
Rearing in an Enriched Environment Ameliorates the ADHD-like Behaviors of Lister Hooded Rats While Suppressing Neuronal Activities in the Medial Prefrontal Cortex
 , , and
, , and
Abstract
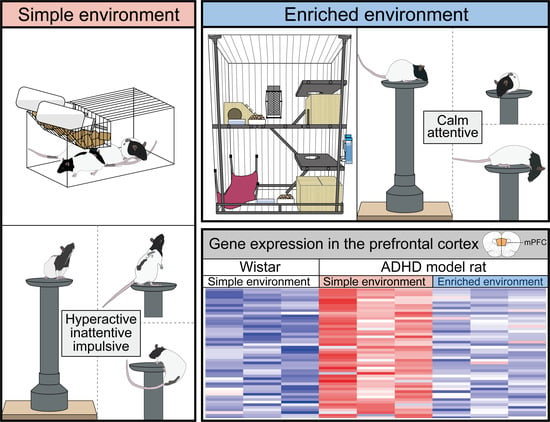
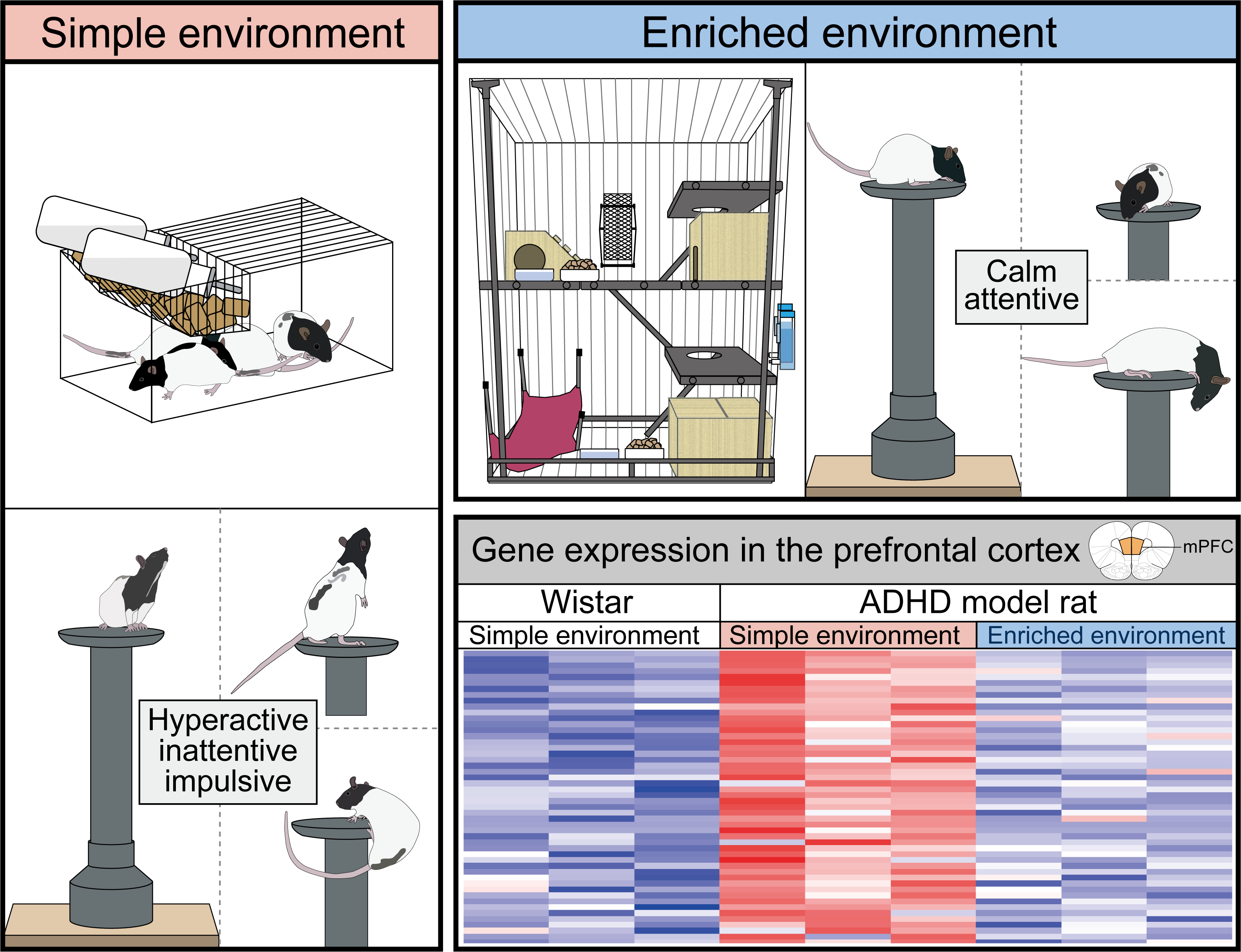
:
1. Introduction
2. Materials and Methods
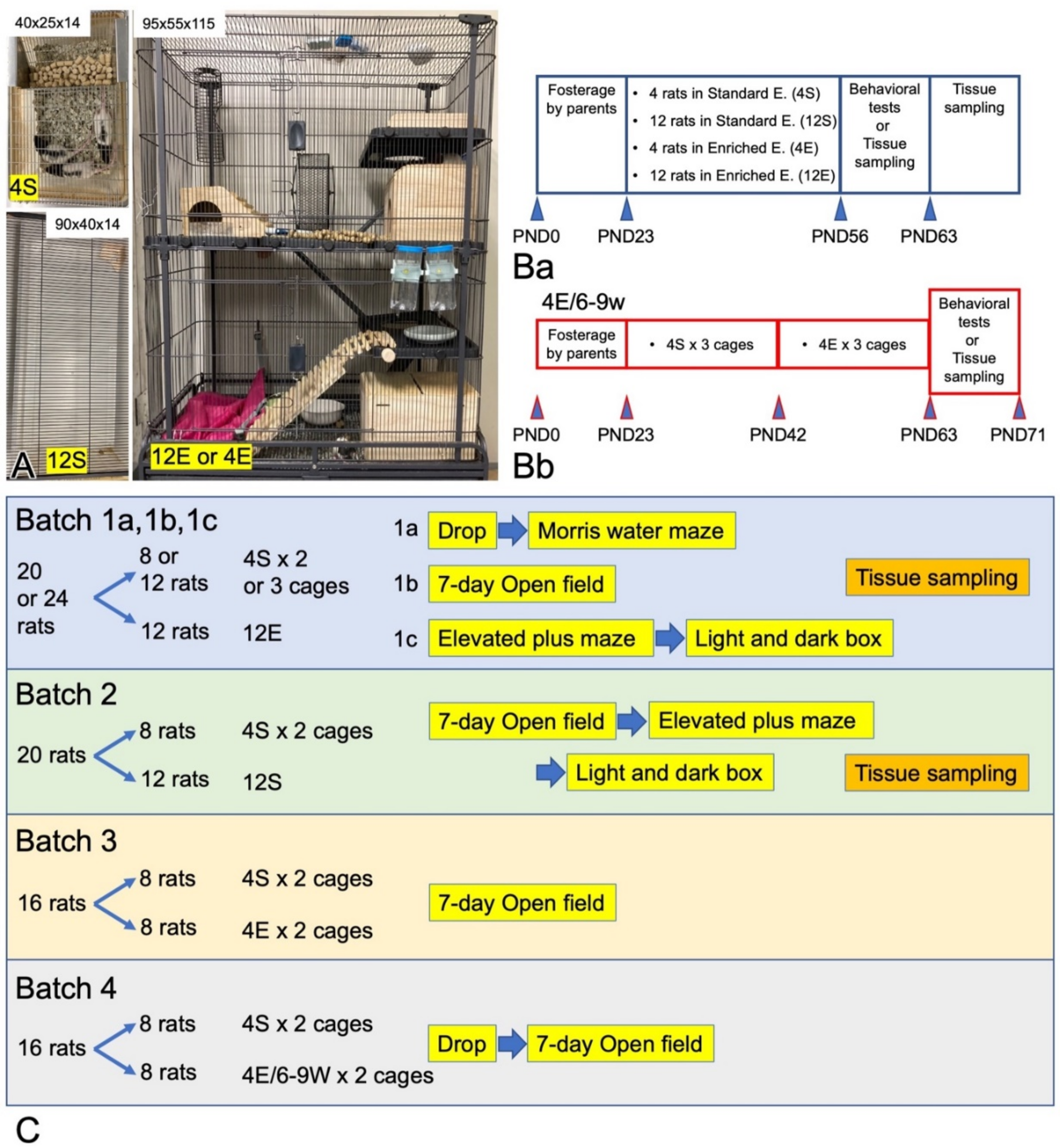
2.1. Animals
2.2. Behavioral Tests
2.2.1. OF
2.2.2. EPM
2.2.3. L/D Box
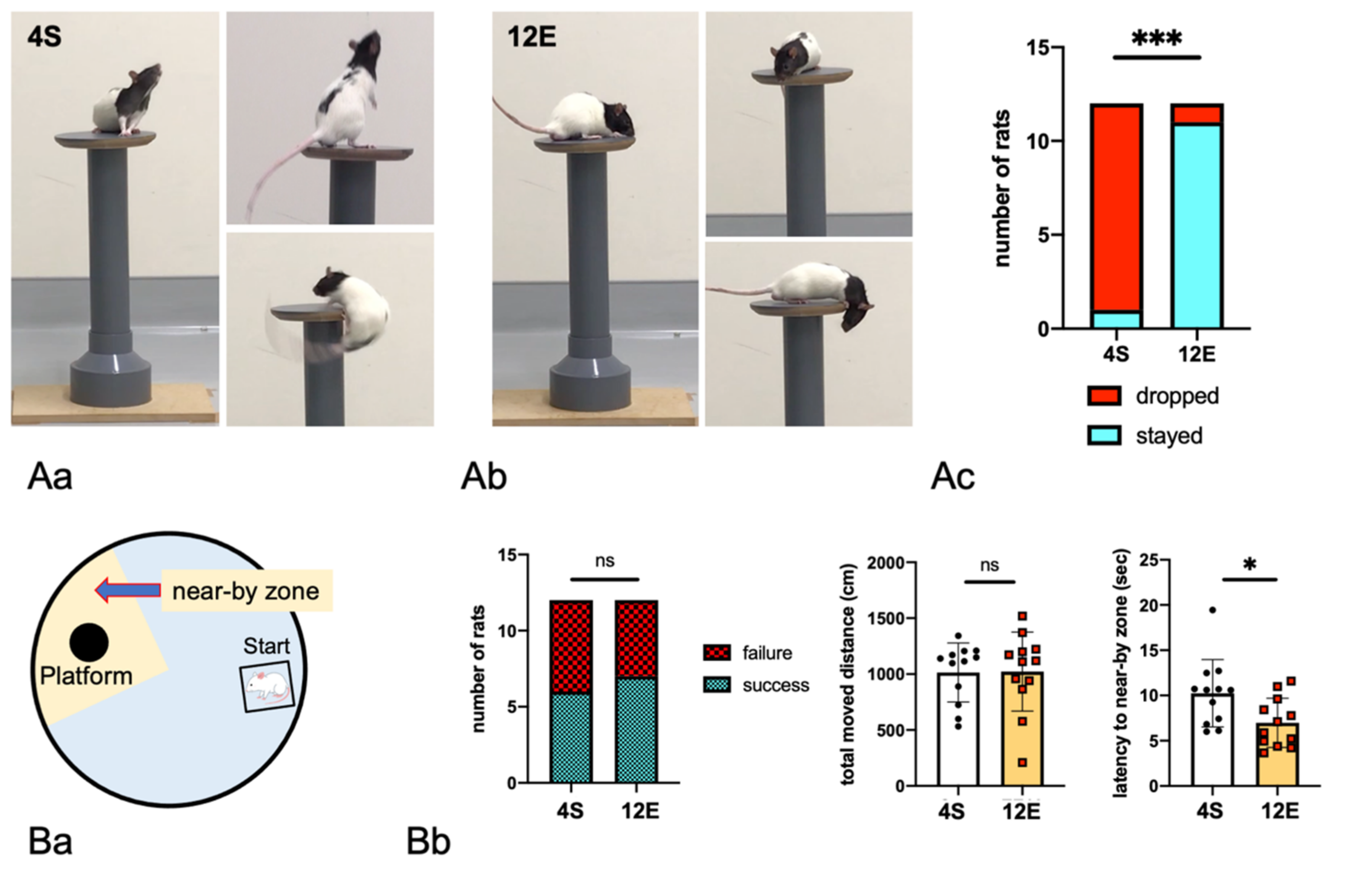
2.2.4. Drop
2.2.5. MWM
2.3. Tissue Dissection
2.4. Enzyme-Linked Immunosorbent Assay
2.5. Quantitative Real-Time RT-PCR
2.6. RNA Sequencing (RNAseq)
2.7. IHC Staining
2.8. Statistical Analysis
3. Results
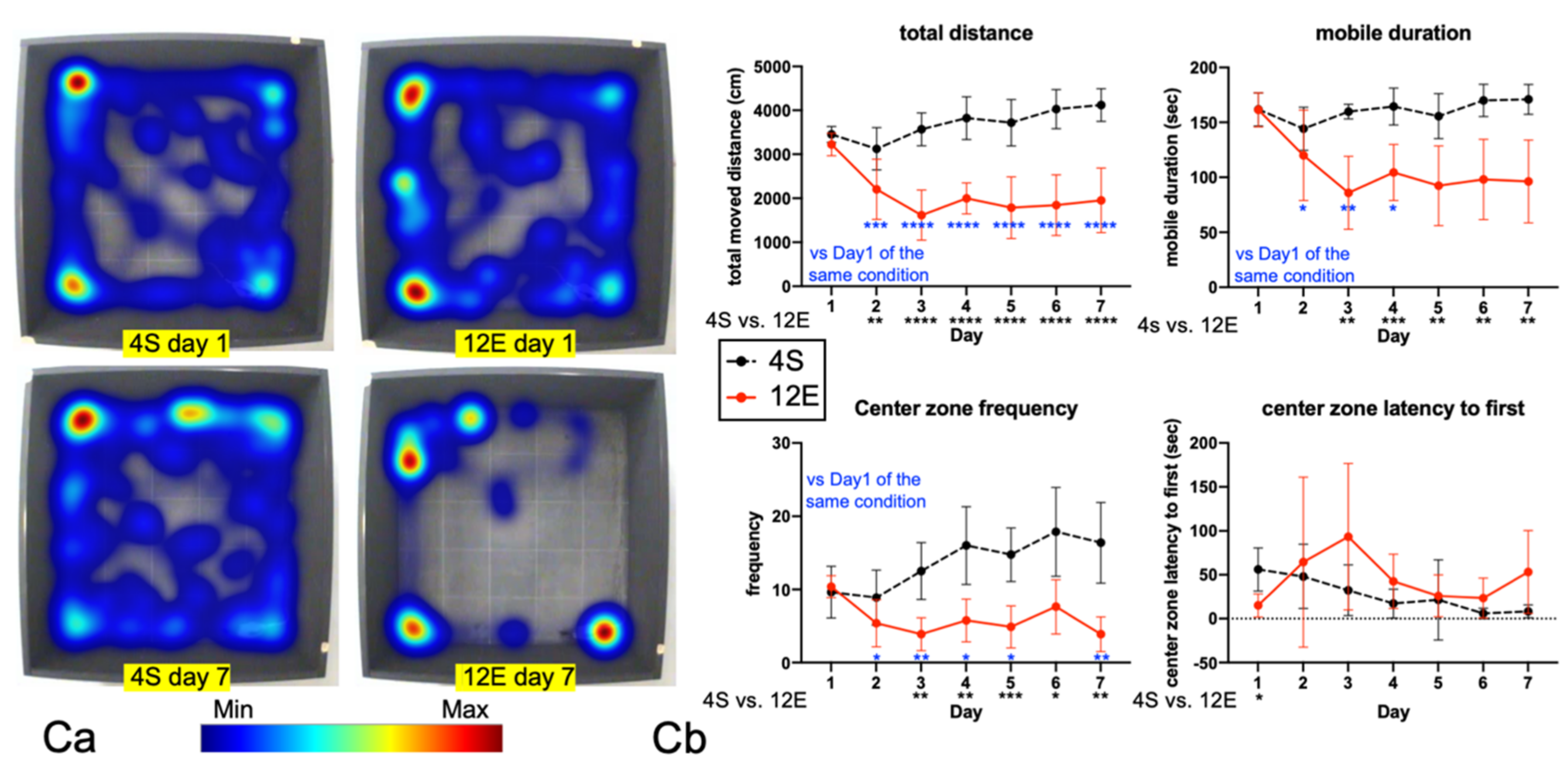
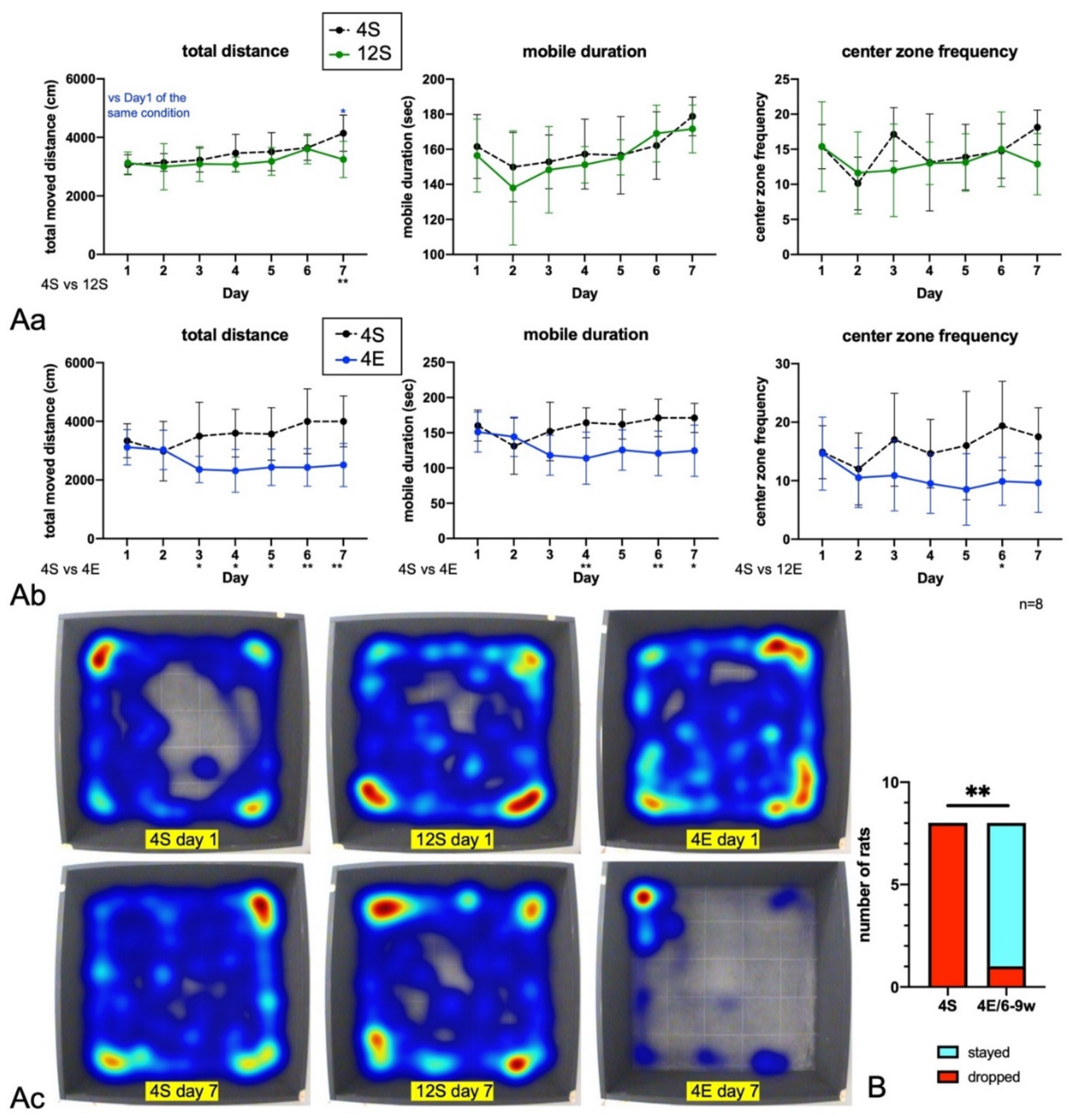
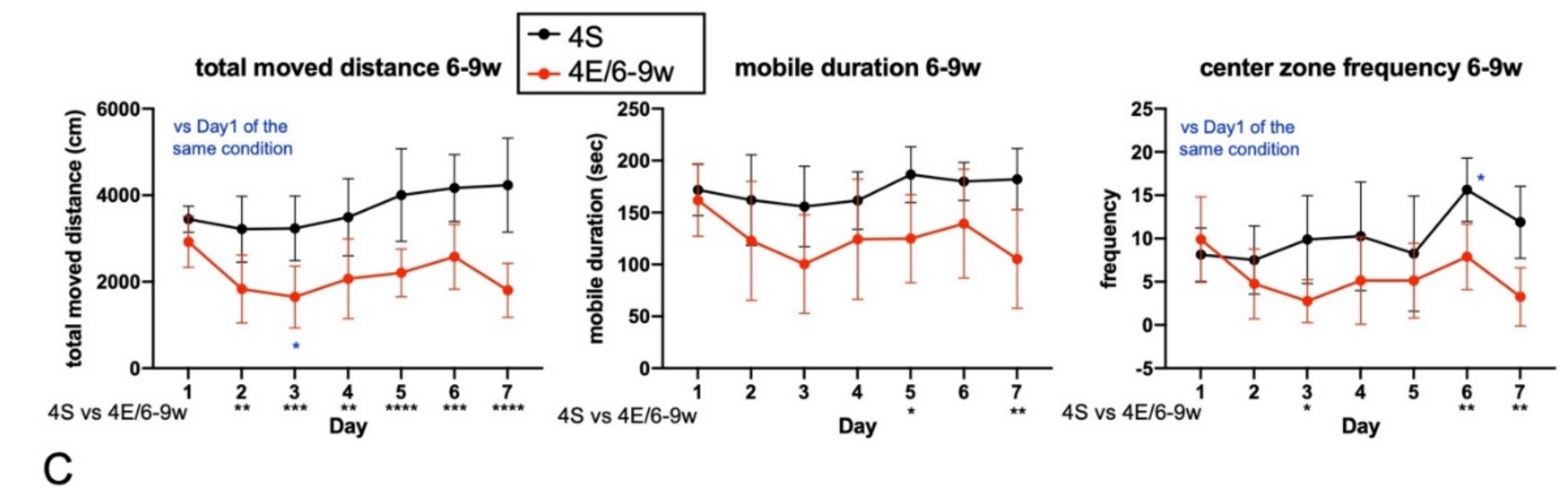
3.1. EE Rearing Ameliorated ADHD-Like Behaviors
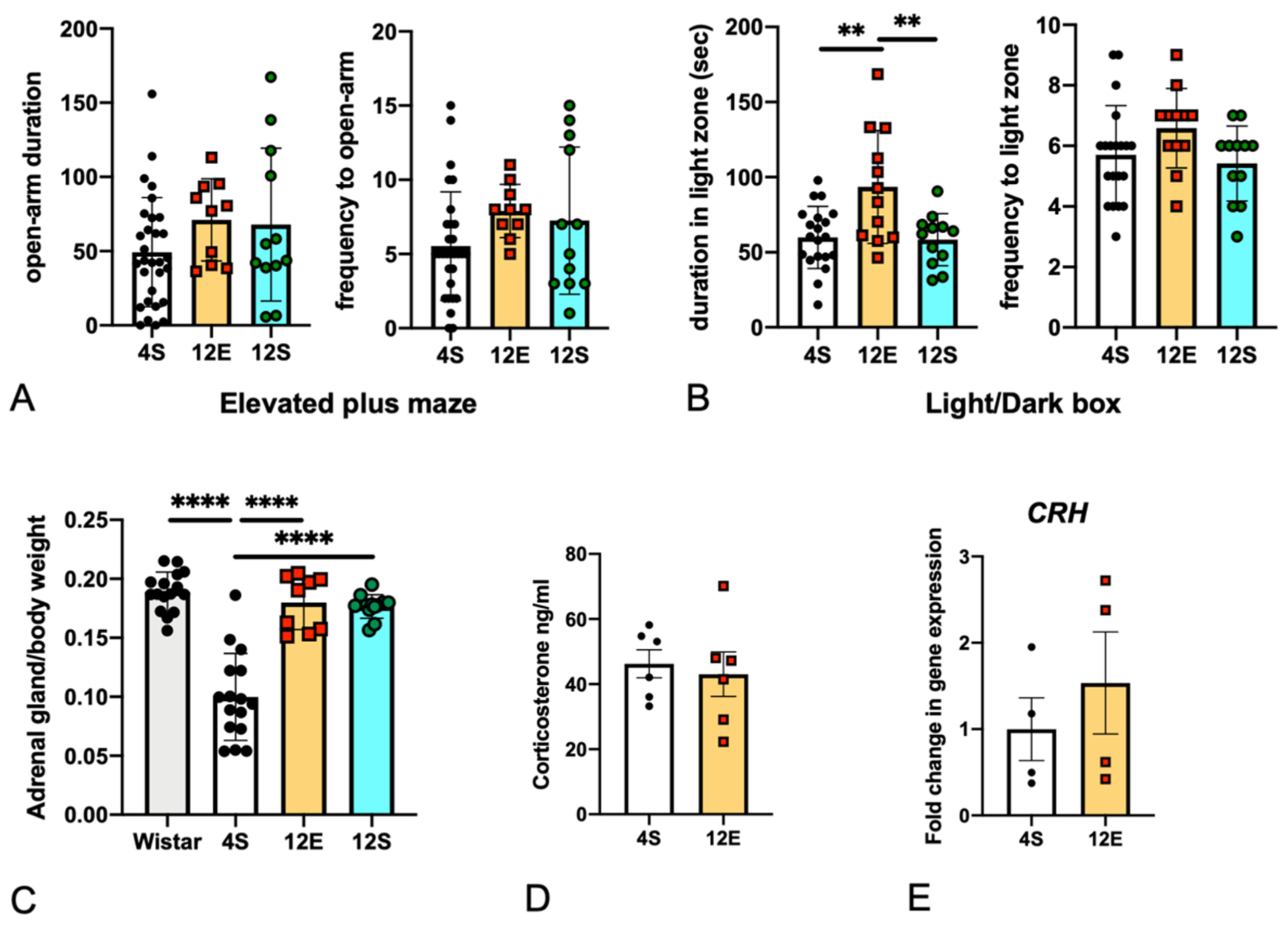
3.2. EE Rearing Did Not Increase Anxiety
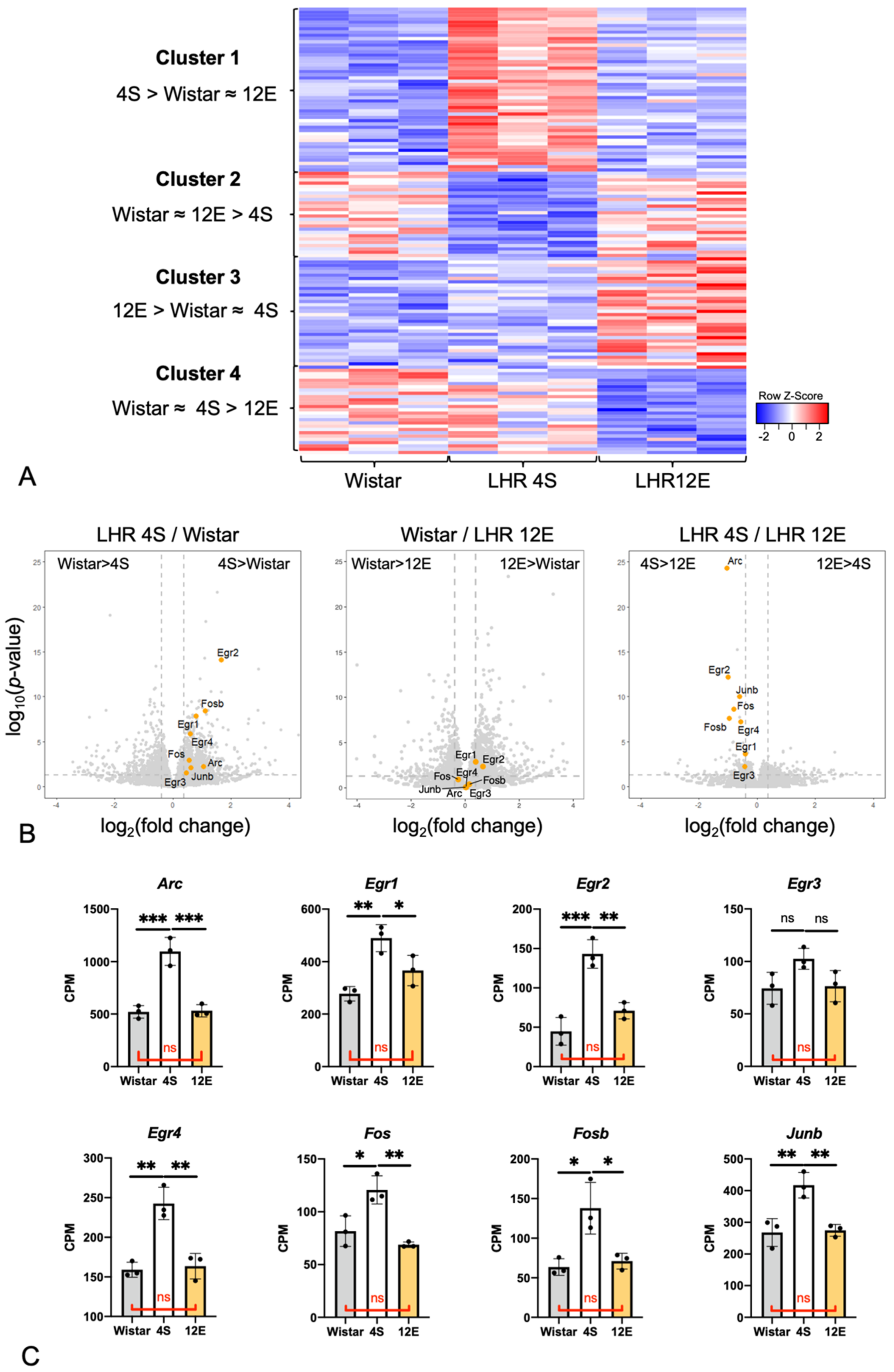
3.3. Effects of the Rearing Conditions on Gene Expression in the PrL
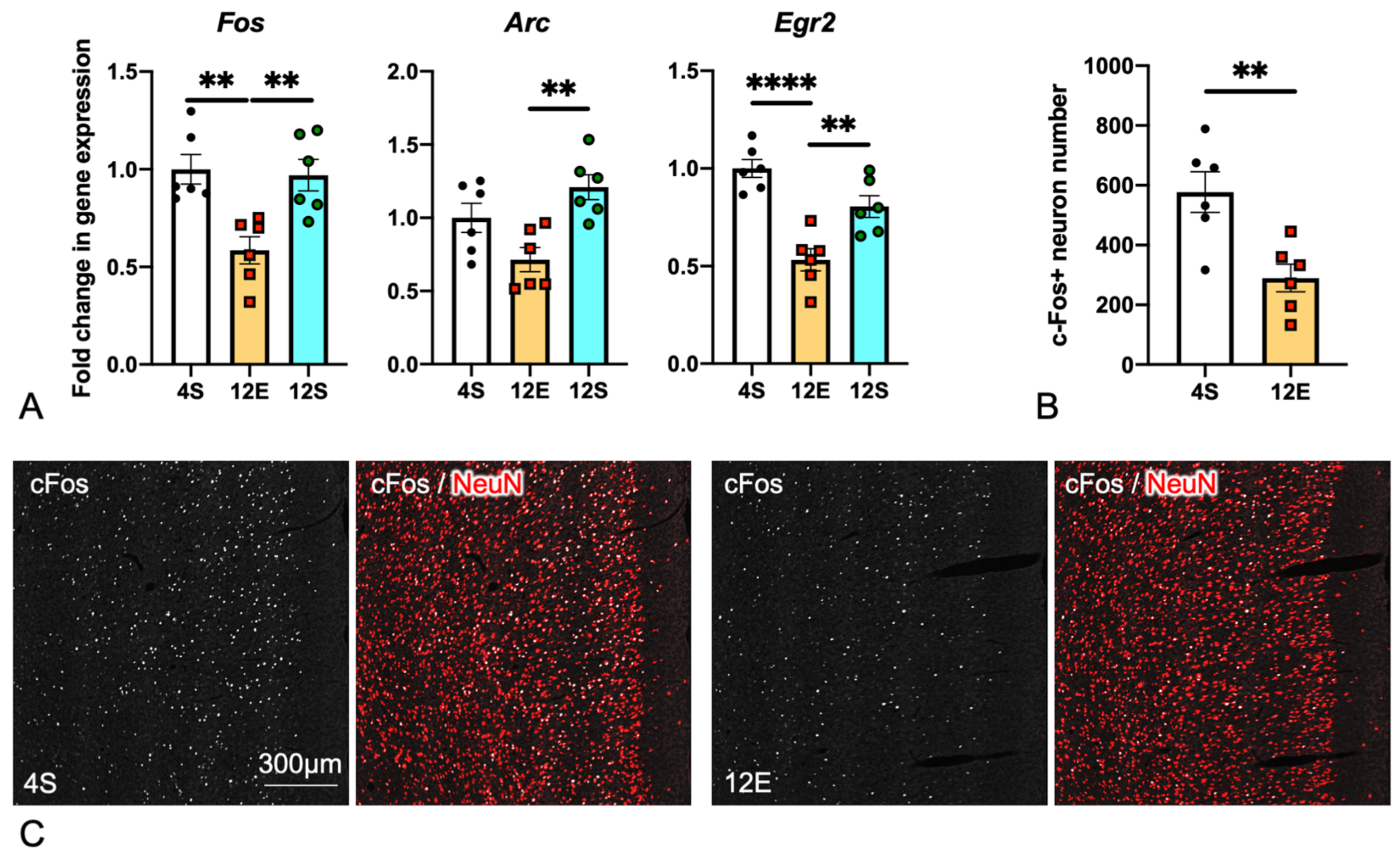
3.4. Comparison of IEG Expression in the PrL among the Three LHR Groups
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Polanczyk, G.V.; Salum, G.A.; Sugaya, L.S.; Caye, A.; Rohde, L.A. Annual research review: A meta-analysis of the worldwide prevalence of mental disorders in children and adolescents. J. Child Psychol. Psychiatry 2015, 56, 345–365. [Google Scholar] [CrossRef]
- Buitelaar, N.J.L.; Posthumus, J.A.; Buitelaar, J.K. ADHD in Childhood and/or Adulthood as a Risk Factor for Domestic Violence or Intimate Partner Violence: A Systematic Review. J. Atten. Disord. 2020, 24, 1203–1214. [Google Scholar] [CrossRef]
- Biederman, J.; Faraone, S.V. Attention-deficit hyperactivity disorder. Lancet 2005, 366, 237–248. [Google Scholar] [CrossRef]
- Faraone, S.V. The pharmacology of amphetamine and methylphenidate: Relevance to the neurobiology of attention-deficit/hyperactivity disorder and other psychiatric comorbidities. Neurosci. Biobehav. Rev. 2018, 87, 255–270. [Google Scholar] [CrossRef]
- Dark, C.; Homman-Ludiye, J.; Bryson-Richardson, R.J. The role of ADHD associated genes in neurodevelopment. Dev. Biol. 2018, 438, 69–83. [Google Scholar] [CrossRef]
- Sagvolden, T.; Russell, V.A.; Aase, H.; Johansen, E.B.; Farshbaf, M. Rodent models of attention-deficit/hyperactivity disorder. Biol. Psychiatry 2005, 57, 1239–1247. [Google Scholar] [CrossRef]
- Yamashita, M.; Sakakibara, Y.; Hall, F.; Numachi, Y.; Yoshida, S.; Kobayashi, H.; Uchiumi, O.; Uhl, G.R.; Kasahara, Y.; Sora, I. Impaired cliff avoidance reaction in dopamine transporter knockout mice. Psychopharmacology 2013, 227, 741–749. [Google Scholar] [CrossRef]
- Ng, Q.X.; Ho, C.Y.X.; Chan, H.W.; Yong, B.Z.J.; Yeo, W.-S. Managing childhood and adolescent attention-deficit/hyperactivity disorder (ADHD) with exercise: A systematic review. Complement. Ther. Med. 2017, 34, 123–128. [Google Scholar] [CrossRef]
- Arime, Y.; Kubo, Y.; Sora, I. Animal models of attention-deficit/hyperactivity disorder. Biol. Pharm. Bull. 2011, 34, 1373–1376. [Google Scholar] [CrossRef] [Green Version]
- Jogamoto, T.; Utsunomiya, R.; Sato, A.; Kihara, N.; Choudhury, M.E.; Miyanishi, K.; Kubo, M.; Nagai, M.; Nomoto, M.; Yano, H.; et al. Lister hooded rats as a novel animal model of attention-deficit/hyperactivity disorder. Neurochem. Int. 2020, 141, 104857. [Google Scholar] [CrossRef]
- Faraone, S.V.; Perlis, R.H.; Doyle, A.E.; Smoller, J.W.; Goralnick, J.J.; Holmgren, M.A.; Sklar, P. Molecular genetics of attention-deficit/hyperactivity disorder. Biol. Psychiatry 2005, 57, 1313–1323. [Google Scholar] [CrossRef] [Green Version]
- Usui, N.; Tian, X.; Harigai, W.; Togawa, S.; Utsunomiya, R.; Doi, T.; Miyoshi, K.; Shinoda, K.; Tanaka, J.; Shimada, S.; et al. Length impairments of the axon initial segment in rodent models of attention-deficit hyperactivity disorder and autism spectrum disorder. Neurochem. Int. 2022, 153, 105273. [Google Scholar] [CrossRef]
- Davidovitch, M.; Koren, G.; Fund, N.; Shrem, M.; Porath, A. Challenges in defining the rates of ADHD diagnosis and treatment: Trends over the last decade. BMC Pediatr. 2017, 17, 218. [Google Scholar] [CrossRef] [Green Version]
- Ra, C.; Cho, J.; Stone, M.D.; De La Cerda, J.; Goldenson, N.; Moroney, E.; Tung, I.; Lee, S.S.; Leventhal, A.M. Association of Digital Media Use with Subsequent Symptoms of Attention-Deficit/Hyperactivity Disorder among Adolescents. JAMA 2018, 320, 255–263. [Google Scholar] [CrossRef] [Green Version]
- Swing, E.L.; Gentile, D.A.; Anderson, C.A.; Walsh, D.A. Television and video game exposure and the development of attention problems. Pediatrics 2010, 126, 214–221. [Google Scholar] [CrossRef] [Green Version]
- André, F.; Munck, I.; Håkansson, A.; Claesdotter-Knutsson, E. Game Addiction Scale for Adolescents-Psychometric Analyses of Gaming Behavior, Gender Differences and ADHD. Front. Psychiatry 2022, 13, 791254. [Google Scholar] [CrossRef]
- Tiraboschi, G.A.; West, G.L.; Boers, E.; Bohbot, V.D.; Fitzpatrick, C. Associations between Video Game Engagement and ADHD Symptoms in Early Adolescence. J. Atten. Disord. 2022, 26, 10870547211073473. [Google Scholar] [CrossRef]
- Zheng, F.; Gao, P.; He, M.; Li, M.; Wang, C.; Zeng, Q.; Zhou, Z.; Yu, Z.; Zhang, L. Association between mobile phone use and inattention in 7102 Chinese adolescents: A population-based cross-sectional study. BMC Public Health 2014, 14, 1022. [Google Scholar] [CrossRef] [Green Version]
- Kuo, F.E.; Taylor, A.F. A potential natural treatment for attention-deficit/hyperactivity disorder: Evidence from a national study. Am. J. Public Health 2004, 94, 1580–1586. [Google Scholar] [CrossRef]
- McCormick, R. Does Access to Green Space Impact the Mental Well-being of Children: A Systematic Review. J. Pediatr. Nurs. 2017, 37, 3–7. [Google Scholar] [CrossRef]
- Vollset, S.E.; Goren, E.; Yuan, C.-W.; Cao, J.; E Smith, A.; Hsiao, T.; Bisignano, C.; Azhar, G.S.; Castro, E.; Chalek, J.; et al. Fertility, mortality, migration, and population scenarios for 195 countries and territories from 2017 to 2100: A forecasting analysis for the Global Burden of Disease Study. Lancet 2020, 396, 1285–1306. [Google Scholar] [CrossRef]
- The MTA Cooperative Group. A 14-Month Randomized Clinical Trial of Treatment Strategies for Attention-Deficit/Hyperactivity Disorder. Arch. Gen. Psychiatry 1999, 56, 1073–1086. [Google Scholar] [CrossRef]
- Ball, N.J.; Mercado, E.I., 3rd; Orduña, I. Enriched Environments as a Potential Treatment for Developmental Disorders: A Critical Assessment. Front. Psychol. 2019, 10, 466. [Google Scholar] [CrossRef] [Green Version]
- Halperin, J.M.; Healey, D.M. The influences of environmental enrichment, cognitive enhancement, and physical exercise on brain development: Can we alter the developmental trajectory of ADHD? Neurosci. Biobehav. Rev. 2011, 35, 621–634. [Google Scholar] [CrossRef] [Green Version]
- Schrijver, N.C.; I Bahr, N.; Weiss, I.C.; Würbel, H. Dissociable effects of isolation rearing and environmental enrichment on exploration, spatial learning and HPA activity in adult rats. Pharmacol. Biochem. Behav. 2002, 73, 209–224. [Google Scholar] [CrossRef]
- Neugebauer, N.M.; Cunningham, S.T.; Zhu, J.; Bryant, R.I.; Middleton, L.S.; Dwoskin, L.P. Effects of environmental enrichment on behavior and dopamine transporter function in medial prefrontal cortex in adult rats prenatally treated with cocaine. Dev. Brain Res. 2004, 153, 213–223. [Google Scholar] [CrossRef]
- Hunziker, M.H.L.; Saldana, R.L.; Neuringer, A. Behavioral variability in SHR and WKY rats as a function of rearing environment and reinforcement contingency. J. Exp. Anal. Behav. 1996, 65, 129–144. [Google Scholar] [CrossRef] [Green Version]
- Mashoodh, R.; Franks, B.; Curley, J.P.; Champagne, F.A. Paternal social enrichment effects on maternal behavior and offspring growth. Proc. Natl. Acad. Sci. USA 2012, 109, 17232–17238. [Google Scholar] [CrossRef] [Green Version]
- Miyanishi, K.; Choudhury, M.E.; Watanabe, M.; Kubo, M.; Nomoto, M.; Yano, H.; Tanaka, J. Behavioral tests predicting striatal dopamine level in a rat hemi-Parkinson’s disease model. Neurochem. Int. 2019, 122, 38–46. [Google Scholar] [CrossRef]
- Choudhury, M.E.; Mikami, K.; Nakanishi, Y.; Matsuura, T.; Utsunomiya, R.; Yano, H.; Kubo, M.; Ando, R.; Iwanami, J.; Yamashita, M.; et al. Insomnia and depressive behavior of MyD88-deficient mice: Relationships with altered microglial functions. J. Neuroimmunol. 2021, 363, 577794. [Google Scholar] [CrossRef]
- Abe, N.; Choudhury, M.E.; Watanabe, M.; Kawasaki, S.; Nishihara, T.; Yano, H.; Matsumoto, S.; Kunieda, T.; Kumon, Y.; Yorozuya, T.; et al. Comparison of the detrimental features of microglia and infiltrated macrophages in traumatic brain injury: A study using a hypnotic bromovalerylurea. Glia 2018, 66, 2158–2173. [Google Scholar] [CrossRef]
- Vorhees, C.V.; Williams, M.T. Morris water maze: Procedures for assessing spatial and related forms of learning and memory. Nat. Protoc. 2006, 1, 848–858. [Google Scholar] [CrossRef] [Green Version]
- Taguchi, S.; Choudhury, M.E.; Miyanishi, K.; Nakanishi, Y.; Kameda, K.; Abe, N.; Yano, H.; Yorozuya, T.; Tanaka, J. Aggravating effects of treadmill exercises during the early-onset period in a rat traumatic brain injury model: When should rehabilitation exercises be initiated? IBRO Rep. 2019, 7, 82–89. [Google Scholar] [CrossRef]
- Nishioka, R.; Sugimoto, K.; Aono, H.; Mise, A.; Choudhury, M.E.; Miyanishi, K.; Islam, A.; Fujita, T.; Takeda, H.; Takahashi, H.; et al. Treadmill exercise ameliorates ischemia-induced brain edema while suppressing Na+/H+ exchanger 1 expression. Exp. Neurol. 2016, 277, 150–161. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Ozaki, S.; Umakoshi, A.; Yano, H.; Ohsumi, S.; Sumida, Y.; Hayase, E.; Usa, E.; Islam, A.; Choudhury, M.E.; Nishi, Y.; et al. Chloride intracellular channel protein 2 is secreted and inhibits MMP14 activity, while preventing tumor cell invasion and metastasis. Neoplasia 2021, 23, 754–765. [Google Scholar] [CrossRef]
- Peña, C.J.; Smith, M.; Ramakrishnan, A.; Cates, H.M.; Bagot, R.C.; Kronman, H.G.; Patel, B.; Chang, A.B.; Purushothaman, I.; Dudley, J.; et al. Early life stress alters transcriptomic patterning across reward circuitry in male and female mice. Nat. Commun. 2019, 10, 5098. [Google Scholar] [CrossRef] [Green Version]
- Falkenberg, T.; Mohammed, A.K.; Henriksson, B.; Persson, H.; Winblad, B.; Lindefors, N. Increased expression of brain-derived neurotrophic factor mRNA in rat hippocampus is associated with improved spatial memory and enriched environment. Neurosci. Lett. 1992, 138, 153–156. [Google Scholar] [CrossRef]
- Nilsson, M.; Perfilieva, E.; Johansson, U.; Orwar, O.; Eriksson, P.S. Enriched environment increases neurogenesis in the adult rat dentate gyrus and improves spatial memory. J. Neurobiol. 1999, 39, 569–578. [Google Scholar] [CrossRef]
- Sainsbury, W.J.; Carrasco, K.; Whitehouse, A.J.O.; McNeil, L.; Waddington, H. Age of Diagnosis for Co-occurring Autism and Attention Deficit Hyperactivity Disorder During Childhood and Adolescence: A Systematic Review. Rev. J. Autism Dev. Disord. 2022, 1–13. [Google Scholar] [CrossRef]
- File, S.E.; Hyde, J.R.G. Can social interaction be used to measure anxiety? Br. J. Pharmacol. 1978, 62, 19–24. [Google Scholar] [CrossRef] [Green Version]
- Blanchard, D.C.; Hynd, A.L.; Minke, K.A.; Minemoto, T.; Blanchard, R.J. Human defensive behaviors to threat scenarios show parallels to fear- and anxiety-related defense patterns of non-human mammals. Neurosci. Biobehav. Rev. 2001, 25, 761–770. [Google Scholar] [CrossRef]
- Erhardt, A.; Ising, M.; Unschuld, P.G.; Kern, N.; Lucae, S.; Putz, B.; Uhr, M.; Binder, E.B.; Holsboer, F.; Keck, M.E. Regulation of the hypothalamic-pituitary-adrenocortical system in patients with panic disorder. Neuropsychopharmacology 2006, 31, 2515–2522. [Google Scholar] [CrossRef] [Green Version]
- Cubillo, A.; Halari, R.; Smith, A.; Taylor, E.; Rubia, K. A review of fronto-striatal and fronto-cortical brain abnormalities in children and adults with Attention Deficit Hyperactivity Disorder (ADHD) and new evidence for dysfunction in adults with ADHD during motivation and attention. Cortex 2011, 48, 194–215. [Google Scholar] [CrossRef]
- Hupalo, S.; Berridge, C.W. Working memory impairing actions of Corticotropin-Releasing Factor (CRF) neurotransmission in the prefrontal cortex. Neuropsychopharmacology 2016, 41, 2733–2740. [Google Scholar] [CrossRef]
- Rappley, M.D. Clinical practice. Attention deficit-hyperactivity disorder. N. Engl. J. Med. 2005, 352, 165–173. [Google Scholar] [CrossRef]
- Pamplona, F.A.; Pandolfo, P.; Savoldi, R.; Prediger, R.D.; Takahashi, R.N. Environmental enrichment improves cognitive deficits in Spontaneously Hypertensive Rats (SHR): Relevance for Attention Deficit/Hyperactivity Disorder (ADHD). Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2009, 33, 1153–1160. [Google Scholar] [CrossRef]
- Botanas, C.J.; Lee, H.; de la Pena, J.B.; Pena, I.J.D.; Woo, T.; Kim, H.J.; Han, D.H.; Kim, B.N.; Cheong, J.H. Rearing in an enriched environment attenuated hyperactivity and inattention in the Spontaneously Hypertensive Rats, an animal model of Attention-Deficit Hyperactivity Disorder. Physiol. Behav. 2016, 155, 30–37. [Google Scholar] [CrossRef]
- Bouchatta, O.; Manouze, H.; Ba-M’Hamed, S.; Landry, M.; Bennis, M. Neonatal 6-OHDA Lesion Model in Mouse Induces Cognitive Dysfunctions of Attention-Deficit/Hyperactivity Disorder (ADHD) During Young Age. Front. Behav. Neurosci. 2020, 14, 27. [Google Scholar] [CrossRef]
- Granon, S.; Passetti, F.; Thomas, K.L.; Dalley, J.W.; Everitt, B.J.; Robbins, T.W. Enhanced and impaired attentional performance after infusion of D1 dopaminergic receptor agents into rat prefrontal cortex. J. Neurosci. 2000, 20, 1208–1215. [Google Scholar] [CrossRef] [Green Version]
- Berridge, C.W.; Devilbiss, D.M.; Andrzejewski, M.E.; Arnsten, A.F.; Kelley, A.E.; Schmeichel, B.; Hamilton, C.; Spencer, R.C. Methylphenidate preferentially increases catecholamine neurotransmission within the prefrontal cortex at low doses that enhance cognitive function. Biol. Psychiatry 2006, 60, 1111–1120. [Google Scholar] [CrossRef]
- McEwen, B.S.; Bowles, N.P.; Gray, J.D.; Hill, M.N.; Hunter, R.G.; Karatsoreos, I.N.; Nasca, C. Mechanisms of stress in the brain. Nat. Neurosci. 2015, 18, 1353–1363. [Google Scholar] [CrossRef]
- Ouanes, S.; Popp, J. High Cortisol and the Risk of Dementia and Alzheimer’s Disease: A Review of the Literature. Front. Aging Neurosci. 2019, 11, 43. [Google Scholar] [CrossRef] [Green Version]
- Sandi, C.; Touyarot, K. Mid-life stress and cognitive deficits during early aging in rats: Individual differences and hippocampal correlates. Neurobiol. Aging 2006, 27, 128–140. [Google Scholar] [CrossRef]
- Desouza, L.A.; Benekareddy, M.; Fanibunda, S.E.; Mohammad, F.; Janakiraman, B.; Ghai, U.; Gur, T.; Blendy, J.A.; Vaidya, V.A. The Hallucinogenic Serotonin2A Receptor Agonist, 2,5-Dimethoxy-4-Iodoamphetamine, Promotes cAMP Response Element Binding Protein-Dependent Gene Expression of Specific Plasticity-Associated Genes in the Rodent Neocortex. Front. Mol. Neurosci. 2021, 14, 790213. [Google Scholar] [CrossRef]
- Xie, L.; Liu, Y.; Hu, Y.; Wang, B.; Zhu, Z.; Jiang, Y.; Suo, Y.; Hu, M.; Gao, J.; Ullah, R.; et al. Neonatal sevoflurane exposure induces impulsive behavioral deficit through disrupting excitatory neurons in the medial prefrontal cortex in mice. Transl. Psychiatry 2020, 10, 202. [Google Scholar] [CrossRef]
- Isaksson, J.; Nilsson, K.W.; Nyberg, F.; Hogmark, A.; Lindblad, F. Cortisol levels in children with attention-deficit/hyperactivity disorder. J. Psychiatr. Res. 2012, 46, 1398–1405. [Google Scholar] [CrossRef]
- El Ghamry, R.H.; Mohamed, M.M.; Azzam, H.M.; Elhabiby, M.M.; Hasan, H.M.; Hashish, A.F.; Elhamshary, M.M.; Barakat, D.M.H. Aggression in ADHD: Relation to salivary cortisol. Middle East Curr. Psychiatry 2021, 28, 25. [Google Scholar] [CrossRef]
- Bernhard, A.; Mayer, J.S.; Fann, N.; Freitag, C.M. Cortisol response to acute psychosocial stress in ADHD compared to conduct disorder and major depressive disorder: A systematic review. Neurosci. Biobehav. Rev. 2021, 127, 899–916. [Google Scholar] [CrossRef]
- Freitag, C.M.; Hanig, S.; Palmason, H.; Meyer, J.; Wust, S.; Seitz, C. Cortisol awakening response in healthy children and children with ADHD: Impact of comorbid disorders and psychosocial risk factors. Psychoneuroendocrinology 2009, 34, 1019–1028. [Google Scholar] [CrossRef]
- Santos-Costa, N.; Baptista-de-Souza, D.; Canto-de-Souza, L.; da Costa, V.F.; Nunes-de-Souza, R.L. Glutamatergic Neurotransmission Controls the Functional Lateralization of the mPFC in the Modulation of Anxiety Induced by Social Defeat Stress in Male Mice. Front. Behav. Neurosci. 2021, 15, 695735. [Google Scholar] [CrossRef]
- Majcher-Maslanka, I.; Solarz, A.; Wedzony, K.; Chocyk, A. Previous Early-life Stress Modifies Acute Corticosterone-induced Synaptic Plasticity in the Medial Prefrontal Cortex of Adolescent Rats. Neuroscience 2018, 379, 316–333. [Google Scholar] [CrossRef]
- Beau-Lejdstrom, R.; Douglas, I.; Evans, S.J.; Smeeth, L. Latest trends in ADHD drug prescribing patterns in children in the UK: Prevalence, incidence and persistence. BMJ Open 2016, 6, e010508. [Google Scholar] [CrossRef] [Green Version]
- Charach, A.; Ickowicz, A.; Schachar, R. Stimulant treatment over five years: Adherence, effectiveness, and adverse Effects. J. Am. Acad. Child Adolesc. Psychiatry 2004, 43, 559–567. [Google Scholar] [CrossRef]
- Graham, J.; Banaschewski, T.; Buitelaar, J.; Coghill, D.; Danckaerts, M.; Dittmann, R.W.; Dopfner, M.; Hamilton, R.; Hollis, C.; Holtmann, M.; et al. European guidelines on managing adverse effects of medication for ADHD. Eur. Child Adolesc. Psychiatry 2010, 20, 17–37. [Google Scholar] [CrossRef] [Green Version]
- Oh, Y.; Joung, Y.S.; Kim, J. Association between Attention Deficit Hyperactivity Disorder Medication and Depression: A 10-year Follow-up Self-controlled Case Study. Clin. Psychopharmacol. Neurosci. 2022, 20, 320–329. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| qPCR Primers | Fwd | Rvs |
|---|---|---|
| Arc (Activity Regulated Cytoskeleton Associated Protein) | GGCATCTGTTGACCGAAGTGT | CACATAGCCGTCCAAGTTGTTCT |
| Fos (Fos Proto-Oncogene) | AGCCGACTCCTTCTCCAGCA | AAGTTGGCACTAGAGACGGACAGAT |
| CRH (Corticotropin Releasing Hormone) | AGGGAAGTCTTGGAAATG | CCGATAATCTCCATCAGTT |
| Egr2 (Early Growth Response 2) | GATCTGCATGCGAAACTTCAG | GCAAACTTACGGCCACAATAG |
| GAPDH | GAGACAGCCGCATCTTCTTG | TGACTGTGCCGTTGAACTTG |
| Antigen | Antibody | Concentration | Source |
|---|---|---|---|
| cFos | rabbit monoclonal (9F6) | 1/500 | Cell signaling, Danvers, MA |
| NeuN | guinea pig polyclonal | 1/500 | Merck millipore, Darmstadt, Germany |
| rabbit IgG | donkey polyclonal; labelled with DyLightL 488 | 1/500 | Jackson ImmunoResearch Labs, West Grove, PA |
| guinea pig IgG | donkey polyclonal; labelled with Cy5 | 1/500 | Jackson ImmunoResearch |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Utsunomiya, R.; Mikami, K.; Doi, T.; Choudhury, M.E.; Jogamoto, T.; Tokunaga, N.; Ishii, E.; Eguchi, M.; Yano, H.; Tanaka, J. Rearing in an Enriched Environment Ameliorates the ADHD-like Behaviors of Lister Hooded Rats While Suppressing Neuronal Activities in the Medial Prefrontal Cortex. Cells 2022, 11, 3649. https://doi.org/10.3390/cells11223649
Utsunomiya R, Mikami K, Doi T, Choudhury ME, Jogamoto T, Tokunaga N, Ishii E, Eguchi M, Yano H, Tanaka J. Rearing in an Enriched Environment Ameliorates the ADHD-like Behaviors of Lister Hooded Rats While Suppressing Neuronal Activities in the Medial Prefrontal Cortex. Cells. 2022; 11(22):3649. https://doi.org/10.3390/cells11223649
Chicago/Turabian StyleUtsunomiya, Ryo, Kanta Mikami, Tomomi Doi, Mohammed E. Choudhury, Toshihiro Jogamoto, Naohito Tokunaga, Eiichi Ishii, Mariko Eguchi, Hajime Yano, and Junya Tanaka. 2022. "Rearing in an Enriched Environment Ameliorates the ADHD-like Behaviors of Lister Hooded Rats While Suppressing Neuronal Activities in the Medial Prefrontal Cortex" Cells 11, no. 22: 3649. https://doi.org/10.3390/cells11223649