
Reactive Oxygen Species in Regulating Lymphangiogenesis and Lymphatic Function
by
 , and
, and
Bhupesh Singla
1,*, 
Ravi Varma Aithabathula
1,
Sonia Kiran
1,
Shweta Kapil
2,
Santosh Kumar
1 and
and
Udai P. Singh
1 1
Department of Pharmaceutical Sciences, The University of Tennessee Health Science Center, Memphis, TN 38017, USA
2
Division of Gastroenterology, Hepatology and Nutrition, Cincinnati Children′s Hospital Medical Center, Cincinnati, OH 45229, USA
*
Author to whom correspondence should be addressed.
Cells 2022, 11(11), 1750; https://doi.org/10.3390/cells11111750
Submission received: 15 April 2022
/
Revised: 18 May 2022
/
Accepted: 24 May 2022
/
Published: 26 May 2022
(This article belongs to the Topic Cellular Redox Homeostasis)
Abstract
:The lymphatic system is pivotal for immunosurveillance and the maintenance of tissue homeostasis. Lymphangiogenesis, the formation of new lymphatic vessels from pre-existing vessels, has both physiological and pathological roles. Recent advances in the molecular mechanisms regulating lymphangiogenesis have opened a new area of research on reparative lymphangiogenesis for the treatment of various pathological disorders comprising neurological disorders, cardiac repair, autoimmune disease, obesity, atherosclerosis, etc. Reactive oxygen species (ROS) produced by the various cell types serve as signaling molecules in several cellular mechanisms and regulate various aspects of growth-factor-mediated responses, including lymphangiogenesis. The ROS, including superoxide anion, hydrogen peroxide, and nitric oxide, play both beneficial and detrimental roles depending upon their levels and cellular microenvironment. Low ROS levels are essential for lymphangiogenesis. On the contrary, oxidative stress due to enhanced ROS generation and/or reduced levels of antioxidants suppresses lymphangiogenesis via promoting lymphatic endothelial cell apoptosis and death. In this review article, we provide an overview of types and sources of ROS, discuss the role of ROS in governing lymphangiogenesis and lymphatic function, and summarize the role of lymphatics in various diseases.
1. Introduction
The lymphatic system, constituted by a network of lymphatic vessels (LVs), lymph nodes (LNs), and lymphoid organs, runs parallel to the blood vascular system. Both lymphatic and blood circulatory systems work in synchrony to maintain tissue homeostasis. The blood vascular system supplies nutrients, oxygen, and hormones to various body organs. In contrast, the lymphatic system plays an important role in transporting extravasated interstitial fluid, immune cells, inflammatory cytokines, antigens, and lipoproteins from the peripheral tissue to the draining LNs and back to the systemic venous circulation. Thus, the lymphatic system is pivotal for the maintenance of interstitial fluid homeostasis, host defense, adaptive immunity, and regulation of inflammatory responses [1,2,3]. The LVs are present throughout the human body with the exception of bone marrow and tissues, such as the epidermis, where blood vessels are also absent. These vessels are categorized in a hierarchical network of vessels, including capillaries, pre-collecting, and collecting lymphatics, based on their specific functions and morphological features [4,5]. Lymph flows through the lymphatic network in a unidirectional manner and at low pressure. The lymph flow rate is influenced by various extrinsic and intrinsic forces. Dysfunctional lymphatic vessels are responsible for several pathological conditions, including inherited and acquired lymphedema, malabsorption syndromes, autoimmune disease, atherosclerosis, neurological disorders, and immune deficiency [6,7,8].
The LVs are lined by a single layer of lymphatic endothelial cells (LECs), which possess specific markers and regulatory molecules, including Prospero homeobox 1 (Prox1), podoplanin, vascular endothelial growth factor receptor 3 (VEGFR3), and lymphatic vessel endothelial hyaluronan receptor 1 (LYVE-1) [9,10,11,12,13]. Like angiogenesis, lymphangiogenesis is the formation of new lymphatic vessels from pre-existing lymphatics. It results from a complex series of cellular events, including proliferation, sprouting, migration, and formation of vessel-like structures by LEC. Unlike developmental lymphangiogenesis, lymphangiogenesis in adults is dysregulated and less coordinated and occurs in pathological conditions such as inflammation, wound healing, and tumor growth [14,15,16]. These pathologies are often characterized by an accumulation of inflammatory cells and tissue edema, which necessitate lymphangiogenesis and LV remodeling for the removal of immune cells, cytokines, and tissue fluid [15,16]. Due to the various functions of LVs, lymphangiogenesis is regulated by multiple signaling pathways, as reviewed earlier [17,18].
Reactive oxygen species (ROS) such as superoxide anion, hydrogen peroxide, and nitric oxide are known to play both physiological and pathophysiological roles. Low levels of ROS are required to mediate LEC proliferation and migration, contributing to lymphangiogenesis. Increased generation of ROS by LEC leads to oxidative stress and inhibits LEC proliferation, migration, and tube formation via inducing apoptosis and cell death [19,20,21]. Based on these and the literature review, in this review article, we provide an overview of types and sources of ROS, discuss the role of ROS in governing lymphangiogenesis and lymphatic function in various pathological conditions, and summarize the role of LVs in various diseases.
2. Reactive Oxygen Species and Free Radicals
Reactive molecules and free radicals derived from molecular oxygen are called ROS. Free radicals are molecular species capable of independent existence that contain one or more unpaired electrons and include nitric oxide (NO•), superoxide anion (O2•−), hydroxyl radical (•OH), and lipid peroxyl radical (LOO•). Examples of non-radical derivatives of molecular oxygen are peroxynitrite (ONOO−), hydrogen peroxide (H2O2), hypochlorous acid (HOCl), and ozone (O3). ROS are synthesized as necessary intermediates in a broad range of biochemical processes and function as second messengers in physiological signaling mechanisms, contributing to the maintenance of tissue homeostasis [22]. In contrast, under pathophysiological conditions, overproduction of ROS and/or diminished antioxidant systems (also known as oxidative stress) may induce oxidative damage to DNA, protein, and lipid molecules. Oxidative-stress-induced alterations from physiological to pathophysiological signal transduction pathways and subsequent cellular damage play a critical role in the initiation, development, and progression of several diseases, including cardiovascular, inflammatory, neurologic, cancer, diabetes, and aging [19,23,24,25]. In this section, we briefly describe the chemistry and biochemistry of ROS and free radicals.
2.1. Nitric Oxide (NO•)
NO• plays a role in several biological processes and has vasodilatory, anti-inflammatory, and anti-thrombotic activities. It is produced by endothelial nitric oxide synthase (eNOS), neuronal NOS (nNOS), and inducible NOS (iNOS) [22]. These isoforms catalyze the formation of L-citrulline from L-arginine, and NO• is produced as a byproduct of the reaction. NOS-mediated NO• production is dependent on the availability of oxygen, so in case of limited oxygen supply, the nitrate–nitrite–NO pathway acts as a backup system to maintain sufficient NO• production [26]. Nitrate is first reduced by gastrointestinal and oral commensal bacteria to nitrite, which is further reduced to NO• by various pathways, as mentioned in Table 1. The formation of the iron–nitrosyl complex by the interaction of NO• to Fe2+ heme protein activates soluble guanylyl cyclase (sGC), which synthesizes second messenger cyclic guanosine monophosphate (cGMP). This sGC-cGMP signaling plays an important role in vasodilation, nerve signaling, mitochondrial biogenesis, angiogenesis, and lymphangiogenesis [27,28]. NO• is also responsible for decreased production of O2•− from complex I and III of the electron transport chain, mitochondrial cytochrome c release, and apoptosis [29,30]. Furthermore, NO• is responsible for limiting calcium availability in vascular smooth muscle cells due to ATP-dependent potassium channel opening, which prevents myosin light chain 2 (MLC2) phosphorylation and inhibits vasoconstriction [31]. The equilibrium between NO• and O2•− production is regulated by the availability of tetrahydrobiopterin (BH4, a cofactor required for NOS activity). The outcomes of reduced availability of BH4 relative to its oxidized form, dihydrobiopterin (BH2), involve increased O2•− release and decreased NO• production [32]. This situation is called eNOS uncoupling, which can be culpable for more superoxide production by the activation of oxidase enzymes, particularly NADPH oxidases (NOXs) and xanthine oxidase (XO); and progressive reduction in NO• bioavailability [33]. Moreover, the interaction between NO• and O2•− forms ONOO−, a strong oxidant that can oxidize BH4 and leads to enhanced eNOS uncoupling.
2.2. Superoxide Anion (O2•−)
O2•− is the precursor of most ROS and is rapidly dismuted to H2O2 spontaneously or via the reaction catalyzed by superoxide dismutase (SOD). The rate of spontaneous dismutation of O2•− is very low compared to enzymatic dismutation (8 × 104 M−1 s−1 versus 2 × 109 M−1 s−1) [34]. Enzymes involved in the biological production of O2•− include NOX isoforms, XO, uncoupled eNOS, and lipoxygenase. Phagocytes involving myeloid cells-neutrophils, macrophages, and monocytes, kill invading pathogens via producing a large amount of O2•−, also called a respiratory burst. It is also formed due to the leakage of one electron from the mitochondrial electron transport chain to molecular oxygen [35]. It has been reported that approximately 1–3% of electrons leak to produce superoxide anions during the mitochondrial transport chain [36].
2.3. Hydrogen Peroxide (H2O2)
H2O2, a highly stable and cell-permeable molecule, is generated because of O2•− dismution catalyzed by SOD. It acts as a signaling molecule in cellular signal transduction pathways [37]. In addition, XO and Nox4 are direct sources of H2O2 [37,38]. It plays a role in various physiological activities involving cell differentiation, proliferation, and migration [22,39]. Due to its cell-permeable nature, it can act in a paracrine manner between various cell types to regulate cellular signaling.
2.4. Peroxynitrite (ONOO−)
It is formed because of the interaction of NO• with O2•− and can lead to increased NOS uncoupling in the endothelium. It is highly toxic and acts as a substrate for the formation of super active nitroso peroxo carboxylate (ONOOCO2−) or peroxynitrous acid (ONOOH) [40]. It is capable of altering the oxidative state of lipids, DNA, and tyrosine and methionine residues in proteins [41]. Peroxynitrite stimulates the nitration of tyrosine residues present in proteins, for instance, SOD, by reaction mediated by transition metals. [42]. Further, peroxynitrite in phagocytic cells acts as a cytotoxic effector molecule against invading pathogens, including bacteria and parasites [43,44,45].
2.5. Hydroxyl Radical (•OH)
The dismution of H2O2 (Fenton reaction) and peroxynitrous acid (ONOOH), which is formed from the oxidation of ONOO−, results in the generation of •OH. Haber–Weiss reaction comprising spontaneous interaction of O2•− and H2O2 is an alternate source of hydroxyl radical production [46]. Generally, •OH does not play any role in cell signaling, but it is an important contributor to oxidative stress [47]. Its cellular levels can be altered by antioxidant enzymes and iron ligands.
2.6. Lipid Peroxyl Radical (LOO•)
Oxidation of unsaturated fatty acids present in cell membranes and lipoproteins leads to the formation of a lipid alkyl radical (L•), which rapidly reacts with molecular oxygen to form lipid peroxyl radical (LOO•). LOO• radical reacts with various proteins and carbohydrates by lowering the activation energy even much more than enzyme-catalyzed reactions, leading to the production of corresponding carbonyl compounds [48]. These radicals are highly destructive to cells, and once produced, they may lead to unrelenting lipid breakdown [49].
2.7. Hypochlorous Acid (HOCl)
It is generated in inflammatory cells such as activated neutrophils, monocytes, and macrophages, which have myeloperoxidase (MPO) enzyme required to produce HOCl. It is synthesized by the interaction of H2O2 with chloride ions. This HOCl has a strong antimicrobial activity and can oxidize lipoproteins, lipids, and proteins [50,51]. HOCl can also be responsible for the formation of monochloramines (NH2Cl), ONOO−, •OH, singlet oxygen (O2), and O3 via interacting with NH3 and other ROS [50].
2.8. Ozone (O3)
Ozone has a powerful oxidizing property and can increase the process of leukocytosis and phagocytosis. It is produced by electric discharge and irradiation of oxygen (air) with short-wavelength ultraviolet radiations. It may be synthesized in vivo by an antibody-mediated H2O oxidation pathway [52]. It can oxidize various biomolecules including lipids, proteins, and nucleic acids [53,54].
3. Role of ROS in Regulating Lymphangiogenesis and Lymphatic Function
Excessive production of ROS, for instance, H2O2, contributes to cell death. ROS-stimulated cell death occurs due to oxidative damage to cellular macromolecules involving proteins, lipids, and nucleic acids, and/or induction of cell-death-signaling pathways [55,56]. Alternatively, high ROS levels activate cell survival molecular-signaling cascades such as mitogen-activated protein kinase and PI3K/Akt [57,58]. The PI3K/Akt signaling prevents oxidative-stress-induced cell death and promotes cell survival. Furthermore, ROS have been shown to trigger cell surface growth factor receptor-mediated cell survival signaling in various cell types [59,60,61,62]. Thus, induction of growth factor receptor-mediated signaling in response to ROS protects against oxidative-stress-induced cell damage. Nitric oxide derived from eNOS in vascular endothelial cells plays an important role in stimulating angiogenesis and maintaining vascular contractility [63,64]. Like vascular endothelial cells, LECs have eNOS, and NO-derived from eNOS has been observed to be essential for lymphangiogenesis [65]. Despite numerous indications of the regulation of blood vessel formation (angiogenesis) by oxidative stress, the effects of ROS in modulating lymphangiogenesis are understudied to date.
3.1. Nitric Oxide
Stimulation of LECs with VEGF-C, a lymphangiogenic factor, has been shown to activate eNOS, which leads to the generation of NO and a subsequent increase in LEC proliferation and lymphangiogenesis [66] (Figure 1). Under physiological conditions, various immune cells, interstitial fluid, cytokines, and antigens present in initial lymphatics are carried by collecting LVs to draining LNs. The lymph flow in collecting LVs depends on the contractility of these collecting LVs. NO-derived from LEC eNOS is important for the maintenance of lymphatic contractility [67]. In addition, NO released by LECs regulates lymphatic permeability [68,69]. Additionally, inhibition of NO production with a NOS inhibitor, L-NMMA, blocks LV regeneration. Expression/activity of NOS in tumor tissues positively correlates with lymphatic metastasis in various types of tumors [70]. Genetic deletion of eNOS and pharmacological inhibition of its activity reduces peritumoral lymphatic hyperplasia in VEGF-C-overexpressing fibrosarcoma and attenuates trafficking of tumor cells to draining LNs, suggesting the role of NO in regulating lymphatic drainage [66]. Inhibition of NO-mediated signaling using an sGC inhibitor abrogates ultraviolet B-irradiation-induced LV enlargement, edema formation, and skin inflammation in mouse models [28]. A recent study by Singla et al. reported matricellular protein R-spondin 2 (RSPO2) as an anti-lymphangiogenic protein [21]. The authors observed that RSPO2 suppresses VEGF-C-stimulated Akt and eNOS phosphorylation, leading to attenuation of NO production by LEC and subsequent impairment in lymphangiogenesis. Moreover, pharmacological NO supplementation using an NO donor prevented the inhibitory effects of RSPO2 on lymphangiogenesis [21]. All this information suggests that NO-sGC signaling is pivotal in regulating lymphangiogenesis and maintenance of LVs. Increased iNOS levels disrupt endogenous NO gradients normally regulated by eNOS and lead to supra-physiological levels of NO, which in turn results in the induction of nitrosative stress [71]. Liao et al. demonstrated that under inflammatory conditions, NO derived from iNOS-overexpressing CD11b+ Gr-1+ myeloid-derived suppresser cells present around the subcutaneous LVs suppresses lymphatic contractions [72] (Figure 1). A study by Rehal et al. reported leaky and dilated LVs surrounded by iNOS+ CD11b+ inflammatory cells in obese mice [73]. These inflammatory cells caused an increased generation of peroxynitrite in obese mice. Additionally, LECs isolated from obese mice exhibited reduced VEGFR3 and podoplanin expression. These findings suggest that enhanced iNOS-derived NO generation in obesity contributes to lymphatic injury and impaired lymphatic pumping via nitrosative stress. Other reports revealed inhibition of lymphatic contractile function with high NO levels and impaired NO signaling in diabetic condition [74,75,76]. Additionally, reduced NO production due to decreased eNOS levels in the lymphatic thoracic duct isolated from rats with metabolic syndrome has been shown to be responsible for reduced vessel contractility [77]. Morris et al. showed KLF2-PPAR-γ-signaling-dependent elevation of NOX-derived ROS production and reduction of bioavailable NO in response to shear stress as observed with chronically increased lymph flow [78]. Further, it has been shown that shear stress reduces eNOS expression and activity in LECs [79].
3.2. Superoxide Anion and H2O2
Increased generation of O2•− scavenges NO and reduces NO bioavailability. Interaction of NO with O2•− forms peroxynitrite (ONOO−), which can further enhance protein modification and DNA damage. Singla et al. observed Nox4 as the major NOX isoform in LECs, which mainly generates H2O2 [21]. Exposure of LECs to H2O2 induces VEGFR3 activation and downstream Akt phosphorylation [80]. VEGFR3 activation following H2O2 treatment occurs as a compensatory mechanism to promote the survival of LECs, and treatments with antioxidants including N-acetylcysteine and catalase prevent H2O2-stimulated VEGFR3 activation [80]. Considering the importance of VEGFR3 activation in the development and maintenance of the lymphatic system, these findings may be the key to understanding the pathogenesis of lymphatic-related diseases such as lymphedema. Hereditary lymphedema patients are susceptible to ROS-induced cell damage. Elevated ROS generation and augmented lipid peroxidation occurring in lymphoedematous tissue are considered to induce lymphatic endothelium damage [81,82]. Wu et al. reported that LECs isolated from diabetic mice have high oxidative stress compared with cells isolated from control mice, and LEC ROS levels inversely correlate with both in vitro and in vivo lymphangiogenesis [19]. Mechanistically, the authors found that hyperglycemia-stimulated ROS generation induces c-Src-dependent upregulation of epsins, which are responsible for the degradation of VEGFR3 in LECs. Further, treatment of LECs with a high concentration of H2O2, as observed in diabetes, causes VEGFR3 phosphorylation and degradation in a ligand-(VEGF-C) independent manner [19]. These studies demonstrate that cell surface VEGFR3 expression in LECs plays a beneficial role in protecting against oxidative-stress-induced cell damage, and loss of VEGFR3 promotes pathological conditions [19,80].
Zawieja et al. reported attenuated lymphatic pumping after exposure to hypoxanthine and xanthine oxidase, which stimulate the generation of O2•− and H2O2. Moreover, treatment with SOD partially prevented the effects of hypoxanthine/xanthine oxidase on lymphatic contractions [83]. In another study, Zawieja and Davis investigated the effects of H2O2 challenge on the active pumping activity of mesenteric collecting lymphatics and observed significantly declined contraction frequency and lymph flow in H2O2-exposed lymphatics [84]. Further, a previous study has shown increased permeability of microvasculature in response to xanthine treatment, and treatment with O2•− and •OH scavengers reduced microvascular permeability [85]. In aging, there is a progressive decrease in NO levels and a concurrent increase in the production of free radicals. Aging-associated oxidative stress may be related to reduced levels of antioxidant enzymes such as SOD and/or increased ROS production [86,87]. A previous report suggested that aging remarkably reduces mesenteric LV contractility, which may limit the ability of these vessels to clear interstitial fluids and inflammatory cells from the site of inflammation in elderly individuals [88]. A subsequent study revealed elevated ROS production and reduced antioxidant enzyme levels in LECs isolated from aged rats, which suggests that aging-associated oxidative stress may contribute to lymphatic pump dysfunction in the elderly [89]. Thus, both O2•− and H2O2 play inhibitory roles in governing lymphatic pumping. A recently published study showed that oxidized low-density lipoprotein (oxLDL) inhibits lymphangiogenesis, blocks cell cycle progression, reduces expression of Akt and eNOS, increases p27 (an inhibitor of the cell cycle) expression, and induces intracellular ROS generation in LECs [20]. Additionally, CD36 knockdown in LECs prevents oxLDL-induced suppression of lymphangiogenesis. However, it is not known whether scavenging ROS can prevent oxLDL′s effects on lymphangiogenesis.
ROS production is high in the colon of IBD patients, and its levels correlate with the incidence of colitis [90,91]. However, in the case of IBD, elevated inflammation-induced LV density and defective lymphatic drainage have been observed compared with controls [92,93]. Further, Nox1 is highly expressed in colon cancer and supports tumor growth. Stalin et al. have recently demonstrated that inhibition of Nox1-mediated signaling via genetic deletion and pharmacological approach (GKT771) reduces lymphangiogenesis, suppresses recruitment of proinflammatory macrophages, and retards tumor growth [94], suggesting that ROS contribute to colon lymphangiogenesis in the cancer setting. Considering the findings of Wang et al., it is possible that ROS contribute to lymphangiogenesis via VEGFR3 activation in tumor tissue [80]. Recently, it has been demonstrated that Nox4 promotes both VEGF-C-dependent and -independent lymphangiogenesis [95]. Additionally, there is evidence of engagement of Nox2-derived ROS in regulating lymphangiogenesis [96]. LPA-stimulated ROS production, which is mediated by phospholipase C and protein kinase C, regulates LPA 1/3-dependent VEGF-C expression in prostate cancer cells [97]. LPA stimulates lymphangiogenesis in the tumor-xenograft mouse models via controlling calreticulin expression. Boehme et al. have recently demonstrated that LECs exposed to prolonged pathologically elevated lymph flow exhibit hyperproliferative growth, which is mediated by elevated hypoxia-inducible factor (HIF)-1α expression and increased mitochondrial NOX-derived ROS production [98]. In addition, the authors report attenuated proliferation of LECs exposed to mitochondrial antioxidants, indicating the importance of mitochondrial ROS in driving LEC proliferation under mechanical stress. Under hypoxic conditions, tumor cells upregulate HIF expression and activity, which induce the transcription of various growth factors′ genes responsible for increasing angiogenesis and lymphangiogenesis [99]. All the above findings suggest that ROS regulates lymphatic permeability, contractility and drainage, and lymphangiogenesis. Further studies are required to investigate the role of ROS in specific diseases and cell types.
4. Role of Lymphatic Vessels in Various Pathologies
4.1. Tumor Metastasis
Lymphatics serve as a route for cancer progression and metastasis. Secretion of well-known lymphangiogenic factor vascular endothelial growth factor (VEGF)-C and other growth factors by tumor cells lead to increased LV density in tumor tissues [100]. Tumor-associated lymphatics aid in the drainage of interstitial fluid containing various cell types and tumor-cell-originated macromolecules to sentinel LNs. Metastatic cells that detach from the primary tumor readily enter into highly permeable surrounding LVs and reach distant organs (Figure 2). Previous studies have indicated positive correlations between the levels of VEGF-C, VEGF-D, and VEGFR3 with an increased incidence of LN and distant organ metastasis [101,102,103]. In addition to metastasis, angiogenesis and lymphangiogenesis support tumor growth by other mechanisms. For example, the blood vascular system supplies nutrients and oxygen to the tumor hypoxic environment, while inflamed lymphatics induce an immunosuppressive microenvironment via reducing dendritic cell (DC)-mediated cytotoxic lymphocyte function by producing T cell inhibitory programmed-death ligand 1 (PD-L1), transforming growth factor-β (TGF-β), iNOS), and indoleamine 2,3-dioxygenase (IDO), thereby stimulating tumor progression [104]. Moreover, tumor tissue LV density has been shown to correlate with poor prognosis in cancer patients [105,106,107]. A previous study employing photodynamic therapy with benzoporphyrin-derivative verteporfin to destroy tumor-associated LVs showed prevention of mouse melanoma cell metastasis and subsequent tumor relapse. In addition, the combination of photodynamic and anti-lymphangiogenic therapies further reduced the invasion of tumor cells into LVs [108]. Conversely, drainage of tumor antigen-carrying migratory DCs to draining LNs is required for an antitumor immune response [109,110]. In addition, Fankhauser et al., in their study using the mouse melanoma model, reported enhanced T cell infiltration and potentiation of immunotherapy with experimental induction of tumor lymphangiogenesis [111]. Similar beneficial effects of augmented intra-tumoral LV density have been shown in colorectal cancer [112]. Therefore, despite the role of LVs in promoting tumor metastasis, LVs are required for the generation of optimum antitumor immune response to potentiate immune therapies. Future studies are required to further explore the role of LVs in specific cancer types.
4.2. Inflammation
Peripheral LVs serve as the principal route for the transportation of soluble antigens, cytokines, and immune cells from peripheral tissue to draining LNs and aid in the resolution of inflammation. LNs possess discrete compartments for T- and B lymphocytes. LECs present in afferent LVs via the expression of various adhesion molecules and chemokines control the entry and transport of immune cells through LVs and their positioning within LNs. LECs secrete chemokines CCL21 and CCL19, which guide and help in homing of CCR7+ activated DCs, macrophages, T- and B lymphocytes to lymphatics and LNs [113,114,115], while LECs with low expression of podoplanin and CCL21 but high CCR10 ligand CCL27 and Duffy antigen receptor for chemokines (DARC) levels direct the movement of CCR10+ T-lymphocytes into LVs [116]. Medullary LECs present in LNs expressing self-antigens and PD-L1 are involved in the depletion of alloreactive CD8+ T cells [117]. Cell adhesion molecules-common lymphatic endothelial and vascular receptor-1 and mannose receptor 1, have been shown to regulate leukocyte trafficking via LVs [118]. Interestingly, a study suggested reduced maturation of DCs via interaction of DC CD11b with intercellular adhesion molecule 1 receptor present in inflamed LECs [119].
Sphingosine 1-phosphate (S1P), synthesized intracellularly by phosphorylation of sphingosine, is a metabolic intermediate linking sphingolipids to glycerophospholipids [120]. The levels of S1P vary between blood (∼1 μM) and lymph (∼0.1 μM). LEC-dependent S1P gradient within LNs is required for egress of T- and B-lymphocytes from LNs into efferent lymph and thus is important for lymphocyte recirculation [121]. Further, S1P enhances the survival of naïve T cells and regulates the organization of LV junctions [122,123]. Lysophosphatidic acid (LPA), a bioactive lipid, has also been reported to support lymphocyte trafficking [124]. Altogether, these discoveries support the pivotal role of lymphatics in controlling immune response and inflammation.
4.3. Gut Homeostasis and Inflammatory Bowel Disease
Intestinal LVs include lacteals present in intestinal villi and mesenteric collecting LVs and constitute the largest lymphatic bed of the human body. These vessels aid in the drainage of lymph from gastrointestinal and lumbar regions into cisterna chyli and promote intestinal homeostasis [125]. Intestinal LVs also play crucial roles in the absorption of dietary lipids and the maintenance of gut immunity [126]. In adults, most of the lymphatics are quiescent under physiological conditions; however, lacteal LECs proliferate slowly and continuously, suggesting an ongoing lymphangiogenesis in adult lacteals [127]. A continuous lacteal regeneration may be a compensatory mechanism for consistent exposure to high lipoprotein levels, other biologically active dietary and microbial products, osmolarity gradients, and high mechanical strain originating from gut peristalsis and villus contractions [128,129,130]. Despite knowing the important role of intestinal lymphatics, the molecular factors regulating maintenance, renewal, and above functions of intestinal lymphatics are not fully understood.
Intestinal LVs are pivotal for gut immunosurveillance involved in inducing mucosal immunity and tolerance. These vessels regulate homing of CCR7+ DCs to mesenteric LNs in response to LEC-secreted CCL21, thereby establishing oral tolerance [131]. The intestinal epithelium has a high renewal rate and renews every 4–5 days in rodents and 5–7 days in humans. Intestinal LVs help in the transportation of apoptotic intestinal epithelial cells to mesenteric LNs and induce regulatory T (Treg) cells [132]. There are distinct mesenteric LNs to which different regions of the intestine drain and orchestrate adaptive immune responses specific to different intestinal segments [133].
Inflammatory bowel disease (IBD), including Crohn′s disease (CD) and ulcerative colitis (UC), is characterized by chronic intestinal inflammation. Its primary causes include genetic factors, dysregulation in immune response, diet, and other environmental cues [134]. Increased permeability of blood vessels in CD leads to accumulation of interstitial fluid that results in enhanced lymph flow to prevent edema. In human IBD patients, studies have reported augmented lymphangiogenesis, LV obstruction, dilation, and submucosal edema [135,136,137]. The presence of obstructive lymphoid aggregates consisting of T cells, B cells, and macrophages formed within lymphatic vasculature and expansion of mesenteric fat on the intestinal wall (also called creeping fat) are the major characteristics observed in patients with CD and result from impaired lymphatic drainage [138,139,140].
4.4. Lymphatics in Neurological Disorders
Under normal physiological conditions, the brain parenchyma is devoid of LVs, and clearance of cellular debris and waste products of the brain are mediated by the glymphatic system [141]. This system was proposed to aid in the exchange of small molecules between cerebrospinal fluid (CSF) and interstitial fluid [142,143]. Recent studies have characterized the architecture and importance of LVs present in the brain and spinal cord meninges [144,145,146,147]. In contrast to most peripheral lymphatic vasculature, which develops during mouse embryonic stage, meningeal LVs mature postnatally [148]. In adults, the development and maintenance of meningeal LVs depend on VEGF-C/VEGFR3 signaling [148]. The development, cytoarchitecture and morphology of meningeal LVs have been reviewed recently [149]. Meningeal LVs are identified as the major route of CSF outflow into deep and superficial cervical LNs in both humans and rodents [145,150,151,152,153]. These lymphatics are also required for the drainage of brain-associated interstitial fluid, solutes, and various immune cells (lymphocytes and CD11c+ CCR7+ DC) egress from CSF to the peripheral lymphatic system [145,151,153,154]. Citing the role of meningeal LVs in the above-mentioned processes, these lymphatics are involved in the pathogenesis of various age-related neurodegenerative diseases, such as Alzheimer′s disease. Ablation of meningeal LVs in mouse models of Alzheimer′s disease has been shown to increase brain β-amyloid levels and promotes its deposition in the meninges, demonstrating the role of these vessels in the clearance of β-amyloids [145,155]. Further, meningeal LVs are essential for the removal of extracellular tau aggregates from the central nervous system (CNS), which is another neuropathological hallmark of Alzheimer′s disease [156]. In addition, impaired meningeal lymphatic drainage leads to the accumulation of brain α-synuclein aggregates in Parkinson′s disease models [157,158] and induces neuroinflammation [159,160].
Contrary to the beneficial roles of meningeal LVs in Alzheimer′s and Parkinson′s disease and attenuating neuroinflammation, these lymphatics contribute to the pathogenesis of autoimmune demyelinating disease of the CNS, multiple sclerosis. Further, by serving as a route for the drainage of neurological immune cells and antigens to cervical LNs, meningeal LVs may support CNS infection [153]. All these studies demonstrate the key role of lymphatics in neurological pathologies.
4.5. Cardiovascular Disease
Atherosclerosis is a vascular inflammatory disease characterized by the accumulation of lipids in large and medium-sized arteries. Hoggan et al. first reported the presence of LVs in the arterial wall >100 years ago [161]. Subsequent studies described the existence of lymphatics in the blood vessels of dogs, pigs, and humans [162,163,164,165,166]. Despite the presence of LVs in the arterial wall, the functional role of arterial lymphatics in atherosclerosis development has not been investigated until recently. Martel et al. demonstrated that these arterial LVs play a critical role in the removal of high-density lipoprotein cholesterol from atherosclerotic arteries and skin via macrophage reverse cholesterol transport (mRCT) [167]. Lymphatic insufficiency has been shown to elevate plasma cholesterol levels [168]. In addition, hypercholesterolemia downregulates the expression of lymphangiogenic factors, including VEGF-C, Ang2, and FoxC2 in peripheral tissues, and impairs lymphatic drainage [169]. ApoA-I and VEGF-C152S treatments have been shown to improve lymphatic transport and attenuate atherosclerotic lesion formation [170,171]. In addition, a recent study by Singla et al. demonstrated the inhibition of atherosclerosis and reduced accumulation of fluorescently-labeled cholesterol in the left carotid artery with improved arterial drainage to periarterial LNs [21]. The role of lymphatics in cholesterol transport has been reviewed in the past [172]. Dissections of the left carotid artery draining deep cervical LNs and surrounding LVs in ApoE-/- mice have been observed to augment atherosclerosis and promote the accumulation of CD3+ T cells in the intimal and adventitial arterial layers [173]. In addition, inhibition of lymphangiogenic VEGFR3-mediated signaling led to T cell enrichment in atherosclerotic lesions, suggesting the role of arterial lymphatics in the egression of arterial T cells. Taken together, these studies suggest that lymphatics play a beneficial role in arterial cholesterol efflux and inflammatory cell egress, which suppresses atherosclerosis development [174].
Blockade of coronary arteries due to atherosclerosis is responsible for the reduced supply of blood to the cardiac muscles, consequently leading to cardiac tissue damage, cell death, and heart failure. This process is accompanied by increased permeability of myocardial microvasculature and results in interstitial fluid accumulation (myocardial edema) and elevated lymph flow [175,176]. Previous studies have suggested that impairment of cardiac lymphatic drainage stimulates cardiac edema, and stimulated cardiac lymphangiogenesis associates with improved cardiac function via supporting the resolution of inflammation and reducing edema [177,178,179,180]. This information advocates that stimulated cardiac lymphangiogenesis may be a novel and potential therapeutic approach for treating cardiac disease.
4.6. Lymphedema
Tissue edema caused by an accumulation of protein-rich fluid due to impaired lymphatic drainage is referred to as lymphedema. It mostly occurs in limbs but can also affect the chest wall, abdomen, neck, and genital regions. Primary lymphedema is a genetic disease, e.g., Nonne–Milroy disease and Meige disease [181]. Secondary lymphedema is usually caused by infection (Wuchereria bancrofti) and surgical and radiotherapy procedures [182,183,184]. These diseases result from insufficient lymph drainage.
5. Conclusions
LVs have significantly different structures and functions from blood vessels due to their roles in the maintenance of tissue fluid homeostasis, uptake of dietary lipids and macromolecules, and immunosurveillance. Significant advances have been made in the last decade regarding the molecular mechanisms regulating LV formation. Low levels of ROS are required for lymphangiogenesis, while excess generation of ROS contributes to inhibition of lymphangiogenesis and impaired lymphatic drainage. However, the effects of ROS derived from LECs and other neighboring cell types (s) in regulating lymphangiogenesis are understudied. Existing data support a pivotal role of NOX-derived ROS in regulating LV formation, and future mechanistic studies are necessary to identify novel molecular stimulators of NOXs, investigate the mechanisms of NOX activation and downstream targets of ROS, and examine the effects of antioxidants in modulating lymphangiogenesis. Understanding these mechanisms may provide new potential therapeutic targets for the treatment of lymphatic-related pathological disorders.
Author Contributions
B.S. wrote, edited, and proofread the manuscript. R.V.A. assisted in the manuscript writing. S.K. (Shweta Kapil) and S.K. (Sonia Kiran). helped with the illustrations. Figures were created using BioRender. S.K. (Shweta Kapil), S.K. (Santosh Kumar), and U.P.S. reviewed and edited the article. All authors have read and agreed to submit the manuscript to the journal Cells for publication. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Institutes of Health grants (K99HL146954 and R00HL146954) awarded to B.S.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Girard, J.P.; Moussion, C.; Forster, R. HEVs, lymphatics and homeostatic immune cell trafficking in lymph nodes. Nat. Rev. Immunol. 2012, 12, 762–773. [Google Scholar] [CrossRef] [PubMed]
- Miller, N.E.; Michel, C.C.; Nanjee, M.N.; Olszewski, W.L.; Miller, I.P.; Hazell, M.; Olivecrona, G.; Sutton, P.; Humphreys, S.M.; Frayn, K.N. Secretion of adipokines by human adipose tissue in vivo: Partitioning between capillary and lymphatic transport. Am. J. Physiol. Endocrinol. Metab. 2011, 301, E659–E667. [Google Scholar] [CrossRef] [Green Version]
- Randolph, G.J.; Miller, N.E. Lymphatic transport of high-density lipoproteins and chylomicrons. J. Clin. Investig. 2014, 124, 929–935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrova, T.V.; Koh, G.Y. Biological functions of lymphatic vessels. Science 2020, 369, eaax4063. [Google Scholar] [CrossRef]
- Ulvmar, M.H.; Makinen, T. Heterogeneity in the lymphatic vascular system and its origin. Cardiovasc. Res. 2016, 111, 310–321. [Google Scholar] [CrossRef] [Green Version]
- Aspelund, A.; Robciuc, M.R.; Karaman, S.; Makinen, T.; Alitalo, K. Lymphatic System in Cardiovascular Medicine. Circ. Res. 2016, 118, 515–530. [Google Scholar] [CrossRef] [PubMed]
- Tammela, T.; Alitalo, K. Lymphangiogenesis: Molecular mechanisms and future promise. Cell 2010, 140, 460–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alitalo, K.; Tammela, T.; Petrova, T.V. Lymphangiogenesis in development and human disease. Nature 2005, 438, 946–953. [Google Scholar] [CrossRef]
- Banerji, S.; Ni, J.; Wang, S.X.; Clasper, S.; Su, J.; Tammi, R.; Jones, M.; Jackson, D.G. LYVE-1, a new homologue of the CD44 glycoprotein, is a lymph-specific receptor for hyaluronan. J. Cell Biol. 1999, 144, 789–801. [Google Scholar] [CrossRef]
- Breiteneder-Geleff, S.; Soleiman, A.; Kowalski, H.; Horvat, R.; Amann, G.; Kriehuber, E.; Diem, K.; Weninger, W.; Tschachler, E.; Alitalo, K.; et al. Angiosarcomas express mixed endothelial phenotypes of blood and lymphatic capillaries: Podoplanin as a specific marker for lymphatic endothelium. Am. J. Pathol. 1999, 154, 385–394. [Google Scholar] [CrossRef]
- Kriehuber, E.; Breiteneder-Geleff, S.; Groeger, M.; Soleiman, A.; Schoppmann, S.F.; Stingl, G.; Kerjaschki, D.; Maurer, D. Isolation and characterization of dermal lymphatic and blood endothelial cells reveal stable and functionally specialized cell lineages. J. Exp. Med. 2001, 194, 797–808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wigle, J.T.; Harvey, N.; Detmar, M.; Lagutina, I.; Grosveld, G.; Gunn, M.D.; Jackson, D.G.; Oliver, G. An essential role for Prox1 in the induction of the lymphatic endothelial cell phenotype. EMBO J. 2002, 21, 1505–1513. [Google Scholar] [CrossRef] [PubMed]
- Kaipainen, A.; Korhonen, J.; Mustonen, T.; van Hinsbergh, V.W.; Fang, G.H.; Dumont, D.; Breitman, M.; Alitalo, K. Expression of the fms-like tyrosine kinase 4 gene becomes restricted to lymphatic endothelium during development. Proc. Natl. Acad. Sci. USA 1995, 92, 3566–3570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, W.; Tammela, T.; Yamamoto, M.; Anisimov, A.; Holopainen, T.; Kaijalainen, S.; Karpanen, T.; Lehti, K.; Yla-Herttuala, S.; Alitalo, K. Notch restricts lymphatic vessel sprouting induced by vascular endothelial growth factor. Blood 2011, 118, 1154–1162. [Google Scholar] [CrossRef] [PubMed]
- Betterman, K.L.; Harvey, N.L. The lymphatic vasculature: Development and role in shaping immunity. Immunol. Rev. 2016, 271, 276–292. [Google Scholar] [CrossRef]
- Christiansen, A.; Detmar, M. Lymphangiogenesis and cancer. Genes Cancer 2011, 2, 1146–1158. [Google Scholar] [CrossRef] [Green Version]
- Bui, K.; Hong, Y.K. Ras Pathways on Prox1 and Lymphangiogenesis: Insights for Therapeutics. Front. Cardiovasc. Med. 2020, 7, 597374. [Google Scholar] [CrossRef]
- Csanyi, G.; Singla, B. Arterial Lymphatics in Atherosclerosis: Old Questions, New Insights, and Remaining Challenges. J. Clin. Med. 2019, 8, 495. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Rahman, H.N.A.; Dong, Y.; Liu, X.; Lee, Y.; Wen, A.; To, K.H.; Xiao, L.; Birsner, A.E.; Bazinet, L.; et al. Epsin deficiency promotes lymphangiogenesis through regulation of VEGFR3 degradation in diabetes. J. Clin. Investig. 2018, 128, 4025–4043. [Google Scholar] [CrossRef] [Green Version]
- Singla, B.; Lin, H.P.; Ahn, W.; White, J.; Csanyi, G. Oxidatively Modified LDL Suppresses Lymphangiogenesis via CD36 Signaling. Antioxidants 2021, 10, 331. [Google Scholar] [CrossRef]
- Singla, B.; Lin, H.P.; Chen, A.; Ahn, W.; Ghoshal, P.; Cherian-Shaw, M.; White, J.; Stansfield, B.K.; Csanyi, G. Role of R-spondin 2 in arterial lymphangiogenesis and atherosclerosis. Cardiovasc. Res. 2021, 117, 1489–1509. [Google Scholar] [CrossRef] [PubMed]
- Droge, W. Free radicals in the physiological control of cell function. Physiol. Rev. 2002, 82, 47–95. [Google Scholar] [CrossRef]
- Charles, R.L.; Eaton, P. Redox signalling in cardiovascular disease. Proteom. Clin. Appl. 2008, 2, 823–836. [Google Scholar] [CrossRef]
- Yang, S.; Lian, G. ROS and diseases: Role in metabolism and energy supply. Mol. Cell Biochem. 2020, 467, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Papaharalambus, C.A.; Griendling, K.K. Basic mechanisms of oxidative stress and reactive oxygen species in cardiovascular injury. Trends Cardiovasc. Med. 2007, 17, 48–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lundberg, J.O.; Weitzberg, E.; Gladwin, M.T. The nitrate-nitrite-nitric oxide pathway in physiology and therapeutics. Nat. Rev. Drug Discov. 2008, 7, 156–167. [Google Scholar] [CrossRef]
- Hobbs, A.J. Soluble guanylate cyclase: The forgotten sibling. Trends Pharmacol. Sci. 1997, 18, 484–491. [Google Scholar] [CrossRef]
- Kajiya, K.; Huggenberger, R.; Drinnenberg, I.; Ma, B.; Detmar, M. Nitric oxide mediates lymphatic vessel activation via soluble guanylate cyclase alpha1beta1-impact on inflammation. FASEB J. 2008, 22, 530–537. [Google Scholar] [CrossRef]
- Lundberg, J.O.; Gladwin, M.T.; Weitzberg, E. Strategies to increase nitric oxide signalling in cardiovascular disease. Nat. Rev. Drug Discov. 2015, 14, 623–641. [Google Scholar] [CrossRef]
- Chung, H.T.; Pae, H.O.; Choi, B.M.; Billiar, T.R.; Kim, Y.M. Nitric oxide as a bioregulator of apoptosis. Biochem. Biophys Res. Commun. 2001, 282, 1075–1079. [Google Scholar] [CrossRef]
- Rogers, N.M.; Sharifi-Sanjani, M.; Csanyi, G.; Pagano, P.J.; Isenberg, J.S. Thrombospondin-1 and CD47 regulation of cardiac, pulmonary and vascular responses in health and disease. Matrix Biol. 2014, 37, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Pacher, P.; Beckman, J.S.; Liaudet, L. Nitric oxide and peroxynitrite in health and disease. Physiol. Rev. 2007, 87, 315–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forstermann, U.; Munzel, T. Endothelial nitric oxide synthase in vascular disease: From marvel to menace. Circulation 2006, 113, 1708–1714. [Google Scholar] [CrossRef] [Green Version]
- Fridovich, I. Superoxide radical: An endogenous toxicant. Annu. Rev. Pharmacol. Toxicol. 1983, 23, 239–257. [Google Scholar] [CrossRef]
- Turrens, J.F. Mitochondrial formation of reactive oxygen species. J. Physiol. 2003, 552, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef] [PubMed]
- Meijles, D.N.; Pagano, P.J. Nox and Inflammation in the Vascular Adventitia. Hypertension 2016, 67, 14–19. [Google Scholar] [CrossRef] [Green Version]
- Kelley, E.E.; Khoo, N.K.; Hundley, N.J.; Malik, U.Z.; Freeman, B.A.; Tarpey, M.M. Hydrogen peroxide is the major oxidant product of xanthine oxidase. Free Radic. Biol. Med. 2010, 48, 493–498. [Google Scholar] [CrossRef] [Green Version]
- Ushio-Fukai, M.; Alexander, R.W. Reactive oxygen species as mediators of angiogenesis signaling: Role of NAD(P)H oxidase. Mol. Cell Biochem. 2004, 264, 85–97. [Google Scholar] [CrossRef]
- Beckman, J.S.; Koppenol, W.H. Nitric oxide, superoxide, and peroxynitrite: The good, the bad, and ugly. Am. J. Physiol. 1996, 271, C1424–C1437. [Google Scholar] [CrossRef] [Green Version]
- Douki, T.; Cadet, J. Peroxynitrite mediated oxidation of purine bases of nucleosides and isolated DNA. Free Radic. Res. 1996, 24, 369–380. [Google Scholar] [CrossRef] [PubMed]
- Ischiropoulos, H.; Zhu, L.; Chen, J.; Tsai, M.; Martin, J.C.; Smith, C.D.; Beckman, J.S. Peroxynitrite-mediated tyrosine nitration catalyzed by superoxide dismutase. Arch. Biochem. Biophys. 1992, 298, 431–437. [Google Scholar] [CrossRef]
- Zhu, L.; Gunn, C.; Beckman, J.S. Bactericidal activity of peroxynitrite. Arch. Biochem. Biophys. 1992, 298, 452–457. [Google Scholar] [CrossRef]
- Denicola, A.; Rubbo, H.; Rodriguez, D.; Radi, R. Peroxynitrite-mediated cytotoxicity to Trypanosoma cruzi. Arch. Biochem. Biophys. 1993, 304, 279–286. [Google Scholar] [CrossRef]
- Radi, R. Oxygen radicals, nitric oxide, and peroxynitrite: Redox pathways in molecular medicine. Proc. Natl. Acad. Sci. USA 2018, 115, 5839–5848. [Google Scholar] [CrossRef] [Green Version]
- Kehrer, J.P. The Haber-Weiss reaction and mechanisms of toxicity. Toxicology 2000, 149, 43–50. [Google Scholar] [CrossRef]
- Thomas, C.; Mackey, M.M.; Diaz, A.A.; Cox, D.P. Hydroxyl radical is produced via the Fenton reaction in submitochondrial particles under oxidative stress: Implications for diseases associated with iron accumulation. Redox. Rep. 2009, 14, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Spiteller, G.; Afzal, M. The action of peroxyl radicals, powerful deleterious reagents, explains why neither cholesterol nor saturated fatty acids cause atherogenesis and age-related diseases. Chemistry 2014, 20, 14928–14945. [Google Scholar] [CrossRef] [PubMed]
- Cheeseman, K.H. Tissue injury by free radicals. Toxicol. Ind. Health 1993, 9, 39–51. [Google Scholar] [CrossRef]
- Hampton, M.B.; Kettle, A.J.; Winterbourn, C.C. Inside the neutrophil phagosome: Oxidants, myeloperoxidase, and bacterial killing. Blood 1998, 92, 3007–3017. [Google Scholar] [CrossRef]
- Hawkins, C.L.; Davies, M.J. Role of myeloperoxidase and oxidant formation in the extracellular environment in inflammation-induced tissue damage. Free Radic. Biol. Med. 2021, 172, 633–651. [Google Scholar] [CrossRef] [PubMed]
- Lerner, R.A.; Eschenmoser, A. Ozone in biology. Proc. Natl. Acad. Sci. USA 2003, 100, 3013–3015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freeman, B.A.; Mudd, J.B. Reaction of ozone with sulfhydryls of human erythrocytes. Arch. Biochem. Biophys. 1981, 208, 212–220. [Google Scholar] [CrossRef]
- Mustafa, M.G. Biochemical basis of ozone toxicity. Free Radic. Biol. Med. 1990, 9, 245–265. [Google Scholar] [CrossRef]
- Martindale, J.L.; Holbrook, N.J. Cellular response to oxidative stress: Signaling for suicide and survival. J. Cell Physiol. 2002, 192, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Irani, K. Oxidant signaling in vascular cell growth, death, and survival: A review of the roles of reactive oxygen species in smooth muscle and endothelial cell mitogenic and apoptotic signaling. Circ. Res. 2000, 87, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Martindale, J.L.; Liu, Y.; Holbrook, N.J. The cellular response to oxidative stress: Influences of mitogen-activated protein kinase signalling pathways on cell survival. Biochem. J. 1998, 333 Pt 2, 291–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, S.; Chock, P.B. Implication of phosphatidylinositol 3-kinase membrane recruitment in hydrogen peroxide-induced activation of PI3K and Akt. Biochemistry 2003, 42, 2995–3003. [Google Scholar] [CrossRef]
- Huang, R.P.; Peng, A.; Golard, A.; Hossain, M.Z.; Huang, R.; Liu, Y.G.; Boynton, A.L. Hydrogen peroxide promotes transformation of rat liver non-neoplastic epithelial cells through activation of epidermal growth factor receptor. Mol. Carcinog. 2001, 30, 209–217. [Google Scholar] [CrossRef]
- Nair, V.D.; Olanow, C.W.; Sealfon, S.C. Activation of phosphoinositide 3-kinase by D2 receptor prevents apoptosis in dopaminergic cell lines. Biochem. J. 2003, 373, 25–32. [Google Scholar] [CrossRef]
- Saito, S.; Frank, G.D.; Mifune, M.; Ohba, M.; Utsunomiya, H.; Motley, E.D.; Inagami, T.; Eguchi, S. Ligand-independent trans-activation of the platelet-derived growth factor receptor by reactive oxygen species requires protein kinase C-delta and c-Src. J. Biol. Chem. 2002, 277, 44695–44700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koshio, O.; Akanuma, Y.; Kasuga, M. Hydrogen peroxide stimulates tyrosine phosphorylation of the insulin receptor and its tyrosine kinase activity in intact cells. Biochem. J. 1988, 250, 95–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Loutzenhiser, R.D.; Cupples, W.A. Frequency modulation of renal myogenic autoregulation by perfusion pressure. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 293, R1199–R1204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forstermann, U.; Sessa, W.C. Nitric oxide synthases: Regulation and function. Eur. Heart J. 2012, 33, 829–837. [Google Scholar] [CrossRef] [Green Version]
- Marchetti, C.; Casasco, A.; Di Nucci, A.; Reguzzoni, M.; Rosso, S.; Piovella, F.; Calligaro, A.; Polak, J.M. Endothelin and nitric oxide synthase in lymphatic endothelial cells: Immunolocalization in vivo and in vitro. Anat. Rec. 1997, 248, 490–497. [Google Scholar] [CrossRef]
- Lahdenranta, J.; Hagendoorn, J.; Padera, T.P.; Hoshida, T.; Nelson, G.; Kashiwagi, S.; Jain, R.K.; Fukumura, D. Endothelial nitric oxide synthase mediates lymphangiogenesis and lymphatic metastasis. Cancer Res. 2009, 69, 2801–2808. [Google Scholar] [CrossRef] [Green Version]
- Schmid-Schonbein, G.W. Nitric oxide (NO) side of lymphatic flow and immune surveillance. Proc. Natl. Acad. Sci. USA 2012, 109, 3–4. [Google Scholar] [CrossRef] [Green Version]
- Ohhashi, T.; Mizuno, R.; Ikomi, F.; Kawai, Y. Current topics of physiology and pharmacology in the lymphatic system. Pharmacol. Ther. 2005, 105, 165–188. [Google Scholar] [CrossRef] [Green Version]
- Hagendoorn, J.; Padera, T.P.; Kashiwagi, S.; Isaka, N.; Noda, F.; Lin, M.I.; Huang, P.L.; Sessa, W.C.; Fukumura, D.; Jain, R.K. Endothelial nitric oxide synthase regulates microlymphatic flow via collecting lymphatics. Circ. Res. 2004, 95, 204–209. [Google Scholar] [CrossRef]
- Fukumura, D.; Kashiwagi, S.; Jain, R.K. The role of nitric oxide in tumour progression. Nat. Rev. Cancer 2006, 6, 521–534. [Google Scholar] [CrossRef]
- Soskic, S.S.; Dobutovic, B.D.; Sudar, E.M.; Obradovic, M.M.; Nikolic, D.M.; Djordjevic, J.D.; Radak, D.J.; Mikhailidis, D.P.; Isenovic, E.R. Regulation of Inducible Nitric Oxide Synthase (iNOS) and its Potential Role in Insulin Resistance, Diabetes and Heart Failure. Open Cardiovasc. Med. J. 2011, 5, 153–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, S.; Cheng, G.; Conner, D.A.; Huang, Y.; Kucherlapati, R.S.; Munn, L.L.; Ruddle, N.H.; Jain, R.K.; Fukumura, D.; Padera, T.P. Impaired lymphatic contraction associated with immunosuppression. Proc. Natl. Acad. Sci. USA 2011, 108, 18784–18789. [Google Scholar] [CrossRef] [Green Version]
- Rehal, S.; Kataru, R.P.; Hespe, G.E.; Baik, J.E.; Park, H.J.; Ly, C.; Shin, J.; Mehrara, B.J. Regulation of lymphatic function and injury by nitrosative stress in obese mice. Mol. Metab. 2020, 42, 101081. [Google Scholar] [CrossRef] [PubMed]
- Gasheva, O.Y.; Zawieja, D.C.; Gashev, A.A. Contraction-initiated NO-dependent lymphatic relaxation: A self-regulatory mechanism in rat thoracic duct. J. Physiol. 2006, 575, 821–832. [Google Scholar] [CrossRef] [PubMed]
- Scallan, J.P.; Davis, M.J. Genetic removal of basal nitric oxide enhances contractile activity in isolated murine collecting lymphatic vessels. J. Physiol. 2013, 591, 2139–2156. [Google Scholar] [CrossRef]
- Scallan, J.P.; Hill, M.A.; Davis, M.J. Lymphatic vascular integrity is disrupted in type 2 diabetes due to impaired nitric oxide signalling. Cardiovasc. Res. 2015, 107, 89–97. [Google Scholar] [CrossRef] [Green Version]
- Zawieja, S.D.; Gasheva, O.; Zawieja, D.C.; Muthuchamy, M. Blunted flow-mediated responses and diminished nitric oxide synthase expression in lymphatic thoracic ducts of a rat model of metabolic syndrome. Am. J. Physiol. Heart. Circ. Physiol. 2016, 310, H385–H393. [Google Scholar] [CrossRef] [Green Version]
- Morris, C.J.; Kameny, R.J.; Boehme, J.; Gong, W.; He, Y.; Zhu, T.; Maltepe, E.; Raff, G.W.; Fineman, J.R.; Datar, S.A. KLF2-mediated disruption of PPAR-gamma signaling in lymphatic endothelial cells exposed to chronically increased pulmonary lymph flow. Am. J. Physiol. Heart Circ. Physiol. 2018, 315, H173–H181. [Google Scholar] [CrossRef]
- Datar, S.A.; Gong, W.; He, Y.; Johengen, M.; Kameny, R.J.; Raff, G.W.; Maltepe, E.; Oishi, P.E.; Fineman, J.R. Disrupted NOS signaling in lymphatic endothelial cells exposed to chronically increased pulmonary lymph flow. Am. J. Physiol. Heart Circ. Physiol. 2016, 311, H137–H145. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.F.; Zhang, X.; Groopman, J.E. Activation of vascular endothelial growth factor receptor-3 and its downstream signaling promote cell survival under oxidative stress. J. Biol. Chem. 2004, 279, 27088–27097. [Google Scholar] [CrossRef] [Green Version]
- Siems, W.G.; Brenke, R.; Beier, A.; Grune, T. Oxidative stress in chronic lymphoedema. QJM 2002, 95, 803–809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohkuma, M. Lipoperoxide in the dermis of patients with lymph stasis. Lymphology 1993, 26, 38–41. [Google Scholar] [PubMed]
- Zawieja, D.C.; Greiner, S.T.; Davis, K.L.; Hinds, W.M.; Granger, H.J. Reactive oxygen metabolites inhibit spontaneous lymphatic contractions. Am. J. Physiol. 1991, 260, H1935–H1943. [Google Scholar] [CrossRef] [PubMed]
- Zawieja, D.C.; Davis, K.L. Inhibition of the active lymph pump in rat mesenteric lymphatics by hydrogen peroxide. Lymphology 1993, 26, 135–142. [Google Scholar] [PubMed]
- Del Maestro, R.F.; Bjork, J.; Arfors, K.E. Increase in microvascular permeability induced by enzymatically generated free radicals. II. Role of superoxide anion radical, hydrogen peroxide, and hydroxyl radical. Microvasc. Res. 1981, 22, 255–270. [Google Scholar] [CrossRef]
- Brown, K.A.; Chu, Y.; Lund, D.D.; Heistad, D.D.; Faraci, F.M. Gene transfer of extracellular superoxide dismutase protects against vascular dysfunction with aging. Am. J. Physiol. Heart Circ. Physiol. 2006, 290, H2600–H2605. [Google Scholar] [CrossRef] [Green Version]
- Brown, K.A.; Didion, S.P.; Andresen, J.J.; Faraci, F.M. Effect of aging, MnSOD deficiency, and genetic background on endothelial function: Evidence for MnSOD haploinsufficiency. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 1941–1946. [Google Scholar] [CrossRef] [Green Version]
- Nagai, T.; Bridenbaugh, E.A.; Gashev, A.A. Aging-associated alterations in contractility of rat mesenteric lymphatic vessels. Microcirculation 2011, 18, 463–473. [Google Scholar] [CrossRef]
- Thangaswamy, S.; Bridenbaugh, E.A.; Gashev, A.A. Evidence of increased oxidative stress in aged mesenteric lymphatic vessels. Lymphat. Res. Biol. 2012, 10, 53–62. [Google Scholar] [CrossRef] [Green Version]
- Sedghi, S.; Keshavarzian, A.; Klamut, M.; Eiznhamer, D.; Zarling, E.J. Elevated breath ethane levels in active ulcerative colitis: Evidence for excessive lipid peroxidation. Am. J. Gastroenterol. 1994, 89, 2217–2221. [Google Scholar]
- Nishikawa, M.; Oshitani, N.; Matsumoto, T.; Nishigami, T.; Arakawa, T.; Inoue, M. Accumulation of mitochondrial DNA mutation with colorectal carcinogenesis in ulcerative colitis. Br. J. Cancer 2005, 93, 331–337. [Google Scholar] [CrossRef] [Green Version]
- Lee, A.S.; Hur, H.J.; Sung, M.J. The Effect of Artemisinin on Inflammation-Associated Lymphangiogenesis in Experimental Acute Colitis. Int. J. Mol. Sci. 2020, 21, 8068. [Google Scholar] [CrossRef] [PubMed]
- D’Alessio, S.; Tacconi, C.; Fiocchi, C.; Danese, S. Advances in therapeutic interventions targeting the vascular and lymphatic endothelium in inflammatory bowel disease. Curr. Opin. Gastroenterol. 2013, 29, 608–613. [Google Scholar] [CrossRef] [PubMed]
- Stalin, J.; Garrido-Urbani, S.; Heitz, F.; Szyndralewiez, C.; Jemelin, S.; Coquoz, O.; Ruegg, C.; Imhof, B.A. Inhibition of host NOX1 blocks tumor growth and enhances checkpoint inhibitor-based immunotherapy. Life Sci. Alliance 2019, 2, e201800265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Liu, Z.; Sun, J.; Song, X.; Bian, M.; Wang, F.; Yan, F.; Yu, Z. Inhibition of NADPH oxidase 4 attenuates lymphangiogenesis and tumor metastasis in breast cancer. FASEB J. 2021, 35, e21531. [Google Scholar] [CrossRef]
- Coso, S. Molecular mechanisms regulating lymphangiogenesis. Ph.D. Thesis, Monash University, Melbourne, Australia, 2017. [Google Scholar] [CrossRef]
- Lin, C.C.; Lin, C.E.; Lin, Y.C.; Ju, T.K.; Huang, Y.L.; Lee, M.S.; Chen, J.H.; Lee, H. Lysophosphatidic acid induces reactive oxygen species generation by activating protein kinase C in PC-3 human prostate cancer cells. Biochem. Biophys Res. Commun. 2013, 440, 564–569. [Google Scholar] [CrossRef] [PubMed]
- Boehme, J.T.; Morris, C.J.; Chiacchia, S.R.; Gong, W.; Wu, K.Y.; Kameny, R.J.; Raff, G.W.; Fineman, J.R.; Maltepe, E.; Datar, S.A. HIF-1alpha promotes cellular growth in lymphatic endothelial cells exposed to chronically elevated pulmonary lymph flow. Sci. Rep. 2021, 11, 1468. [Google Scholar] [CrossRef]
- Semenza, G.L. Cancer-stromal cell interactions mediated by hypoxia-inducible factors promote angiogenesis, lymphangiogenesis, and metastasis. Oncogene 2013, 32, 4057–4063. [Google Scholar] [CrossRef] [Green Version]
- Karaman, S.; Detmar, M. Mechanisms of lymphatic metastasis. J. Clin. Investig. 2014, 124, 922–928. [Google Scholar] [CrossRef] [Green Version]
- Stacker, S.A.; Caesar, C.; Baldwin, M.E.; Thornton, G.E.; Williams, R.A.; Prevo, R.; Jackson, D.G.; Nishikawa, S.; Kubo, H.; Achen, M.G. VEGF-D promotes the metastatic spread of tumor cells via the lymphatics. Nat. Med. 2001, 7, 186–191. [Google Scholar] [CrossRef]
- Hoshida, T.; Isaka, N.; Hagendoorn, J.; di Tomaso, E.; Chen, Y.L.; Pytowski, B.; Fukumura, D.; Padera, T.P.; Jain, R.K. Imaging steps of lymphatic metastasis reveals that vascular endothelial growth factor-C increases metastasis by increasing delivery of cancer cells to lymph nodes: Therapeutic implications. Cancer Res. 2006, 66, 8065–8075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karnezis, T.; Shayan, R.; Caesar, C.; Roufail, S.; Harris, N.C.; Ardipradja, K.; Zhang, Y.F.; Williams, S.P.; Farnsworth, R.H.; Chai, M.G.; et al. VEGF-D promotes tumor metastasis by regulating prostaglandins produced by the collecting lymphatic endothelium. Cancer Cell 2012, 21, 181–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garnier, L.; Gkountidi, A.O.; Hugues, S. Tumor-Associated Lymphatic Vessel Features and Immunomodulatory Functions. Front. Immunol. 2019, 10, 720. [Google Scholar] [CrossRef] [Green Version]
- Wilczak, W.; Wittmer, C.; Clauditz, T.; Minner, S.; Steurer, S.; Buscheck, F.; Krech, T.; Lennartz, M.; Harms, L.; Leleu, D.; et al. Marked Prognostic Impact of Minimal Lymphatic Tumor Spread in Prostate Cancer. Eur. Urol. 2018, 74, 376–386. [Google Scholar] [CrossRef] [PubMed]
- Stacker, S.A.; Williams, S.P.; Karnezis, T.; Shayan, R.; Fox, S.B.; Achen, M.G. Lymphangiogenesis and lymphatic vessel remodelling in cancer. Nat. Rev. Cancer 2014, 14, 159–172. [Google Scholar] [CrossRef]
- Dadras, S.S.; Paul, T.; Bertoncini, J.; Brown, L.F.; Muzikansky, A.; Jackson, D.G.; Ellwanger, U.; Garbe, C.; Mihm, M.C.; Detmar, M. Tumor lymphangiogenesis: A novel prognostic indicator for cutaneous melanoma metastasis and survival. Am. J. Pathol. 2003, 162, 1951–1960. [Google Scholar] [CrossRef]
- Tammela, T.; Saaristo, A.; Holopainen, T.; Yla-Herttuala, S.; Andersson, L.C.; Virolainen, S.; Immonen, I.; Alitalo, K. Photodynamic ablation of lymphatic vessels and intralymphatic cancer cells prevents metastasis. Sci. Transl. Med. 2011, 3, 69ra11. [Google Scholar] [CrossRef]
- Fransen, M.F.; Schoonderwoerd, M.; Knopf, P.; Camps, M.G.; Hawinkels, L.J.; Kneilling, M.; van Hall, T.; Ossendorp, F. Tumor-draining lymph nodes are pivotal in PD-1/PD-L1 checkpoint therapy. JCI Insight. 2018, 3, e124507. [Google Scholar] [CrossRef] [Green Version]
- Salmon, H.; Idoyaga, J.; Rahman, A.; Leboeuf, M.; Remark, R.; Jordan, S.; Casanova-Acebes, M.; Khudoynazarova, M.; Agudo, J.; Tung, N.; et al. Expansion and Activation of CD103(+) Dendritic Cell Progenitors at the Tumor Site Enhances Tumor Responses to Therapeutic PD-L1 and BRAF Inhibition. Immunity 2016, 44, 924–938. [Google Scholar] [CrossRef] [Green Version]
- Fankhauser, M.; Broggi, M.A.S.; Potin, L.; Bordry, N.; Jeanbart, L.; Lund, A.W.; Da Costa, E.; Hauert, S.; Rincon-Restrepo, M.; Tremblay, C.; et al. Tumor lymphangiogenesis promotes T cell infiltration and potentiates immunotherapy in melanoma. Sci. Transl. Med. 2017, 9, eaal4712. [Google Scholar] [CrossRef] [Green Version]
- Mlecnik, B.; Bindea, G.; Angell, H.K.; Maby, P.; Angelova, M.; Tougeron, D.; Church, S.E.; Lafontaine, L.; Fischer, M.; Fredriksen, T.; et al. Integrative Analyses of Colorectal Cancer Show Immunoscore Is a Stronger Predictor of Patient Survival Than Microsatellite Instability. Immunity 2016, 44, 698–711. [Google Scholar] [CrossRef] [Green Version]
- Card, C.M.; Yu, S.S.; Swartz, M.A. Emerging roles of lymphatic endothelium in regulating adaptive immunity. J. Clin. Investig. 2014, 124, 943–952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forster, R.; Davalos-Misslitz, A.C.; Rot, A. CCR7 and its ligands: Balancing immunity and tolerance. Nat. Rev. Immunol. 2008, 8, 362–371. [Google Scholar] [CrossRef] [PubMed]
- Xuan, W.; Qu, Q.; Zheng, B.; Xiong, S.; Fan, G.H. The chemotaxis of M1 and M2 macrophages is regulated by different chemokines. J. Leukoc. Biol. 2015, 97, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Wick, N.; Haluza, D.; Gurnhofer, E.; Raab, I.; Kasimir, M.T.; Prinz, M.; Steiner, C.W.; Reinisch, C.; Howorka, A.; Giovanoli, P.; et al. Lymphatic precollectors contain a novel, specialized subpopulation of podoplanin low, CCL27-expressing lymphatic endothelial cells. Am. J. Pathol. 2008, 173, 1202–1209. [Google Scholar] [CrossRef] [Green Version]
- Cohen, J.N.; Guidi, C.J.; Tewalt, E.F.; Qiao, H.; Rouhani, S.J.; Ruddell, A.; Farr, A.G.; Tung, K.S.; Engelhard, V.H. Lymph node-resident lymphatic endothelial cells mediate peripheral tolerance via Aire-independent direct antigen presentation. J. Exp. Med. 2010, 207, 681–688. [Google Scholar] [CrossRef] [Green Version]
- Salmi, M.; Jalkanen, S. Cell-surface enzymes in control of leukocyte trafficking. Nat. Rev. Immunol. 2005, 5, 760–771. [Google Scholar] [CrossRef]
- Podgrabinska, S.; Kamalu, O.; Mayer, L.; Shimaoka, M.; Snoeck, H.; Randolph, G.J.; Skobe, M. Inflamed lymphatic endothelium suppresses dendritic cell maturation and function via Mac-1/ICAM-1-dependent mechanism. J. Immunol. 2009, 183, 1767–1779. [Google Scholar] [CrossRef]
- Kihara, A. Sphingosine 1-phosphate is a key metabolite linking sphingolipids to glycerophospholipids. Biochim. Biophys. Acta 2014, 1841, 766–772. [Google Scholar] [CrossRef]
- Yanagida, K.; Hla, T. Vascular and Immunobiology of the Circulatory Sphingosine 1-Phosphate Gradient. Annu. Rev. Physiol. 2017, 79, 67–91. [Google Scholar] [CrossRef] [Green Version]
- Pham, T.H.; Baluk, P.; Xu, Y.; Grigorova, I.; Bankovich, A.J.; Pappu, R.; Coughlin, S.R.; McDonald, D.M.; Schwab, S.R.; Cyster, J.G. Lymphatic endothelial cell sphingosine kinase activity is required for lymphocyte egress and lymphatic patterning. J. Exp. Med. 2010, 207, 17–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendoza, A.; Fang, V.; Chen, C.; Serasinghe, M.; Verma, A.; Muller, J.; Chaluvadi, V.S.; Dustin, M.L.; Hla, T.; Elemento, O.; et al. Lymphatic endothelial S1P promotes mitochondrial function and survival in naive T cells. Nature 2017, 546, 158–161. [Google Scholar] [CrossRef] [PubMed]
- Hisano, Y.; Kono, M.; Cartier, A.; Engelbrecht, E.; Kano, K.; Kawakami, K.; Xiong, Y.; Piao, W.; Galvani, S.; Yanagida, K.; et al. Lysolipid receptor cross-talk regulates lymphatic endothelial junctions in lymph nodes. J. Exp. Med. 2019, 216, 1582–1598. [Google Scholar] [CrossRef] [PubMed]
- Jeltsch, M.; Tammela, T.; Alitalo, K.; Wilting, J. Genesis and pathogenesis of lymphatic vessels. Cell Tissue Res. 2003, 314, 69–84. [Google Scholar] [CrossRef]
- Bernier-Latmani, J.; Petrova, T.V. Intestinal lymphatic vasculature: Structure, mechanisms and functions. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 510–526. [Google Scholar] [CrossRef]
- Bernier-Latmani, J.; Cisarovsky, C.; Demir, C.S.; Bruand, M.; Jaquet, M.; Davanture, S.; Ragusa, S.; Siegert, S.; Dormond, O.; Benedito, R.; et al. DLL4 promotes continuous adult intestinal lacteal regeneration and dietary fat transport. J. Clin. Investig. 2015, 125, 4572–4586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abreu, M.T. Toll-like receptor signalling in the intestinal epithelium: How bacterial recognition shapes intestinal function. Nat. Rev. Immunol. 2010, 10, 131–144. [Google Scholar] [CrossRef] [PubMed]
- Hallback, D.A.; Jodal, M.; Mannischeff, M.; Lundgren, O. Tissue osmolality in intestinal villi of four mammals in vivo and in vitro. Acta Physiol. Scand. 1991, 143, 271–277. [Google Scholar] [CrossRef]
- Womack, W.A.; Barrowman, J.A.; Graham, W.H.; Benoit, J.N.; Kvietys, P.R.; Granger, D.N. Quantitative assessment of villous motility. Am. J. Physiol. 1987, 252, G250–G256. [Google Scholar] [CrossRef]
- Pabst, O.; Mowat, A.M. Oral tolerance to food protein. Mucosal. Immunol. 2012, 5, 232–239. [Google Scholar] [CrossRef]
- Cummings, R.J.; Barbet, G.; Bongers, G.; Hartmann, B.M.; Gettler, K.; Muniz, L.; Furtado, G.C.; Cho, J.; Lira, S.A.; Blander, J.M. Different tissue phagocytes sample apoptotic cells to direct distinct homeostasis programs. Nature 2016, 539, 565–569. [Google Scholar] [CrossRef]
- Esterhazy, D.; Canesso, M.C.C.; Mesin, L.; Muller, P.A.; de Castro, T.B.R.; Lockhart, A.; ElJalby, M.; Faria, A.M.C.; Mucida, D. Compartmentalized gut lymph node drainage dictates adaptive immune responses. Nature 2019, 569, 126–130. [Google Scholar] [CrossRef] [PubMed]
- D’Alessio, S.; Tacconi, C.; Danese, S. Targeting lymphatics in inflammatory bowel disease. Oncotarget 2015, 6, 34047–34048. [Google Scholar] [CrossRef] [PubMed]
- Heatley, R.V.; Bolton, P.M.; Hughes, L.E.; Owen, E.W. Mesenteric lymphatic obstruction in Crohn’s disease. Digestion 1980, 20, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Pedica, F.; Ligorio, C.; Tonelli, P.; Bartolini, S.; Baccarini, P. Lymphangiogenesis in Crohn’s disease: An immunohistochemical study using monoclonal antibody D2-40. Virchows Arch. 2008, 452, 57–63. [Google Scholar] [CrossRef]
- Geleff, S.; Schoppmann, S.F.; Oberhuber, G. Increase in podoplanin-expressing intestinal lymphatic vessels in inflammatory bowel disease. Virchows Arch. 2003, 442, 231–237. [Google Scholar] [CrossRef]
- Thaunat, O.; Kerjaschki, D.; Nicoletti, A. Is defective lymphatic drainage a trigger for lymphoid neogenesis? Trends Immunol. 2006, 27, 441–445. [Google Scholar] [CrossRef]
- von der Weid, P.Y.; Rehal, S.; Ferraz, J.G. Role of the lymphatic system in the pathogenesis of Crohn’s disease. Curr. Opin. Gastroenterol. 2011, 27, 335–341. [Google Scholar] [CrossRef]
- Randolph, G.J.; Bala, S.; Rahier, J.F.; Johnson, M.W.; Wang, P.L.; Nalbantoglu, I.; Dubuquoy, L.; Chau, A.; Pariente, B.; Kartheuser, A.; et al. Lymphoid Aggregates Remodel Lymphatic Collecting Vessels that Serve Mesenteric Lymph Nodes in Crohn Disease. Am. J. Pathol. 2016, 186, 3066–3073. [Google Scholar] [CrossRef] [Green Version]
- Plog, B.A.; Nedergaard, M. The Glymphatic System in Central Nervous System Health and Disease: Past, Present, and Future. Annu. Rev. Pathol. 2018, 13, 379–394. [Google Scholar] [CrossRef] [Green Version]
- Iliff, J.J.; Wang, M.; Liao, Y.; Plogg, B.A.; Peng, W.; Gundersen, G.A.; Benveniste, H.; Vates, G.E.; Deane, R.; Goldman, S.A.; et al. A paravascular pathway facilitates CSF flow through the brain parenchyma and the clearance of interstitial solutes, including amyloid beta. Sci. Transl. Med. 2012, 4, 147ra111. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, M.K.; Mestre, H.; Nedergaard, M. The glymphatic pathway in neurological disorders. Lancet Neurol. 2018, 17, 1016–1024. [Google Scholar] [CrossRef] [Green Version]
- Aspelund, A.; Antila, S.; Proulx, S.T.; Karlsen, T.V.; Karaman, S.; Detmar, M.; Wiig, H.; Alitalo, K. A dural lymphatic vascular system that drains brain interstitial fluid and macromolecules. J. Exp. Med. 2015, 212, 991–999. [Google Scholar] [CrossRef] [PubMed]
- Da Mesquita, S.; Louveau, A.; Vaccari, A.; Smirnov, I.; Cornelison, R.C.; Kingsmore, K.M.; Contarino, C.; Onengut-Gumuscu, S.; Farber, E.; Raper, D.; et al. Functional aspects of meningeal lymphatics in ageing and Alzheimer’s disease. Nature 2018, 560, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Absinta, M.; Ha, S.K.; Nair, G.; Sati, P.; Luciano, N.J.; Palisoc, M.; Louveau, A.; Zaghloul, K.A.; Pittaluga, S.; Kipnis, J.; et al. Human and nonhuman primate meninges harbor lymphatic vessels that can be visualized noninvasively by MRI. Elife 2017, 6, e29738. [Google Scholar] [CrossRef] [PubMed]
- Visanji, N.P.; Lang, A.E.; Munoz, D.G. Lymphatic vasculature in human dural superior sagittal sinus: Implications for neurodegenerative proteinopathies. Neurosci. Lett. 2018, 665, 18–21. [Google Scholar] [CrossRef]
- Antila, S.; Karaman, S.; Nurmi, H.; Airavaara, M.; Voutilainen, M.H.; Mathivet, T.; Chilov, D.; Li, Z.; Koppinen, T.; Park, J.H.; et al. Development and plasticity of meningeal lymphatic vessels. J. Exp. Med. 2017, 214, 3645–3667. [Google Scholar] [CrossRef]
- das Neves, S.P.; Delivanoglou, N.; Da Mesquita, S. CNS-Draining Meningeal Lymphatic Vasculature: Roles, Conundrums and Future Challenges. Front. Pharmacol. 2021, 12, 655052. [Google Scholar] [CrossRef]
- Ma, Q.; Ineichen, B.V.; Detmar, M.; Proulx, S.T. Outflow of cerebrospinal fluid is predominantly through lymphatic vessels and is reduced in aged mice. Nat. Commun. 2017, 8, 1434. [Google Scholar] [CrossRef] [Green Version]
- Ahn, J.H.; Cho, H.; Kim, J.H.; Kim, S.H.; Ham, J.S.; Park, I.; Suh, S.H.; Hong, S.P.; Song, J.H.; Hong, Y.K.; et al. Meningeal lymphatic vessels at the skull base drain cerebrospinal fluid. Nature 2019, 572, 62–66. [Google Scholar] [CrossRef]
- Melin, E.; Eide, P.K.; Ringstad, G. In vivo assessment of cerebrospinal fluid efflux to nasal mucosa in humans. Sci. Rep. 2020, 10, 14974. [Google Scholar] [CrossRef] [PubMed]
- Louveau, A.; Herz, J.; Alme, M.N.; Salvador, A.F.; Dong, M.Q.; Viar, K.E.; Herod, S.G.; Knopp, J.; Setliff, J.C.; Lupi, A.L.; et al. CNS lymphatic drainage and neuroinflammation are regulated by meningeal lymphatic vasculature. Nat. Neurosci. 2018, 21, 1380–1391. [Google Scholar] [CrossRef] [PubMed]
- Song, E.; Mao, T.; Dong, H.; Boisserand, L.S.B.; Antila, S.; Bosenberg, M.; Alitalo, K.; Thomas, J.L.; Iwasaki, A. VEGF-C-driven lymphatic drainage enables immunosurveillance of brain tumours. Nature 2020, 577, 689–694. [Google Scholar] [CrossRef]
- Da Mesquita, S.; Papadopoulos, Z.; Dykstra, T.; Brase, L.; Farias, F.G.; Wall, M.; Jiang, H.; Kodira, C.D.; de Lima, K.A.; Herz, J.; et al. Meningeal lymphatics affect microglia responses and anti-Abeta immunotherapy. Nature 2021, 593, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Patel, T.K.; Habimana-Griffin, L.; Gao, X.; Xu, B.; Achilefu, S.; Alitalo, K.; McKee, C.A.; Sheehan, P.W.; Musiek, E.S.; Xiong, C.; et al. Dural lymphatics regulate clearance of extracellular tau from the CNS. Mol. Neurodegener. 2019, 14, 11. [Google Scholar] [CrossRef]
- Zou, W.; Pu, T.; Feng, W.; Lu, M.; Zheng, Y.; Du, R.; Xiao, M.; Hu, G. Blocking meningeal lymphatic drainage aggravates Parkinson’s disease-like pathology in mice overexpressing mutated alpha-synuclein. Transl. Neurodegener. 2019, 8, 7. [Google Scholar] [CrossRef]
- Ding, X.B.; Wang, X.X.; Xia, D.H.; Liu, H.; Tian, H.Y.; Fu, Y.; Chen, Y.K.; Qin, C.; Wang, J.Q.; Xiang, Z.; et al. Impaired meningeal lymphatic drainage in patients with idiopathic Parkinson’s disease. Nat. Med. 2021, 27, 411–418. [Google Scholar] [CrossRef]
- Bolte, A.C.; Dutta, A.B.; Hurt, M.E.; Smirnov, I.; Kovacs, M.A.; McKee, C.A.; Ennerfelt, H.E.; Shapiro, D.; Nguyen, B.H.; Frost, E.L.; et al. Meningeal lymphatic dysfunction exacerbates traumatic brain injury pathogenesis. Nat. Commun. 2020, 11, 4524. [Google Scholar] [CrossRef]
- Hsu, S.J.; Zhang, C.; Jeong, J.; Lee, S.I.; McConnell, M.; Utsumi, T.; Iwakiri, Y. Enhanced Meningeal Lymphatic Drainage Ameliorates Neuroinflammation and Hepatic Encephalopathy in Cirrhotic Rats. Gastroenterology 2021, 160, 1315–1329.e1313. [Google Scholar] [CrossRef]
- Hoggan, G.; Hoggan, F.E. The Lymphatics of the Walls of the Larger Blood-Vessels and Lymphatics. J. Anat. Physiol. 1882, 17, 1–23. [Google Scholar]
- Drozdz, K.; Janczak, D.; Dziegiel, P.; Podhorska, M.; Patrzalek, D.; Ziolkowski, P.; Andrzejak, R.; Szuba, A. Adventitial lymphatics of internal carotid artery in healthy and atherosclerotic vessels. Folia Histochem. Cytobiol. 2008, 46, 433–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, R.A. Lymphatics of blood vessels. Lymphology 1969, 2, 44–56. [Google Scholar] [PubMed]
- Miller, A.J.; DeBoer, A.; Palmer, A. The role of the lymphatic system in coronary atherosclerosis. Med. Hypotheses 1992, 37, 31–36. [Google Scholar] [CrossRef]
- Nakano, T.; Nakashima, Y.; Yonemitsu, Y.; Sumiyoshi, S.; Chen, Y.X.; Akishima, Y.; Ishii, T.; Iida, M.; Sueishi, K. Angiogenesis and lymphangiogenesis and expression of lymphangiogenic factors in the atherosclerotic intima of human coronary arteries. Hum. Pathol. 2005, 36, 330–340. [Google Scholar] [CrossRef] [PubMed]
- Sacchi, G.; Weber, E.; Comparini, L. Histological framework of lymphatic vasa vasorum of major arteries: An experimental study. Lymphology 1990, 23, 135–139. [Google Scholar] [PubMed]
- Martel, C.; Li, W.; Fulp, B.; Platt, A.M.; Gautier, E.L.; Westerterp, M.; Bittman, R.; Tall, A.R.; Chen, S.H.; Thomas, M.J.; et al. Lymphatic vasculature mediates macrophage reverse cholesterol transport in mice. J. Clin. Investig. 2013, 123, 1571–1579. [Google Scholar] [CrossRef] [Green Version]
- Vuorio, T.; Nurmi, H.; Moulton, K.; Kurkipuro, J.; Robciuc, M.R.; Ohman, M.; Heinonen, S.E.; Samaranayake, H.; Heikura, T.; Alitalo, K.; et al. Lymphatic vessel insufficiency in hypercholesterolemic mice alters lipoprotein levels and promotes atherogenesis. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 1162–1170. [Google Scholar] [CrossRef] [Green Version]
- Lim, H.Y.; Thiam, C.H.; Yeo, K.P.; Bisoendial, R.; Hii, C.S.; McGrath, K.C.; Tan, K.W.; Heather, A.; Alexander, J.S.; Angeli, V. Lymphatic vessels are essential for the removal of cholesterol from peripheral tissues by SR-BI-mediated transport of HDL. Cell Metab. 2013, 17, 671–684. [Google Scholar] [CrossRef] [Green Version]
- Milasan, A.; Jean, G.; Dallaire, F.; Tardif, J.C.; Merhi, Y.; Sorci-Thomas, M.; Martel, C. Apolipoprotein A-I Modulates Atherosclerosis Through Lymphatic Vessel-Dependent Mechanisms in Mice. J. Am. Heart Assoc. 2017, 6, e006892. [Google Scholar] [CrossRef]
- Milasan, A.; Smaani, A.; Martel, C. Early rescue of lymphatic function limits atherosclerosis progression in Ldlr-/- mice. Atherosclerosis 2019, 283, 106–119. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.H.; Elvington, A.; Randolph, G.J. The role of the lymphatic system in cholesterol transport. Front. Pharmacol. 2015, 6, 182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rademakers, T.; van der Vorst, E.P.; Daissormont, I.T.; Otten, J.J.; Theodorou, K.; Theelen, T.L.; Gijbels, M.; Anisimov, A.; Nurmi, H.; Lindeman, J.H.; et al. Adventitial lymphatic capillary expansion impacts on plaque T cell accumulation in atherosclerosis. Sci. Rep. 2017, 7, 45263. [Google Scholar] [CrossRef]
- Alitalo, K. The lymphatic vasculature in disease. Nat. Med. 2011, 17, 1371–1380. [Google Scholar] [CrossRef]
- Dongaonkar, R.M.; Stewart, R.H.; Geissler, H.J.; Laine, G.A. Myocardial microvascular permeability, interstitial oedema, and compromised cardiac function. Cardiovasc. Res. 2010, 87, 331–339. [Google Scholar] [CrossRef] [PubMed]
- Feola, M.; Lefer, A.M. Alterations in cardiac lymph dynamics in acute myocardial ischemia in dogs. J. Surg. Res. 1977, 23, 299–305. [Google Scholar] [CrossRef]
- Henri, O.; Pouehe, C.; Houssari, M.; Galas, L.; Nicol, L.; Edwards-Levy, F.; Henry, J.P.; Dumesnil, A.; Boukhalfa, I.; Banquet, S.; et al. Selective Stimulation of Cardiac Lymphangiogenesis Reduces Myocardial Edema and Fibrosis Leading to Improved Cardiac Function Following Myocardial Infarction. Circulation 2016, 133, 1484–1497, discussion 1497. [Google Scholar] [CrossRef] [Green Version]
- Klotz, L.; Norman, S.; Vieira, J.M.; Masters, M.; Rohling, M.; Dube, K.N.; Bollini, S.; Matsuzaki, F.; Carr, C.A.; Riley, P.R. Cardiac lymphatics are heterogeneous in origin and respond to injury. Nature 2015, 522, 62–67. [Google Scholar] [CrossRef] [Green Version]
- Vieira, J.M.; Norman, S.; Villa Del Campo, C.; Cahill, T.J.; Barnette, D.N.; Gunadasa-Rohling, M.; Johnson, L.A.; Greaves, D.R.; Carr, C.A.; Jackson, D.G.; et al. The cardiac lymphatic system stimulates resolution of inflammation following myocardial infarction. J. Clin. Investig. 2018, 128, 3402–3412. [Google Scholar] [CrossRef]
- Liu, X.; De la Cruz, E.; Gu, X.; Balint, L.; Oxendine-Burns, M.; Terrones, T.; Ma, W.; Kuo, H.H.; Lantz, C.; Bansal, T.; et al. Lymphoangiocrine signals promote cardiac growth and repair. Nature 2020, 588, 705–711. [Google Scholar] [CrossRef]
- Oliver, G.; Kipnis, J.; Randolph, G.J.; Harvey, N.L. The Lymphatic Vasculature in the 21(st) Century: Novel Functional Roles in Homeostasis and Disease. Cell 2020, 182, 270–296. [Google Scholar] [CrossRef]
- Rockson, S.G. Lymphedema. Am. J. Med. 2001, 110, 288–295. [Google Scholar] [CrossRef]
- Lourens, G.B.; Ferrell, D.K. Lymphatic Filariasis. Nurs. Clin. N. Am. 2019, 54, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Witte, M.H.; Bernas, M.J.; Martin, C.P.; Witte, C.L. Lymphangiogenesis and lymphangiodysplasia: From molecular to clinical lymphology. Microsc. Res. Tech. 2001, 55, 122–145. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Role of ROS in regulating lymphangiogenesis and lymphatic function. Activation of VEGFR3 present in LECs by its ligand VEGF-C induces optimal Nox4-derived H2O2 production, which in turn enhances VEGFR3 autophosphorylation and stimulates downstream pro-lymphangiogenic signaling (upper panel). Oxidized LDL and RSPO2 inhibit lymphangiogenesis via suppression of Akt/eNOS pathway. Under diabetic condition, excessive H2O2 generation elevates epsin expression and promotes VEGFR3 degradation, leading to attenuated lymphangiogenesis and reduced lymphatic transport. In inflammatory condition, supra-physiological NO production by CD11b+ myeloid immune cells surrounding LVs contributes to nitrosative stress leading to the suppression of lymphatic contractions and inducing LV leakiness (lower panel).
Figure 1.
Role of ROS in regulating lymphangiogenesis and lymphatic function. Activation of VEGFR3 present in LECs by its ligand VEGF-C induces optimal Nox4-derived H2O2 production, which in turn enhances VEGFR3 autophosphorylation and stimulates downstream pro-lymphangiogenic signaling (upper panel). Oxidized LDL and RSPO2 inhibit lymphangiogenesis via suppression of Akt/eNOS pathway. Under diabetic condition, excessive H2O2 generation elevates epsin expression and promotes VEGFR3 degradation, leading to attenuated lymphangiogenesis and reduced lymphatic transport. In inflammatory condition, supra-physiological NO production by CD11b+ myeloid immune cells surrounding LVs contributes to nitrosative stress leading to the suppression of lymphatic contractions and inducing LV leakiness (lower panel).

Figure 2.
Role of the lymphatic system in health and disease.


{kind=link}
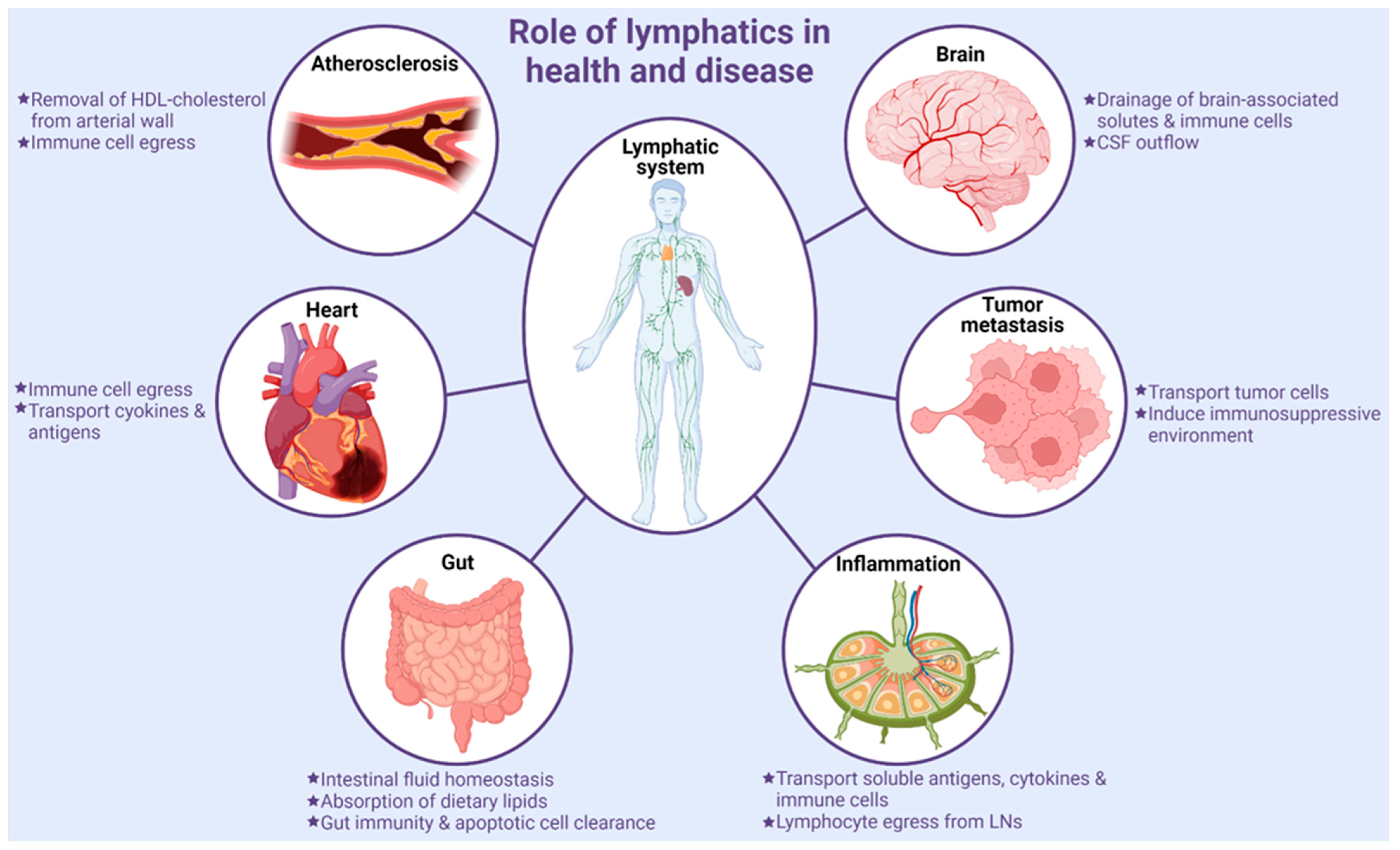{kind=link}
Table 1.
Types and generation of reactive oxygen species.
| Name of Molecule | Half-Life of Molecule | Generation of Molecule |
|---|---|---|
| Nitric oxide (NO•) | 10−5 to 10−3 s | Nitric oxide synthase L-arginine + O2 + NADPH → L-citrulline + NO• + NADP+ Reduction of nitrite Deoxyhaemoglobin/myoglobin NO2− + Fe2+ + H+ → NO• + Fe3+ + OH− Xanthine oxidoreductase NO2− + Mo4+ + H+ → NO• + Mo5+ + OH− Protons NO2− + H+ → HNO2 2 HNO2 → 2 N2O3 + H2O N2O3 → NO• + •NO2 Ascorbate NO2− + H+ → HNO2 2 HNO2 + Asc → 2 NO• + dehydroAsc + 2 H2O Polyphenols (Ph-OH) NO2− + H+ → HNO2 Ph-OH + HNO2 → Ph-•O + NO• + H2O |
| Superoxide (O2•−) | 10−11 to 10−9 s | NADPH oxidase NADPH + 2O2 → NADP+ + 2O2•− + 2H− Xanthine oxidase Hypoxanthine + H2O + 2O2 → Xanthine + 2O2•− + 2H− Xanthine + H2O + 2O2 → Uric acid + 2O2•− + 2H− Uncoupled endothelial nitric oxide synthase NADPH + 2O2 → NADP+ + 2O2•− + 2H− Mitochondrial electron transport chain complexes I and III O2 → O2•− Lipooxygenase Arachidonic acid + O2 → HPETE+ O2•− |
| Hydroxyl radical (•OH) | 10−9 s | Fenton reaction Fe2+ + H2O2 → Fe3+ + •OH + OH− Haber-Weiss reaction •O2− + H2O2 → •OH + OH− + O2 HOONO → •OH + NO2• |
| Lipid peroxyl radical (LOO•) | 7 s | L-H + •X → L•+ XH LOO• + L-H → LOOH + L• L• + O2 → LOO• L-H: polyunsaturated fatty acid •X: oxidizing character (i.e., •OH or O2•−) L•: lipid radical |
| Peroxynitrite (ONOO−) | 10−2 s | O2•− + NO• → ONOO− |
| Hydrogen peroxide (H2O2) | 10−8 (in presence of catalase) or 10−3 s | 2O2•− + 2H+ → H2O2 + O2 |
| Hypochlorous acid (HOCl) | <1 min | Myeloperoxidase H2O2 + Cl− → HOCl + OH− |
| Ozone (O3) | 1 min | x1O2 + yH2O  [H2O3(y−1)H2O] → H2O2 + (x−1)3O2 + O3 [H2O3(y−1)H2O] → H2O2 + (x−1)3O2 + O3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Singla, B.; Aithabathula, R.V.; Kiran, S.; Kapil, S.; Kumar, S.; Singh, U.P. Reactive Oxygen Species in Regulating Lymphangiogenesis and Lymphatic Function. Cells 2022, 11, 1750. https://doi.org/10.3390/cells11111750
AMA Style
Singla B, Aithabathula RV, Kiran S, Kapil S, Kumar S, Singh UP. Reactive Oxygen Species in Regulating Lymphangiogenesis and Lymphatic Function. Cells. 2022; 11(11):1750. https://doi.org/10.3390/cells11111750
Chicago/Turabian StyleSingla, Bhupesh, Ravi Varma Aithabathula, Sonia Kiran, Shweta Kapil, Santosh Kumar, and Udai P. Singh. 2022. "Reactive Oxygen Species in Regulating Lymphangiogenesis and Lymphatic Function" Cells 11, no. 11: 1750. https://doi.org/10.3390/cells11111750
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.