
Mechanical Force Modulates Alveolar Bone Marrow Mesenchymal Cells Characteristics for Bone Remodeling during Orthodontic Tooth Movement through Lactate Production

Abstract
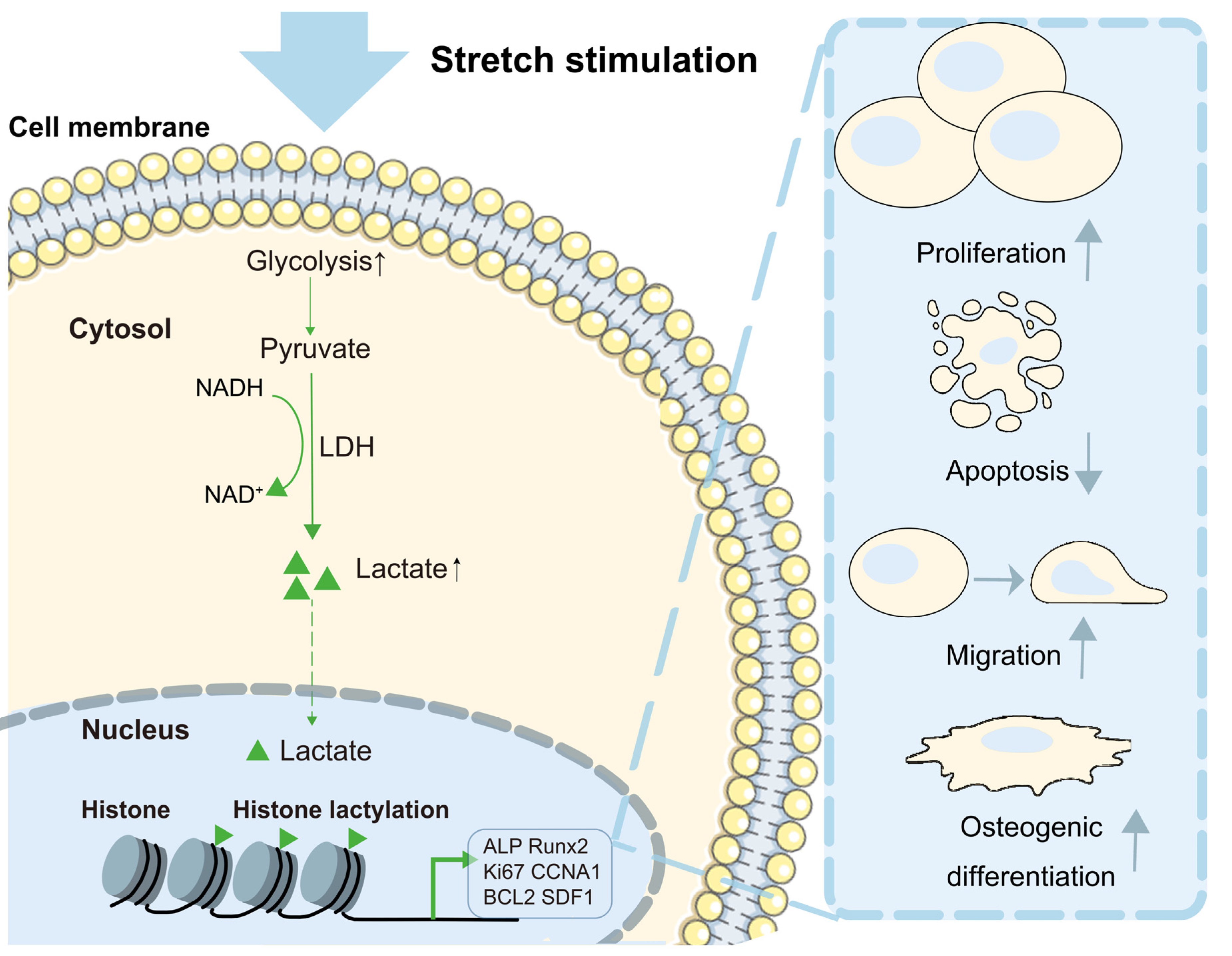
:1. Introduction
2. Materials and Methods
2.1. Animals and Ethic Statement
2.2. Immunohistochemistry (IHC)
2.3. Isolation and Identification of ABMMCs
2.4. Osteogenic Differentiation and Adipogenic Differentiation of ABMMCs
2.5. Application of Cyclic Mechanical Stretch
2.6. Lactate Assay
2.7. Cell Cycle Assessment
2.8. Hoechst 33,258 Staining
2.9. Annexin V-FITC/PI Assay
2.10. RNA Extraction and Real-Time Polymerase Chain Reaction
2.11. Western Blot (WB)
2.12. GNE-140 Application
2.13. Pyruvate Detection
2.14. Micro-CT Scanning
2.15. Statistical Analysis
3. Results
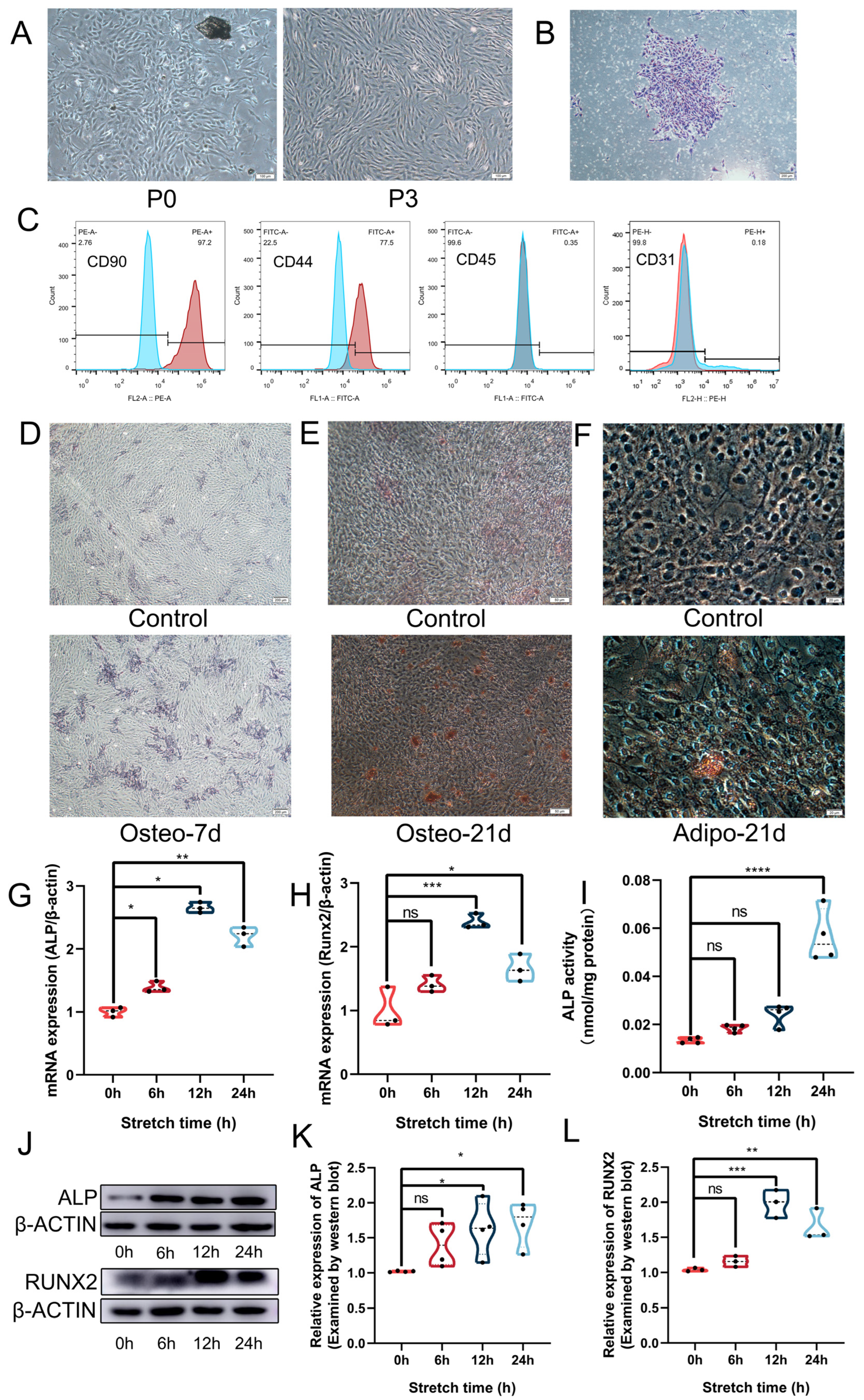
3.1. Isolated ABMMCs Maintained Stem Cell Properties and Cyclic Stretch Promoted Their Osteogenic Differentiation
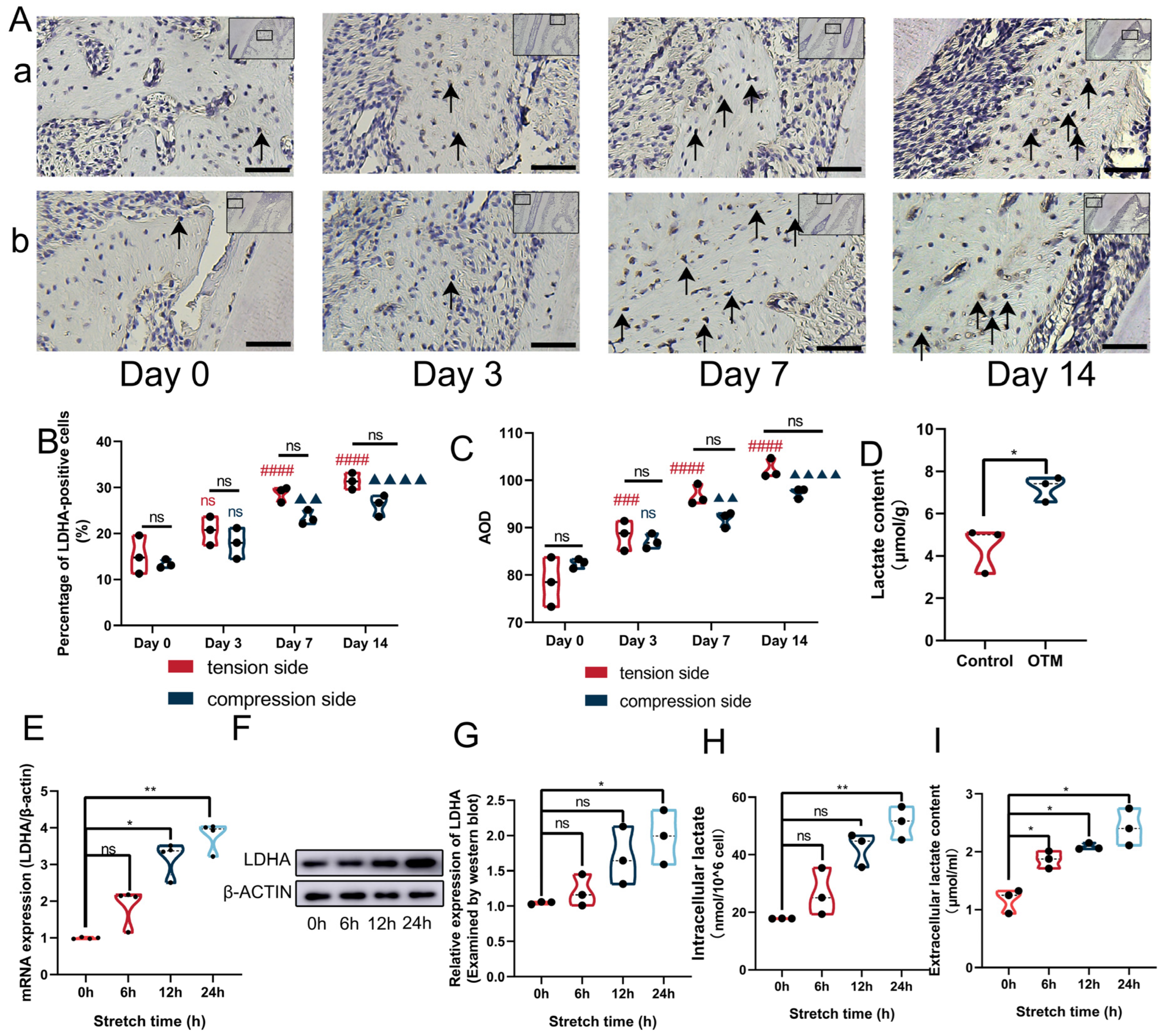
3.2. Mechanical Force Increased the Expression of LDHA and Lactate Production
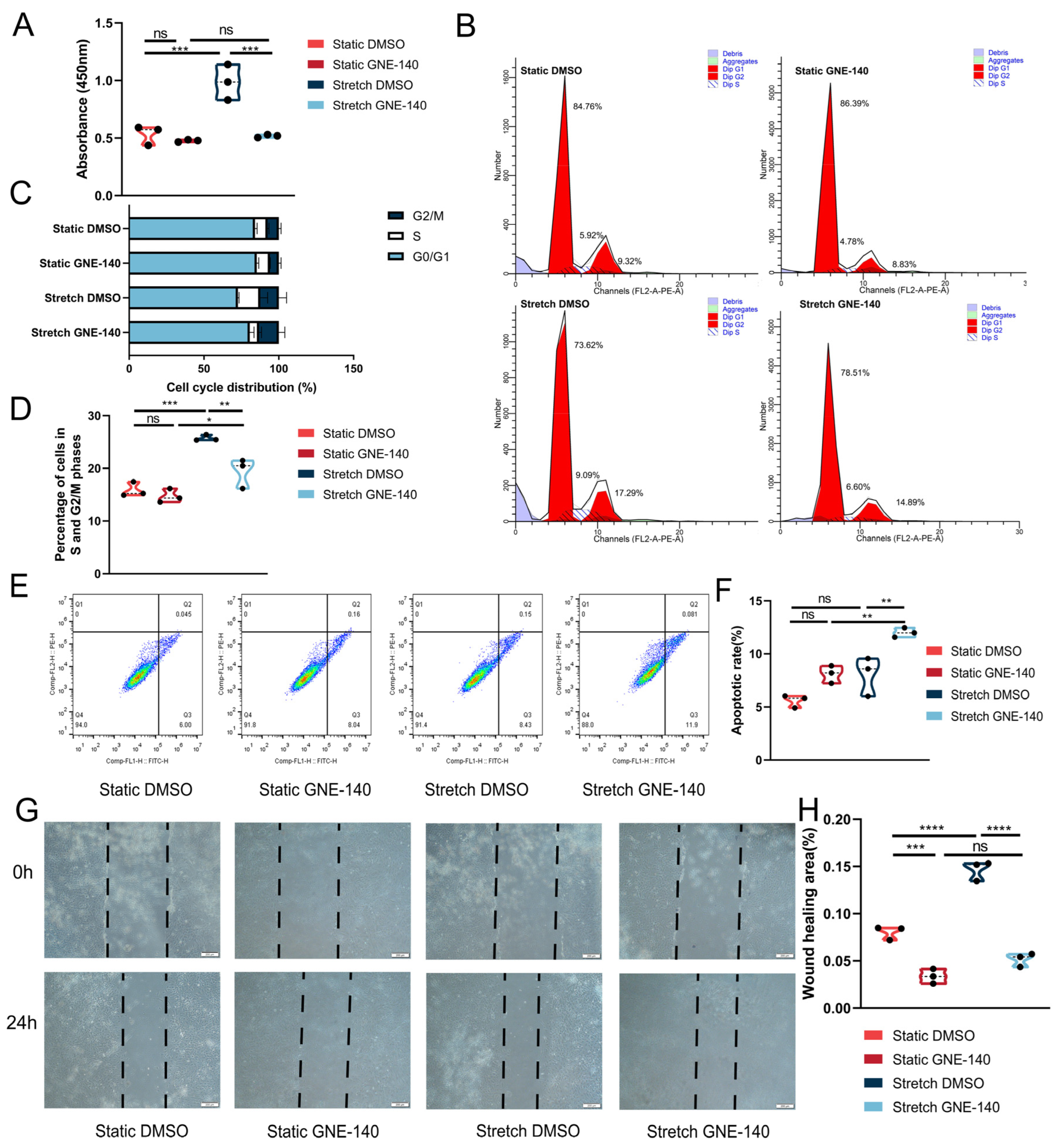
3.3. GNE-140 Inhibited the Osteogenic Differentiation of ABMMCs under Cyclic Mechanical Stretch
3.4. GNE-140 Suppressed the Proliferation and Migration Yet Promoted Apoptosis of ABMMCs under Cyclic Mechanical Stretch
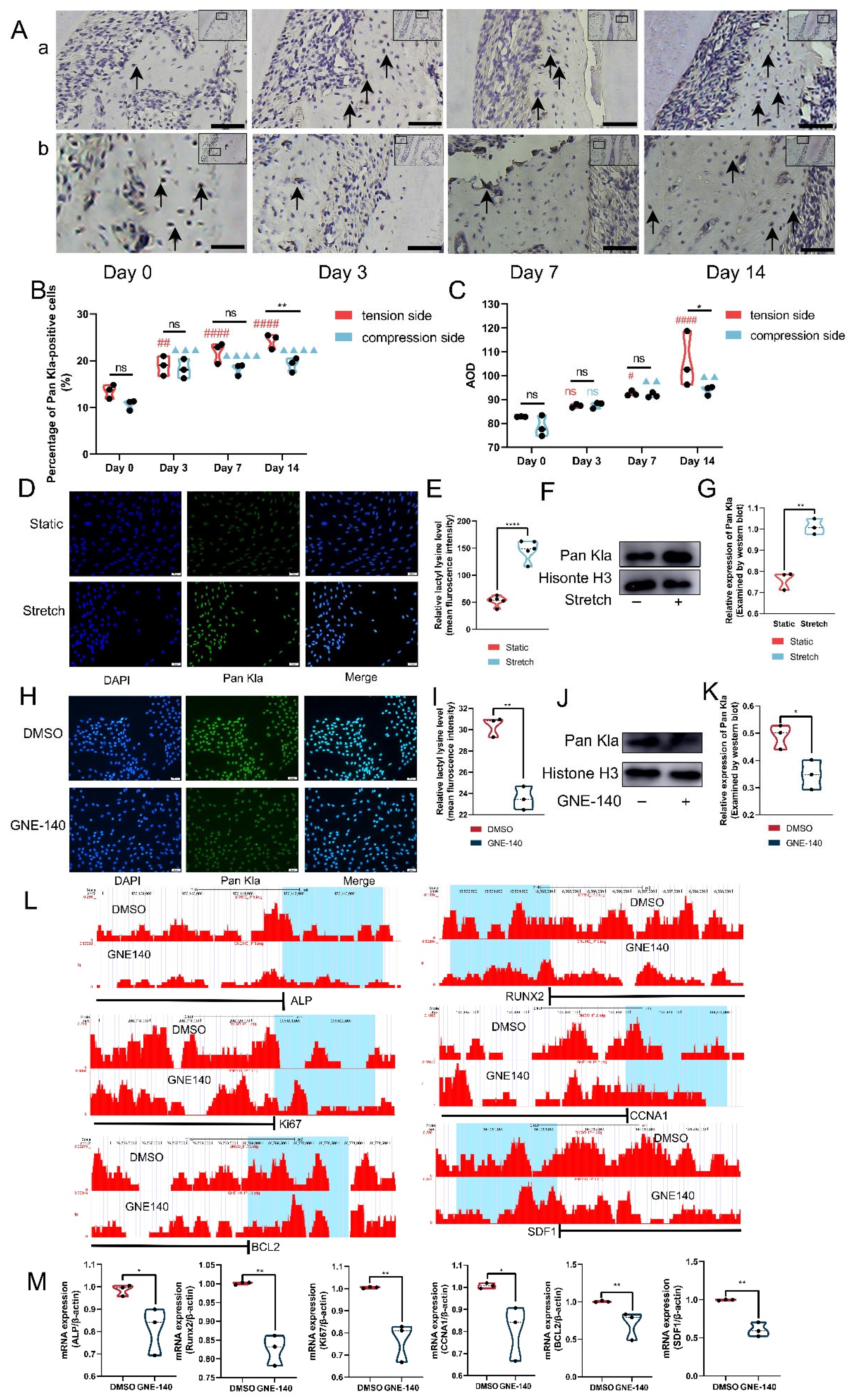
3.5. Histone Lactylation Mediated the Regulatory Effects of Lactate on ABMMCs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, Y.; Jacox, L.A.; Coats, S.; Kwon, J.; Xue, P.; Tang, N.; Rui, Z.; Wang, X.; Kim, Y.I.; Wu, T.J.; et al. Roles of autophagy in orthodontic tooth movement. Am. J. Orthod. Dentofac. 2021, 159, 582–593. [Google Scholar] [CrossRef] [PubMed]
- Jin, A.; Hong, Y.; Yang, Y.; Xu, H.; Huang, X.; Gao, X.; Gong, X.; Dai, Q.; Jiang, L. FOXO3 Mediates Tooth Movement by Regulating Force-Induced Osteogenesis. J. Dent. Res. 2022, 101, 196–205. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Liu, Y.; Ding, W.; Shi, J.; Li, S.; Liu, Y.; Wu, M.; Wang, H. Mechanical stretch-induced osteogenic differentiation of human jaw bone marrow mesenchymal stem cells (hJBMMSCs) via inhibition of the NF-κB pathway. Cell Death Dis. 2018, 9, 207. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Yuan, Y.; Wu, W.; Lei, L.; Zhang, L. The effects of locomotion on bone marrow mesenchymal stem cell fate: Insight into mechanical regulation and bone formation. Cell Biosci. 2021, 11, 88. [Google Scholar] [CrossRef] [PubMed]
- Brockhaus, J.; Craveiro, R.B.; Azraq, I.; Niederau, C.; Schröder, S.K.; Weiskirchen, R.; Jankowski, J.; Wolf, M. In Vitro Compression Model for Orthodontic Tooth Movement Modulates Human Periodontal Ligament Fibroblast Proliferation, Apoptosis and Cell Cycle. Biomolecules 2021, 11, 932. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.; Lin, H.; Fu, H.; Wang, J.; Yang, Y.; Wan, Z.; Han, G. CREB activation affects mesenchymal stem cell migration and differentiation in periodontal tissues due to orthodontic force. Int. J. Biochem. Cell Biol. 2020, 129, 105862. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Liang, H.; Sun, B.; Mi, J.; Tong, X.; Wang, Y.; Chen, M.; Yu, L.; Pan, J.; Liu, S.; et al. Role of TRPC6 in periodontal tissue reconstruction mediated by appropriate stress. Stem Cell Res. Ther. 2022, 13, 401. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Yu, W.; Liu, Y.; Yuan, X.; Yuan, R.; Guo, Q. Expression of HIF-1α in cycling stretch-induced osteogenic differentiation of bone mesenchymal stem cells. Mol. Med. Rep. 2019, 20, 4489–4498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, L.; Hu, G.; Karner, C.M. Bioenergetic Metabolism In Osteoblast Differentiation. Curr. Osteoporos. Rep. 2022, 20, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Karner, C.M.; Long, F. Glucose metabolism in bone. Bone 2018, 115, 2–7. [Google Scholar] [CrossRef] [PubMed]
- Borle, A.B.; Nichols, N.; Nichols, G., Jr. Metabolic studies of bone in vitro. I. Normal bone. J. Biol. Chem. 1960, 235, 1206–1210. [Google Scholar] [CrossRef] [PubMed]
- Dirckx, N.; Moorer, M.C.; Clemens, T.L.; Riddle, R.C. The role of osteoblasts in energy homeostasis. Nat. Rev. Endocrinol. 2019, 15, 651–665. [Google Scholar] [CrossRef] [PubMed]
- Park-Min, K.H. Metabolic reprogramming in osteoclasts. Semin. Immunopathol. 2019, 41, 565–572. [Google Scholar] [CrossRef]
- Wu, D.; Huang, R.T.; Hamanaka, R.B.; Krause, M.; Oh, M.J.; Kuo, C.H.; Nigdelioglu, R.; Meliton, A.Y.; Witt, L.; Dai, G.; et al. HIF-1α is required for disturbed flow-induced metabolic reprogramming in human and porcine vascular endothelium. eLife 2017, 6, e25217. [Google Scholar] [CrossRef]
- Park, J.S.; Burckhardt, C.J.; Lazcano, R.; Solis, L.M.; Isogai, T.; Li, L.; Chen, C.S.; Gao, B.; Minna, J.D.; Bachoo, R.; et al. Mechanical regulation of glycolysis via cytoskeleton architecture. Nature 2020, 578, 621–626. [Google Scholar] [CrossRef] [PubMed]
- Wilches-Buitrago, L.; Viacava, P.R.; Cunha, F.Q.; Alves-Filho, J.C.; Fukada, S.Y. Fructose 1,6-bisphosphate inhibits osteoclastogenesis by attenuating RANKL-induced NF-κB/NFATc-1. Agents Actions 2019, 68, 415–421. [Google Scholar] [CrossRef]
- Moriguchi, N.; Hinoi, E.; Tsuchihashi, Y.; Fujimori, S.; Iemata, M.; Takarada, T.; Yoneda, Y. Cytoprotection by pyruvate through an anti-oxidative mechanism in cultured rat calvarial osteoblasts. Histol. Histopathol. 2006, 21, 969–977. [Google Scholar] [CrossRef]
- Magistretti, P.J.; Allaman, I. Lactate in the brain: From metabolic end-product to signalling molecule. Nat. Rev. Neurosci. 2018, 19, 235–249. [Google Scholar] [CrossRef]
- Sun, S.; Li, H.; Chen, J.; Qian, Q. Lactic Acid: No Longer an Inert and end-Product of Glycolysis. Physiology 2017, 32, 453–463. [Google Scholar] [CrossRef] [Green Version]
- Haas, R.; Cucchi, D.; Smith, J.; Pucino, V.; Macdougall, C.E.; Mauro, C. Intermediates of Metabolism: From Bystanders to Signalling Molecules. Trends Biochem. Sci. 2016, 41, 460–471. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, M.; Feng, H.; Peng, Y.; Sun, J.; Qu, X.; Li, C. Lactate induces osteoblast differentiation by stabilization of HIF1α. Mol. Cell. Endocrinol. 2017, 452, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Gou, H.; Chen, X.; Li, L.; Wang, X.; Xu, Y. Lactate inhibits osteogenic differentiation of human periodontal ligament stem cells via autophagy through the MCT1-mTOR signaling pathway. Bone 2022, 162, 116444. [Google Scholar] [CrossRef]
- Srinivas, S.R.; Gopal, E.; Zhuang, L.; Itagaki, S.; Martin, P.M.; Fei, Y.J.; Ganapathy, V.; Prasad, P.D. Cloning and functional identification of slc5a12 as a sodium-coupled low-affinity transporter for monocarboxylates (SMCT2). Biochem. J. 2005, 392, 655–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirschhaeuser, F.; Sattler, U.G.; Mueller-Klieser, W. Lactate: A metabolic key player in cancer. Cancer Res. 2011, 71, 6921–6925. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; He, Q.; Yang, S.; Zhao, X.; Li, X.; Wei, F. Mechanical force-sensitive lncRNA SNHG8 inhibits osteogenic differentiation by regulating EZH2 in hPDLSCs. Cell. Signal. 2022, 93, 110285. [Google Scholar] [CrossRef] [PubMed]
- Yamaza, T.; Ren, G.; Akiyama, K.; Chen, C.; Shi, Y.; Shi, S. Mouse mandible contains distinctive mesenchymal stem cells. J. Dent. Res. 2011, 90, 317–324. [Google Scholar] [CrossRef]
- Gu, X.; Li, X.; Jin, Y.; Zhang, Z.; Li, M.; Liu, D.; Wei, F. CDR1as regulated by hnRNPM maintains stemness of periodontal ligament stem cells via miR-7/KLF4. J. Cell. Mol. Med. 2021, 25, 4501–4515. [Google Scholar] [CrossRef]
- Tang, Y.; Feinberg, T.; Keller, E.T.; Li, X.Y.; Weiss, S.J. Snail/Slug binding interactions with YAP/TAZ control skeletal stem cell self-renewal and differentiation. Nat. Cell Biol. 2016, 18, 917–929. [Google Scholar] [CrossRef]
- Xu, Y.; Jiang, Y.; Wang, Y.; Ren, Y.; Zhao, Z.; Wang, T.; Li, T. LINC00473 regulated apoptosis, proliferation and migration but could not reverse cell cycle arrest of human bone marrow mesenchymal stem cells induced by a high-dosage of dexamethasone. Stem Cell Res. 2020, 48, 101954. [Google Scholar] [CrossRef]
- Zhang, D.; Tang, Z.; Huang, H.; Zhou, G.; Cui, C.; Weng, Y.; Liu, W.; Kim, S.; Lee, S.; Perez-Neut, M.; et al. Metabolic regulation of gene expression by histone lactylation. Nature 2019, 574, 575–580. [Google Scholar] [CrossRef]
- Jia, X.; Su, H.; Chen, X.; Huang, Y.; Zheng, Y.; Ji, P.; Gao, C.; Gong, X.; Huang, Y.; Jiang, L.H.; et al. A critical role of the K(Ca) 3.1 channel in mechanical stretch-induced proliferation of rat bone marrow-derived mesenchymal stem cells. J. Cell. Mol. Med. 2020, 24, 3739–3744. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Luo, Q.; Chen, Z.; Sun, J.; Xu, B.; Ju, Y.; Song, G. Cyclic mechanical stretching promotes migration but inhibits invasion of rat bone marrow stromal cells. Stem Cell Res. 2015, 14, 155–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ansaryan, S.; Khayamian, M.A.; Saghafi, M.; Shalileh, S.; Nikshoar, M.S.; Abbasvandi, F.; Mahmoudi, M.; Bahrami, F.; Abdolahad, M. Stretch Induces Invasive Phenotypes in Breast Cells Due to Activation of Aerobic-Glycolysis-Related Pathways. Adv. Biosyst. 2019, 3, e1800294. [Google Scholar] [CrossRef] [PubMed]
- Wan, W.; He, C.; Du, C.; Wang, Y.; Wu, S.; Wang, T.; Zou, R. Effect of ILK on small-molecule metabolism of human periodontal ligament fibroblasts with mechanical stretching. J. Periodontal Res. 2020, 55, 229–237. [Google Scholar] [CrossRef]
- Colegio, O.R.; Chu, N.Q.; Szabo, A.L.; Chu, T.; Rhebergen, A.M.; Jairam, V.; Cyrus, N.; Brokowski, C.E.; Eisenbarth, S.C.; Phillips, G.M.; et al. Functional polarization of tumour-associated macrophages by tumour-derived lactic acid. Nature 2014, 513, 559–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haas, R.; Smith, J.; Rocher-Ros, V.; Nadkarni, S.; Montero-Melendez, T.; D’Acquisto, F.; Bland, E.J.; Bombardieri, M.; Pitzalis, C.; Perretti, M.; et al. Lactate Regulates Metabolic and Pro-inflammatory Circuits in Control of T Cell Migration and Effector Functions. PLoS Biol. 2015, 13, e1002202. [Google Scholar] [CrossRef]
- Manosalva, C.; Quiroga, J.; Hidalgo, A.I.; Alarcón, P.; Anseoleaga, N.; Hidalgo, M.A.; Burgos, R.A. Role of Lactate in Inflammatory Processes: Friend or Foe. Front. Immunol. 2021, 12, 808799. [Google Scholar] [CrossRef]
- Yang, H.; Du, L.; Zhang, Z. Potential biomarkers in septic shock besides lactate. Exp. Biol. Med. 2020, 245, 1066–1072. [Google Scholar] [CrossRef]
- Ippolito, L.; Morandi, A.; Giannoni, E.; Chiarugi, P. Lactate: A Metabolic Driver in the Tumour Landscape. Trends Biochem. Sci. 2019, 44, 153–166. [Google Scholar] [CrossRef]
- Beckert, S.; Farrahi, F.; Aslam, R.S.; Scheuenstuhl, H.; Königsrainer, A.; Hussain, M.Z.; Hunt, T.K. Lactate stimulates endothelial cell migration. Wound Repair Regen. 2006, 14, 321–324. [Google Scholar] [CrossRef]
- Goetze, K.; Walenta, S.; Ksiazkiewicz, M.; Kunz-Schughart, L.A.; Mueller-Klieser, W. Lactate enhances motility of tumor cells and inhibits monocyte migration and cytokine release. Int. J. Oncol. 2011, 39, 453–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, T.P.; Ganapathy, V. Lactate/GPR81 signaling and proton motive force in cancer: Role in angiogenesis, immune escape, nutrition, and Warburg phenomenon. Pharmacol. Ther. 2020, 206, 107451. [Google Scholar] [CrossRef]
- Wang, J.X.; Choi, S.Y.C.; Niu, X.; Kang, N.; Xue, H.; Killam, J.; Wang, Y. Lactic Acid and an Acidic Tumor Microenvironment suppress Anticancer Immunity. Int. J. Mol. Sci. 2020, 21, 8363. [Google Scholar] [CrossRef] [PubMed]
- Boudreau, A.; Purkey, H.E.; Hitz, A.; Robarge, K.; Peterson, D.; Labadie, S.; Kwong, M.; Hong, R.; Gao, M.; Del Nagro, C.; et al. Metabolic plasticity underpins innate and acquired resistance to LDHA inhibition. Nat. Chem. Biol. 2016, 12, 779–786. [Google Scholar] [CrossRef]
- Tsukamoto, S.; Shibasaki, A.; Naka, A.; Saito, H.; Iida, K. Lactate Promotes Myoblast Differentiation and Myotube Hypertrophy via a Pathway Involving MyoD In Vitro and Enhances Muscle Regeneration In Vivo. Int. J. Mol. Sci. 2018, 19, 3649. [Google Scholar] [CrossRef] [Green Version]
- Schwab, M.; Thunborg, K.; Azimzadeh, O.; von Toerne, C.; Werner, C.; Shevtsov, M.; Di Genio, T.; Zdralevic, M.; Pouyssegur, J.; Renner, K.; et al. Targeting Cancer Metabolism Breaks Radioresistance by Impairing the Stress Response. Cancers 2021, 13, 3762. [Google Scholar] [CrossRef]
- Ždralević, M.; Brand, A.; Di Ianni, L.; Dettmer, K.; Reinders, J.; Singer, K.; Peter, K.; Schnell, A.; Bruss, C.; Decking, S.M.; et al. Double genetic disruption of lactate dehydrogenases A and B is required to ablate the “Warburg effect” restricting tumor growth to oxidative metabolism. J. Biol. Chem. 2018, 293, 15947–15961. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Chen, K.; Wang, T.; Wu, Y.; Xing, G.; Chen, M.; Hao, Z.; Zhang, C.; Zhang, J.; Ma, B.; et al. Glis1 facilitates induction of pluripotency via an epigenome-metabolome-epigenome signalling cascade. Nat. Metab. 2020, 2, 882–892. [Google Scholar] [CrossRef]
- Cui, H.; Xie, N.; Banerjee, S.; Ge, J.; Jiang, D.; Dey, T.; Matthews, Q.L.; Liu, R.M.; Liu, G. Lung Myofibroblasts Promote Macrophage Profibrotic Activity through Lactate-induced Histone Lactylation. Am. J. Respir. Cell Mol. Biol. 2021, 64, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Gaffney, D.O.; Jennings, E.Q.; Anderson, C.C.; Marentette, J.O.; Shi, T.; Schou Oxvig, A.M.; Streeter, M.D.; Johannsen, M.; Spiegel, D.A.; Chapman, E.; et al. Non-enzymatic Lysine Lactoylation of Glycolytic Enzymes. Cell Chem. Biol. 2020, 27, 206–213.e206. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Forward Primers (5′–3′) | Reverse Primers (3′–5′) |
|---|---|---|
| ALP | ACAACACCAACGCTCAGGTC | GTGACCTCGTTCCCCTGAGT |
| RUNX2 | ACGAATGCACTATCCAGCCA | GCAGGTACGTGTGGTAGTGA |
| LDHA | CCTCAGCGTCCCATGTATCC | TCTGCACTCTTCTTCAGGCG |
| Ki67 | TATCCAGCTGCCTGTAGTGT | AGGAGCGGTCAATGATGGT |
| CCNA1 | GCTTGTCAGATGCTCACCCG | GCCCTCCTGTACTGCTCATTT |
| BCL2 | TGGCCTTCTTTGAGTTCGGT | GTTCCACAAAGGCATCCCAGC |
| SDF-1 | TACAGATGCCCCTGCCGATT | CTGAAGGGCACAGTTTGGAGT |
| β-ACTIN | CTCTGTGTGGATTGGTGGCT | CGCAGCTCAGTAACAGTCCG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhai, M.; Cui, S.; Li, L.; Cheng, C.; Zhang, Z.; Liu, J.; Wei, F. Mechanical Force Modulates Alveolar Bone Marrow Mesenchymal Cells Characteristics for Bone Remodeling during Orthodontic Tooth Movement through Lactate Production. Cells 2022, 11, 3724. https://doi.org/10.3390/cells11233724
Zhai M, Cui S, Li L, Cheng C, Zhang Z, Liu J, Wei F. Mechanical Force Modulates Alveolar Bone Marrow Mesenchymal Cells Characteristics for Bone Remodeling during Orthodontic Tooth Movement through Lactate Production. Cells. 2022; 11(23):3724. https://doi.org/10.3390/cells11233724
Chicago/Turabian StyleZhai, Mingrui, Shuyue Cui, Lan Li, Chen Cheng, Zijie Zhang, Jiani Liu, and Fulan Wei. 2022. "Mechanical Force Modulates Alveolar Bone Marrow Mesenchymal Cells Characteristics for Bone Remodeling during Orthodontic Tooth Movement through Lactate Production" Cells 11, no. 23: 3724. https://doi.org/10.3390/cells11233724