- Shim, M.; Foley, J.; Anna, C.; Mishina, Y.; Eling, T. Embryonic Expression of Cyclooxygenase-2 Causes Malformations in Axial Skeleton. J. Biol. Chem. 2010, 285, 16206–16217. [Google Scholar] [CrossRef] [Green Version]
- Moftah, M.Z.; Downie, S.A.; Bronstein, N.B.; Mezentseva, N.; Pu, J.; Maher, P.A.; Newman, S.A. Ectodermal FGFs Induce Perinodular Inhibition of Limb Chondrogenesis in Vitro and in Vivo via FGF Receptor 2. Dev. Biol. 2002, 249, 270–282. [Google Scholar] [CrossRef] [Green Version]
- Kumar, M.; Ray, P.; Chapman, S.C. Fibroblast Growth Factor and Bone Morphogenetic Protein Signaling Are Required for Specifying Prechondrogenic Identity in Neural Crest-Derived Mesenchyme and Initiating the Chondrogenic Program. Dev. Dyn. 2012, 241, 1091–1103. [Google Scholar] [CrossRef]
- Kumar, D.; Lassar, A.B. Fibroblast Growth Factor Maintains Chondrogenic Potential of Limb Bud Mesenchymal Cells by Modulating DNMT3A Recruitment. Cell Rep. 2014, 8, 1419–1431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buckland, R.A.; Collinson, J.M.; Graham, E.; Davidson, D.R.; Hill, R.E. Antagonistic Effects of FGF4 on BMP Induction of Apoptosis and Chondrogenesis in the Chick Limb Bud. Mech. Dev. 1998, 71, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Semba, I.; Nonaka, K.; Takahashi, I.; Takahashi, K.; Dashner, R.; Shum, L.; Nuckolls, G.H.; Slavkin, H.C. Positionally-Dependent Chondrogenesis Induced by BMP4 Is Co-Regulated by Sox9 and Msx2. Dev. Dyn. 2000, 217, 401–414. [Google Scholar] [CrossRef]
- Zhao, W.; Allen, S.; Dhoot, G.K. FGF Mediated Sulf1 Regulation. FEBS Lett. 2007, 581, 4960–4964. [Google Scholar] [CrossRef] [Green Version]
- Reinhardt, R.; Gullotta, F.; Nusspaumer, G.; Ünal, E.; Ivanek, R.; Zuniga, A.; Zeller, R. Molecular Signatures Identify Immature Mesenchymal Progenitors in Early Mouse Limb Buds That Respond Differentially to Morphogen Signaling. Development 2019, 146, 173328. [Google Scholar] [CrossRef] [Green Version]
- Klumpers, D.D.; Mooney, D.J.; Smit, T.H. From Skeletal Development to Tissue Engineering: Lessons from the Micromass Assay. Tissue Eng. Part B. Rev. 2015, 21, 427–437. [Google Scholar] [CrossRef] [Green Version]
- Raspopovic, J.; Marcon, L.; Russo, L.; Sharpe, J. Digit Patterning Is Controlled by a Bmp-Sox9-Wnt Turing Network Modulated by Morphogen Gradients. Science 2014, 345, 566–570. [Google Scholar] [CrossRef]
- Omi, M.; Sato-Maeda, M.; Ide, H. Role of Chondrogenic Tissue in Programmed Cell Death and BMP Expression in Chick Limb Buds. Int. J. Dev. Biol. 2000, 44, 381–388. [Google Scholar]
- Yamashita, A.; Krawetz, R.; Rancourt, D.E. Loss of Discordant Cells during Micro-Mass Differentiation of Embryonic Stem Cells into the Chondrocyte Lineage. Cell Death Differ. 2009, 16, 278–286. [Google Scholar] [CrossRef]
- Montero, J.A.; Lorda-Diez, C.I.; Hurle, J.M. Confluence of Cellular Degradation Pathways During Interdigital Tissue Remodeling in Embryonic Tetrapods. Front. Cell Dev. Biol. 2020, 8, 1217. [Google Scholar] [CrossRef]
- Debacq-Chainiaux, F.; Erusalimsky, J.D.; Campisi, J.; Toussaint, O. Protocols to Detect Senescence-Associated Beta-Galactosidase (SA-Βgal) Activity, a Biomarker of Senescent Cells in Culture and in Vivo. Nat. Protoc. 2009, 4, 1798–1806. [Google Scholar] [CrossRef] [PubMed]
- Rolfe, R.A.; Shea, C.A.; Murphy, P. Geometric Analysis of Chondrogenic Self-Organisation of Embryonic Limb Bud Cells in Micromass Culture. Cell Tissue Res. 2022, 388, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Downie, S.A.; Newman, S.A. Morphogenetic Differences between Fore and Hind Limb Precartilage Mesenchyme: Relation to Mechanisms of Skeletal Pattern Formation. Dev. Biol. 1994, 162, 195–208. [Google Scholar] [CrossRef] [PubMed]
- Merino, R.; Macias, D.; Gañan, Y.; Rodriguez-Leon, J.; Economides, A.N.; Rodriguez-Esteban, C.; Izpisua-Belmonte, J.C.; Hurle, J.M. Control of Digit Formation by Activin Signalling. Development 1999, 126, 2161–2170. [Google Scholar] [CrossRef]
- Kiskowski, M.A.; Alber, M.S.; Thomas, G.L.; Glazier, J.A.; Bronstein, N.B.; Pu, J.; Newman, S.A. Interplay between Activator–Inhibitor Coupling and Cell-Matrix Adhesion in a Cellular Automaton Model for Chondrogenic Patterning. Dev. Biol. 2004, 271, 372–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yadav, P.; Chatterjee, K.; Saini, D.K. Senescent Cells in 3D Culture Show Suppressed Senescence Signatures. Biomater. Sci. 2021, 9, 6461–6473. [Google Scholar] [CrossRef]
- Montero, J.A.; Sanchez-Fernandez, C.; Lorda-Diez, C.I.; Garcia-Porrero, J.A.; Hurle, J.M. DNA Damage Precedes Apoptosis during the Regression of the Interdigital Tissue in Vertebrate Embryos. Sci. Rep. 2016, 6, 35478. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Fernandez, C.; Lorda-Diez, C.I.; Hurlé, J.M.; Montero, J.A. The Methylation Status of the Embryonic Limb Skeletal Progenitors Determines Their Cell Fate in Chicken. Commun. Biol. 2020, 3, 283. [Google Scholar] [CrossRef]
- Orlando, L.; Tanasijevic, B.; Nakanishi, M.; Reid, J.C.; García-Rodríguez, J.L.; Chauhan, K.D.; Porras, D.P.; Aslostovar, L.; Lu, J.D.; Shapovalova, Z.; et al. Phosphorylation State of the Histone Variant H2A.X Controls Human Stem and Progenitor Cell Fate Decisions. Cell Rep. 2021, 34, 108818. [Google Scholar] [CrossRef]
- Zuzarte-Luis, V.; Berciano, M.T.; Lafarga, M.; Hurlé, J.M. Caspase Redundancy and Release of Mitochondrial Apoptotic Factors Characterize Interdigital Apoptosis. Apoptosis 2006, 11, 701–715. [Google Scholar] [CrossRef]
- Macias, D.; Gañan, Y.; Ros, M.A.; Hurle, J.M. In Vivo Inhibition of Programmed Cell Death by Local Administration of FGF-2 and FGF-4 in the Interdigital Areas of the Embryonic Chick Leg Bud. Anat. Embryol. 1996, 193, 533–541. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Martínez, R.; Castro-Obregón, S.; Covarrubias, L. Progressive Interdigital Cell Death: Regulation by the Antagonistic Interaction between Fibroblast Growth Factor 8 and Retinoic Acid. Development 2009, 136, 3669–3678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montero, J.A.; Gañan, Y.; Macias, D.; Rodriguez-Leon, J.; Sanz-Ezquerro, J.J.; Merino, R.; Chimal-Monroy, J.; Nieto, M.A.; Hurle, J.M. Role of FGFs in the Control of Programmed Cell Death during Limb Development. Development 2001, 128, 2075–2084. [Google Scholar] [CrossRef]
- Sanchez-Fernandez, C.; Lorda-Diez, C.I.; García-Porrero, J.A.; Montero, J.A.; Hurlé, J.M. UHRF Genes Regulate Programmed Interdigital Tissue Regression and Chondrogenesis in the Embryonic Limb. Cell Death Dis. 2019, 10, 347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ornitz, D.M.; Itoh, N. The Fibroblast Growth Factor Signaling Pathway. Wiley Interdiscip. Rev. Dev. Biol. 2015, 4, 215–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, W.; Cao, Y.; Zhang, Z.; Du, F.; Shi, Y.; Li, X.; Zhang, Q. Targeted Delivery of FGF2 to Subchondral Bone Enhanced the Repair of Articular Cartilage Defect. Acta Biomater. 2018, 69, 170–182. [Google Scholar] [CrossRef]
- Macias, D.; Gañan, Y.; Sampath, T.K.; Piedra, M.E.; Ros, M.A.; Hurle, J.M. Role of BMP-2 and OP-1 (BMP-7) in Programmed Cell Death and Skeletogenesis during Chick Limb Development. Development 1997, 124, 1109–1117. [Google Scholar] [CrossRef]
- Díaz-Hernández, M.E.; Galván-Hernández, C.I.; Marín-Llera, J.C.; Camargo-Sosa, K.; Bustamante, M.; Wischin, S.; Chimal-Monroy, J. Activation of the WNT-BMP-FGF Regulatory Network Induces the Onset of Cell Death in Anterior Mesodermal Cells to Establish the ANZ. Front. Cell Dev. Biol. 2021, 9, 703836. [Google Scholar] [CrossRef]
- Kaltcheva, M.M.; Anderson, M.J.; Harfe, B.D.; Lewandoski, M. BMPs Are Direct Triggers of Interdigital Programmed Cell Death. Dev. Biol. 2016, 411, 266–276. [Google Scholar] [CrossRef]
- Bandyopadhyay, A.; Tsuji, K.; Cox, K.; Harfe, B.D.; Rosen, V.; Tabin, C.J. Genetic Analysis of the Roles of BMP2, BMP4, and BMP7 in Limb Patterning and Skeletogenesis. PLoS Genet. 2006, 2, 2116–2130. [Google Scholar] [CrossRef] [Green Version]
- Zuzarte-Luís, V.; Montero, J.A.; Rodriguez-León, J.; Merino, R.; Rodríguez-Rey, J.C.; Hurlé, J.M. A New Role for BMP5 during Limb Development Acting through the Synergic Activation of Smad and MAPK Pathways. Dev. Biol. 2004, 272, 39–52. [Google Scholar] [CrossRef] [PubMed]
- Villacorte, M.; Suzuki, K.; Hayashi, K.; de Sousa Lopes, S.C.; Haraguchi, R.; Taketo, M.M.; Nakagata, N.; Yamada, G. Antagonistic Crosstalk of Wnt/Beta-Catenin/Bmp Signaling within the Apical Ectodermal Ridge (AER) Regulates Interdigit Formation. Biochem. Biophys. Res. Commun. 2010, 391, 1653–1657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, B.; Ajima, R.; Yang, W.; Li, C.; Song, H.; Anderson, M.J.; Liu, R.R.; Lewandoski, M.B.; Yamaguchi, T.P.; Yang, Y. Coordinated Directional Outgrowth and Pattern Formation by Integration of Wnt5a and Fgf Signaling in Planar Cell Polarity. Dev. 2018, 145, 163824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Topol, L.; Jiang, X.; Choi, H.; Garrett-Beal, L.; Carolan, P.J.; Yang, Y. Wnt-5a Inhibits the Canonical Wnt Pathway by Promoting GSK-3-Independent Beta-Catenin Degradation. J. Cell Biol. 2003, 162, 899–908. [Google Scholar] [CrossRef] [PubMed]
- Grotewold, L.; Rüther, U. The Wnt Antagonist Dickkopf-1 Is Regulated by Bmp Signaling and c-Jun and Modulates Programmed Cell Death. EMBO J. 2002, 21, 966–975. [Google Scholar] [CrossRef] [Green Version]
- Farrera-Hernández, A.; Marín-Llera, J.C.; Chimal-Monroy, J. WNT5A-Ca2+-CaN-NFAT Signalling Plays a Permissive Role during Cartilage Differentiation in Embryonic Chick Digit Development. Dev. Biol. 2021, 469, 86–95. [Google Scholar] [CrossRef]
- Christley, S.; Alber, M.S.; Newman, S.A. Patterns of Mesenchymal Condensation in a Multiscale, Discrete Stochastic Model. PLOS Comput. Biol. 2007, 3, e76. [Google Scholar] [CrossRef]
- de Mera-Rodríguez, J.A.; Álvarez-Hernán, G.; Gañán, Y.; Martín-Partido, G.; Rodríguez-León, J.; Francisco-Morcillo, J. Is Senescence-Associated β-Galactosidase a Reliable in Vivo Marker of Cellular Senescence During Embryonic Development? Front. Cell Dev. Biol. 2021, 9, 623175. [Google Scholar] [CrossRef]
- Lorda-Diez, C.I.; Garcia-Riart, B.; Montero, J.A.; Rodriguez-León, J.; Garcia-Porrero, J.A.; Hurlé, J.M. Apoptosis during Embryonic Tissue Remodeling Is Accompanied by Cell Senescence. Aging 2015, 7, 974–985. [Google Scholar] [CrossRef] [Green Version]
- Muñoz-Espín, D.; Cañamero, M.; Maraver, A.; Gómez-López, G.; Contreras, J.; Murillo-Cuesta, S.; Rodríguez-Baeza, A.; Varela-Nieto, I.; Ruberte, J.; Collado, M.; et al. XProgrammed Cell Senescence during Mammalian Embryonic Development. Cell 2013, 155, 1104. [Google Scholar] [CrossRef] [Green Version]
- Rhinn, M.; Ritschka, B.; Keyes, W.M. Cellular Senescence in Development, Regeneration and Disease. Development 2019, 146, 151837. [Google Scholar] [CrossRef] [PubMed]
- Varela-Nieto, I.; Palmero, I.; Magariños, M. Complementary and Distinct Roles of Autophagy, Apoptosis and Senescence during Early Inner Ear Development. Hear. Res. 2019, 376, 86–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Belly, H.; Paluch, E.K.; Chalut, K.J. Interplay between Mechanics and Signalling in Regulating Cell Fate. Nat. Rev. Mol. Cell Biol. 2022, 23, 465–480. [Google Scholar] [CrossRef] [PubMed]
- Parada, C.; Banavar, S.P.; Khalilian, P.; Rigaud, S.; Michaut, A.; Liu, Y.; Joshy, D.M.; Campàs, O.; Gros, J. Mechanical Feedback Defines Organizing Centers to Drive Digit Emergence. Dev. Cell 2022, 57, 854–866.e6. [Google Scholar] [CrossRef]
- Weng, S.; Huebner, R.J.; Wallingford, J.B. Convergent Extension Requires Adhesion-Dependent Biomechanical Integration of Cell Crawling and Junction Contraction. Cell Rep. 2022, 39, 110666. [Google Scholar] [CrossRef] [PubMed]
- Barna, M.; Niswander, L. Visualization of Cartilage Formation: Insight into Cellular Properties of Skeletal Progenitors and Chondrodysplasia Syndromes. Dev. Cell 2007, 12, 931–941. [Google Scholar] [CrossRef] [Green Version]
- Lim, J.; Tu, X.; Choi, K.; Akiyama, H.; Mishina, Y.; Long, F. BMP-Smad4 Signaling Is Required for Precartilaginous Mesenchymal Condensation Independent of Sox9 in the Mouse. Dev. Biol. 2015, 400, 132–138. [Google Scholar] [CrossRef] [Green Version]
- Juhász, T.; Matta, C.; Somogyi, C.; Katona, É.; Takács, R.; Soha, R.F.; Szabó, I.A.; Cserháti, C.; Sződy, R.; Karácsonyi, Z.; et al. Mechanical Loading Stimulates Chondrogenesis via the PKA/CREB-Sox9 and PP2A Pathways in Chicken Micromass Cultures. Cell. Signal. 2014, 26, 468–482. [Google Scholar] [CrossRef] [Green Version]
- Akiyama, H.; Chaboissier, M.C.; Martin, J.F.; Schedl, A.; De Crombrugghe, B. The Transcription Factor Sox9 Has Essential Roles in Successive Steps of the Chondrocyte Differentiation Pathway and Is Required for Expression of Sox5 and Sox6. Genes Dev. 2002, 16, 2813–2828. [Google Scholar] [CrossRef] [Green Version]
- Coutu, D.L.; Galipeau, J. Roles of FGF Signaling in Stem Cell Self-Renewal, Senescence and Aging. Aging 2011, 3, 920–933. [Google Scholar] [CrossRef] [Green Version]
- Martin, G.R. The Roles of FGFs in the Early Development of Vertebrate Limbs. Genes Dev. 1998, 12, 1571–1586. [Google Scholar] [CrossRef] [PubMed]
- Verheyden, J.M.; Lewandoski, M.; Deng, C.; Harfe, B.D.; Sun, X. Conditional Inactivation of Fgfr1 in Mouse Defines Its Role in Limb Bud Establishment, Outgrowth and Digit Patterning. Development 2005, 132, 4235–4245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Weinstein, M.; Li, C.; Naski, M.; Cohen, R.I.; Ornitz, D.M.; Leder, P.; Deng, C. Fibroblast Growth Factor Receptor 2 (FGFR2)-Mediated Reciprocal Regulation Loop between FGF8 and FGF10 Is Essential for Limb Induction. Development 1998, 125, 753–765. [Google Scholar] [CrossRef] [PubMed]
- Gañan, Y.; Macias, D.; Basco, R.D.; Merino, R.; Hurle, J.M. Morphological Diversity of the Avian Foot Is Related with the Pattern of Msx Gene Expression in the Developing Autopod. Dev. Biol. 1998, 196, 33–41. [Google Scholar] [CrossRef] [Green Version]
- Chimal-Monroy, J.; Rodriguez-Leon, J.; Montero, J.A.; Gañan, Y.; Macias, D.; Merino, R.; Hurle, J.M. Analysis of the Molecular Cascade Responsible for Mesodermal Limb Chondrogenesis: Sox Genes and BMP Signaling. Dev. Biol. 2003, 257, 292–301. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Shepherd, J.; Zhao, D.; Bollu, L.R.; Tahaney, W.M.; Hill, J.; Zhang, Y.; Mazumdar, A.; Brown, P.H. SOX9 Is Essential for Triple-Negative Breast Cancer Cell Survival and Metastasis. Mol. Cancer Res. 2020, 18, 1825–1838. [Google Scholar] [CrossRef]
- Hernández-Martínez, R.; Covarrubias, L. Interdigital Cell Death Function and Regulation: New Insights on an Old Programmed Cell Death Model. Dev. Growth Differ. 2011, 53, 245–258. [Google Scholar] [CrossRef]
- Chimal-Monroy, J.; Montero, J.A.; Gañan, Y.; Macias, D.; Garcia-Porrero, J.A.; Hurle, J.M. Comparative Analysis of the Expression and Regulation of Wnt5a, Fz4, and Frzb1 during Digit Formation and in Micromass Cultures. Dev. Dyn. 2002, 224, 314–320. [Google Scholar] [CrossRef]
- Mukhopadhyay, M.; Shtrom, S.; Rodriguez-Esteban, C.; Chen, L.; Tsukui, T.; Gomer, L.; Dorward, D.W.; Glinka, A.; Grinberg, A.; Huang, S.P.; et al. Dickkopf1 Is Required for Embryonic Head Induction and Limb Morphogenesis in the Mouse. Dev. Cell 2001, 1, 423–434. [Google Scholar] [CrossRef] [Green Version]
- Ellies, D.L.; Church, V.; Francis-West, P.; Lumsden, A. The WNT Antagonist CSFRP2 Modulates Programmed Cell Death in the Developing Hindbrain. Development 2000, 127, 5285–5295. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene. | FGF2 vs Control. | Gene. | FGF2 vs Control. |
|---|---|---|---|
| Differentiation markers. | FGF signaling. | ||
| Sclx | 1.06 ± 0.48 | Sprouty1 | 21.46 ± 13.53 *** |
| Sox9 | 1.40 ± 0.99 | FgfR1 | 1.11 ± 0.46 |
| Senescence markers. | FgfR2 | 0.54 ± 0.20 *** | |
| p21 | 2.70 ± 1.08 *** | FgfR3 | 0.78 ± 0.50 |
| GBL1 | 1.42 ± 0.46 | FgfR4 | 0.94 ± 0.47 |
| Cathepsin D | 2.38 ± 0.84 * | BMP signaling. | |
| Mmp2 | 1.30 ± 0.26 | Bmp2 | 2.30 ± 0.86 ** |
| Il-6 | 2.59 ± 0.93 *,a | Bmp4 | 2.67 ± 1.05 ** |
| Cell Death markers. | Bmp5 | 0.40 ± 0.23 *** | |
| Bcl2 | 1.19 ± 0.46 | Bmp7 | 3.75 ± 1.63 *** |
| Bak1 | 2.56 ± 0.91 *** | Noggin | 0.73 ± 0.52 |
| Undifferentiation markers. | WNT signaling. | ||
| Msx1 | 1.27 ± 0.58 | Wnt5a | 0.90 ± 0.34 |
| Msx2 | 1.34 ± 0.57 | Dkk1 | 4.71 ± 1.80 ** |
| Oct4 | 1.40 ± 0.23 a | ||
| Epigenetic modulators. | |||
| Dnmt1 | 0.56 ± 0.24 * | ||
| Dnmt3A | 0.61 ± 0.31 | ||
| Dnmt3B | 0.96 ± 0.35 | ||
| TET3 | 0.69 ± 0.19 | ||
| HDAC2 | 0.68 ± 0.27 | Note: a very low expression level CTs > 30 vs Gapdh (CT ≈ 18). | |
| HDAC3 | 0.55 ± 0.24 * | ||
| HDAC8 | 0.77 ± 0.26 | ||
| Prmt5 | 1.08 ± 0.10 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duarte-Olivenza, C.; Hurle, J.M.; Montero, J.A.; Lorda-Diez, C.I. Modeling the Differentiation of Embryonic Limb Chondroprogenitors by Cell Death and Cell Senescence in High Density Micromass Cultures and Their Regulation by FGF Signaling. Cells 2023, 12, 175. https://doi.org/10.3390/cells12010175
Duarte-Olivenza C, Hurle JM, Montero JA, Lorda-Diez CI. Modeling the Differentiation of Embryonic Limb Chondroprogenitors by Cell Death and Cell Senescence in High Density Micromass Cultures and Their Regulation by FGF Signaling. Cells. 2023; 12(1):175. https://doi.org/10.3390/cells12010175
Chicago/Turabian StyleDuarte-Olivenza, Cristina, Juan M. Hurle, Juan A. Montero, and Carlos I. Lorda-Diez. 2023. "Modeling the Differentiation of Embryonic Limb Chondroprogenitors by Cell Death and Cell Senescence in High Density Micromass Cultures and Their Regulation by FGF Signaling" Cells 12, no. 1: 175. https://doi.org/10.3390/cells12010175