
An Early Disturbance in Serotonergic Neurotransmission Contributes to the Onset of Parkinsonian Phenotypes in Drosophila melanogaster

 , , , , , and
, , , , , and 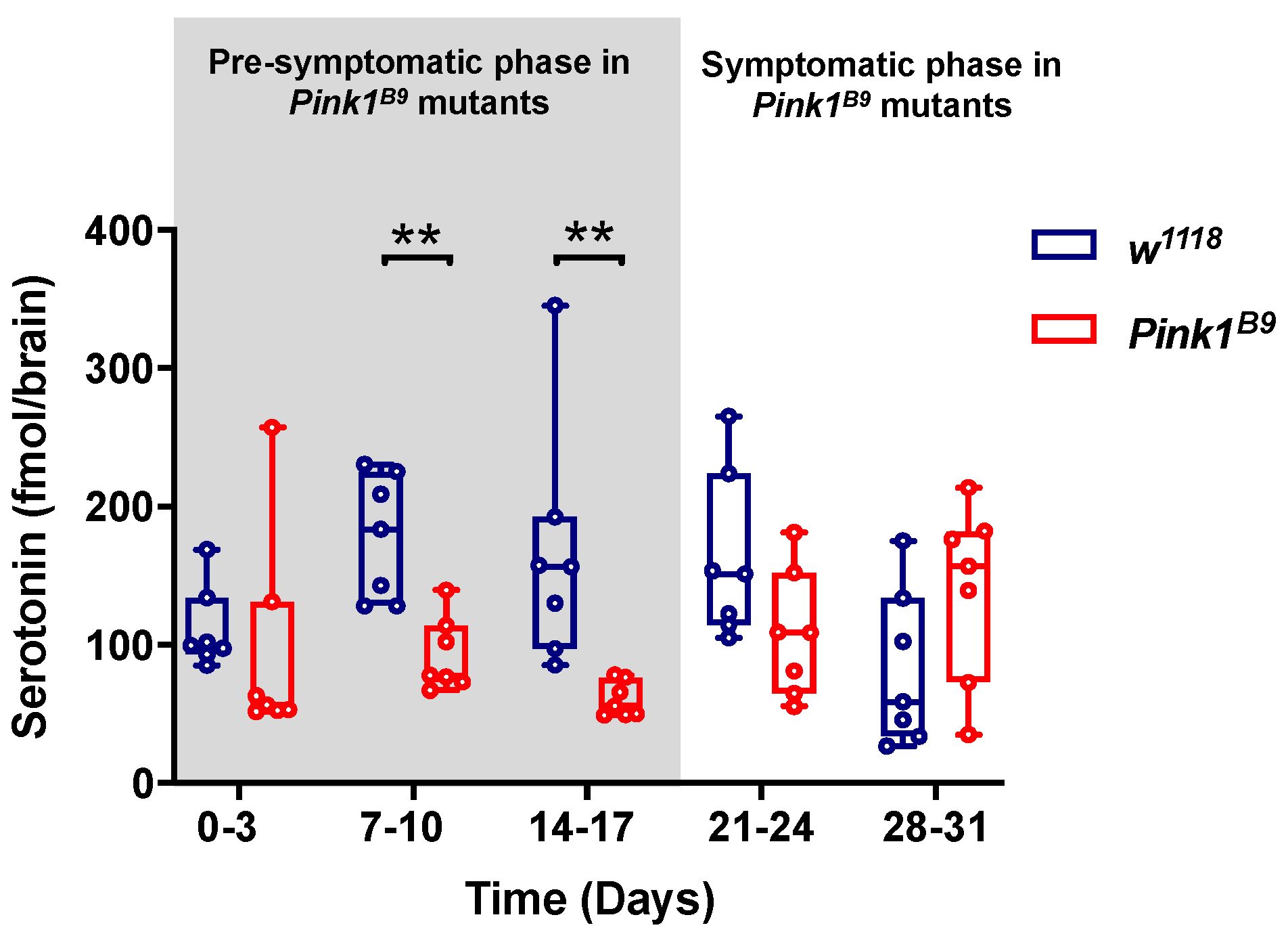{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fly Stock and Maintenance
2.2. Bioethical and Biosafety Issues
2.3. Serotonin Measurement in Fly Brains by HPLC
2.4. Fast-Scan Cyclic Voltammetry (FSCV)
2.5. Drug Exposure
2.6. Video Recording of Motor Behavior
2.7. Immunofluorescence
2.8. Imaging and Quantification of Trh and TH-Positive Cells
2.9. Western Blot
2.10. Real-Time Quantitative PCR
2.11. Statistical Analysis
3. Results
3.1. Pink1B9 Mutant Flies Exhibit Lower 5-HT Brain Content during the Pre-Symptomatic Phase
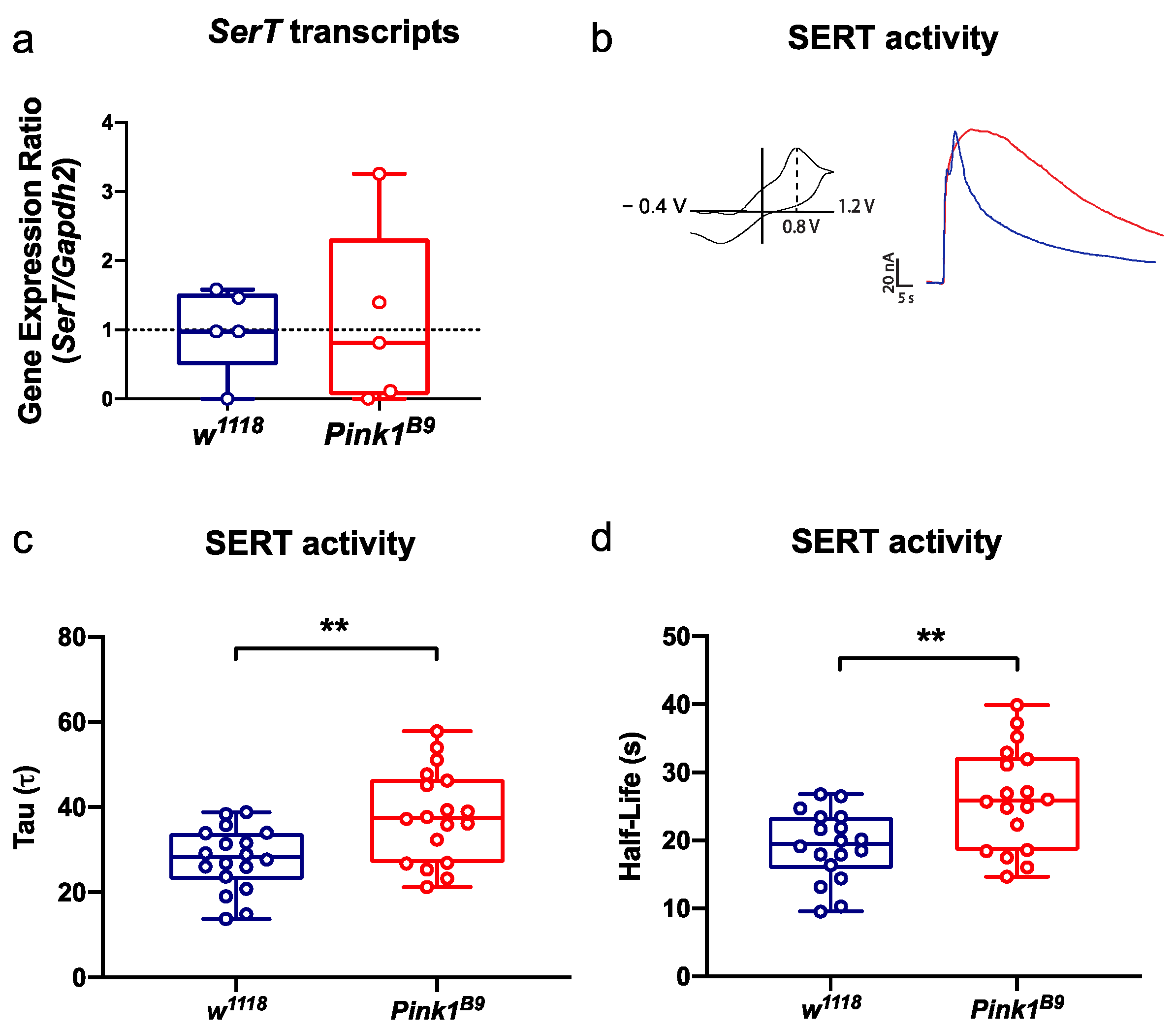
3.2. SERT Activity Declines in Pink1B9 Mutant Flies during the Pre-Symptomatic Phase
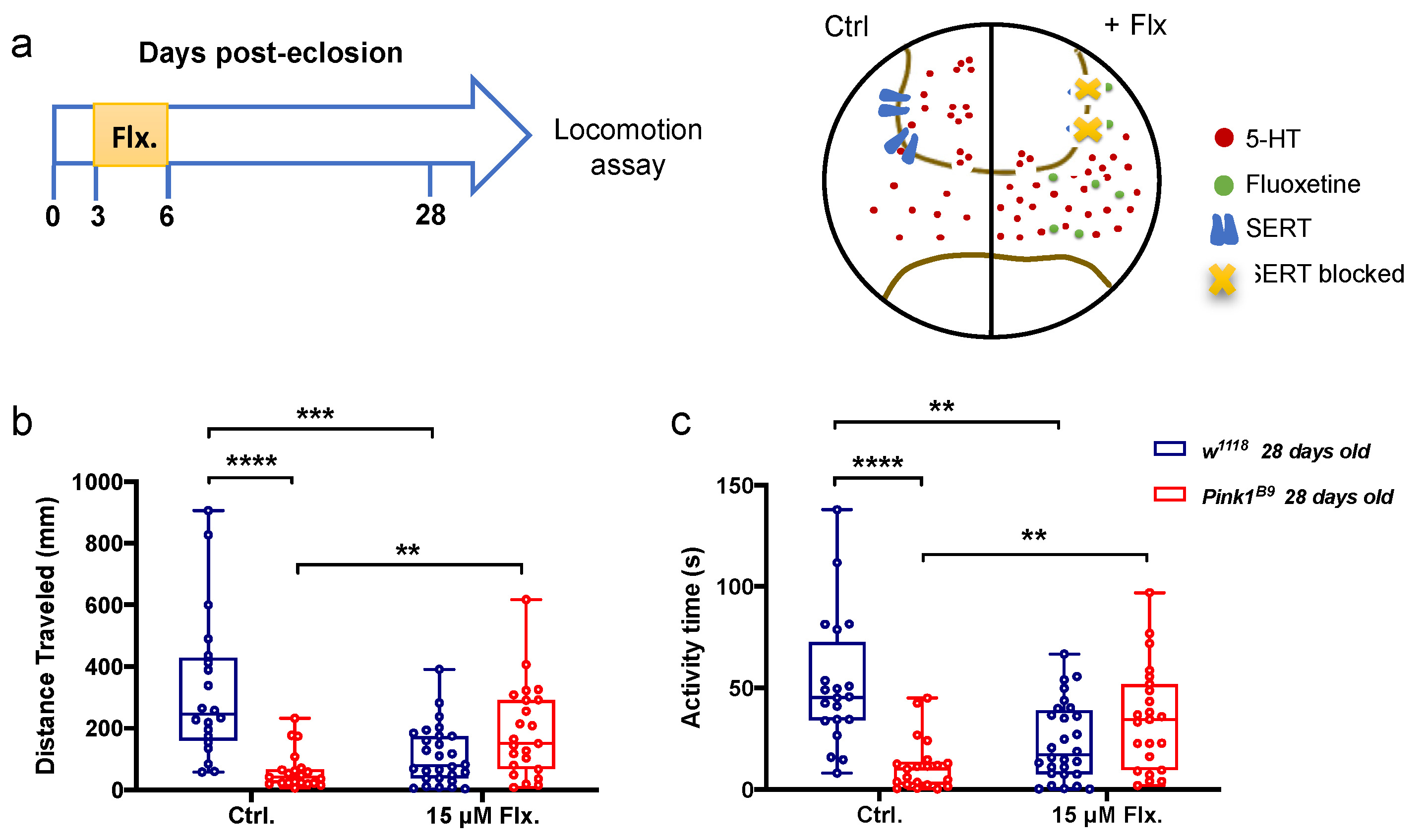
3.3. A Transient Increase in Serotonergic Signaling in Young Control and Pink1B9 Mutant Flies: Opposite Effects on Locomotion
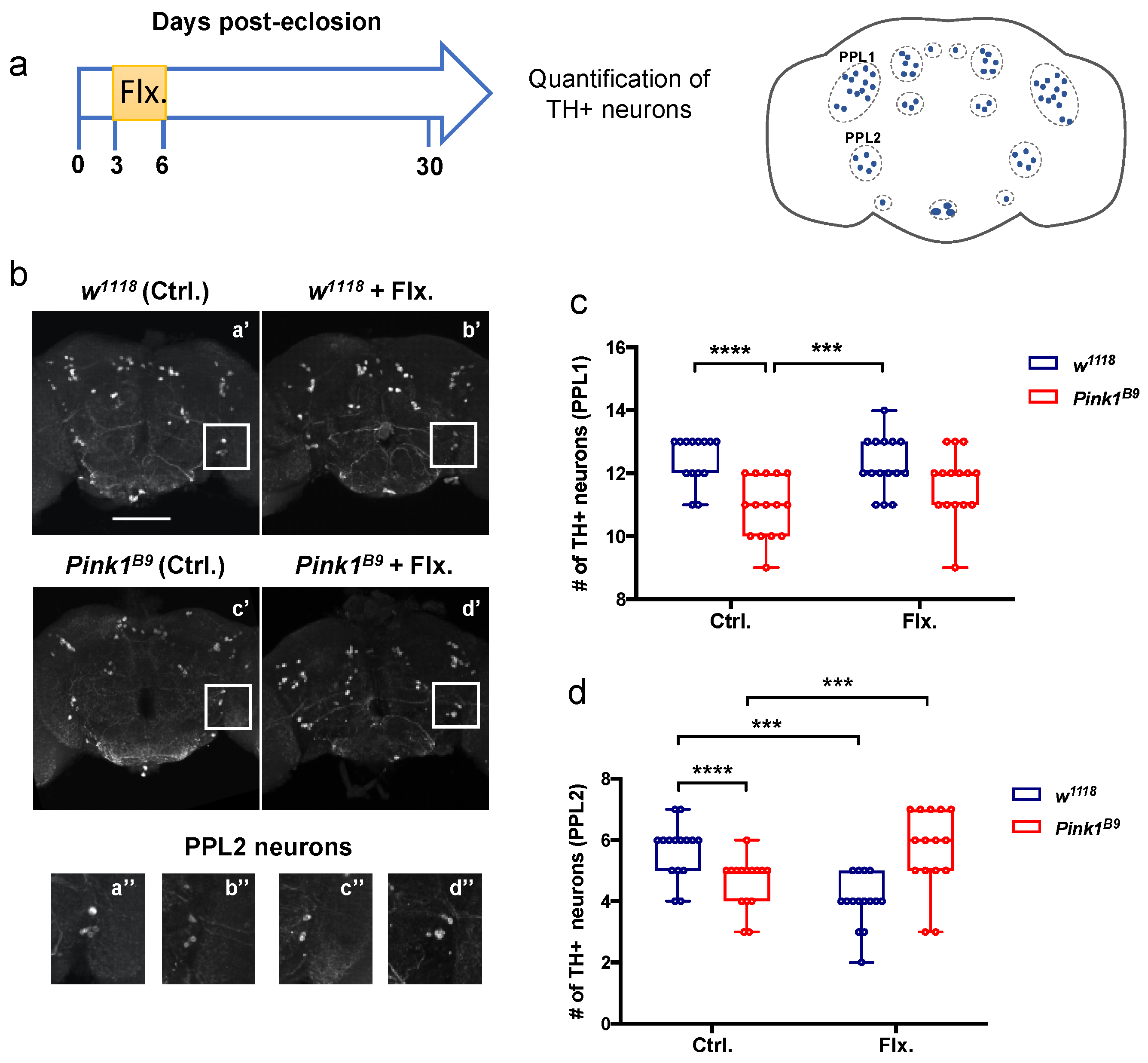
3.4. A Transient Increase in Serotonergic Signaling in Young Control and Pink1B9 Mutant Flies: Opposite Effects on Survival of PPL2 Dopaminergic Neurons
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bernheimer, H.; Birkmayer, W.; Hornykiewicz, O.; Jellinger, K.; Seitelberger, F. Brain dopamine and the syndromes of Parkinson and Huntington Clinical, morphological and neurochemical correlations. J. Neurol. Sci. 1973, 20, 415–455. [Google Scholar] [CrossRef]
- Kish, S.J.; Shannak, K.; Hornykiewicz, O. Uneven Pattern of Dopamine Loss in the Striatum of Patients with Idiopathic Parkinson’s Disease. N. Engl. J. Med. 1988, 318, 876–880. [Google Scholar] [CrossRef] [PubMed]
- Kalia, L.V.; Lang, A.E. Parkinson’s disease. Lancet 2015, 386, 896–912. [Google Scholar] [CrossRef]
- Postuma, R.B.; Berg, D. Prodromal Parkinson’s Disease: The Decade Past, the Decade to Come. Mov. Disord. 2019, 34, 665–675. [Google Scholar] [CrossRef]
- Buddhala, C.; Loftin, S.K.; Kuley, B.M.; Cairns, N.J.; Campbell, M.C.; Perlmutter, J.S.; Kotzbauer, P.T. Dopaminergic, serotonergic, and noradrenergic deficits in Parkinson disease. Ann. Clin. Transl. Neurol. 2015, 2, 949–959. [Google Scholar] [CrossRef]
- Politis, M.; Niccolini, F. Serotonin in Parkinson’s disease. Behav. Brain Res. 2015, 277, 136–145. [Google Scholar] [CrossRef]
- Doder, M.; Rabiner, E.A.; Turjanski, N.; Lees, A.J.; Brooks, D.J. Tremor in Parkinson’s disease and serotonergic dysfunction. Neurology 2003, 60, 601–605. [Google Scholar] [CrossRef]
- Loane, C.; Wu, K.; Bain, P.; David, J.; Piccini, P.; Politis, M. Serotonergic loss in motor circuitries correlates with severity of action-postural tremor in PD. Neurology 2013, 80, 1850–1855. [Google Scholar] [CrossRef] [Green Version]
- Kish, S.J.; Tong, J.; Hornykiewicz, O.; Rajput, A.; Chang, L.J.; Guttman, M.; Furukawa, Y. Preferential loss of serotonin markers in caudate versus putamen in Parkinson’s disease. Brain 2008, 131, 120–131. [Google Scholar] [CrossRef]
- Pasquini, J.; Ceravolo, R.; Qamhawi, Z.; Lee, J.Y.; Deuschl, G.; Brooks, D.J.; Bonuccelli, U.; Pavese, N. Progression of tremor in early stages of Parkinson’s disease: A clinical and neuroimaging study. Brain 2018, 141, 811–821. [Google Scholar] [CrossRef] [Green Version]
- Wile, D.J.; Agarwal, P.A.; Schulzer, M.; Mak, E.; Dinelle, K.; Shahinfard, E.; Vafai, N.; Hasegawa, K.; Zhang, J.; McKenzie, J.; et al. Serotonin and dopamine transporter PET changes in the premotor phase of LRRK2 parkinsonism: Cross-sectional studies. Lancet Neurol. 2017, 16, 351–359. [Google Scholar] [CrossRef] [Green Version]
- Wilson, H.; Dervenoulas, G.; Pagano, G.; Koros, C.; Yousaf, T.; Picillo, M.; Polychronis, S.; Simitsi, A.; Giordano, B.; Chappell, Z.; et al. Serotonergic pathology and disease burden in the premotor and motor phase of A53T α-synuclein parkinsonism: A cross-sectional study. Lancet Neurol. 2019, 18, 748–759. [Google Scholar] [CrossRef] [Green Version]
- Azmitia, E.C.; Segal, M. An autoradiographic analysis of the differential ascending projections of the dorsal and median raphe nuclei in the rat. J. Comp. Neurol. 1978, 179, 641–667. [Google Scholar] [CrossRef] [PubMed]
- Di Giovanni, G.; Di Matteo, V.; Pierucci, M.; Esposito, E. Serotonin-dopamine interaction: Electrophysiological evidence. Prog. Brain Res. 2008, 172, 45–71. [Google Scholar]
- Navailles, S.; De Deurwaerdère, P. Presynaptic control of serotonin on striatal dopamine function. Psychopharmacology 2011, 213, 213–242. [Google Scholar] [CrossRef]
- Clark, I.E.; Dodson, M.W.; Jiang, C.; Cao, J.H.; Huh, J.R.; Seol, J.H.; Yoo, S.J.; Hay, B.A.; Guo, M. Drosophila pink1 is required for mitochondrial function and interacts genetically with parkin. Nature 2006, 441, 1162–1166. [Google Scholar] [CrossRef]
- Park, J.; Lee, S.B.; Lee, S.; Kim, Y.; Song, S.; Kim, S.; Bae, E.; Kim, J.; Shong, M.; Kim, J.M.; et al. Mitochondrial dysfunction in Drosophila PINK1 mutants is complemented by parkin. Nature 2006, 441, 1157–1161. [Google Scholar] [CrossRef]
- Matsuda, N.; Sato, S.; Shiba, K.; Okatsu, K.; Saisho, K.; Gautier, C.A.; Sou, Y.; Saiki, S.; Kawajiri, S.; Sato, F.; et al. PINK1 stabilized by mitochondrial depolarization recruits Parkin to damaged mitochondria and activates latent Parkin for mitophagy. J. Cell Biol. 2010, 189, 211. [Google Scholar] [CrossRef]
- Vives-Bauza, C.; Zhou, C.; Huang, Y.; Cui, M.; de Vries, R.L.A.; Kim, J.; May, J.; Tocilescu, M.A.; Liu, W.; Ko, H.S.; et al. PINK1-dependent recruitment of Parkin to mitochondria in mitophagy. Proc. Natl. Acad. Sci. USA 2010, 107, 378–383. [Google Scholar] [CrossRef] [Green Version]
- Molina-Mateo, D.; Fuenzalida-Uribe, N.; Hidalgo, S.; Molina-Fernández, C.; Abarca, J.; Zárate, R.V.; Escandón, M.; Figueroa, R.; Tevy, M.F.; Campusano, J.M. Characterization of a presymptomatic stage in a Drosophila Parkinson’s disease model: Unveiling dopaminergic compensatory mechanisms. Biochim. Biophys. Acta-Mol. Basis Dis. 2017, 1863, 2882–2890. [Google Scholar] [CrossRef]
- Hidalgo, S.; Castro, C.; Zárate, R.V.; Valderrama, B.P.; Hodge, J.J.L.; Campusano, J.M. The behavioral and neurochemical characterization of a Drosophila dysbindin mutant supports the contribution of serotonin to schizophrenia negative symptoms. Neurochem. Int. 2020, 138, 104753. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, S.; Molina-Mateo, D.; Escobedo, P.; Zárate, R.V.; Fritz, E.; Fierro, A.; Perez, E.G.; Iturriaga-Vasquez, P.; Reyes-Parada, M.; Varas, R.; et al. Characterization of a Novel Drosophila SERT Mutant: Insights on the Contribution of the Serotonin Neural System to Behaviors. ACS Chem. Neurosci. 2017, 8, 2168–2179. [Google Scholar] [CrossRef] [PubMed]
- Majeed, Z.R.; Ritter, K.; Robinson, J.; Blümich, S.L.E.; Brailoiu, E.; Cooper, R.L. New insights into the acute actions from a high dosage of fluoxetine on neuronal and cardiac function: Drosophila, crayfish and rodent models. Comp. Biochem. Physiol. Part-C Toxicol. Pharmacol. 2015, 176–177, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Bao, X.; Wang, B.; Zhang, J.; Yan, T.; Yang, W.; Jiao, F.; Liu, J.; Wang, S. Localization of serotonin/tryptophan-hydroxylase-immunoreactive cells in the brain and suboesophageal ganglion of Drosophila melanogaster. Cell Tissue Res. 2010, 340, 51–59. [Google Scholar] [CrossRef]
- Colomb, J.; Reiter, L.; Blaszkiewicz, J.; Wessnitzer, J.; Brembs, B. Open Source Tracking and Analysis of Adult Drosophila Locomotion in Buridan’s Paradigm with and without Visual Targets. PLoS ONE 2012, 7, e42247. [Google Scholar] [CrossRef]
- Alekseyenko, O.V.; Lee, C.; Kravitz, E.A. Targeted Manipulation of Serotonergic Neurotransmission Affects the Escalation of Aggression in Adult Male Drosophila melanogaster. PLoS ONE 2010, 5, e10806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitworth, A.J. Drosophila Models of Parkinson’s Disease. Adv. Genet. 2011, 73, 1–50. [Google Scholar] [CrossRef]
- Pooryasin, A.; Fiala, A. Identified Serotonin-Releasing Neurons Induce Behavioral Quiescence and Suppress Mating in Drosophila. J. Neurosci. 2015, 35, 12792–12812. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Guttman, M.; Boileau, I.; Warsh, J.; Saint-Cyr, J.A.; Ginovart, N.; McCluskey, T.; Houle, S.; Wilson, A.; Mundo, E.; Rusjan, P.; et al. Brain serotonin transporter binding in non-depressed patients with Parkinson’s disease. Eur. J. Neurol. 2007, 14, 523–528. [Google Scholar] [CrossRef]
- Qiu, B.Y.; Qiao, J.X.; Yong, J. Meta-analysis of selective serotonin reuptake inhibitors (SSRIs) compared to tricyclic antidepressants (TCAs) in the efficacy and safety of anti-depression therapy in Parkinson’s disease(PD) patients. Iran. J. Pharm. Res. 2014, 13, 1213–1219. [Google Scholar] [CrossRef] [PubMed]
- Sandoval-Rincón, M.; Sáenz-Farret, M.; Miguel-Puga, A.; Micheli, F.; Arias-Carrión, O. Rational pharmacological approaches for cognitive dysfunction and depression in Parkinson’s disease. Front. Neurol. 2015, 6, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armstrong, M.J.; Okun, M.S. Diagnosis and Treatment of Parkinson Disease: A Review. JAMA 2020, 323, 548–560. [Google Scholar] [CrossRef] [PubMed]
- Bu, M.; Farrer, M.J.; Khoshbouei, H. Dynamic control of the dopamine transporter in neurotransmission and homeostasis. Npj Park. Dis. 2021, 7, 22. [Google Scholar] [CrossRef] [PubMed]
- Boulet, S.; Mounayar, S.; Poupard, A.; Bertrand, A.; Jan, C.; Pessiglione, M.; Hirsch, E.C.; Feuerstein, C.; François, C.; Féger, J.; et al. Behavioral Recovery in MPTP-Treated Monkeys: Neurochemical Mechanisms Studied by Intrastriatal Microdialysis. J. Neurosci. 2008, 28, 9575–9584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, Y.C.; Kim, S.R.; Park, J.-Y.; Chung, E.S.; Park, K.W.; Won, S.Y.; Bok, E.; Jin, M.; Park, E.S.; Yoon, S.-H.; et al. Fluoxetine prevents MPTP-induced loss of dopaminergic neurons by inhibiting microglial activation. Neuropharmacology 2011, 60, 963–974. [Google Scholar] [CrossRef]
- Scheunemann, L.; Plaçais, P.Y.; Dromard, Y.; Schwärzel, M.; Preat, T. Dunce Phosphodiesterase Acts as a Checkpoint for Drosophila Long-Term Memory in a Pair of Serotonergic Neurons. Neuron 2018, 98, 350–365.e5. [Google Scholar] [CrossRef] [Green Version]
- Fanibunda, S.E.; Deb, S.; Maniyadath, B.; Tiwari, P.; Ghai, U.; Gupta, S.; Figueiredo, D.; Weisstaub, N.; Gingrich, J.A.; Vaidya, A.D.B.; et al. Serotonin regulates mitochondrial biogenesis and function in rodent cortical neurons via the 5-HT 2A receptor and SIRT1–PGC-1α axis. Proc. Natl. Acad. Sci. USA 2019, 116, 11028–11037. [Google Scholar] [CrossRef] [Green Version]
- Lipton, S.A.; Kater, S.B. Neurotransmitter regulation of neuronal outgrowth, plasticity and survival. Trends Neurosci. 1989, 12, 265–270. [Google Scholar] [CrossRef]
- Carvajal-Oliveros, A.; Campusano, J.M. Studying the Contribution of Serotonin to Neurodevelopmental Disorders. Can This Fly? Front. Behav. Neurosci. 2021, 14, 262. [Google Scholar] [CrossRef]
- Pratelli, M.; Migliarini, S.; Pelosi, B.; Napolitano, F.; Usiello, A.; Pasqualetti, M. Perturbation of Serotonin Homeostasis during Adulthood Affects Serotonergic Neuronal Circuitry. Eneuro 2017, 4, ENEURO.0376-16.2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simpson, K.L.; Weaver, K.J.; De Villers-Sidani, E.; Lu, J.Y.F.; Cai, Z.; Pang, Y.; Rodriguez-Porcel, F.; Paul, I.A.; Merzenich, M.; Lin, R.C.S. Perinatal antidepressant exposure alters cortical network function in rodents. Proc. Natl. Acad. Sci. USA 2011, 108, 18465–18470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugie, A.; Marchetti, G.; Tavosanis, G. Structural aspects of plasticity in the nervous system of I. Neural Dev. 2018, 13, 14. [Google Scholar] [CrossRef] [PubMed]
- Devaud, J.-M.; Acebes, A.; Ramaswami, M.; Ferrús, A. Structural and functional changes in the olfactory pathway of adult Drosophila take place at a critical age. J. Neurobiol. 2003, 56, 13–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sachse, S.; Rueckert, E.; Keller, A.; Okada, R.; Tanaka, N.K.; Ito, K.; Vosshall, L.B.B. Activity-Dependent Plasticity in an Olfactory Circuit. Neuron 2007, 56, 838–850. [Google Scholar] [CrossRef] [Green Version]
- Golovin, R.M.; Vest, J.; Vita, D.J.; Broadie, K. Activity-dependent remodeling of Drosophila olfactory sensory neuron brain innervation during an early-life critical period. J. Neurosci. 2019, 39, 2995–3012. [Google Scholar] [CrossRef] [Green Version]
- Poewe, W.; Seppi, K.; Tanner, C.M.; Halliday, G.M.; Brundin, P.; Volkmann, J.; Schrag, A.E.; Lang, A.E. Parkinson disease. Nat. Rev. Dis. Prim. 2017, 3, 17013. [Google Scholar] [CrossRef]
- Nielsen, S.S.; Warden, M.N.; Camacho-Soto, A.; Willis, A.W.; Wright, B.A.; Racette, B.A. A predictive model to identify Parkinson disease from administrative claims data. Neurology 2017, 89, 1448–1456. [Google Scholar] [CrossRef]
- Heinzel, S.; Berg, D.; Gasser, T.; Chen, H.; Yao, C.; Postuma, R.B. Update of the MDS research criteria for prodromal Parkinson’s disease. Mov. Disord. 2019, 34, 1464–1470. [Google Scholar] [CrossRef]
- Faust, I.M.; Racette, B.A.; Nielsen, S.S. Validation of a Parkinson Disease Predictive Model in a Population-Based Study. Parkinsons Dis. 2020, 2020, 2857608. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zárate, R.V.; Hidalgo, S.; Navarro, N.; Molina-Mateo, D.; Arancibia, D.; Rojo-Cortés, F.; Oliva, C.; Andrés, M.E.; Zamorano, P.; Campusano, J.M. An Early Disturbance in Serotonergic Neurotransmission Contributes to the Onset of Parkinsonian Phenotypes in Drosophila melanogaster. Cells 2022, 11, 1544. https://doi.org/10.3390/cells11091544
Zárate RV, Hidalgo S, Navarro N, Molina-Mateo D, Arancibia D, Rojo-Cortés F, Oliva C, Andrés ME, Zamorano P, Campusano JM. An Early Disturbance in Serotonergic Neurotransmission Contributes to the Onset of Parkinsonian Phenotypes in Drosophila melanogaster. Cells. 2022; 11(9):1544. https://doi.org/10.3390/cells11091544
Chicago/Turabian StyleZárate, Rafaella V., Sergio Hidalgo, Nicole Navarro, Daniela Molina-Mateo, Duxan Arancibia, Francisca Rojo-Cortés, Carlos Oliva, María Estela Andrés, Pedro Zamorano, and Jorge M. Campusano. 2022. "An Early Disturbance in Serotonergic Neurotransmission Contributes to the Onset of Parkinsonian Phenotypes in Drosophila melanogaster" Cells 11, no. 9: 1544. https://doi.org/10.3390/cells11091544