
Inhibition of EZH2 Ameliorates Sepsis Acute Lung Injury (SALI) and Non-Small-Cell Lung Cancer (NSCLC) Proliferation through the PD-L1 Pathway

Abstract
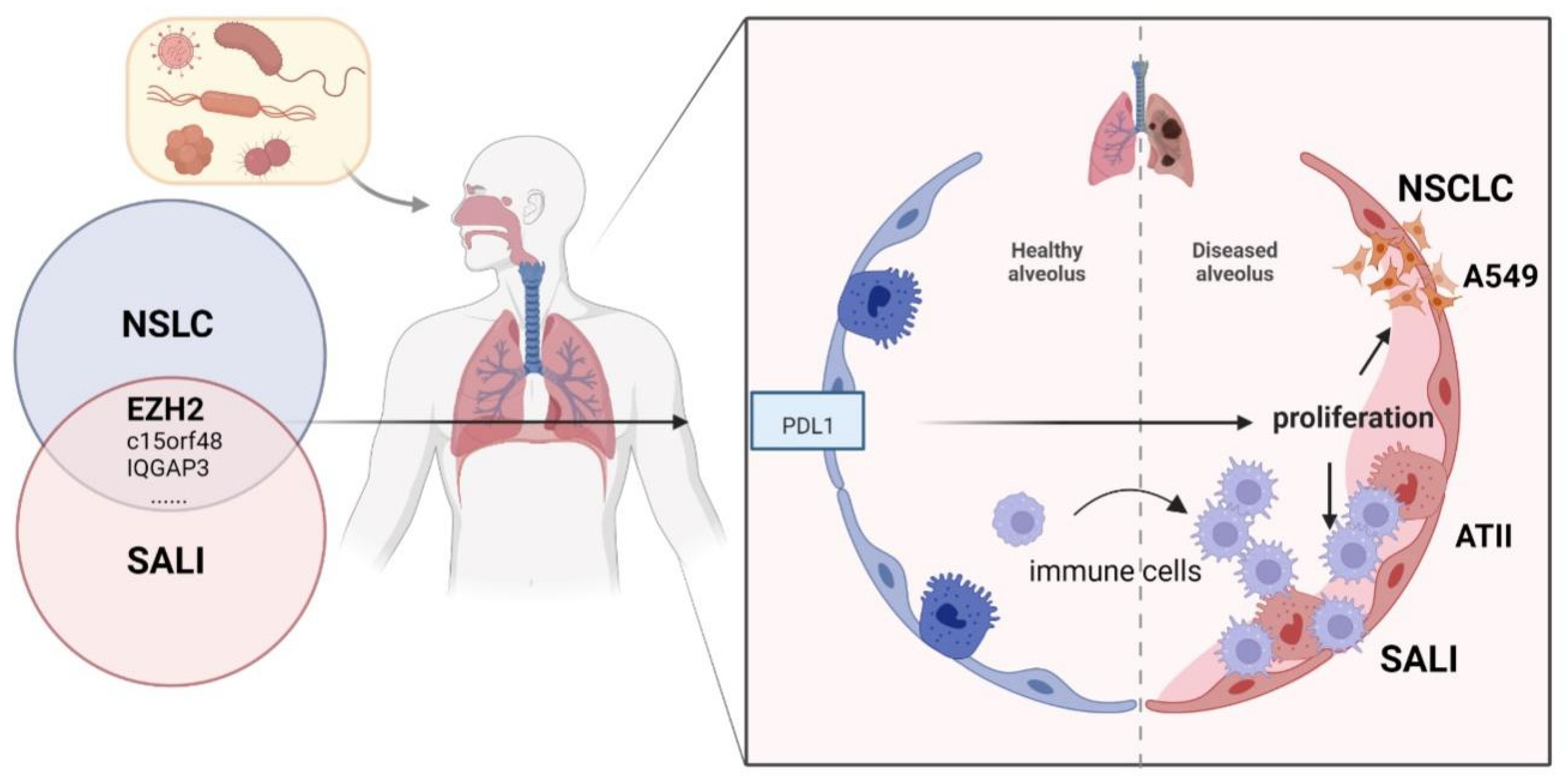
:1. Introduction
2. Materials and Methods
2.1. Bioinformatics Analysis
2.1.1. Dataset Download
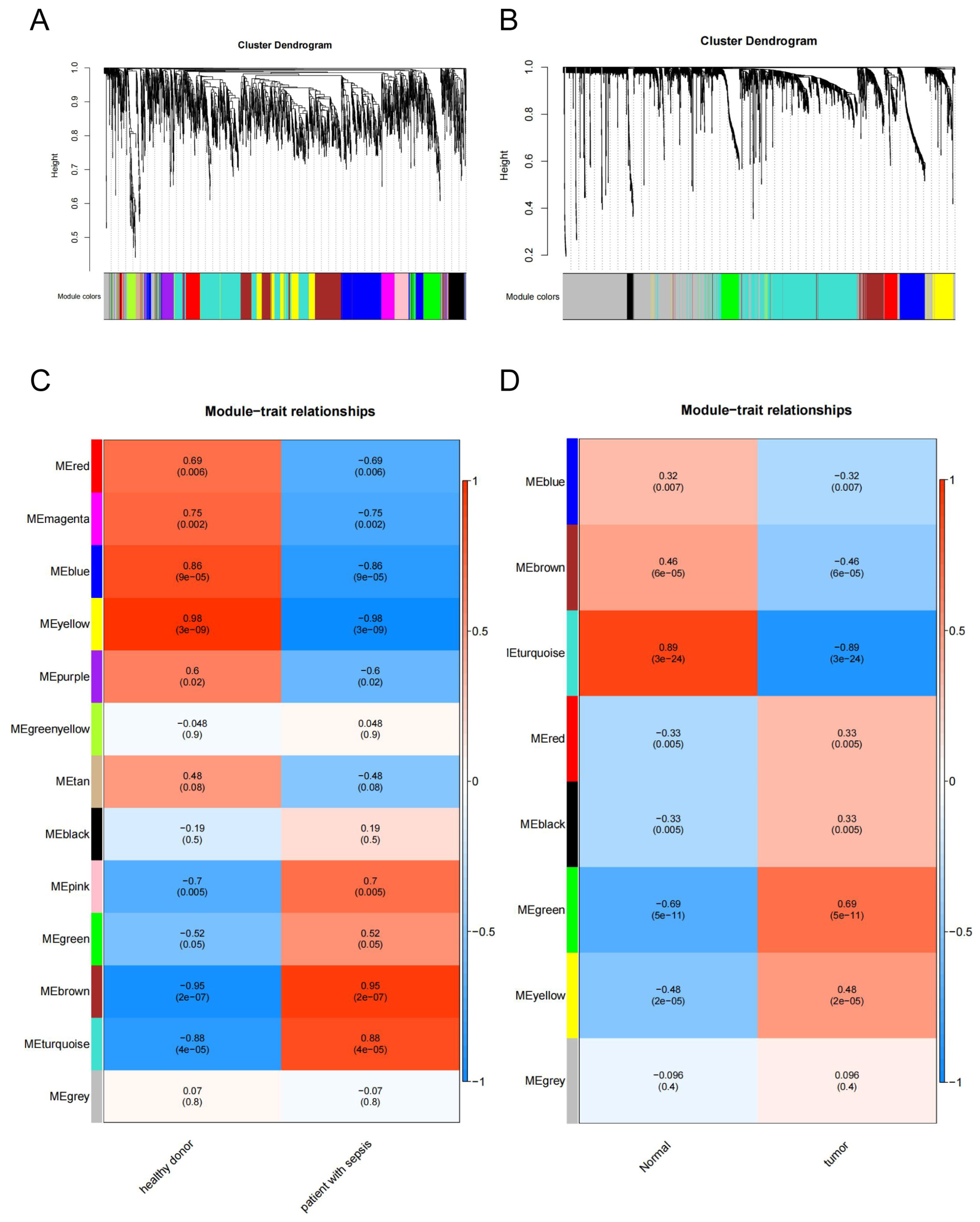
2.1.2. Weighted Gene Co-Expression Network Analysis
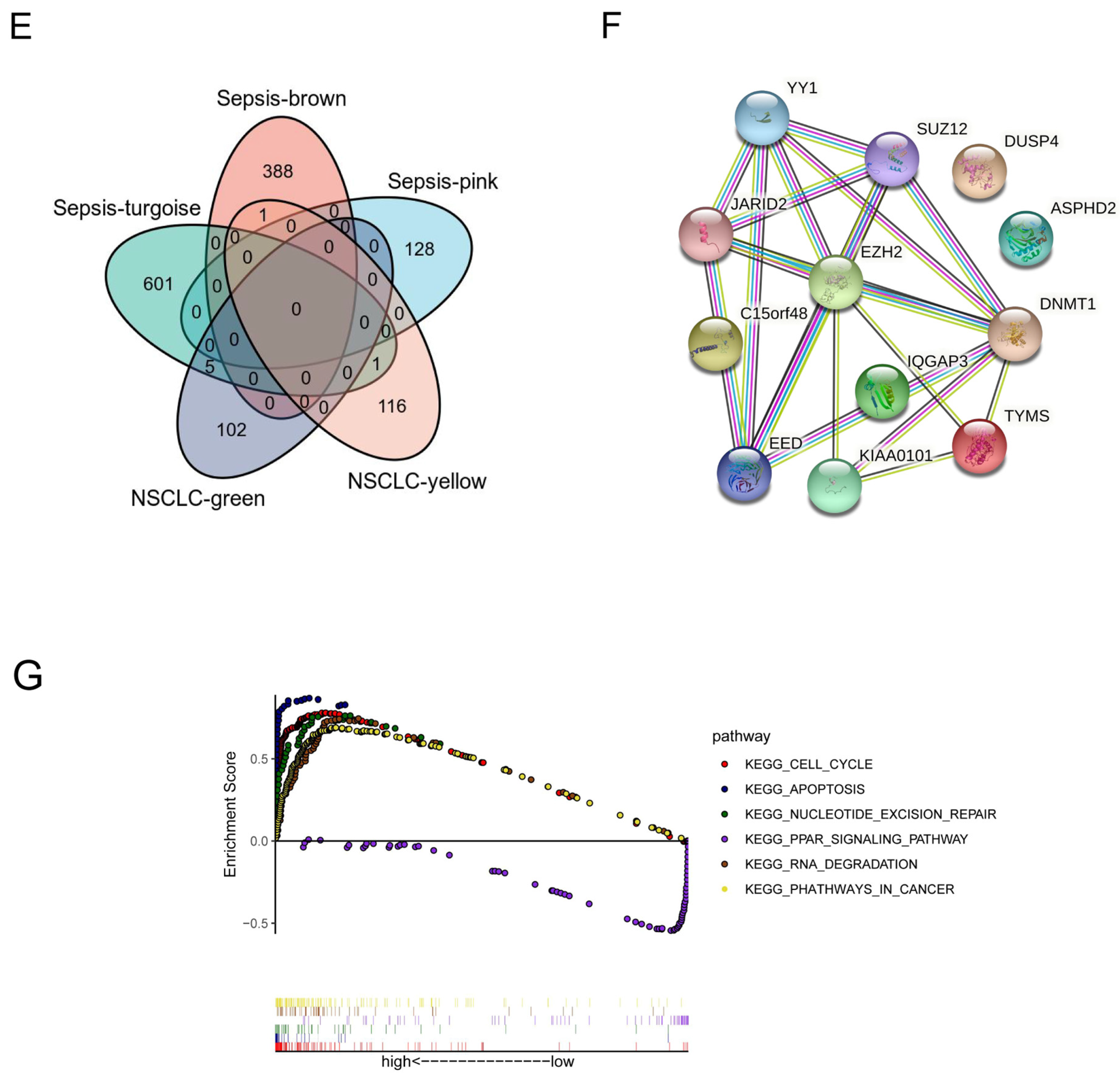
2.1.3. PPI Network Construction and Key Target Screening
2.1.4. Assessment of the Immune Landscape
2.2. Molecular Biological Verification
2.2.1. Basic Information
2.2.2. Entry and Exclusion Criteria
2.2.3. Cell Culture
2.2.4. Drug
2.2.5. Reagents
2.2.6. Western Blot
2.2.7. Cell Counting Kit-8
2.2.8. Colony Formation
2.2.9. Transwell Co-Culture Assay
2.2.10. Flow Cytometry Analysis
2.3. Data Processing and Statistical Analysis
3. Results
3.1. Identification of a Shared Molecular Connection between SALI and NSCLC
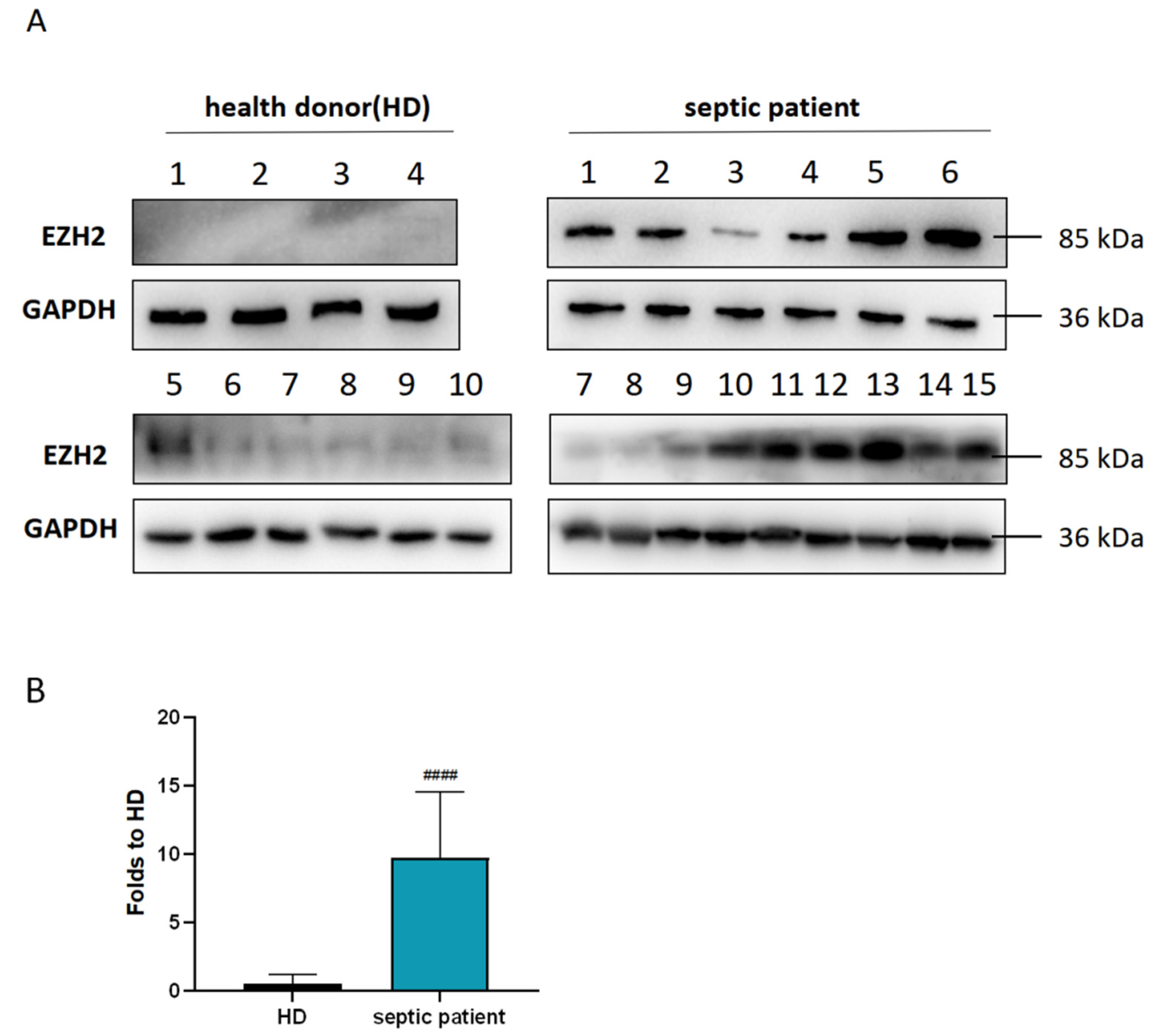
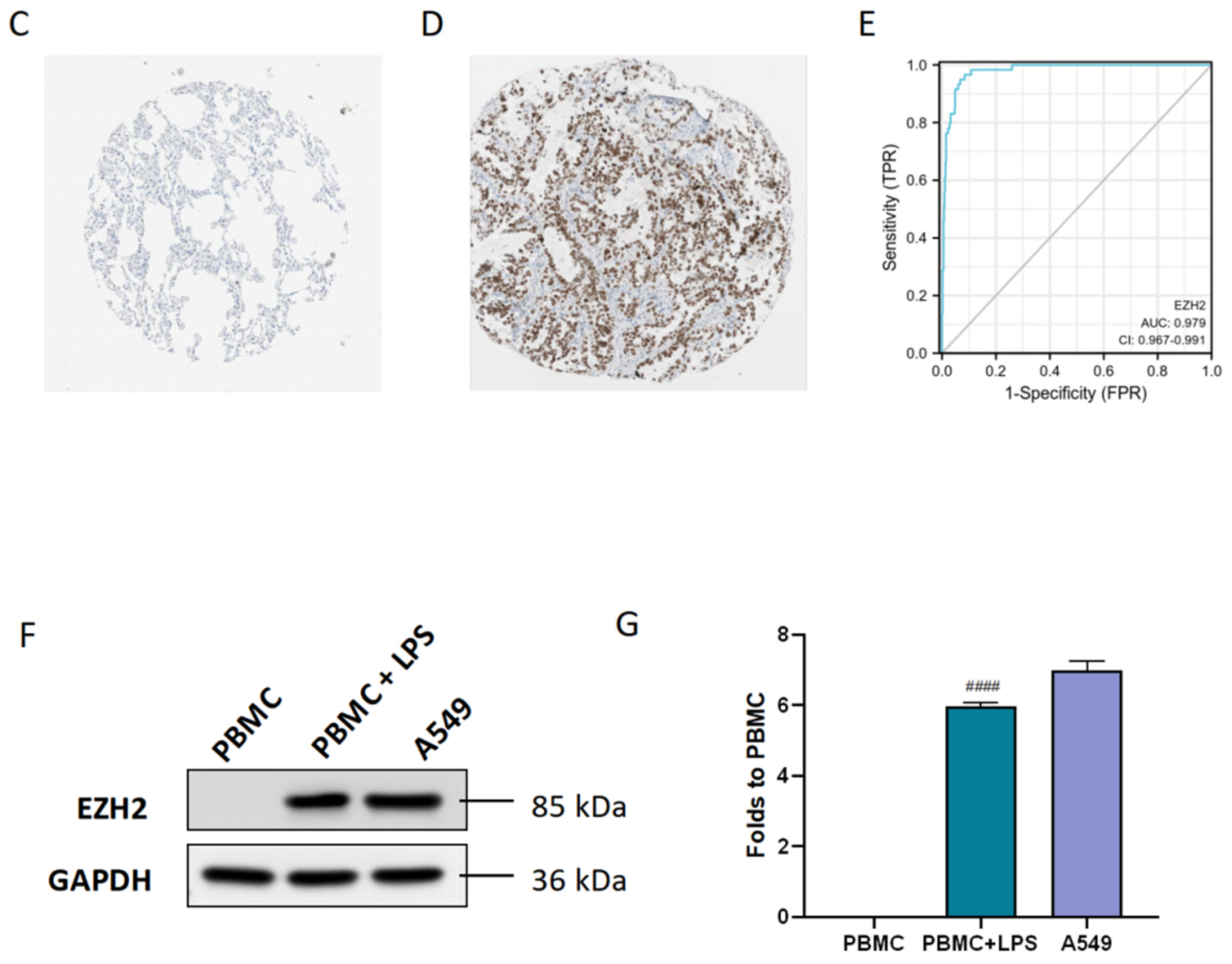
3.2. Diagnostic and Prognostic Value of the Hub Gene EZH2

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HD | Septic Patient | p-Value | ||
|---|---|---|---|---|
| Low EZH2 | High EZH2 | |||
| Basic Information | ||||
| Patients (number of patients) | 10 | 7 | 8 | - |
| Gender: number of male patients (% within gender) | 5(50%) | 3(42.86%) | 3(37.5%) | 0.866 |
| Age range | 30–86 | 44–94 | 49–84 | - |
| BMI | 23.23 ± 2.32 | 23.63 ± 2.37 | 23.81 ± 2.01 | 0.8560 |
| Age (Mean ± SD) | 56.1 ± 20.89 | 71.71 ± 12.55 | 72.38 ± 14.94 | 0.0955 |
| Protopathy disease [n (%)] | ||||
| Respiratory diseases | - | 0.75 | 0.6 | 0.833 |
| Digestive diseases | - | 0.25 | 0.4 | - |
| Hospital day, HOD (Mean ± SD) | - | 59.71 ± 24.11 | 98.38 ± 34.16 | 0.0269 |
| PaO2/FiO2, P/F (Mean ± SD) | - | 281.6 ± 49.3 | 198.5 ± 42.17 | 0.0038 |
3.3. Association between EZH2 and the IME
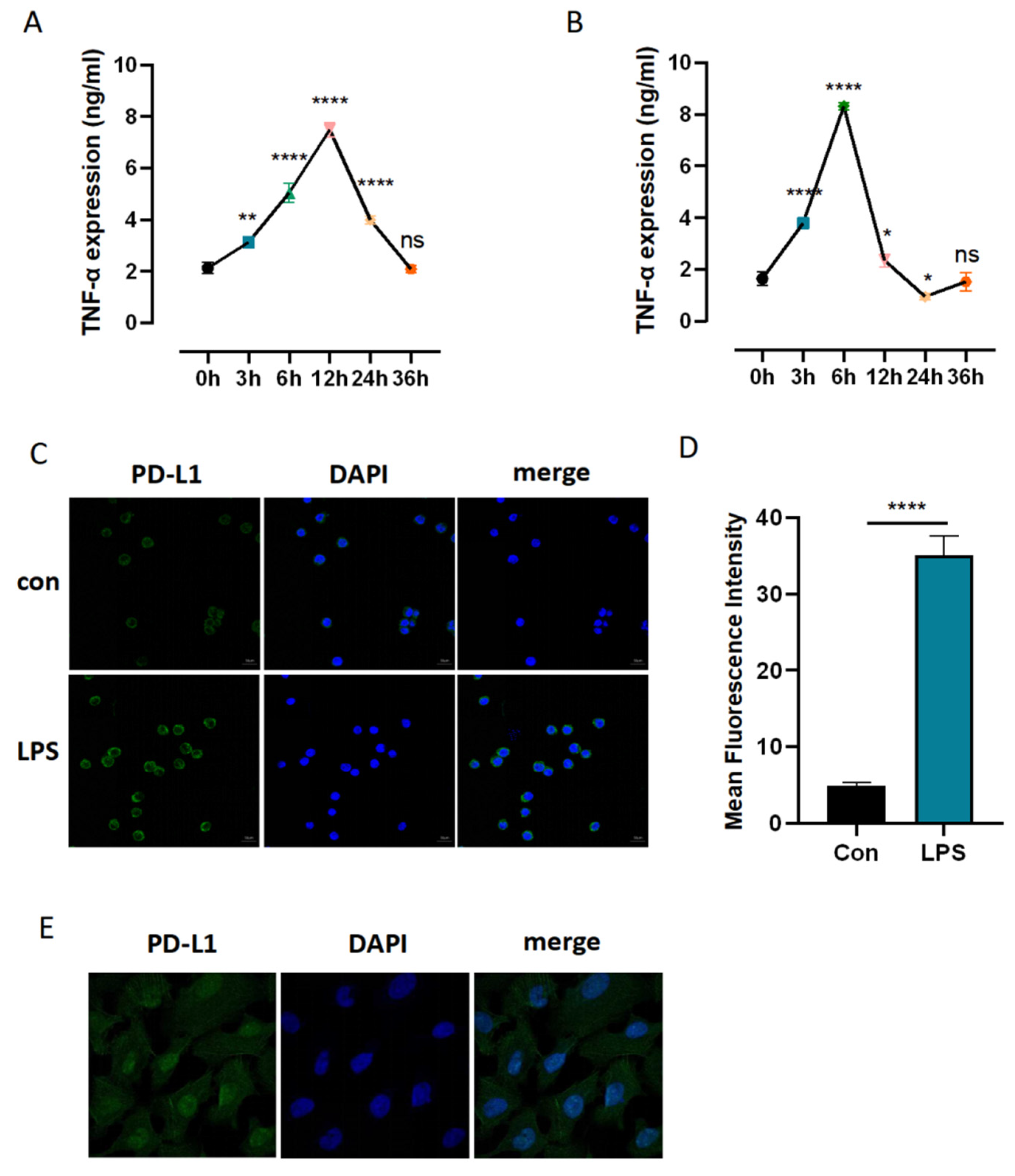
3.4. Identification of Sufficient Concentrations and Time Points of LPS Challenge
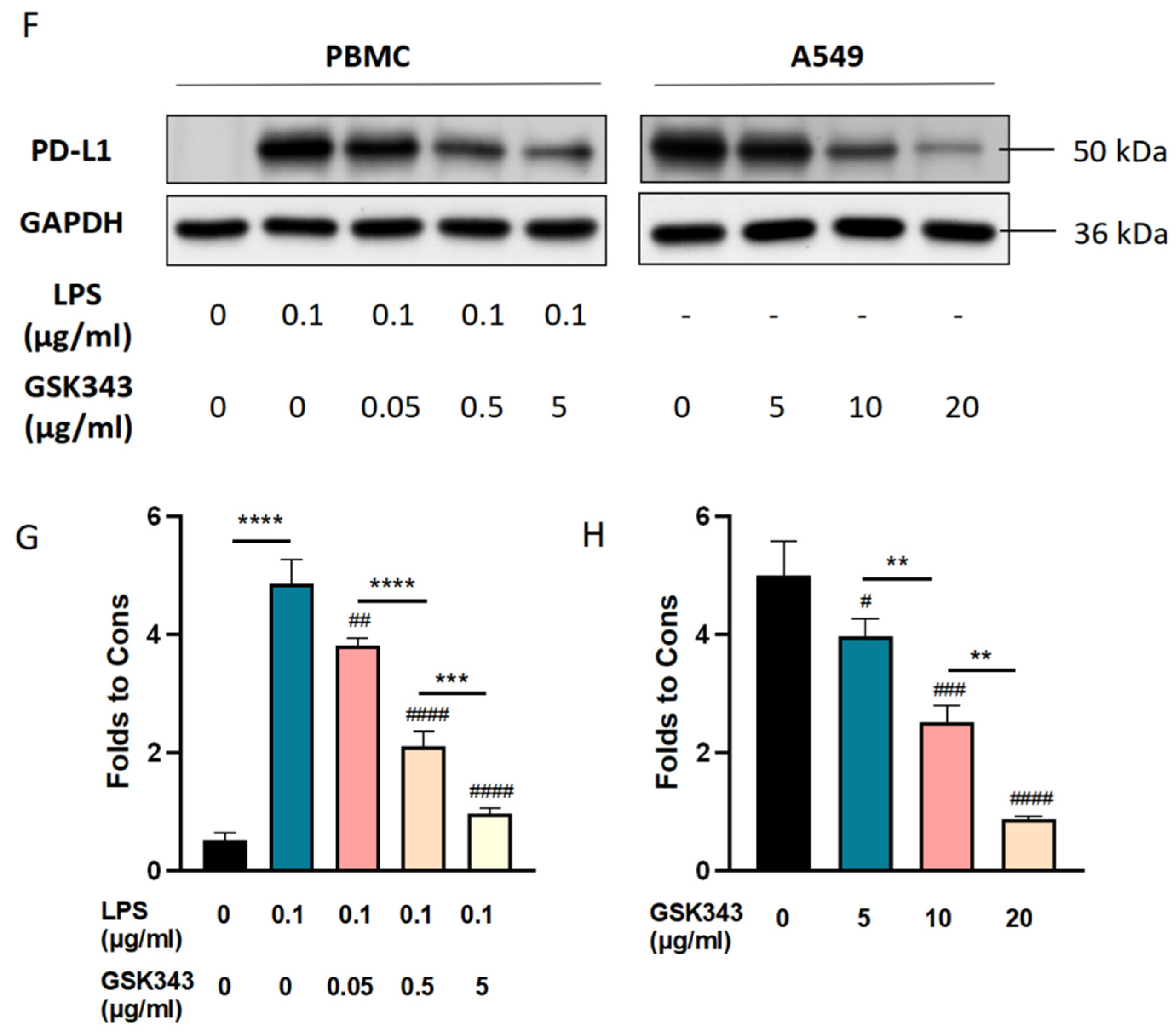
3.5. Inhibition of EZH2 Suppressed PD-L1 Expression in LPS-Induced PBMCs and A549
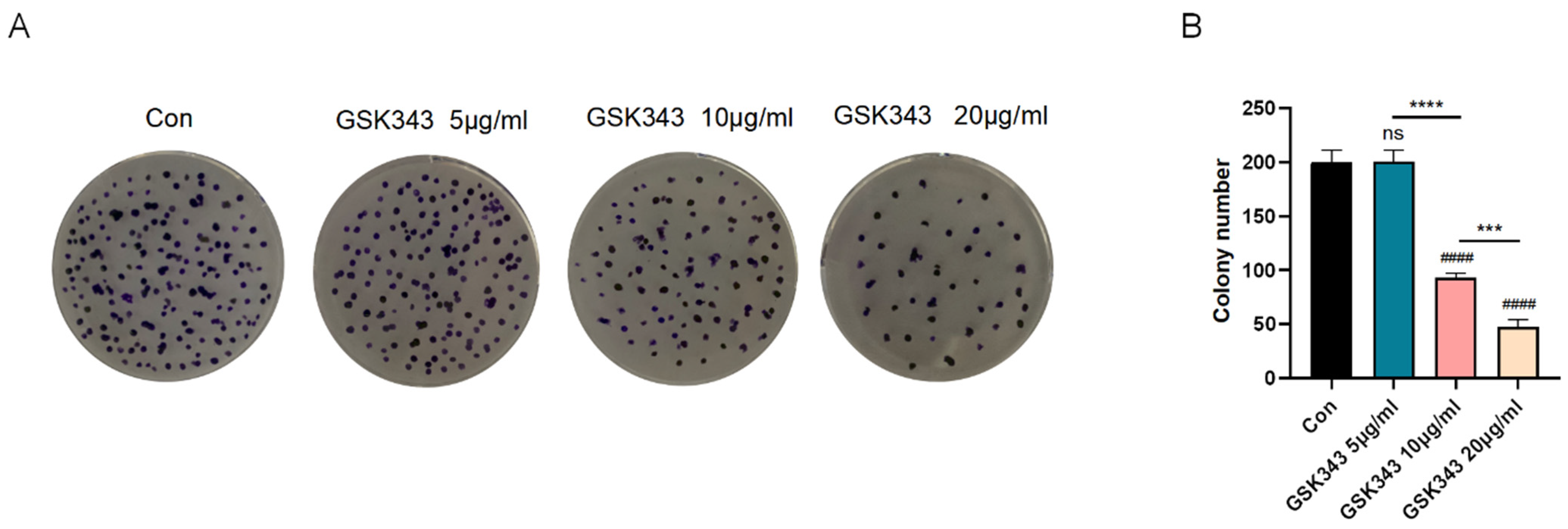
3.6. Inhibition of EZH2 Ameliorated A549 Proliferation
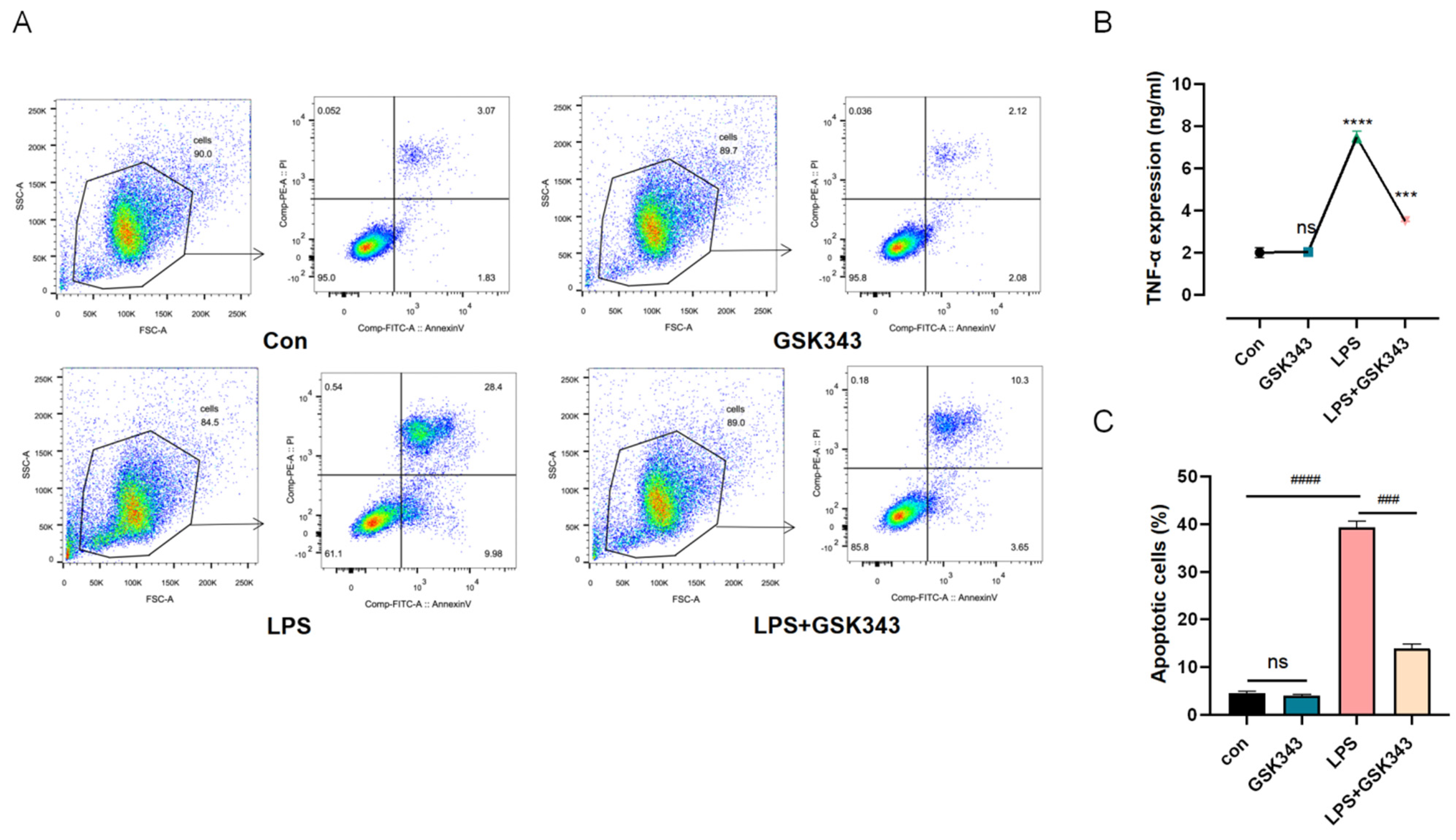
3.7. Inhibition of EZH2 Ameliorated LPS-Induced PBMC Apoptosis
4. Discussion
5. Conclusions
6. Limitations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, L.; Zhao, Y.; Lai, D.; Zhang, P.; Yang, Y.; Li, Y.; Fei, K.; Jiang, G.; Fan, J. Neutrophil extracellular traps promote macrophage pyroptosis in sepsis. Cell Death Dis. 2018, 9, 597. [Google Scholar] [CrossRef]
- Zhang, H.; Zhou, Y.; Qu, M.; Yu, Y.; Chen, Z.; Zhu, S.; Guo, K.; Chen, W.; Miao, C. Tissue Factor-Enriched Neutrophil Extracellular Traps Promote Immunothrombosis and Disease Progression in Sepsis-Induced Lung Injury. Front. Cell Infect. Microbiol. 2021, 11, 677902. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wei, J.; Zhu, H.; Cao, Y.; Yu, X.; Chen, Y.; Lv, C.; Zhao, X. Consensus of Chinese experts on early prevention and blocking of sepsis. Zhonghua Wei Zhong Bing Ji Jiu Yi Xue 2020, 32, 518–530. [Google Scholar]
- Chen, Z.; Fillmore, C.M.; Hammerman, P.S.; Kim, C.F.; Wong, K. Non-small-cell lung cancers: A heterogeneous set of diseases. Nat. Rev. Cancer 2014, 14, 535–546. [Google Scholar] [CrossRef] [PubMed]
- Altorki, N.K.; Markowitz, G.J.; Gao, D.; Port, J.L.; Saxena, A.; Stiles, B.; McGraw, T.; Mittal, V. The lung microenvironment: An important regulator of tumour growth and metastasis. Nat. Rev. Cancer 2019, 19, 9–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, C.; Wang, J.; Wang, B.; Cheng, Y.; Wang, Z.; Han, B.; Lu, Y.; Wu, G.; Zhang, L.; Song, Y.; et al. Chinese Experts Consensus on Immune Checkpoint Inhibitors for Non-small Cell Lung Cancer (2020 Version). Zhongguo Fei Ai Za Zhi 2021, 24, 217–235. [Google Scholar]
- Wang, H.; Zhang, Z.; Ma, Y.; Jia, Y.; Ma, B.; Gu, J.; Chen, O.; Yue, S. Construction of Severe Eosinophilic Asthma Related Competing Endogenous RNA Network by Weighted Gene Co-Expression Network Analysis. Front. Pharmacol. 2022, 13, 852536. [Google Scholar] [CrossRef]
- Shen, Z.; Liu, S.; Liu, J.; Liu, J.; Yao, C. Weighted Gene Co-Expression Network Analysis and Treatment Strategies of Tumor Recurrence-Associated Hub Genes in Lung Adenocarcinoma. Front. Gene 2021, 12, 756235. [Google Scholar] [CrossRef]
- Zhang, T.; Yuan, K.; Wang, Y.; Xu, M.; Cai, S.; Chen, C.; Ma, J. Identification of Candidate Biomarkers and Prognostic Analysis in Colorectal Cancer Liver Metastases. Front. Oncol. 2021, 11, 652354. [Google Scholar] [CrossRef]
- Chen, A.X.; Simpson, S.Q.; Pallin, D.J. Sepsis Guidelines. N. Engl. J. Med. 2019, 380, 1369–1371. [Google Scholar] [CrossRef]
- Chinese Society of Critical Care Medicine; Chinese Medical Association. Guidelines for management of acute lung injury/acute respiratory distress syndrome: An evidence-based update by the Chinese Society of Critical Care Medicine (2006). Zhongguo Wei Zhong Bing Ji Jiu Yi Xue 2006, 18, 706–710. [Google Scholar]
- Duan, R.; Du, W.; Guo, W. EZH2: A novel target for cancer treatment. J. Hematol. Oncol. 2020, 13, 104. [Google Scholar] [CrossRef]
- Xiao, G.; Jin, L.; Liu, C.; Wang, Y.; Meng, Y.; Zhou, Z.; Chen, J.; Yu, X.; Zhang, Y.; Xu, J.; et al. EZH2 negatively regulates PD-L1 expression in hepatocellular carcinoma. J. Immun. Cancer 2019, 7, 300. [Google Scholar] [CrossRef]
- Zang, X.; Gu, J.; Zhang, J.; Shi, H.; Hou, S.; Xu, X.; Chen, Y.; Zhang, Y.; Mao, F.; Qian, H.; et al. Exosome-transmitted lncRNA UFC1 promotes non-small-cell lung cancer progression by EZH2-mediated epigenetic silencing of PTEN expression. Cell Death Dis. 2020, 11, 215. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, M.E.; Moore, H.M.; Li, X.; Toy, K.A.; Huang, W.; Sabel, M.S.; Kidwell, K.M.; Kleer, C.G. EZH2 expands breast stem cells through activation of NOTCH1 signaling. Proc. Natl. Acad. Sci. USA 2014, 111, 3098–3103. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Zhou, X.; Tang, L. Effects of histone methyltransferase inhibitor on the polarization of peritoneal macrophages in septic mice. Zhonghua Wei Zhong Bing Ji Jiu Yi Xue 2019, 31, 187–192. [Google Scholar]
- Zhen, H.; Hu, H.; Rong, G.; Huang, X.; Tan, C.; Yu, X. VitA or VitD ameliorates bronchopulmonary dysplasia by regulating the balance between M1 and M2 macrophages. Biomed. Pharm. 2021, 141, 111836. [Google Scholar] [CrossRef] [PubMed]
- Jiang, R.; Xu, J.; Zhang, Y.; Zhu, X.; Liu, J.; Tan, Y. Ligustrazine Alleviate Acute Lung Injury Through Suppressing Pyroptosis and Apoptosis of Alveolar Macrophages. Front. Pharmacol. 2021, 12, 680512. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhao, D.; Zhou, X.; Tang, L. Effects and molecular mechanism of histone methyltransferase enhancer of zeste homolog 2 on regulating sepsis-induced T cell dysfunction. Zhonghua Wei Zhong Bing Ji Jiu Yi Xue 2022, 34, 357–361. [Google Scholar] [PubMed]
- Yue, D.; Wang, Z.; Yang, Y.; Hu, Z.; Luo, G.; Wang, F. EZH2 inhibitor GSK343 inhibits sepsis-induced intestinal disorders. Exp. Ther. Med. 2021, 21, 437. [Google Scholar] [CrossRef]
- Bah, I.; Alkhateeb, T.; Youssef, D.; Yao, Z.Q.; McCall, C.E.; El Gazzar, M. KDM6A Lysine Demethylase Directs Epigenetic Polarity of MDSCs during Murine Sepsis. J. Innate. Immun. 2022, 14, 112–123. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Sun, H.; Zhuang, S.; Liu, N.; Bao, X.; Liu, X.; Ren, H.; Lv, D.; Li, Z.; Bai, J.; et al. Novel pharmacological inhibition of EZH2 attenuates septic shock by altering innate inflammatory responses to sepsis. Int. Immun. 2019, 76, 105899. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Rayile, A.; Zhang, X.; Li, Y.; Zhao, Q. Ulinastatin protects against lipopolysaccharide-induced cardiac microvascular endothelial cell dysfunction via downregulation of lncRNA MALAT1 and EZH2 in sepsis. Int. J. Mol. Med. 2017, 39, 1269–1276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nova, Z.; Skovierova, H.; Calkovska, A. Alveolar-Capillary Membrane-Related Pulmonary Cells as a Target in Endotoxin-Induced Acute Lung Injury. Int. J. Mol. Sci. 2019, 20, 831. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, W. Tim3 regulates the ability of macrophages to counter lipopolysaccharideinduced pulmonary epithelial barrier dysfunction via the PI3K/Akt pathway in epithelial cells. Mol. Med. Rep. 2020, 22, 534–542. [Google Scholar] [CrossRef]
- Arneth, B. Tumor Microenvironment. Medicina 2019, 56, 15. [Google Scholar] [CrossRef] [Green Version]
- Xia, C.; Dong, X.; Li, H.; Cao, M.; Sun, D.; He, S.; Yang, F.; Yan, X.; Zhang, S.; Li, N.; et al. Cancer statistics in China and United States, 2022: Profiles, trends, and determinants. Chin. Med. J. 2022, 135, 584–590. [Google Scholar] [CrossRef]
- Pelly, V.S.; Moeini, A.; Roelofsen, L.M.; Bonavita, E.; Bell, C.R.; Hutton, C.; Blanco-Gomez, A.; Banyard, A.; Bromley, C.P.; Flanagan, E.; et al. Anti-Inflammatory Drugs Remodel the Tumor Immune Environment to Enhance Immune Checkpoint Blockade Efficacy. Cancer Dis. 2021, 11, 2602–2619. [Google Scholar] [CrossRef]
- Nutt, S.L.; Keenan, C.; Chopin, M.; Allan, R.S. EZH2 function in immune cell development. Biol. Chem. 2020, 401, 933–943. [Google Scholar] [CrossRef]
- Kang, N.; Eccleston, M.; Clermont, P.; Latarani, M.; Male, D.K.; Wang, Y.; Crea, F. EZH2 inhibition: A promising strategy to prevent cancer immune editing. Epigenomics 2020, 12, 1457–1476. [Google Scholar] [CrossRef]
- Yin, Y.; Qiu, S.; Li, X.; Huang, B.; Xu, Y.; Peng, Y. EZH2 suppression in glioblastoma shifts microglia toward M1 phenotype in tumor microenvironment. J. Neuroinflammation 2017, 14, 220. [Google Scholar] [CrossRef] [Green Version]
- Bugide, S.; Gupta, R.; Green, M.R.; Wajapeyee, N. EZH2 inhibits NK cell-mediated antitumor immunity by suppressing CXCL10 expression in an HDAC10-dependent manner. Proc. Natl. Acad. Sci. USA 2021, 118, e2102718118. [Google Scholar] [CrossRef]
- Hu, Z.; Chen, Y.; He, J.; Liu, H.; Hui, T. EZH2 Might Affect Macrophage Chemotaxis and Anti-Inflammatory Factors by Regulating CCL2 in Dental Pulp Inflammation. Stem. Cells Int. 2021, 2021, 3060480. [Google Scholar] [CrossRef] [PubMed]
- Bisserier, M.; Wajapeyee, N. Mechanisms of resistance to EZH2 inhibitors in diffuse large B-cell lymphomas. Blood 2018, 131, 2125–2137. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Pan, X.; Zhang, W.; Guo, H.; Cheng, S.; He, Q.; Yang, B.; Ding, L. Epigenetic strategies synergize with PD-L1/PD-1 targeted cancer immunotherapies to enhance antitumor responses. Acta Pharm. Sin. B 2020, 10, 723–733. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, Y.; Xie, J.; Zhao, Z.; Gupta, S.; Guo, Y.; Jia, S.; Parodo, J.; Marshall, J.C.; Deng, X. Upregulated PD-L1 delays human neutrophil apoptosis and promotes lung injury in an experimental mouse model of sepsis. Blood 2021, 138, 806–810. [Google Scholar] [CrossRef]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Guo, Z.; Wang, X.; Liao, H.; Chai, Y.; Wang, Z.; Wang, Z. Inhibition of EZH2 Ameliorates Sepsis Acute Lung Injury (SALI) and Non-Small-Cell Lung Cancer (NSCLC) Proliferation through the PD-L1 Pathway. Cells 2022, 11, 3958. https://doi.org/10.3390/cells11243958
Wang Z, Guo Z, Wang X, Liao H, Chai Y, Wang Z, Wang Z. Inhibition of EZH2 Ameliorates Sepsis Acute Lung Injury (SALI) and Non-Small-Cell Lung Cancer (NSCLC) Proliferation through the PD-L1 Pathway. Cells. 2022; 11(24):3958. https://doi.org/10.3390/cells11243958
Chicago/Turabian StyleWang, Ziyi, Zhe Guo, Xuesong Wang, Haiyan Liao, Yan Chai, Ziwen Wang, and Zhong Wang. 2022. "Inhibition of EZH2 Ameliorates Sepsis Acute Lung Injury (SALI) and Non-Small-Cell Lung Cancer (NSCLC) Proliferation through the PD-L1 Pathway" Cells 11, no. 24: 3958. https://doi.org/10.3390/cells11243958