
Vitamin D Receptor Influences Intestinal Barriers in Health and Disease


1
Department of Medicine, University of Illinois at Chicago, Chicago, IL 60612, USA
2
Department of Microbiology/Immunology, University of Illinois at Chicago, Chicago, IL 60612, USA
3
Jesse Brown VA Medical Center Chicago (537), 820 S Damen Ave, Chicago, IL 60612, USA
*
Author to whom correspondence should be addressed.
Cells 2022, 11(7), 1129; https://doi.org/10.3390/cells11071129
Submission received: 20 February 2022
/
Revised: 23 March 2022
/
Accepted: 24 March 2022
/
Published: 27 March 2022
(This article belongs to the Special Issue Cell-Cell Interactions and Cell Adhesion Signaling in Disease States)
Abstract
:Vitamin D receptor (VDR) executes most of the biological functions of vitamin D. Beyond this, VDR is a transcriptional factor regulating the expression levels of many target genes, such as genes for tight junction proteins claudin-2, -5, -12, and -15. In this review, we discuss the progress of research on VDR that influences intestinal barriers in health and disease. We searched PubMed and Google Scholar using key words vitamin D, VDR, tight junctions, cancer, inflammation, and infection. We summarize the literature and progress reports on VDR regulation of tight junction distribution, cellular functions, and mechanisms (directly or indirectly). We review the impacts of VDR on barriers in various diseases, e.g., colon cancer, infection, inflammatory bowel disease, and chronic inflammatory lung diseases. We also discuss the limits of current studies and future directions. Deeper understanding of the mechanisms by which the VDR signaling regulates intestinal barrier functions allow us to develop efficient and effective therapeutic strategies based on levels of tight junction proteins and vitamin D/VDR statuses for human diseases.
1. Introduction
The intestinal epithelium cells (IECs) are essential in barrier function, structural function, and host defense. The tight junctions (TJs) seal the space between adjacent epithelial cells. TJs are made by a number of proteins, including claudins, occludin, junctional adhesion molecules (JAM), and tricellulin and cytoplasmic plaque proteins, e.g., zonula occludens (ZO) proteins. Interestingly, VDR is known to transcriptionally regulate several genes for TJs, including claudin-2, -5, -12, and -15 in the intestinal epithelial cells [1,2,3,4]. VDR deletion in IEC cells leads to decreased protein levels of claudins, such as claudin-2 and -12, which contribute to vitamin D-dependent homeostasis. However, VDR regulation of some TJ proteins may be vitamin D-independent.
In 2013, we reviewed the roles of vitamin D and VDR in tissue barriers [5]. Back then, our understanding of VDR in influencing intestinal barriers was limited to its role as a transcriptional factor and a receptor for vitamin D. We did not have sufficient evidence on its tissue-, gender-, and time-specificity in regulating barrier function. We did not sufficiently consider the physiological relevance in vivo and in disease models. We did not know that the human gut microbiome is determined by the variation of vdr gene [6]. Indeed, ~3% of the mouse and human genomes are regulated directly or indirectly by vitamin D, suggesting widespread effects of vitamin D/VDR in various disease mechanisms [7,8,9]. Thus, the traditional model of treating cultured cells with vitamin D3 is now giving way to models with a more complex mechanism of action.
In the current review, we discuss the progress of research on VDR that influences intestinal barriers in health and disease. We searched PubMed and Google Scholar using key words “vitamin D, VDR, tight junctions, cancer, inflammation, and infection”. We summarize studies related to VDR/TJ distribution, cellular functions, and mechanisms; we discuss the role of VDR and barriers in various diseases, e.g., IBD, colon cancer, chronic obstructive pulmonary disease (COPD), infection, and other diseases, mainly focusing on the intestinal diseases. We also discuss the limits of current studies and future directions. New insights into VDR and TJs will allow us to better understand the pathophysiology of diseases and develop novel strategies for prevention and treatment.
Intestinal TJs for Barrier Functions
The intestinal barriers are provided by several layers: mucus, epithelial cells and TJs, and the local immune system. The intestinal mucus layer has a primary role in host protection against mechanical, chemical, and biological injuries. The intestinal epithelium is the second layer, which serve as a barrier, provide structure, and play a role in host defense [10,11,12,13,14,15,16]. TJs seal the space between adjacent epithelial cells and regulate epithelial polarity and vectorial movement of solutes and fluids in the intercellular space. The microbiome contributes to maintaining the integrity of the intestinal epithelium. Bacteria can induce proliferation of IECs, which is evidenced by lower proliferative IECs in germ-free (GF) than those in conventional mice [17]. A recent study [18] showed evidence on microbiota and their product butyrate in regulating expression of actin-binding protein synaptopodin for intestinal homeostasis. Synaptopodin is localized to the TJs and within F-actin stress fibers of intestinal epithelial cells. It is critical for barrier integrity and cell motility. Butyrate regulation of synaptopodin reveals a direct mechanistic link between microbiota-derived metabolite and intestinal barrier functions.
TJ structural components, which determine epithelial polarization and intestinal barrier function, can be regulated by homeodomain proteins that control the differentiation of the intestinal epithelium [19,20]. TJ proteins include claudins, occludin, JAM, and ZO. Many TJ proteins, such as ZO and occludin, tighten cell structure and maintain a barrier [21]. Some TJ proteins, such as claudin-2, are considered as “leaky” proteins because they form paracellular water channels [22] and increased their expression in inflammation and cancer [23,24,25,26,27,28]. Mikio Furuse was the first to purify and identify claudins in the laboratory of the late Shoichiro Tsukita [29]. The name is taken from the Latin word “claudere,” meaning to close, because it was anticipated that these proteins might constitute the tight junctional barrier [30]. Occludin, tricellulin, and JAMs genes exist as a limited number of different isoforms; they are unlikely to account for the diversity of paracellular permeability characteristics in different epithelia [30]. There are 27 claudin family members that contribute to tight junctions [31], and not all claudins are the same. Thus, many studies are on claudins to determine paracellular selectivity. Some claudins have other roles in cell signaling and differentiation. For example, a recent study reported that claudin-7 controls intestinal crypt stem cell survival, self-renewal, and epithelial differentiation through Wnt/β-catenin signaling in mice [32]. claudin-2 and claudin-12 form paracellular Ca2+ channels in human intestinal epithelial cell lines [1].
In a healthy adult intestine, the expression and apical tight junction localization of zonula occludens 1 (ZO-1) and occludin are unchanging. In contrast, most claudins are expressed in decreasing or increasing gradients or in more complex patterns along the longitudinal axis of the intestine and the crypt to villus/surface differentiation axis, based on the observations in mice [33]. During neonatal development, several claudins altered in transcript expression as well as changes in localization along the crypt–villus axis. claudin-19 was only detected at days 1 and 14 in mice [33]. In Figure 1, claudin-2 restricts to the crypt at the basal lateral side of the mouse intestine, whereas claudin-7 localizes in the epithelial cell membrane along the crypt–villus axis, based on our previous reports [2,27]. From the website Protein Atlas, we can find some claudins related to their distribution, expression levels in different human organs, disease-related information, and validated antibodies for research (https://www.proteinatlas.org/ENSG00000184113-CLDN5, accessed on 2 February 2022).
2. Vitamin D/VDR and TJs in Intestinal Homeostasis
Classically, 1,25-dihydroxy vitamin D (1,25(OH)2D3), the active form of vitamin D, plays a key role in calcium homeostasis and in electrolyte and blood pressure regulation [34]. VDR mediates most known physiological functions of 1,25(OH)2D3. Once VDR binds with 1,25(OH)2D3, VDR heterodimerizes with the retinoid X receptor (RXR) in nuclei, then binds to the vitamin D-response element (VDRE) in the promoter of the target gene, thus activating gene transcription [35]. VDR target genes include genes for the antimicrobial peptide (AMP) cathelicidin precursor (LL-37) [36], β defensin [37], autophagy regulator autophagy related 16 Like 1 (ATG16L1) [38], and proliferation regulator Axin1 [39], in addition to the genes for claudin-2, -5, -12, and -15 [1,2,3,4].
VDR is critical in regulating intestinal homeostasis by preventing pathogenic bacterial invasion, inhibiting inflammation, and maintaining barrier functions [36,40,41,42,43,44]. In the Protein Atlas (https://www.proteinatlas.org/ENSG00000111424-VDR, uploaded on June 2018; accessed on 23 February 2022), we can find that the presence of VDR in various human and mouse tissues, along with its ability to exert differentiation [45,46], growth, and inhibitory [46] and anti-inflammatory actions [43,47,48,49], sets the stage for therapeutic exploitation of VDR ligands for the treatment of various inflammatory conditions. VDR deletion in human IEC cells lead to decreased claudin-2 and -12 [1]. However, little is known about how the intestinal VDR are involved in TJs and intestinal inflammation and infection, especially in in vivo systems.
In Table 1, we summarize the studies on vitamin D/VDR directly or indirectly influence the expression and function of cell junction proteins in different tissues from human and mouse samples. Although the primary focus of this review is on the intestinal epithelial TJ barrier, the studies in other cell types, including lung, skin, kidney, cornea, brain, oral, and urinary bladder, are also included in the current table.
3. Novel Roles of Vitamin D, VDR, and TJs in Diseases
Emerging evidence demonstrates that vitamin D/VDR deficiency is a critical factor in the pathology of many diseases, such as IBD and colon cancer, among others [45,74,75,76,77,78,79,80,81]. In the following section, we discuss the tissue-specific regulatory functions of VDR on TJ proteins in different diseases, based on clinical data and laboratory evidence.
3.1. Vitamin D, VDR, and TJs in Intestinal Inflammation
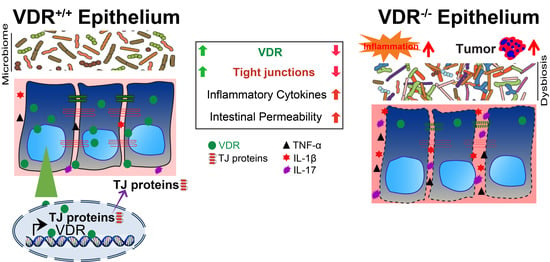
Multiple factors (e.g., risk genes, environmental triggers, immunity, and microbiome) contribute to the pathogenesis of IBD [16,82,83,84,85]. Susceptibility to IBD is associated with polymorphisms in the vdr gene [86,87,88,89,90]. Low vitamin D status and reduced VDR is observed in patients with IBD [91,92,93,94]. Vitamin D influences the course and severity of human IBD [92]. Vitamin D/VDR appears to be an important immunological regulator of IBD [94,95]. In experimental models, VDR knockout (KO) mice spontaneously developed colitis [96,97,98,99,100,101] and VDR/IL-10 double KO mice developed severe IBD [102]. Our studies further demonstrated that mice with conditional deletion of intestinal epithelial VDR are susceptible to colitis, due to dysbiosis, dysfunction of Paneth cells, dysregulated autophagy, and disrupted TJs [27,38].
Luminal stimuli and epithelial cell dysfunction are known to contribute to IBD pathogenesis and progression [103,104]. Defective epithelial barrier function has been implicated in IBD and can predict relapse during clinical remission. Increased permeability is also present in a subset of unaffected first-degree relatives of patients with Crohn’s disease [105], a type of IBD [104]. Claudin-2 forms a paracellular water channel and thus mediates paracellular water transport in leaky epithelia [22,33,106,107,108]. Changes in the expression and distribution of claudin-2, -5, and -8 lead to discontinuous TJs and barrier dysfunction in active CD [26,106]. Loss-of-function mutations in protein tyrosine phosphatase nonreceptor type 2 [109] increase the risk of IBD and celiac disease. Interestingly, T-cell protein tyrosine phosphatase (TCPTP) protects against intestinal barrier dysfunction induced by the inflammatory cytokine IFN-gamma. The mechanisms are that it maintained localization of ZO-1 and occludin at apical tight junctions and restricted both expression and insertion of the cation pore-forming transmembrane protein, claudin-2, at TJs through upregulation of matriptase, the inhibitory cysteine protease, in experimental mouse models [109].
Intestinal epithelial VDR deletion led to decreased claudin-2 at both mRNA and protein levels in mice [2]. Previous studies used the whole VDR−/− mouse model, which is known to have problems in various organs. To determine the role of intestinal epithelial VDR and mechanisms by which VDR regulates claudin-2, intestinal epithelial-specific VDR knockout VDR∆IEC mice were used. It was demonstrated that claudin-2 is transcriptionally regulated by VDR in the healthy intestine; however, claudin-2 is hyper regulated through inflammatory signaling in colitis with reduced intestinal epithelial VDR [2,27].
Claudin-15 is significantly lower in patients with IBD, compared to the health controls [3,110]. VDR conditional knockout VDR∆IEC or overexpressing mouse models, cultured human cell lines, and organoids were used to investigate the mechanisms of altered claudin-15 in intestinal inflammation [3]. Overexpression of intestinal epithelial VDR resulted in significantly increased claudin-15 and decreased susceptibility to chemically and bacterially induced colitis. Colonic claudin-15 was reduced in VDR∆IEC mice, which were susceptible to colitis. Direct binding of VDR to the claudin-15 promoter was identified. Abnormal gut barrier function may serve as a biomarker for the risk of IBD onset [111]. This study provides an explanation of reduced claudin-15 and VDR observed in the human IBD and deeper understanding of mechanisms by which tissue-specific VDR regulating barrier functions in the intestine.
3.2. Disrupted Intestinal Barrier in Colorectal Cancer (CRC)
The disruption of TJs is a common manifestation of CRC. Multiple factors, including host genetic background, immunity, environment, and microbiome contribute to the development of colon cancer [81,112,113]. At baseline, obesity, history of a sessile-serrated adenoma, and a family history of CRC were associated with claudin-1, occludin, and mucin-12 (MUC12) [114]. Mandle et al. investigated the effects of supplemental calcium (1200 mg, daily) and/or vitamin D3 (1000 IU daily) on intestinal barrier function-related biomarkers in 105 participants from a large colorectal adenoma recurrence chemoprevention clinical trial. They tested expression of claudin-1, occludin, and MUC12 in the normal-appearing colorectal mucosa. Following 1 year of treatment, in the calcium relative to the no-calcium group, the CLDN1, OCLD, and MUC12 expression increased. The estimated calcium treatment effects were greater among participants with baseline serum 25-OH-vitamin D concentrations below the median value of 22.69 ng/mL (claudin-1: 29%, p = 0.04; OCLD: 36%, p = 0.06; MUC12: 35%, p = 0.05), suggesting the link between vitamin D deficiency and TJ proteins in the human CRC. VDR is known to protect against tumorigenesis in the colon [4,115]. However, this study did not test the level of VDR in these patients. There are no data of microbiome from these patients.
We retrieved the expression of 20 claudins in normal and colon cancer patients by using Sequence Read Archive (SRA) microarray data (Gene Expression Omnibus (GEO) database GSE8671). Among these claudins, the expression of claudin-1, -2, -12, and -19 were increased in colon cancer patients; mRNA expression of claudin-5, -8, -15, -17, -20, and -23 were significantly decreased in patients with colon cancer; and the expression of claudin-3, -4, -6, -7, -9, -10, -11, -14, -16, and -18 did not show distinct changes in patients with colon cancer and normal. These data indicate the significance to study claudins in well-controlled experimental models and understand their different roles in the development of cancer.
Claudin-5 forms paracellular barriers and pores that determine permeability in the epithelia and endothelia. It is downregulated in colon cancer [116,117]. We found that reduction of colonic VDR expression is positively correlated with the low level of claudin-5 in patients with CRC [4]. We determined that VDR acts as a transcriptional regulator for the maintenance of physiological levels of the target gene claudin-5 in the colon. Furthermore, in a conditional intestinal epithelial VDR-overexpressed mouse model, we found the protective role of VDR in the maintenance of TJs in the context of inflammation and colitis-associated colon cancer. High VDR level contributed to high claudin-5, reduced intestinal inflammation, and fewer tumors in the mouse colon, similar as the observation in the human CRC [4]. Interestingly, claudin-7 was not altered in the VDR-deficient colon, compared with the VDR+/+ colon in mice [4]. Our findings offer an insight into the VDR regulation on certain claudins, depending on the disease types.
VDR plays multiple functional roles of in the development of colon cancer [115,118]; thus, it is important to dissect the mechanisms by which VDR contributes to barrier function in protecting the host from tumorigenesis. The positively correlated status of VDR and claudins, such as claudin-5 and -15, could be potentially applied to risk assessment, early detection, and prevention of colitis and colitis-associated colon cancer. Hence, new insights into the mechanisms responsible for VDR and barrier dysfunction are needed. These studies will provide a new avenue to restore barrier functions and develop a novel protocol for risk assessment and prevention of CRC and other human cancers.
3.3. TJs and VDR in Lungs
Vitamin D/VDR signaling plays an important role in regulating the components of junctions and maintaining the integrity of epithelial barriers in multiple organs. In lungs, the permeability of the alveolar epithelial barrier is mainly regulated by the intercellular junctions [119,120]. In chronic inflammatory lung diseases, epithelial barrier function is impaired, disruption of the epithelial barrier resulted in alveolar permeability, i.e., paracellular movements of fluid from the interstitium to the pulmonary airspace, and infiltration of inflammatory cells [64]. Figure 2 shows well-organized ZO-1 in epithelial cells of the healthy lungs and lipopolysaccharide (LPS) treatment that disrupted the ZO-1 distribution in a mouse mode in vivo [65].
Vitamin D can promote epithelial barrier integrity or protect against epithelial barrier destruction in lungs. In the bronchial epithelial cell line 16HBE, vitamin D is able to counteract the effects of cigarette smoke extract–induced bronchial epithelial barrier disruption, such as transepithelial electrical resistance (TER) reduction, permeability increase, distribution anomalies, and increased cleavage of E-cadherin and β-catenin [63]. Vitamin D-treated BALB/c mice increased the expression of ZO-1 and E-cadherin, and reduced signs of asthma induced by toluene di-isocyanate [121]. Vitamin D treatment alleviated LPS-induced lung injury and preserved alveolar barrier function by inducing the expression of occludin and ZO-1 to maintain the pulmonary barrier in lungs [64,122], whereas in healthy mice fed with a vitamin D-poor diet indicated that vitamin D supplementation had little effect on epithelial integrity [123]. Vitamin D also promotes epithelial barrier function through its ability to increase expression of cystic fibrosis transmembrane conductance regulator in airway epithelial cells [124]. These studies indicate that vitamin D promotes the integrity and function of the epithelial barrier and protects against epithelial damage by dampening inflammatory response.
Lung VDR plays an important role in maintaining the pulmonary barrier integrity. We reported that VDR deletion increased lung permeability by altering the expression of TJ molecules, particularly claudin-2, -4, -10, -12, and -18 [65]. VDR KO mice showed significantly decreased expression of junctional proteins, including ZO-1, occludin, claudin-1, -2, -4, -12, -18, β-catenin, and VE-cadherin in lungs, compared with WT mice. claudin-2, -4, -12, and -18 mRNA and protein levels were significantly decreased in the lungs of VDR−/− mice. However, VDR deletion did not change the mRNA levels of ZO-1, occludin, claudin-1, -3, -7, a-catenin, β-catenin, and VE-cadherin, suggesting the different mechanisms by which VDR regulates junctional proteins. These changes of TJs are closely correlated with airway epithelial barrier destruction in chronic pneumonia because of vitamin D/VDR deficiency [65]. Interestingly, claudin-15 mRNA level was significantly decreased in lungs of VDR−/− mice, but total claudin-15 protein was unchanged. Immunoblotting of claudin-15 protein in total lung tissue may not reflect its changes in certain types of lung cells [65]. Vitamin D supplementation alleviated LPS-induced lung injury and preserved alveolar barrier function through maintenance of the pulmonary barrier by inducing expression of occludin and ZO-1 in whole lung homogenates [64].
Several studies revealed the link of vitamin D deficiency to chronic inflammatory lung diseases, e.g., chronic obstructive pulmonary disease (COPD) and asthma [125,126,127,128]. COPD is a complex and progressive lung disease that is characterized by persistent airflow limitation resulting from chronic inflammation and structural changes [129]. The lower vitamin D levels and the more severe airflow obstruction were observed in patients with COPD [130] and accelerated decline in lung function [131]. VDR-deficient or knockout mice develop a COPD phenotype, whereas VDR overexpression can ameliorate inflammation in the lungs [132,133]. Vitamin D treatment reduces exacerbation rates in vitamin D-deficient patients with COPD or asthma, thus decreasing the incidence of acute respiratory tract infections [134,135,136].
3.4. Infection
The major obstacle pathogens must overcome is the intestinal epithelial barrier [137]. TJs are critical to providing host defense against pathogens. One of the beneficial roles vitamin D/VDR is to act as an important mediator of intestinal epithelial defenses against infectious agents. Vitamin D deficiency predisposes to more severe intestinal injury in an infectious model of colitis [138]. Vitamin D-deficient mice challenged with C. rodentium demonstrated increased colonic hyperplasia and epithelial barrier dysfunction. Vitamin D deficiency resulted in an altered composition of the fecal microbiome with or without C. rodentium infection [138], and 1,25(OH)2D3 altered E. coli O157:H7-induced reductions in TER, decreased permeability, and preserved barrier integrity in mice [139].
Salmonella are the causative agents of a variety of diseases ranging from diarrhea-generating gastroenteritis to systemic typhoid fever. Salmonella induces the disruption of TJs during infection. This is exemplified through decreases in TER, increases in tracer permeability, and TJ protein alterations in the infected intestinal cell lines and mouse tissues [137,140,141]. We and others have demonstrated that Salmonella targets TJ proteins [137,140,141], e.g., ZO-1 (Figure 3) [142,143,144], occludin [142,143,144], and claudins [28,105], and facilitates pathogenic enteric bacterial invasion [28]. The expression of claudin-7 is very stable in the colon in mouse colitis or bacterial infection [2,145].
Pathological bacterial translocation from the disrupted intestinal barrier leads to substantial complications and mortality in other organs, such as liver cirrhosis. Lee et al. [53] investigated the effects of calcitriol on bacterial translocation in cirrhotic rats. Cirrhotic rats were administrated with a 2-week course of active vitamin D3 (calcitriol, 0.1 µg/kg per day) or vehicle by oral gavage after thioacetamide (TAA) injection for 16 weeks. Vitamin D3 treatment significantly attenuated bacterial translocation and reduced intestinal permeability in TAA-induced cirrhotic rats. It upregulates the expressions of occludin in the small intestine and claudin-1 in the colon of cirrhotic rats directly independent of intrahepatic status. Vitamin D3 treatment also enriched Muribaculaceae, Bacteroidales, Allobaculum, Anaerovorax, and Ruminococcaceae in mice. This study showed that vitamin D3 attenuates intestinal leakage, reduces bacterial translocation, and enriches potentially beneficial gut microbiota, suggesting a potential therapeutic agent to prevent cirrhotic complications [53].
Vitamin D upregulates TJ proteins occludin and claudin-14 during E. coli urinary tract infection [73], likely improving the epithelial integrity, which in turn may ameliorate the protection against infection. This finding is relevant especially among patients with recurrent urinary tract infection (UTI) and where low vitamin D levels are anticipated. Bladder biopsies were obtained from post-menopausal women before and after a 3-month period of supplementation with 25-hydroxyvitamin D3 and ex vivo infected with E. coli. In biopsies, obtained before E. coli infection, vitamin D had no effect on TJ proteins. However, during E. coli infection, vitamin D induced occludin and claudin-14 in mature superficial umbrella cells of the urinary bladder. Vitamin D increased cell–cell adhesion, thus consolidating the epithelial integrity during infection [73].
We have started to collect more evidence of the tissue-specific role of VDR in intestinal health and inflammation. The studies in UTI [73] support the gender-difference and time-dependent (e.g., post-menopausal women) roles of VDR on TJs. We also noted gender differences in microbial metabolites [146] and virome in the mouse models [147]. We still do not have sufficient evidence for the gender- and time- specificity in the VDR-regulating barrier function in human samples.
The prevention of gut-barrier dysfunction is a viable approach for anti-infection, including COVID-19 [148]. Vitamin D deficiency is associated with adverse outcomes in infections [149]. There are compelling epidemiological associations between incidence and severity of COVID-19 and vitamin D deficiency [150]; one of the mechanisms is that the vitamin D pathway accelerates shutdown of TH1 cells in severe COVID-19. However, the mechanisms through VDR and tight junctions remain unknown. Even with the clear evidence that TJ function is influenced by pathogens, much more work is needed to understand the microbial regulation of intestinal TJs and to develop novel therapeutic targets for alleviating infection and infection-associated inflammation.
4. Elucidate Cellular and Molecular Mechanisms of Intestinal VDR in Regulating TJ Proteins
4.1. VDR Transcriptional Regulation of the Genes of TJ Proteins
Based on the genetic studies, VDR is a transcriptional factor regulating the genes of some TJ proteins, including claudin-2, -5, -12, and -15 in the intestinal epithelial cells [1,2,3,4]. For example, the sequence of functional vitamin D response element (AGATAACAAAGGTCA) is identified in the promoter of claudin-5 gene. VDR deletion reduced the mRNA and protein levels of claudin-5 and upregulation of VDR increased the expression of claudin-5. These data demonstrate the genetic regulation of VDR on certain genes for TJs.
Another important question concerning VDR regulation is whether it is vitamin D dependent or independent. Although in normal cells with sufficient VDR expression, we observe the vitamin D-induced TJ expression through the VDR, the inflamed cells may use a totally different mechanism to alter TJs because the TJ proteins are also regulated by cytokines and other signaling pathways. In inflamed intestine of ulcerative colitis patients, VDR expression was low and claudin-2 was enhanced. Mechanistically, the enhanced claudin-2 promoter activity through the binding sites of nuclear factor kappa B (NF-κB) and signal transducer and activator of transcription (STAT) in inflamed cells without VDR [27]. Cytokines play an important role in the modulation of the intestinal epithelial TJ barrier. The pro-inflammatory cytokine, such as interleukin-6 (IL-6), interferon gamma (IFN-γ) and tumor necrosis factor alpha (TNF-α), induced increase in intestinal TJ permeability, which is an important pathogenic mechanism contributing to the development of intestinal inflammation [151]. IL-6 upregulates claudin-2 expression in intestinal epithelium [152]. On one hand, VDR is known to physically interact with NF-κBp65 and transcriptionally regulates IκBa to execute the anti-inflammatory role and suppress the inflammatory cytokines [153,154]. On the other hand, there is limited study on whether and how high inflammatory cytokines might directly suppress the expression of VDR.
4.2. Cellular Changes of TJs by 1,25(OH)2D3/VDR Status
TJ proteins, such as ZO-1, are upregulated in enterocytes by 1,25(OH)2D3 in vitro [1,43]. Severely disrupted TJs and increased permeability were seen in DSS-treated VDR−/− colonic epithelial cells [43]. However, in the VDR−/− mice, VDR deletion did not change the mRNA levels of ZO1, occludin, claudin-1, -3, -7, α-catenin, β-catenin, and VE-cadherin, suggesting the different mechanisms by which VDR regulates junctional proteins. We found that claudin-15 mRNA levels were significantly decreased in lungs of VDR−/− mice, but total claudin-15 protein levels were not changed in the total lung tissue by Western blot. However, in the intestinal tissue, VDR deletion led to reduction of claudin-15 [3]. These results suggested that claudins have a different biological function in the different cells or tissues. Hence, further insights into the mechanisms responsible for intestinal VDR and barrier dysfunction are needed, especially in in vivo systems and human samples focusing on the tissue specificity and gender difference of VDR.
4.3. Other Regulators, e.g., Probiotics and Microtome, on TJs and VDR
Other factors, such as environmental factor and microbiome, also contribute to the changes of VDR and TJs. Most commensal organisms are kept segregated from the epithelium by the mucosa. Goblet cells are responsible for mucus production. Mucus consists of secretory immunoglobulin type A (sIgA) and AMPs (e.g., α-defensins), which are secreted by Paneth cells located in the small intestinal crypts and released to the colon. IgA regulates the composition and metabolic function of gut microbiota by promoting symbiosis between bacteria [155]. Macrophages in the underlying intestinal tissue produce milk fat globule-EGF factor 8, which directly targets IECs and regulates the integrity of intestinal barrier function [156]. The presence of these components helps to maintain the balance in the composition of the gut microbiota. Dysfunction in the production/secretion of individual proteins can lead to not only disturbances in the proportions of individual bacteria, but also the functions of barriers.
Whereas vitamin D has been intensively studied, the role of bacteria in modulating the effects of vitamin D and VDR signaling is not well known. We integrate our findings with other studies and, more importantly, understand how probiotics coordinate the effects of vitamin D/VDR and maintain the barrier function [157,158,159,160,161,162]. The novel information on the combined role of vitamin D and probiotics in anti-inflammation will contribute new concepts to the therapeutic methods for IBD and other inflammatory diseases. Although the potential importance of VDR as a therapeutic target has been recognized [163], no approach to date has safely and effectively altered its activity. Overuse of vitamin D to activate VDR signaling has potent hypercalcemic effect. In contrast, probiotic usage has little downside. Although probiotics can be dangerous if administered to very sick patients, the worst aspect is that probiotics will not produce any obvious benefit. By taking probiotics and/or vitamin D, it may indicate an inexpensive method to prevent chronic diseases.
5. Limits and Future Directions
We have made significant progress in the past decade to understand the role that vitamin D/VDR has in regulating tissue barriers. A series of molecular and biochemical experiments in vivo and in vitro, using VDR−/− mice, conditional knockout models, organoids, and human database, allow us to investigate the in-depth mechanisms by which barriers are modulated in health and disease. However, the traditional model is to treat cultured cells or animals with vitamin D3. Our understanding of VDR in influencing intestinal barriers was limited to its role as a transcriptional factor and a receptor for vitamin D.
We now know the diverse roles and tissue specificity of VDR and its ligands [164,165]. We have begun to appreciate their clinical application in various human diseases [165]. For example, VDR ligands (e.g., calcipotriol) suppress pancreatitis and VDR acts as a master transcriptional regulator in pancreatic stellate cells [166]. VDR expressed in stroma from human pancreatic tumors and calcipotriol could reduce markers of inflammation and fibrosis in pancreatitis and human tumor stroma, suggesting vitamin D priming as an adjunct in the treatment of pancreatic ductal adenocarcinoma. In the kidney, 1,25(OH)2 VitD influences claudin-16-mediated Mg2+ transport [150]. However, in lungs, claudin-16 is very low and may not be necessary for respiratory function, or, other claudins may complement the function of claudin-16 in the lungs [167]. Thus, we need more studies on VDR’s tissue-, gender-, and time-specificity in regulating barrier function in health and disease. We need to consider sufficiently the physiological relevance in vivo and in disease models. We need to consider the other factors, e.g., microbiome and inflammatory triggers, which could take over in the vitamin D deficient context to alter TJ proteins, thus manipulating the barrier functions. We need to recognize that PCR data on mRNA alteration could not reflect the cellular distribution and functions of TJs. Functional studies and the syngenetic roles of TJ proteins are needed.
A series of molecular and biochemical experiments were performed in vivo and in vitro by using VDR transgenic mice and cultured human intestinal epithelial cells. However, the traditional model is to treat cultured cells or animals with vitamin D3. Our understanding of VDR in influencing intestinal barriers is limited to its role as a transcriptional factor and a receptor for vitamin D. In many clinical studies, the status of VDR was not considered in the study design or data interpretation. To move forward, special attention is needed on the roles of VDR in a gender-different and time-dependent manner, e.g., post-menopausal women, aging, and different stages of diseases.
Many researchers already notice the limits of cell lines. The recent development of organoids will help us to study tight junctions, physiological relevance, host–microbial interactions, and drug discovery [145,161,168,169,170,171,172]. As shown in Figure 4, claudin-7 is well maintained and distributes in the cell membranes in the mouse organoids. We found that TER of mouse colonoid-derived monolayers remained unchanged between the O-VDR overexerting colonoids and VDRloxP colonoids [3]. We still need more studies on its tissue, gender, and time specificity in regulating barrier function. We need to consider sufficiently the physiological relevance of in vivo and in disease models, and the advancement of technologies and methods for the overall investigation of the intestinal barrier [173].
6. Conclusions
The recent progress reveals a novel activity of VDR in regulation of many tight junction proteins in primate cell structure and intestinal homeostasis and diseases (as shown in the Graphic Abstract). We aim to show the current state of knowledge on this topic and its potential therapeutic applications. This knowledge can be used to develop intestinal VDR-associated TJ proteins, e.g., claudin-5 and -15, as clinical biomarkers for identifying patients who may benefit from currently available interventions and could be used for the eventual development of novel strategies for the prevention and treatment of diseases. VDR signaling is also highly significant in regulating other proliferation and anti-inflammatory pathways [74,157,162,174]. We hope to integrate our findings with other studies and, more importantly, understand how the microbiome, probiotics, and metabolites coordinate the effects of vitamin D/VDR [146]. Our long-term goal is to develop individualized therapeutic strategies based on tight junction proteins [175] and vitamin D/VDR statuses for efficient and effective prevention and treatment of chronic diseases.
Author Contributions
Y.-G.Z. and J.S. performed the literature search, detailed analyses, summarized related literature, and prepared the draft text. J.S. designed the study/project, obtained funds, summarized the literature, and directed the project. Both authors contributed to writing the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
We would like to acknowledge the VA Merit Award 1 I01BX004824-01, DOD CDMRP BC191198, the NIDDK/National Institutes of Health grants R01 DK105118 and R01DK114126 to Jun Sun. The study sponsors performed no role in the study design, data collection, analysis, and interpretation of data. The contents do not represent the views of the United States Department of Veterans Affairs or the United States Government.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| 1,25(OH)2D3 | 1,25-dihydroxy vitamin D |
| AMP | Antimicrobial Peptide |
| ATG16L1 | Autophagy Related 16 Like 1 |
| CD | Crohn’s Disease |
| COPD | Chronic Obstructive Pulmonary Disease |
| CRC | Colorectal Cancer |
| DSS | Dextran Sulfate Sodium |
| GEO | Gene Expression Omnibus |
| GI | Gastrointestinal |
| GF | Germ-Free |
| H&E | Hematoxylin and Eosin |
| IBD | Inflammatory Bowel Disease |
| IEC | Intestinal Epithelial Cells |
| IF | Immunofluorescence |
| IHC | Immunohistochemistry |
| IL-6 | Interleukin-6 |
| IFN-γ | Interferon Gamma |
| JAM | Junctional Adhesion Molecules |
| KO | Knockout |
| LPS | Lipopolysaccharide |
| LoxP | Locus of X-over, P1 |
| MMP7 | Matrix Metalloproteinase 7 |
| MUC2 | Mucin 2 |
| NF-κB | Nuclear Factor Kappa B |
| PC | Paneth Cell |
| qRT-PCR | Quantitative Real Time Polymerase Chain Reaction |
| RXR | Retinoid X Receptor |
| TAA | Thioacetamide |
| TER | Transepithelial Electrical Resistance |
| SRA | Sequence Read Archive |
| STAT | Signal Transducer and Activator of Transcription |
| sIgA | Secretory Immunoglobulin Type A |
| TJ | Tight Junction |
| TNF-α | Tumor Necrosis Factor |
| VDR | Vitamin D Receptor |
| UC | Ulcerative Colitis |
| UTI | Urinary Tract Infection |
| ZO | Zonula Occludens |
References
- Fujita, H.; Sugimoto, K.; Inatomi, S.; Maeda, T.; Osanai, M.; Uchiyama, Y.; Yamamoto, Y.; Wada, T.; Kojima, T.; Yokozaki, H.; et al. Tight junction proteins claudin-2 and -12 are critical for vitamin D-dependent Ca2+ absorption between enterocytes. Mol. Biol. Cell 2008, 19, 1912–1921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.G.; Wu, S.; Lu, R.; Zhou, D.; Zhou, J.; Carmeliet, G.; Petrof, E.; Claud, E.C.; Sun, J. Tight junction CLDN2 gene is a direct target of the vitamin D receptor. Sci. Rep. 2015, 5, 10642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatterjee, I.; Zhang, Y.; Zhang, J.; Lu, R.; Xia, Y.; Sun, J. Overexpression of Vitamin D Receptor in Intestinal Epithelia Protects Against Colitis via Upregulating Tight Junction Protein Claudin 15. J. Crohns Colitis 2021, 15, 1720–1736. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Garrett, S.; Carroll, R.E.; Xia, Y.; Sun, J. Vitamin D Receptor Upregulates Tight Junction Protein Claudin-5 against Tumorigenesis. bioRxiv 2021. [Google Scholar] [CrossRef]
- Zhang, Y.G.; Wu, S.; Sun, J. Vitamin D, Vitamin D Receptor, and Tissue Barriers. Tissue Barriers 2013, 1, e23118. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Thingholm, L.B.; Skieceviciene, J.; Rausch, P.; Kummen, M.; Hov, J.R.; Degenhardt, F.; Heinsen, F.A.; Ruhlemann, M.C.; Szymczak, S.; et al. Genome-wide association analysis identifies variation in vitamin D receptor and other host factors influencing the gut microbiota. Nat. Genet. 2016, 48, 1396–1406. [Google Scholar] [CrossRef]
- Wang, Y.; Becklund, B.R.; DeLuca, H.F. Identification of a highly specific and versatile vitamin D receptor antibody. Arch. Biochem. Biophys. 2010, 494, 166–177. [Google Scholar] [CrossRef]
- Bouillon, R.; Carmeliet, G.; Verlinden, L.; van Etten, E.; Verstuyf, A.; Luderer, H.F.; Lieben, L.; Mathieu, C.; Demay, M. Vitamin D and human health: Lessons from vitamin D receptor null mice. Endocr. Rev. 2008, 29, 726–776. [Google Scholar] [CrossRef]
- Carlberg, C.; Seuter, S. A genomic perspective on vitamin D signaling. Anticancer Res. 2009, 29, 3485–3493. [Google Scholar]
- Laukoetter, M.G.; Nava, P.; Lee, W.Y.; Severson, E.A.; Capaldo, C.T.; Babbin, B.A.; Williams, I.R.; Koval, M.; Peatman, E.; Campbell, J.A.; et al. JAM-A regulates permeability and inflammation in the intestine in vivo. J. Exp. Med. 2007, 204, 3067–3076. [Google Scholar] [CrossRef]
- Blikslager, A.T.; Moeser, A.J.; Gookin, J.L.; Jones, S.L.; Odle, J. Restoration of barrier function in injured intestinal mucosa. Physiol. Rev. 2007, 87, 545–564. [Google Scholar] [CrossRef] [PubMed]
- Farhadi, A.; Banan, A.; Fields, J.; Keshavarzian, A. Intestinal barrier: An interface between health and disease. J. Gastroenterol. Hepatol. 2003, 18, 479–497. [Google Scholar] [CrossRef] [PubMed]
- Rajapaksa, T.E.; Stover-Hamer, M.; Fernandez, X.; Eckelhoefer, H.A.; Lo, D.D. Claudin 4-targeted protein incorporated into PLGA nanoparticles can mediate M cell targeted delivery. J. Control. Release 2010, 142, 196–205. [Google Scholar] [CrossRef] [Green Version]
- Ling, J.; Liao, H.; Clark, R.; Wong, M.S.; Lo, D.D. Structural constraints for the binding of short peptides to claudin-4 revealed by surface plasmon resonance. J. Biol. Chem. 2008, 283, 30585–30595. [Google Scholar] [CrossRef] [Green Version]
- Ivanov, A.I. Structure and regulation of intestinal epithelial tight junctions: Current concepts and unanswered questions. Adv. Exp. Med. Biol. 2012, 763, 132–148. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, A.I.; Nusrat, A.; Parkos, C.A. The epithelium in inflammatory bowel disease: Potential role of endocytosis of junctional proteins in barrier disruption. Novartis Found Symp 2004, 263, 115–124, discussion 124–132, 211–118. [Google Scholar] [PubMed]
- Chung, H.; Kasper, D.L. Microbiota-stimulated immune mechanisms to maintain gut homeostasis. Curr. Opin. Immunol. 2010, 22, 455–460. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.X.; Lee, J.S.; Campbell, E.L.; Colgan, S.P. Microbiota-derived butyrate dynamically regulates intestinal homeostasis through regulation of actin-associated protein synaptopodin. Proc. Natl. Acad. Sci. USA 2020, 117, 11648–11657. [Google Scholar] [CrossRef]
- Rahner, C.; Mitic, L.L.; Anderson, J.M. Heterogeneity in expression and subcellular localization of claudins 2, 3, 4, and 5 in the rat liver, pancreas, and gut. Gastroenterology 2001, 120, 411–422. [Google Scholar] [CrossRef]
- Sakaguchi, T.; Gu, X.; Golden, H.M.; Suh, E.; Rhoads, D.B.; Reinecker, H.C. Cloning of the human claudin-2 5’-flanking region revealed a TATA-less promoter with conserved binding sites in mouse and human for caudal-related homeodomain proteins and hepatocyte nuclear factor-1alpha. J. Biol. Chem. 2002, 277, 21361–21370. [Google Scholar] [CrossRef] [Green Version]
- Chiba, H.; Osanai, M.; Murata, M.; Kojima, T.; Sawada, N. Transmembrane proteins of tight junctions. Biochim. Biophys. Acta 2008, 1778, 588–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenthal, R.; Milatz, S.; Krug, S.M.; Oelrich, B.; Schulzke, J.D.; Amasheh, S.; Gunzel, D.; Fromm, M. Claudin-2, a component of the tight junction, forms a paracellular water channel. J. Cell Sci. 2010, 123, 1913–1921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abu-Farsakh, S.; Wu, T.; Lalonde, A.; Sun, J.; Zhou, Z. High expression of Claudin-2 in esophageal carcinoma and precancerous lesions is significantly associated with the bile salt receptors VDR and TGR5. BMC Gastroenterol. 2017, 17, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchert, M.; Papin, M.; Bonnans, C.; Darido, C.; Raye, W.S.; Garambois, V.; Pelegrin, A.; Bourgaux, J.F.; Pannequin, J.; Joubert, D.; et al. Symplekin promotes tumorigenicity by up-regulating claudin-2 expression. Proc. Natl. Acad. Sci. USA 2010, 107, 2628–2633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhawan, P.; Ahmad, R.; Chaturvedi, R.; Smith, J.J.; Midha, R.; Mittal, M.K.; Krishnan, M.; Chen, X.; Eschrich, S.; Yeatman, T.J.; et al. Claudin-2 expression increases tumorigenicity of colon cancer cells: Role of epidermal growth factor receptor activation. Oncogene 2011, 30, 3234–3247. [Google Scholar] [CrossRef] [Green Version]
- Weber, C.R.; Nalle, S.C.; Tretiakova, M.; Rubin, D.T.; Turner, J.R. Claudin-1 and claudin-2 expression is elevated in inflammatory bowel disease and may contribute to early neoplastic transformation. Lab. Investig. 2008, 88, 1110–1120. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.G.; Lu, R.; Xia, Y.; Zhou, D.; Petrof, E.; Claud, E.C.; Sun, J. Lack of Vitamin D Receptor Leads to Hyperfunction of Claudin-2 in Intestinal Inflammatory Responses. Inflamm. Bowel Dis. 2019, 25, 97–110. [Google Scholar] [CrossRef]
- Zhang, Y.G.; Wu, S.; Xia, Y.; Sun, J. Salmonella infection upregulates the leaky protein claudin-2 in intestinal epithelial cells. PLoS ONE 2013, 8, e58606. [Google Scholar] [CrossRef] [Green Version]
- Furuse, M.; Fujita, K.; Hiiragi, T.; Fujimoto, K.; Tsukita, S. Claudin-1 and -2: Novel integral membrane proteins localizing at tight junctions with no sequence similarity to occludin. J. Cell Biol. 1998, 141, 1539–1550. [Google Scholar] [CrossRef]
- Angelow, S.; Ahlstrom, R.; Yu, A.S. Biology of claudins. Am. J. Physiol. Ren. Physiol. 2008, 295, F867–F876. [Google Scholar] [CrossRef]
- Mineta, K.; Yamamoto, Y.; Yamazaki, Y.; Tanaka, H.; Tada, Y.; Saito, K.; Tamura, A.; Igarashi, M.; Endo, T.; Takeuchi, K.; et al. Predicted expansion of the claudin multigene family. FEBS Lett. 2011, 585, 606–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xing, T.; Benderman, L.J.; Sabu, S.; Parker, J.; Yang, J.; Lu, Q.; Ding, L.; Chen, Y.H. Tight Junction Protein Claudin-7 Is Essential for Intestinal Epithelial Stem Cell Self-Renewal and Differentiation. Cell. Mol. Gastroenterol. Hepatol. 2020, 9, 641–659. [Google Scholar] [CrossRef] [PubMed]
- Holmes, J.L.; Van Itallie, C.M.; Rasmussen, J.E.; Anderson, J.M. Claudin profiling in the mouse during postnatal intestinal development and along the gastrointestinal tract reveals complex expression patterns. Gene Expr. Patterns 2006, 6, 581–588. [Google Scholar] [CrossRef] [PubMed]
- Demay, M.B. Mechanism of vitamin D receptor action. Ann. N. Y. Acad. Sci. 2006, 1068, 204–213. [Google Scholar] [CrossRef] [PubMed]
- Kato, S. The function of vitamin D receptor in vitamin D action. J. Biochem. 2000, 127, 717–722. [Google Scholar] [CrossRef] [PubMed]
- Gombart, A.F.; Borregaard, N.; Koeffler, H.P. Human cathelicidin antimicrobial peptide (CAMP) gene is a direct target of the vitamin D receptor and is strongly up-regulated in myeloid cells by 1,25-dihydroxyvitamin D3. FASEB J. 2005, 19, 1067–1077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.T.; Nestel, F.P.; Bourdeau, V.; Nagai, Y.; Wang, Q.; Liao, J.; Tavera-Mendoza, L.; Lin, R.; Hanrahan, J.W.; Mader, S.; et al. Cutting edge: 1,25-dihydroxyvitamin D3 is a direct inducer of antimicrobial peptide gene expression. J. Immunol. 2004, 173, 2909–2912. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Zhang, Y.G.; Lu, R.; Xia, Y.; Zhou, D.; Petrof, E.O.; Claud, E.C.; Chen, D.; Chang, E.B.; Carmeliet, G.; et al. Intestinal epithelial vitamin D receptor deletion leads to defective autophagy in colitis. Gut 2015, 64, 1082–1094. [Google Scholar] [CrossRef]
- Jin, D.; Zhang, Y.G.; Wu, S.; Lu, R.; Lin, Z.; Zheng, Y.; Chen, H.; Cs-Szabo, G.; Sun, J. Vitamin D receptor is a novel transcriptional regulator for Axin1. J. Steroid Biochem. Mol. Biol. 2017, 165, 430–437. [Google Scholar] [CrossRef] [Green Version]
- Kamen, D.L.; Tangpricha, V. Vitamin D and molecular actions on the immune system: Modulation of innate and autoimmunity. J. Mol. Med. 2010, 88, 441–450. [Google Scholar] [CrossRef] [Green Version]
- Waterhouse, J.C.; Perez, T.H.; Albert, P.J. Reversing bacteria-induced vitamin D receptor dysfunction is key to autoimmune disease. Ann. N. Y. Acad. Sci. 2009, 1173, 757–765. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.T.; Krutzik, S.R.; Modlin, R.L. Therapeutic implications of the TLR and VDR partnership. Trends Mol. Med. 2007, 13, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Kong, J.; Zhang, Z.; Musch, M.W.; Ning, G.; Sun, J.; Hart, J.; Bissonnette, M.; Li, Y.C. Novel role of the vitamin D receptor in maintaining the integrity of the intestinal mucosal barrier. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 294, G208–G216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lagishetty, V.; Misharin, A.V.; Liu, N.Q.; Lisse, T.S.; Chun, R.F.; Ouyang, Y.; McLachlan, S.M.; Adams, J.S.; Hewison, M. Vitamin d deficiency in mice impairs colonic antibacterial activity and predisposes to colitis. Endocrinology 2010, 151, 2423–2432. [Google Scholar] [CrossRef] [Green Version]
- Gocek, E.; Studzinski, G.P. Vitamin D and differentiation in cancer. Crit. Rev. Clin. Lab. Sci. 2009, 46, 190–209. [Google Scholar] [CrossRef] [Green Version]
- Samuel, S.; Sitrin, M.D. Vitamin D’s role in cell proliferation and differentiation. Nutr. Rev. 2008, 66, S116–S124. [Google Scholar] [CrossRef]
- Yu, S.; Bruce, D.; Froicu, M.; Weaver, V.; Cantorna, M.T. Failure of T cell homing, reduced CD4/CD8alphaalpha intraepithelial lymphocytes, and inflammation in the gut of vitamin D receptor KO mice. Proc. Natl. Acad. Sci. USA 2008, 105, 20834–20839. [Google Scholar] [CrossRef] [Green Version]
- Ogura, M.; Nishida, S.; Ishizawa, M.; Sakurai, K.; Shimizu, M.; Matsuo, S.; Amano, S.; Uno, S.; Makishima, M. Vitamin D3 modulates the expression of bile acid regulatory genes and represses inflammation in bile duct-ligated mice. J. Pharm. Exp. 2009, 328, 564–570. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Kong, J.; Duan, Y.; Szeto, F.L.; Liao, A.; Madara, J.L.; Li, Y.C. Increased NF-kappaB activity in fibroblasts lacking the vitamin D receptor. Am. J. Physiol. Endocrinol. Metab. 2006, 291, E315–E322. [Google Scholar] [CrossRef]
- Liu, F.H.; Li, S.S.; Li, X.X.; Wang, S.; Li, M.G.; Guan, L.; Luan, T.G.; Liu, Z.G.; Liu, Z.J.; Yang, P.C. Vitamin D3 induces vitamin D receptor and HDAC11 binding to relieve the promoter of the tight junction proteins. Oncotarget 2017, 8, 58781–58789. [Google Scholar] [CrossRef]
- Wang, Z.; Yang, H.; Jin, M.; Zhang, H.M.; Chen, X.F.; Wu, M.X.; Guo, M.Y.; Huang, C.Z.; Qian, J.M. Effects of Vitamin D Receptor on Mucosal Barrier Proteins in Colon Cells under Hypoxic Environment. Zhongguo Yi Xue Ke Xue Yuan Xue Bao 2019, 41, 506–511. [Google Scholar] [CrossRef]
- Palmer, H.G.; Gonzalez-Sancho, J.M.; Espada, J.; Berciano, M.T.; Puig, I.; Baulida, J.; Quintanilla, M.; Cano, A.; de Herreros, A.G.; Lafarga, M.; et al. Vitamin D(3) promotes the differentiation of colon carcinoma cells by the induction of E-cadherin and the inhibition of beta-catenin signaling. J. Cell Biol. 2001, 154, 369–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, P.C.; Hsieh, Y.C.; Huo, T.I.; Yang, U.C.; Lin, C.H.; Li, C.P.; Huang, Y.H.; Hou, M.C.; Lin, H.C.; Lee, K.C. Active Vitamin D3 Treatment Attenuated Bacterial Translocation via Improving Intestinal Barriers in Cirrhotic Rats. Mol. Nutr. Food Res. 2021, 65, e2000937. [Google Scholar] [CrossRef] [PubMed]
- Ordonez-Moran, P.; Alvarez-Diaz, S.; Valle, N.; Larriba, M.J.; Bonilla, F.; Munoz, A. The effects of 1,25-dihydroxyvitamin D3 on colon cancer cells depend on RhoA-ROCK-p38MAPK-MSK signaling. J. Steroid Biochem. Mol. Biol. 2010, 121, 355–361. [Google Scholar] [CrossRef]
- Haussler, M.R.; Whitfield, G.K.; Kaneko, I.; Haussler, C.A.; Hsieh, D.; Hsieh, J.C.; Jurutka, P.W. Molecular Mechanisms of Vitamin D Action. Calcif. Tissue Int. 2012, 92, 77–98. [Google Scholar] [CrossRef] [PubMed]
- Pereira, F.; Barbachano, A.; Silva, J.; Bonilla, F.; Campbell, M.J.; Munoz, A.; Larriba, M.J. KDM6B/JMJD3 histone demethylase is induced by vitamin D and modulates its effects in colon cancer cells. Hum. Mol. Genet. 2011, 20, 4655–4665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christakos, S.; Dhawan, P.; Ajibade, D.; Benn, B.S.; Feng, J.; Joshi, S.S. Mechanisms involved in vitamin D mediated intestinal calcium absorption and in non-classical actions of vitamin D. J. Steroid Biochem. Mol. Biol. 2010, 121, 183–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kutuzova, G.D.; Deluca, H.F. Gene expression profiles in rat intestine identify pathways for 1,25-dihydroxyvitamin D(3) stimulated calcium absorption and clarify its immunomodulatory properties. Arch. Biochem. Biophys. 2004, 432, 152–166. [Google Scholar] [CrossRef]
- Chen, S.W.; Ma, Y.Y.; Zhu, J.; Zuo, S.; Zhang, J.L.; Chen, Z.Y.; Chen, G.W.; Wang, X.; Pan, Y.S.; Liu, Y.C.; et al. Protective effect of 1,25-dihydroxyvitamin D3 on ethanol-induced intestinal barrier injury both in vitro and in vivo. Toxicol. Lett. 2015, 237, 79–88. [Google Scholar] [CrossRef]
- He, L.; Liu, T.; Shi, Y.; Tian, F.; Hu, H.; Deb, D.K.; Chen, Y.; Bissonnette, M.; Li, Y.C. Gut Epithelial Vitamin D Receptor Regulates Microbiota-Dependent Mucosal Inflammation by Suppressing Intestinal Epithelial Cell Apoptosis. Endocrinology 2018, 159, 967–979. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Zhang, H.; Wu, H.; Li, H.; Liu, L.; Guo, J.; Li, C.; Shih, D.Q.; Zhang, X. Protective role of 1,25(OH)2vitamin D3 in the mucosal injury and epithelial barrier disruption in DSS-induced acute colitis in mice. BMC Gastroenterol. 2012, 12, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sari, E.; Oztay, F.; Tasci, A.E. Vitamin D modulates E-cadherin turnover by regulating TGF-beta and Wnt signalings during EMT-mediated myofibroblast differentiation in A459 cells. J. Steroid Biochem. Mol. Biol. 2020, 202, 105723. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Zhao, H.; Dong, H.; Zou, F.; Cai, S. 1alpha,25-dihydroxyvitamin D(3) counteracts the effects of cigarette smoke in airway epithelial cells. Cell Immunol. 2015, 295, 137–143. [Google Scholar] [CrossRef]
- Shi, Y.Y.; Liu, T.J.; Fu, J.H.; Xu, W.; Wu, L.L.; Hou, A.N.; Xue, X.D. Vitamin D/VDR signaling attenuates lipopolysaccharide-induced acute lung injury by maintaining the integrity of the pulmonary epithelial barrier. Mol. Med. Rep. 2016, 13, 1186–1194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Lu, R.; Zhang, Y.G.; Sun, J. Vitamin D Receptor Deletion Leads to the Destruction of Tight and Adherens Junctions in Lungs. Tissue Barriers 2018, 6, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Migliori, M.; Giovannini, L.; Panichi, V.; Filippi, C.; Taccola, D.; Origlia, N.; Mannari, C.; Camussi, G. Treatment with 1,25-dihydroxyvitamin D3 preserves glomerular slit diaphragm-associated protein expression in experimental glomerulonephritis. Int. J. Immunopathol. Pharm. 2005, 18, 779–790. [Google Scholar] [CrossRef]
- Kladnitsky, O.; Rozenfeld, J.; Azulay-Debby, H.; Efrati, E.; Zelikovic, I. The claudin-16 channel gene is transcriptionally inhibited by 1,25-dihydroxyvitamin D. Exp. Physiol. 2015, 100, 79–94. [Google Scholar] [CrossRef]
- Elizondo, R.A.; Yin, Z.; Lu, X.; Watsky, M.A. Effect of vitamin D receptor knockout on cornea epithelium wound healing and tight junctions. Investig. Ophthalmol. Vis. Sci. 2014, 55, 5245–5251. [Google Scholar] [CrossRef] [Green Version]
- Yin, Z.; Pintea, V.; Lin, Y.; Hammock, B.D.; Watsky, M.A. Vitamin D enhances corneal epithelial barrier function. Investig. Ophthalmol. Vis. Sci. 2011, 52, 7359–7364. [Google Scholar] [CrossRef] [Green Version]
- Visconti, B.; Paolino, G.; Carotti, S.; Pendolino, A.L.; Morini, S.; Richetta, A.G.; Calvieri, S. Immunohistochemical expression of VDR is associated with reduced integrity of tight junction complex in psoriatic skin. J. Eur. Acad. Dermatol. Venereol. 2015, 29, 2038–2042. [Google Scholar] [CrossRef]
- Won, S.; Sayeed, I.; Peterson, B.L.; Wali, B.; Kahn, J.S.; Stein, D.G. Vitamin D prevents hypoxia/reoxygenation-induced blood-brain barrier disruption via vitamin D receptor-mediated NF-kB signaling pathways. PLoS ONE 2015, 10, e0122821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, C.; Kim, H.J.; Kim, H.M. Vitamin D maintains E-cadherin intercellular junctions by downregulating MMP-9 production in human gingival keratinocytes treated by TNF-alpha. J. Periodontal. Implant Sci. 2019, 49, 270–286. [Google Scholar] [CrossRef] [PubMed]
- Mohanty, S.; Kamolvit, W.; Hertting, O.; Brauner, A. Vitamin D strengthens the bladder epithelial barrier by inducing tight junction proteins during E. coli urinary tract infection. Cell Tissue Res. 2020, 380, 669–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blaney, G.P.; Albert, P.J.; Proal, A.D. Vitamin D metabolites as clinical markers in autoimmune and chronic disease. Ann. N. Y. Acad. Sci. 2009, 1173, 384–390. [Google Scholar] [CrossRef]
- Adorini, L.; Penna, G. Control of autoimmune diseases by the vitamin D endocrine system. Nat. Clin. Pract. Rheumatol. 2008, 4, 404–412. [Google Scholar] [CrossRef] [PubMed]
- Grau, M.V.; Baron, J.A.; Sandler, R.S.; Haile, R.W.; Beach, M.L.; Church, T.R.; Heber, D. Vitamin D, calcium supplementation, and colorectal adenomas: Results of a randomized trial. J. Natl. Cancer Inst. 2003, 95, 1765–1771. [Google Scholar] [CrossRef]
- Heaney, R.P. Vitamin D in health and disease. Clin. J. Am. Soc. Nephrol. 2008, 3, 1535–1541. [Google Scholar] [CrossRef] [Green Version]
- Cannell, J.J.; Hollis, B.W.; Zasloff, M.; Heaney, R.P. Diagnosis and treatment of vitamin D deficiency. Expert Opin. Pharm. 2008, 9, 107–118. [Google Scholar] [CrossRef]
- Campbell, F.C.; Xu, H.; El-Tanani, M.; Crowe, P.; Bingham, V. The yin and yang of vitamin D receptor (VDR) signaling in neoplastic progression: Operational networks and tissue-specific growth control. Biochem. Pharm. 2010, 79, 1–9. [Google Scholar] [CrossRef]
- Zhang, Y.G.; Lu, R.; Wu, S.; Chatterjee, I.; Zhou, D.; Xia, Y.; Sun, J. Vitamin D Receptor Protects Against Dysbiosis and Tumorigenesis via the JAK/STAT Pathway in Intestine. Cell. Mol. Gastroenterol. Hepatol. 2020, 10, 729–746. [Google Scholar] [CrossRef]
- Song, M.; Chan, A.T.; Sun, J. Influence of the Gut Microbiome, Diet, and Environment on Risk of Colorectal Cancer. Gastroenterology 2020, 158, 322–340. [Google Scholar] [CrossRef]
- Corridoni, D.; Arseneau, K.O.; Cifone, M.G.; Cominelli, F. The dual role of nod-like receptors in mucosal innate immunity and chronic intestinal inflammation. Front. Immunol. 2014, 5, 317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corridoni, D.; Arseneau, K.O.; Cominelli, F. Inflammatory bowel disease. Immunol. Lett. 2014, 161, 231–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNamee, E.N.; Masterson, J.C.; Veny, M.; Collins, C.B.; Jedlicka, P.; Byrne, F.R.; Ng, G.Y.; Rivera-Nieves, J. Chemokine receptor CCR7 regulates the intestinal TH1/TH17/Treg balance during Crohn’s-like murine ileitis. J. Leukoc. Biol. 2015, 97, 1011–1022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sartor, R.B. Mechanisms of disease: Pathogenesis of Crohn’s disease and ulcerative colitis. Nat. Clin. Pract. Gastroenterol. Hepatol. 2006, 3, 390–407. [Google Scholar] [CrossRef] [PubMed]
- Dresner-Pollak, R.; Ackerman, Z.; Eliakim, R.; Karban, A.; Chowers, Y.; Fidder, H.H. The BsmI vitamin D receptor gene polymorphism is associated with ulcerative colitis in Jewish Ashkenazi patients. Genet. Test. 2004, 8, 417–420. [Google Scholar] [CrossRef]
- Simmons, J.D.; Mullighan, C.; Welsh, K.I.; Jewell, D.P. Vitamin D receptor gene polymorphism: Association with Crohn’s disease susceptibility. Gut 2000, 47, 211–214. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.H.; Song, G.G. Pathway analysis of a genome-wide association study of ileal Crohn’s disease. DNA Cell Biol. 2012, 31, 1549–1554. [Google Scholar] [CrossRef]
- Azad, A.K.; Sadee, W.; Schlesinger, L.S. Innate immune gene polymorphisms in tuberculosis. Infect. Immun. 2012, 80, 3343–3359. [Google Scholar] [CrossRef] [Green Version]
- Eloranta, J.J.; Wenger, C.; Mwinyi, J.; Hiller, C.; Gubler, C.; Vavricka, S.R.; Fried, M.; Kullak-Ublick, G.A. Association of a common vitamin D-binding protein polymorphism with inflammatory bowel disease. Pharm. Genom. 2011, 21, 559–564. [Google Scholar] [CrossRef] [Green Version]
- Abreu, M.T.; Kantorovich, V.; Vasiliauskas, E.A.; Gruntmanis, U.; Matuk, R.; Daigle, K.; Chen, S.; Zehnder, D.; Lin, Y.C.; Yang, H.; et al. Measurement of vitamin D levels in inflammatory bowel disease patients reveals a subset of Crohn’s disease patients with elevated 1,25-dihydroxyvitamin D and low bone mineral density. Gut 2004, 53, 1129–1136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, W.C.; Hanauer, S.B.; Li, Y.C. Mechanisms of disease: Vitamin D and inflammatory bowel disease. Nat. Clin. Pract. Gastroenterol. Hepatol. 2005, 2, 308–315. [Google Scholar] [CrossRef] [PubMed]
- Sentongo, T.A.; Semaeo, E.J.; Stettler, N.; Piccoli, D.A.; Stallings, V.A.; Zemel, B.S. Vitamin D status in children, adolescents, and young adults with Crohn disease. Am. J. Clin. Nutr. 2002, 76, 1077–1081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakke, D.; Sun, J. Ancient Nuclear Receptor VDR with New Functions: Microbiome and Inflammation. Inflamm. Bowel Dis. 2018, 24, 1149–1154. [Google Scholar] [CrossRef] [Green Version]
- Stio, M.; Martinesi, M.; Bruni, S.; Treves, C.; d’Albasio, G.; Bagnoli, S.; Bonanomi, A.G. Interaction among vitamin D(3) analogue KH 1060, TNF-alpha, and vitamin D receptor protein in peripheral blood mononuclear cells of inflammatory bowel disease patients. Int. Immunopharmacol. 2006, 6, 1083–1092. [Google Scholar] [CrossRef]
- Cantorna, M.T.; Zhu, Y.; Froicu, M.; Wittke, A. Vitamin D status, 1,25-dihydroxyvitamin D3, and the immune system. Am. J. Clin. Nutr. 2004, 80, 1717S–1720S. [Google Scholar] [CrossRef]
- Cantorna, M.T. Vitamin D and its role in immunology: Multiple sclerosis, and inflammatory bowel disease. Prog. Biophys. Mol. Biol. 2006, 92, 60–64. [Google Scholar] [CrossRef]
- Kong, J.; Qiao, G.; Zhang, Z.; Liu, S.Q.; Li, Y.C. Targeted vitamin D receptor expression in juxtaglomerular cells suppresses renin expression independent of parathyroid hormone and calcium. Kidney Int. 2008, 74, 1577–1581. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.T.; Modlin, R.L. Human macrophage host defense against Mycobacterium tuberculosis. Curr. Opin. Immunol. 2008, 20, 371–376. [Google Scholar] [CrossRef]
- Laverny, G.; Penna, G.; Vetrano, S.; Correale, C.; Nebuloni, M.; Danese, S.; Adorini, L. Efficacy of a potent and safe vitamin D receptor agonist for the treatment of inflammatory bowel disease. Immunol. Lett. 2010, 113, 49–58. [Google Scholar] [CrossRef]
- Liu, W.; Chen, Y.; Golan, M.A.; Annunziata, M.L.; Du, J.; Dougherty, U.; Kong, J.; Musch, M.; Huang, Y.; Pekow, J.; et al. Intestinal epithelial vitamin D receptor signaling inhibits experimental colitis. J. Clin. Investig. 2013, 123, 3983–3996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Froicu, M.; Zhu, Y.; Cantorna, M.T. Vitamin D receptor is required to control gastrointestinal immunity in IL-10 knockout mice. Immunology 2006, 117, 310–318. [Google Scholar] [CrossRef] [PubMed]
- Lechuga, S.; Ivanov, A.I. Disruption of the epithelial barrier during intestinal inflammation: Quest for new molecules and mechanisms. Biochim. Biophys. Acta Mol. Cell Res. 2017, 1864, 1183–1194. [Google Scholar] [CrossRef] [PubMed]
- Katz, K.D.; Hollander, D.; Vadheim, C.M.; McElree, C.; Delahunty, T.; Dadufalza, V.D.; Krugliak, P.; Rotter, J.I. Intestinal permeability in patients with Crohn’s disease and their healthy relatives. Gastroenterology 1989, 97, 927–931. [Google Scholar] [CrossRef]
- Martz, S.L.; McDonald, J.A.; Sun, J.; Zhang, Y.G.; Gloor, G.B.; Noordhof, C.; He, S.M.; Gerbaba, T.K.; Blennerhassett, M.; Hurlbut, D.J.; et al. Administration of defined microbiota is protective in a murine Salmonella infection model. Sci. Rep. 2015, 5, 16094. [Google Scholar] [CrossRef]
- Zeissig, S.; Burgel, N.; Gunzel, D.; Richter, J.; Mankertz, J.; Wahnschaffe, U.; Kroesen, A.J.; Zeitz, M.; Fromm, M.; Schulzke, J.D. Changes in expression and distribution of claudin 2, 5 and 8 lead to discontinuous tight junctions and barrier dysfunction in active Crohn’s disease. Gut 2007, 56, 61–72. [Google Scholar] [CrossRef]
- Schulzke, J.D.; Fromm, M. Tight junctions: Molecular structure meets function. Ann. N. Y. Acad. Sci. 2009, 1165, 1–6. [Google Scholar] [CrossRef]
- Van Itallie, C.M.; Holmes, J.; Bridges, A.; Anderson, J.M. Claudin-2-dependent changes in noncharged solute flux are mediated by the extracellular domains and require attachment to the PDZ-scaffold. Ann. N. Y. Acad. Sci. 2009, 1165, 82–87. [Google Scholar] [CrossRef] [Green Version]
- Marchelletta, R.R.; Krishnan, M.; Spalinger, M.R.; Placone, T.W.; Alvarez, R.; Sayoc-Becerra, A.; Canale, V.; Shawki, A.; Park, Y.S.; Bernts, L.H.; et al. T cell protein tyrosine phosphatase protects intestinal barrier function by restricting epithelial tight junction remodeling. J. Clin. Investig. 2021, 131, e138230. [Google Scholar] [CrossRef]
- Darsigny, M.; Babeu, J.P.; Dupuis, A.A.; Furth, E.E.; Seidman, E.G.; Levy, E.; Verdu, E.F.; Gendron, F.P.; Boudreau, F. Loss of hepatocyte-nuclear-factor-4alpha affects colonic ion transport and causes chronic inflammation resembling inflammatory bowel disease in mice. PLoS ONE 2009, 4, e7609. [Google Scholar] [CrossRef]
- Turpin, W.; Lee, S.H.; Raygoza Garay, J.A.; Madsen, K.L.; Meddings, J.B.; Bedrani, L.; Power, N.; Espin-Garcia, O.; Xu, W.; Smith, M.I.; et al. Increased Intestinal Permeability is Associated with Later Development of Crohn’s Disease. Gastroenterology 2020, 159, 2092–2100. [Google Scholar] [CrossRef]
- Sun, J.; Kato, I. Gut microbiota, inflammation and colorectal cancer. Genes Dis. 2016, 3, 130–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J. Impact of bacterial infection and intestinal microbiome on colorectal cancer development. Chin. Med. J. 2021, 135, 400. [Google Scholar] [CrossRef] [PubMed]
- Mandle, H.B.; Jahan, F.A.; Bostick, R.M.; Baron, J.A.; Barry, E.L.; Yacoub, R.; Merrill, J.; Rutherford, R.E.; Seabrook, M.E.; Fedirko, V. Effects of supplemental calcium and vitamin D on tight-junction proteins and mucin-12 expression in the normal rectal mucosa of colorectal adenoma patients. Mol. Carcinog. 2019, 58, 1279–1290. [Google Scholar] [CrossRef] [PubMed]
- Wada, K.; Tanaka, H.; Maeda, K.; Inoue, T.; Noda, E.; Amano, R.; Kubo, N.; Muguruma, K.; Yamada, N.; Yashiro, M.; et al. Vitamin D receptor expression is associated with colon cancer in ulcerative colitis. Oncol. Rep. 2009, 22, 1021–1025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, L.; Han, J.; Li, L.; Wang, Y.; Li, Y.; Zhang, S. Claudin Family Participates in the Pathogenesis of Inflammatory Bowel Diseases and Colitis-Associated Colorectal Cancer. Front. Immunol. 2019, 10, 1441. [Google Scholar] [CrossRef]
- Cherradi, S.; Martineau, P.; Gongora, C.; Del Rio, M. Claudin gene expression profiles and clinical value in colorectal tumors classified according to their molecular subtype. Cancer Manag. Res. 2019, 11, 1337–1348. [Google Scholar] [CrossRef] [Green Version]
- Sun, J. Vitamin D and mucosal immune function. Curr. Opin. Gastroenterol. 2010, 26, 591–595. [Google Scholar] [CrossRef] [Green Version]
- You, K.; Xu, X.; Fu, J.; Xu, S.; Yue, X.; Yu, Z.; Xue, X. Hyperoxia disrupts pulmonary epithelial barrier in newborn rats via the deterioration of occludin and ZO-1. Respir. Res. 2012, 13, 36. [Google Scholar] [CrossRef] [Green Version]
- Overgaard, C.E.; Mitchell, L.A.; Koval, M. Roles for claudins in alveolar epithelial barrier function. Ann. N. Y. Acad. Sci. 2012, 1257, 167–174. [Google Scholar] [CrossRef]
- Li, W.; Dong, H.; Zhao, H.; Song, J.; Tang, H.; Yao, L.; Liu, L.; Tong, W.; Zou, M.; Zou, F.; et al. 1,25-Dihydroxyvitamin D3 prevents toluene diisocyanate-induced airway epithelial barrier disruption. Int. J. Mol. Med. 2015, 36, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.; Yang, J.; Hu, X.; Li, M.; Wang, Q.; Dancer, R.C.A.; Parekh, D.; Gao-Smith, F.; Thickett, D.R.; Jin, S. Vitamin D attenuates lung injury via stimulating epithelial repair, reducing epithelial cell apoptosis and inhibits TGF-beta induced epithelial to mesenchymal transition. Biochem. Pharm. 2020, 177, 113955. [Google Scholar] [CrossRef] [PubMed]
- Gorman, S.; Buckley, A.G.; Ling, K.M.; Berry, L.J.; Fear, V.S.; Stick, S.M.; Larcombe, A.N.; Kicic, A.; Hart, P.H. Vitamin D supplementation of initially vitamin D-deficient mice diminishes lung inflammation with limited effects on pulmonary epithelial integrity. Physiol. Rep. 2017, 5, e13371. [Google Scholar] [CrossRef] [PubMed]
- DiFranco, K.M.; Mulligan, J.K.; Sumal, A.S.; Diamond, G. Induction of CFTR gene expression by 1,25(OH)2 vitamin D3, 25OH vitamin D3, and vitamin D3 in cultured human airway epithelial cells and in mouse airways. J. Steroid Biochem. Mol. Biol. 2017, 173, 323–332. [Google Scholar] [CrossRef]
- Herr, C.; Greulich, T.; Koczulla, R.A.; Meyer, S.; Zakharkina, T.; Branscheidt, M.; Eschmann, R.; Bals, R. The role of vitamin D in pulmonary disease: COPD, asthma, infection, and cancer. Respir. Res. 2011, 12, 31. [Google Scholar] [CrossRef] [Green Version]
- Zhu, M.; Ji, Y. The Association Between Vitamin D And COPD Risk, Severity And Exacerbation: A Systematic Review And Meta-Analysis Update. Am. J. Resp. Crit. Care 2016, 193, A3496. [Google Scholar] [CrossRef] [Green Version]
- Sundar, I.K.; Rahman, I. Vitamin d and susceptibility of chronic lung diseases: Role of epigenetics. Front. Pharm. 2011, 2, 50. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, C.R.; Arum, S.M.; Smith, C.M. Vitamin D deficiency and chronic lung disease. Can. Respir. J. 2009, 16, 75–80. [Google Scholar] [CrossRef] [Green Version]
- Jeffery, P.K. Structural and inflammatory changes in COPD: A comparison with asthma. Thorax 1998, 53, 129–136. [Google Scholar] [CrossRef] [Green Version]
- Janssens, W.; Bouillon, R.; Claes, B.; Carremans, C.; Lehouck, A.; Buysschaert, I.; Coolen, J.; Mathieu, C.; Decramer, M.; Lambrechts, D. Vitamin D deficiency is highly prevalent in COPD and correlates with variants in the vitamin D-binding gene. Thorax 2010, 65, 215–220. [Google Scholar] [CrossRef] [Green Version]
- Kunisaki, K.M.; Niewoehner, D.E.; Singh, R.J.; Connett, J.E. Vitamin D status and longitudinal lung function decline in the Lung Health Study. Eur. Respir. J. 2011, 37, 238–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sundar, I.K.; Hwang, J.W.; Wu, S.; Sun, J.; Rahman, I. Deletion of vitamin D receptor leads to premature emphysema/COPD by increased matrix metalloproteinases and lymphoid aggregates formation. Biochem. Biophys. Res. Commun. 2011, 406, 127–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishii, M.; Yamaguchi, Y.; Isumi, K.; Ogawa, S.; Akishita, M. Transgenic Mice Overexpressing Vitamin D Receptor (VDR) Show Anti-Inflammatory Effects in Lung Tissues. Inflammation 2017, 40, 2012–2019. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; He, J.; Yu, M.; Sun, J. The efficacy of vitamin D therapy for patients with COPD: A meta-analysis of randomized controlled trials. Ann. Palliat. Med. 2020, 9, 286–297. [Google Scholar] [CrossRef]
- Lehouck, A.; Mathieu, C.; Carremans, C.; Baeke, F.; Verhaegen, J.; Van Eldere, J.; Decallonne, B.; Bouillon, R.; Decramer, M.; Janssens, W. High doses of vitamin D to reduce exacerbations in chronic obstructive pulmonary disease: A randomized trial. Ann. Intern. Med. 2012, 156, 105–114. [Google Scholar] [CrossRef] [Green Version]
- Martineau, A.R.; James, W.Y.; Hooper, R.L.; Barnes, N.C.; Jolliffe, D.A.; Greiller, C.L.; Islam, K.; McLaughlin, D.; Bhowmik, A.; Timms, P.M.; et al. Vitamin D3 supplementation in patients with chronic obstructive pulmonary disease (ViDiCO): A multicentre, double-blind, randomised controlled trial. Lancet Respir. Med. 2015, 3, 120–130. [Google Scholar] [CrossRef]
- Hallstrom, K.; McCormick, B.A. Salmonella Interaction with and Passage through the Intestinal Mucosa: Through the Lens of the Organism. Front. Microbiol. 2011, 2, 88. [Google Scholar] [CrossRef] [Green Version]
- Assa, A.; Vong, L.; Pinnell, L.J.; Avitzur, N.; Johnson-Henry, K.C.; Sherman, P.M. Vitamin D deficiency promotes epithelial barrier dysfunction and intestinal inflammation. J. Infect. Dis. 2014, 210, 1296–1305. [Google Scholar] [CrossRef] [Green Version]
- Assa, A.; Vong, L.; Pinnell, L.J.; Rautava, J.; Avitzur, N.; Johnson-Henry, K.C.; Sherman, P.M. Vitamin D deficiency predisposes to adherent-invasive Escherichia coli-induced barrier dysfunction and experimental colonic injury. Inflamm. Bowel Dis. 2015, 21, 297–306. [Google Scholar] [CrossRef]
- Guttman, J.A.; Finlay, B.B. Tight junctions as targets of infectious agents. Biochim. Biophys. Acta 2009, 1788, 832–841. [Google Scholar] [CrossRef] [Green Version]
- Kohler, H.; Sakaguchi, T.; Hurley, B.P.; Kase, B.A.; Reinecker, H.C.; McCormick, B.A. Salmonella enterica serovar Typhimurium regulates intercellular junction proteins and facilitates transepithelial neutrophil and bacterial passage. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 293, G178–G187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyle, E.C.; Brown, N.F.; Finlay, B.B. Salmonella enterica serovar Typhimurium effectors SopB, SopE, SopE2 and SipA disrupt tight junction structure and function. Cell Microbiol. 2006, 8, 1946–1957. [Google Scholar] [CrossRef] [PubMed]
- Bertelsen, L.S.; Paesold, G.; Marcus, S.L.; Finlay, B.B.; Eckmann, L.; Barrett, K.E. Modulation of chloride secretory responses and barrier function of intestinal epithelial cells by the Salmonella effector protein SigD. Am. J. Physiol. Cell Physiol. 2004, 287, C939–C948. [Google Scholar] [CrossRef] [PubMed]
- Liao, A.P.; Petrof, E.O.; Kuppireddi, S.; Zhao, Y.; Xia, Y.; Claud, E.C.; Sun, J. Salmonella type III effector AvrA stabilizes cell tight junctions to inhibit inflammation in intestinal epithelial cells. PLoS ONE 2008, 3, e2369. [Google Scholar] [CrossRef]
- Zhang, Y.G.; Wu, S.; Xia, Y.; Sun, J. Salmonella-infected crypt-derived intestinal organoid culture system for host-bacterial interactions. Physiol. Rep. 2014, 2, e12147. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, I.; Lu, R.; Zhang, Y.; Zhang, J.; Dai, Y.; Xia, Y.; Sun, J. Vitamin D receptor promotes healthy microbial metabolites and microbiome. Sci. Rep. 2020, 10, 7340. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, Y.; Xia, Y.; Sun, J. Imbalance of the intestinal virome and altered viral-bacterial interactions caused by a conditional deletion of the vitamin D receptor. Gut Microbes 2021, 13, 1957408. [Google Scholar] [CrossRef]
- Zhang, J.; Garrett, S.; Sun, J. Gastrointestinal symptoms, pathophysiology, and treatment in COVID-19. Genes Dis. 2021, 8, 385–400. [Google Scholar] [CrossRef]
- Yamshchikov, A.V.; Desai, N.S.; Blumberg, H.M.; Ziegler, T.R.; Tangpricha, V. Vitamin D for Treatment and Prevention of Infectious Diseases; A Systematic Review of Randomized Controlled Trials. Endocr. Pract. 2009, 15, 438–449. [Google Scholar] [CrossRef] [Green Version]
- Akbar, M.R.; Wibowo, A.; Pranata, R.; Setiabudiawan, B. Low serum 25-hydroxyvitamin D (vitamin D) level is associated with susceptibility to COVID-19, severity, and mortality: A systematic review and meta-analysis. Front. Nutr. 2021, 8, 131. [Google Scholar] [CrossRef]
- Al-Sadi, R.; Boivin, M.; Ma, T. Mechanism of cytokine modulation of epithelial tight junction barrier. Front. Biosci. A J. Virtual Libr. 2009, 14, 2765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, T.; Yoshinaga, N.; Tanabe, S. Interleukin-6 (IL-6) regulates claudin-2 expression and tight junction permeability in intestinal epithelium. J. Biol. Chem. 2011, 286, 31263–31271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, S.; Liao, A.P.; Xia, Y.; Li, Y.C.; Li, J.D.; Sartor, R.B.; Sun, J. Vitamin D receptor negatively regulates bacterial-stimulated NF-kappaB activity in intestine. Am. J. Pathol. 2010, 177, 686–697. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Xia, Y.; Liu, X.; Sun, J. Vitamin D receptor deletion leads to reduced level of IkappaBalpha protein through protein translation, protein-protein interaction, and post-translational modification. Int. J. Biochem. Cell Biol. 2010, 42, 329–336. [Google Scholar] [CrossRef] [Green Version]
- Nakajima, A.; Vogelzang, A.; Maruya, M.; Miyajima, M.; Murata, M.; Son, A.; Kuwahara, T.; Tsuruyama, T.; Yamada, S.; Matsuura, M.; et al. IgA regulates the composition and metabolic function of gut microbiota by promoting symbiosis between bacteria. J. Exp. Med. 2018, 215, 2019–2034. [Google Scholar] [CrossRef] [Green Version]
- Akhtar, S.; Wang, X.; Bu, H.-F.; Tan, X.-D. Role of MFG-E8 in protection of intestinal epithelial barrier function and attenuation of intestinal inflammation. In MFG-E8 and Inflammation; Springer: Berlin/Heidelberg, Germany, 2014; pp. 55–63. [Google Scholar]
- Bakke, D.; Chatterjee, I.; Agrawal, A.; Dai, Y.; Sun, J. Regulation of Microbiota by Vitamin D Receptor: A Nuclear Weapon in Metabolic Diseases. Nucl. Recept. Res. 2018, 5, 101377. [Google Scholar] [CrossRef] [Green Version]
- Yoon, S.S.; Sun, J. Probiotics, nuclear receptor signaling, and anti-inflammatory pathways. Gastroenterol. Res. Pract. 2011, 2011, 971938. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Yoon, S.; Zhang, Y.G.; Lu, R.; Xia, Y.; Wan, J.; Petrof, E.O.; Claud, E.C.; Chen, D.; Sun, J. Vitamin D receptor pathway is required for probiotic protection in colitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 309, G341–G349. [Google Scholar] [CrossRef] [Green Version]
- Shang, M.; Sun, J. Vitamin D/VDR, Probiotics, and Gastrointestinal Diseases. Curr. Med. Chem. 2017, 24, 876–887. [Google Scholar] [CrossRef]
- Lu, R.; Shang, M.; Zhang, Y.G.; Jiao, Y.; Xia, Y.; Garrett, S.; Bakke, D.; Bauerl, C.; Martinez, G.P.; Kim, C.H.; et al. Lactic Acid Bacteria Isolated From Korean Kimchi Activate the Vitamin D Receptor-autophagy Signaling Pathways. Inflamm. Bowel Dis. 2020, 26, 1199–1211. [Google Scholar] [CrossRef]
- Battistini, C.; Ballan, R.; Herkenhoff, M.E.; Saad, S.M.I.; Sun, J. Vitamin D Modulates Intestinal Microbiota in Inflammatory Bowel Diseases. Int. J. Mol. Sci. 2020, 22, 362. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Huang, L.; Zhang, W.; Zhang, H. Vitamin D/VDR in Acute Kidney Injury: A Potential Therapeutic Target. Curr. Med. Chem. 2021, 28, 3865–3876. [Google Scholar] [CrossRef] [PubMed]
- Bikle, D.; Feingold, K.R.; Anawalt, B.; Boyce, A.; Chrousos, G.; de Herder, W.W.; Dhatariya, K.; Dungan, K.; Hershman, J.M.; Hofland, J.; et al. (Eds.) Vitamin D: Production, Metabolism and Mechanisms of Action; MDText.com, Inc.: South Dartmouth, MA, USA, 2000. Available online: https://www.ncbi.nlm.nih.gov/books/NBK278935/ (accessed on 10 February 2022).
- Bikle, D.; Adams, J.; Christakos, S. Vitamin D: Production, metabolism, mechanism of action, and clinical requirements. In Primer on the Metabolic Bone Diseases and Disorders of Mineral Metabolism; Wiley: Hoboken, NJ, USA, 2013; pp. 235–247. [Google Scholar]
- Sherman, M.H.; Yu, R.T.; Engle, D.D.; Ding, N.; Atkins, A.R.; Tiriac, H.; Collisson, E.A.; Connor, F.; Van Dyke, T.; Kozlov, S.; et al. Vitamin D receptor-mediated stromal reprogramming suppresses pancreatitis and enhances pancreatic cancer therapy. Cell 2014, 159, 80–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirano, T.; Kobayashi, N.; Itoh, T.; Takasuga, A.; Nakamaru, T.; Hirotsune, S.; Sugimoto, Y. Null mutation of PCLN-1/Claudin-16 results in bovine chronic interstitial nephritis. Genome Res. 2000, 10, 659–663. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Brown, P.C.; Chow, E.C.Y.; Ewart, L.; Ferguson, S.S.; Fitzpatrick, S.; Freedman, B.S.; Guo, G.L.; Hedrich, W.; Heyward, S.; et al. 3D cell culture models: Drug pharmacokinetics, safety assessment, and regulatory consideration. Clin. Transl. Sci. 2021, 14, 1659–1680. [Google Scholar] [CrossRef]
- Lu, R.; Zhang, Y.G.; Xia, Y.; Zhang, J.; Kaser, A.; Blumberg, R.; Sun, J. Paneth Cell Alertness to Pathogens Maintained by Vitamin D Receptors. Gastroenterology 2021, 160, 1269–1283. [Google Scholar] [CrossRef]
- Lu, R.; Zhang, Y.G.; Xia, Y.; Sun, J. Imbalance of autophagy and apoptosis in intestinal epithelium lacking the vitamin D receptor. FASEB J. 2019, 33, 11845–11856. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, T. Recent progress in organoid culture to model intestinal epithelial barrier functions. Int. Immunol. 2019, 31, 13–21. [Google Scholar] [CrossRef] [Green Version]
- Altay, G.; Larrañaga, E.; Tosi, S.; Barriga, F.M.; Batlle, E.; Fernández-Majada, V.; Martínez, E. Self-organized intestinal epithelial monolayers in crypt and villus-like domains show effective barrier function. Sci. Rep. 2019, 9, 10140. [Google Scholar] [CrossRef]
- Schoultz, I.; Keita, A.V. The Intestinal Barrier and Current Techniques for the Assessment of Gut Permeability. Cells 2020, 9, 1909. [Google Scholar] [CrossRef]
- Lu, R.; Wu, S.; Xia, Y.; Sun, J. The vitamin D receptor, inflammatory bowel diseases, and colon cancer. Curr. Colorectal Cancer Rep. 2012, 8, 57–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tajik, N.; Frech, M.; Schulz, O.; Schalter, F.; Lucas, S.; Azizov, V.; Durholz, K.; Steffen, F.; Omata, Y.; Rings, A.; et al. Targeting zonulin and intestinal epithelial barrier function to prevent onset of arthritis. Nat. Commun. 2020, 11, 1995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Location of claudin-2 and claudin-7 in normal mouse colon. (a) Normal claudin-2 distribution in cell membranes at the crypt base in the mouse colon (C57BL6/J mice, 6–8 weeks) (left panel: indicated by yellow arrows). (b) Normal claudin-7 distribution in cell membrane along crypt–villus axis in the mouse colon (right panel: indicated by yellow arrows) [28].
Figure 1.
Location of claudin-2 and claudin-7 in normal mouse colon. (a) Normal claudin-2 distribution in cell membranes at the crypt base in the mouse colon (C57BL6/J mice, 6–8 weeks) (left panel: indicated by yellow arrows). (b) Normal claudin-7 distribution in cell membrane along crypt–villus axis in the mouse colon (right panel: indicated by yellow arrows) [28].

Figure 2.
Disruption of tight junctions in mouse lung with LPS treatment. (a) Normal ZO-1 distribution in healthy mouse lungs (left panel: yellow arrow). (b) Reduced expression and disorganized structure of ZO-1 (right panel: yellow arrow) in the lungs of mice (C57BL6/J, 6–8 weeks) treated with LPS (10 mg/kg, intranasally). Mice were sacrificed after 24 h-infection [65].
Figure 2.
Disruption of tight junctions in mouse lung with LPS treatment. (a) Normal ZO-1 distribution in healthy mouse lungs (left panel: yellow arrow). (b) Reduced expression and disorganized structure of ZO-1 (right panel: yellow arrow) in the lungs of mice (C57BL6/J, 6–8 weeks) treated with LPS (10 mg/kg, intranasally). Mice were sacrificed after 24 h-infection [65].

Figure 3.
ZO-1 distribution before and after Salmonella infection in mouse colon. (a) ZO-1 distribution in the colon of VDRloxp colon and VDRΔIEC mice. The yellow arrows indicate the “zipper” structure of ZO-1 in the apical side of the colon, and red arrows show the disrupted ZO-1 due to the intestinal epithelial VDR deletion in VDRΔIEC mice. (b) Disrupted ZO-1 distribution after Salmonella infection in the colon of VDRloxp and VDRΔIEC mice. VDRloxp and VDRΔIEC mice aged 6–8 weeks were infected with Salmonella and sacrificed 8 h post-infection, as described in our previous study [27,28].
Figure 3.
ZO-1 distribution before and after Salmonella infection in mouse colon. (a) ZO-1 distribution in the colon of VDRloxp colon and VDRΔIEC mice. The yellow arrows indicate the “zipper” structure of ZO-1 in the apical side of the colon, and red arrows show the disrupted ZO-1 due to the intestinal epithelial VDR deletion in VDRΔIEC mice. (b) Disrupted ZO-1 distribution after Salmonella infection in the colon of VDRloxp and VDRΔIEC mice. VDRloxp and VDRΔIEC mice aged 6–8 weeks were infected with Salmonella and sacrificed 8 h post-infection, as described in our previous study [27,28].

Figure 4.
Distribution of claudin-7 in organoids. (a) Claudin-7 distribution in the cell membranes in the mouse organoids in fluorescent setting (left panel, yellow arrows). (b) Mouse organoids in the bright field. Experimental conditions are described in our previous report [145].
Figure 4.
Distribution of claudin-7 in organoids. (a) Claudin-7 distribution in the cell membranes in the mouse organoids in fluorescent setting (left panel, yellow arrows). (b) Mouse organoids in the bright field. Experimental conditions are described in our previous report [145].


{kind=link}
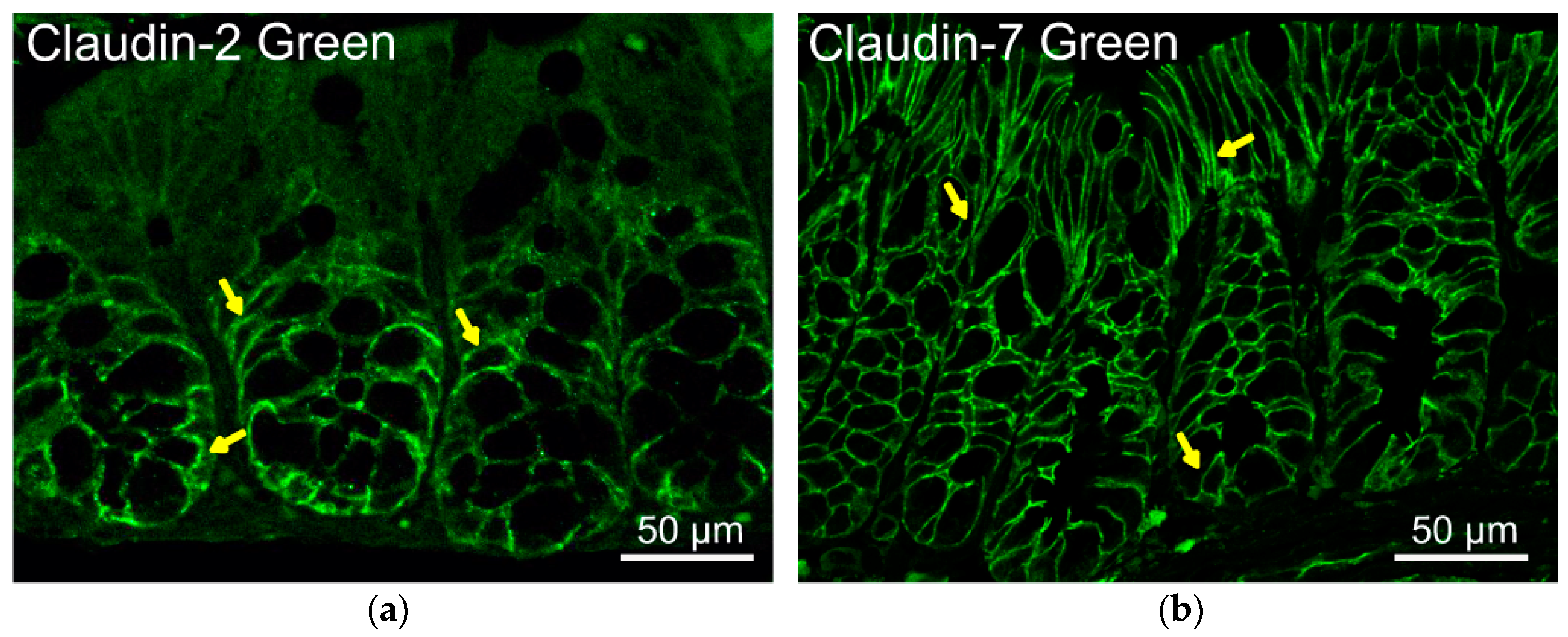{kind=link}
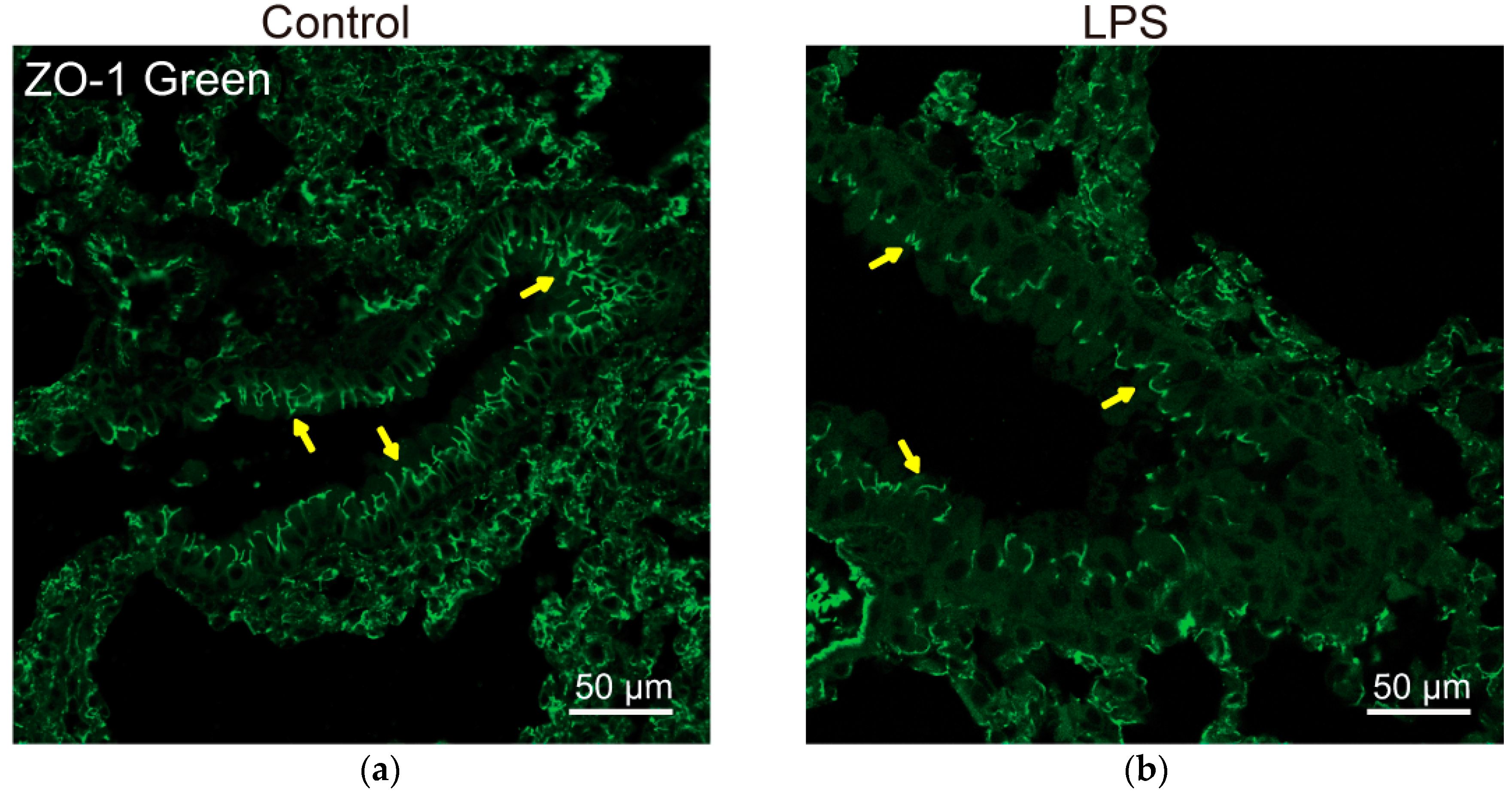{kind=link}
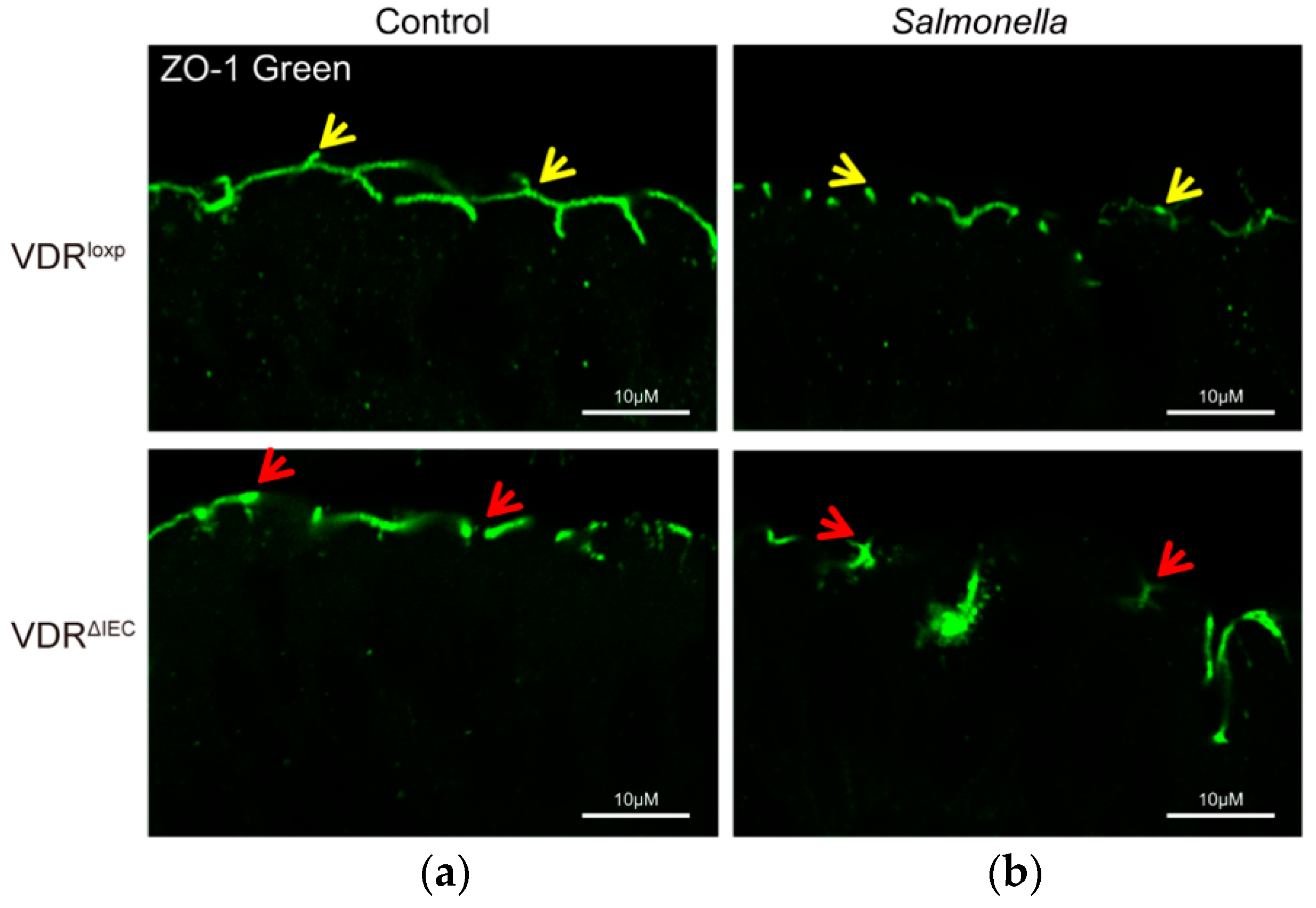{kind=link}
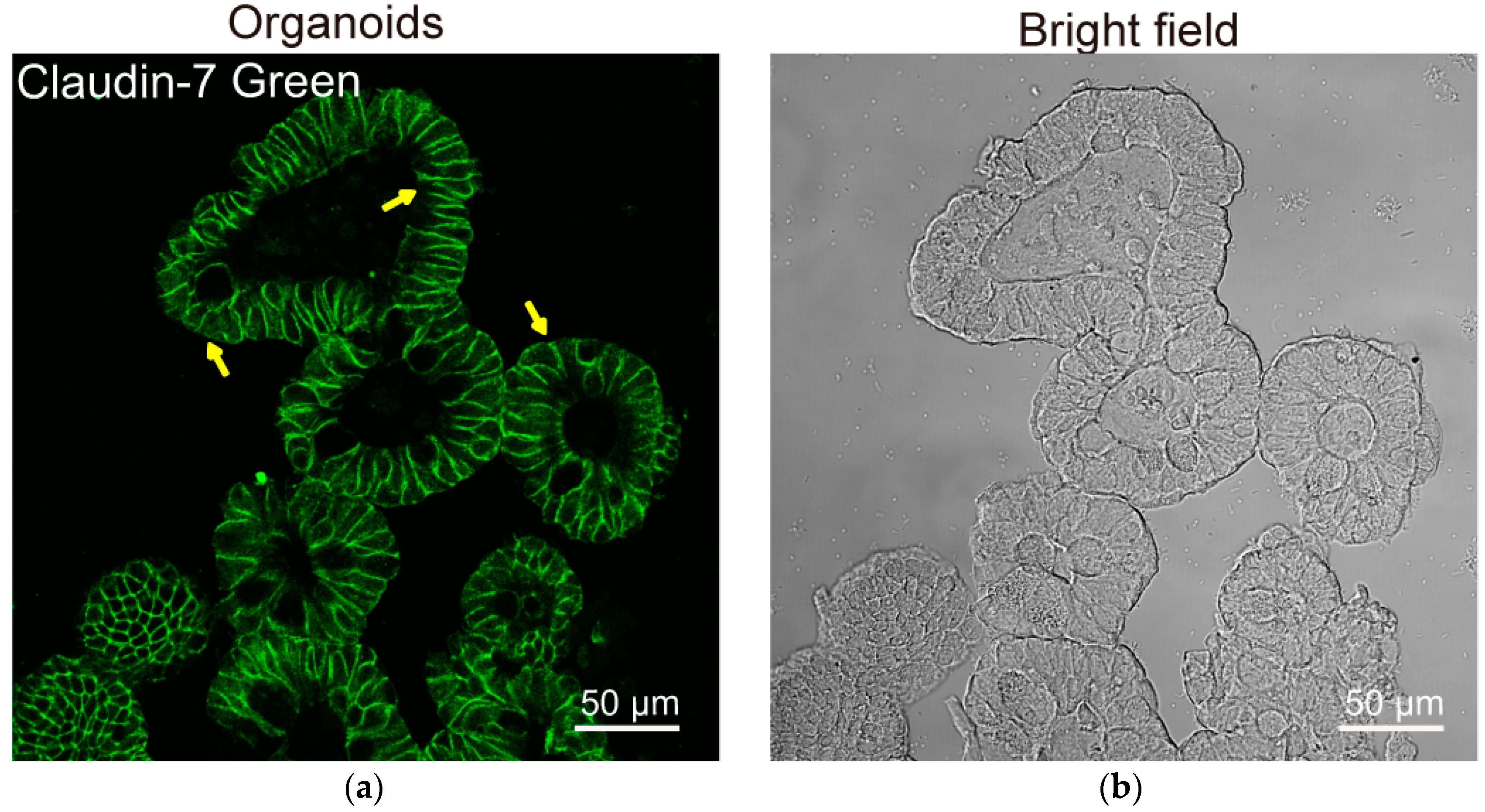{kind=link}
Table 1.
Vitamin D and VDR influence the expression and function of cell junction proteins.
| Tissues & Cell Types | TJ Proteins | Cell Cultures In Vitro | Experimental Models In Vivo | Conclusions | References |
|---|---|---|---|---|---|
| Intestine | ZO-1 E-cadherin β-catenin | T84 | The expression of VDR was positively correlated with the expression of ZO-1, occluding and claudin-5 in T84 cells. | [50] | |
| DLD-1 | VDR acts as a regulator in the expression of intestinal mucosal barrier proteins under hypoxia environment. The expressions of VDR, ZO-1, occludin, claudin-1, and E-cadherin were obviously higher in vitamin D plus hypoxia group than in single vitamin D treatment group. | [51] | |||
| SW480 | 1,25(OH)2D3 induced the expression of adhesion proteins and promoted the translocation of nuclear beta-catenin and ZO-1 to the plasma membrane. | [52] | |||
| Occludin | Cirrhotic rats | Vitamin D3 treatment significantly attenuated bacterial translocation and reduced intestinal permeability in thioacetamide-induced cirrhotic rats. It upregulated the expressions of occludin in the small intestine and claudin-1 in the colon of cirrhotic rats directly independent of intrahepatic status. Vitamin D3 treatment also enriched Muribaculaceae, Bacteroidales, Allobaculum, Anaerovorax, and Ruminococcaceae. | [53] | ||
| SW480-ADH | ROCK and MSK inhibition abrogates the induction of 1,25(OH)2D3 24-hydroxylase (CYP24), E-cadherin, and vinculin, and the repression of cyclin D1 by 1,25(OH)2D3. | [54] | |||
| Caco-2 | VDR−/− Mice | 1,25(OH)2D3 enhanced TJs by increasing junction protein expression and TER and preserved the structural integrity of TJs in the presence of DSS. VDR knockdown reduced the junction proteins and TER. 1,25(OH)2D3 can stimulate epithelial cell migration in vitro. | [43] | ||
| Claudins | Caco-2 SKCO15 | VDR−/− VDRΔIEC mice | 1,25(OH)2D3 treatment upregulates claudin-2 expression in human epithelium cells. VDR deletion in intestinal epithelial cells led to significant decreased claudin-2 expression. CLDN2 gene is a direct target of the transcription factor VDR. VDR enhances claudin-2 promoter activity in a Cdx1 binding, site-dependent manner. | [2] | |
| Enteroids Caco-2 SKCO15 | VDR−/− Mice Salmonella- or DSS colitis model | In inflamed intestines of Salmonella- or DSS-induced colitis model, a lack of VDR regulation led to a robust increase of claudin-2 at the mRNA and protein levels post-infection. In inflamed intestines of ulcerative colitis patients, VDR expression was low and claudin-2 was enhanced. In inflamed VDR−/− cells, 1,25(OH)2D3 enhanced claudin-2 promoter activity through the binding sites of NF-κB and STAT. | [27] | ||
| SKCO15 | O-VDR VDR∆IEC-Mice | Reduced claudin-15 was significantly correlated with decreased VDR in human IBD. O-VDR mice showed decreased susceptibility to chemically and bacterially induced colitis and marked increased claudin-15 expression in the colon. Correspondingly, colonic claudin-15 was reduced in VDR∆IEC mice, which were susceptible to colitis. Overexpression of intestinal epithelial VDR and vitamin D treatment resulted in significantly increased claudin-15. ChIP assays identified claudin-15 gene as a direct target of VDR. | [3] | ||
| Human Colonoids SKCO15 | VDR−/− VDRΔIEC-mice | Colonic VDR expression was low and significantly correlated with a reduction in claudin-5 in human CRC patients. Lack of VDR and a reduction of claudin-5 are associated with an increased number of tumors in the VDR−/− and VDRΔIEC mice. CHIP assay identified CLDN-5 as a downstream target of the VDR signaling pathway. | [4] | ||
| Human tissue microarr-ays | Vitamin D receptor (VDR) enhanced claudin-2 expression in colon and bile salt receptors VDR and Takeda G-protein coupled receptor 5 (TGR5) were highly expressed in esophageal adenocarcinoma and pre-cancerous lesions. | [23] | |||
| 1,25(OH)2D3 induces RANKL, SPP1 (osteopontin), and BGP (osteocalcin) to govern bone mineral remodeling; TRPV6, CaBP(9k), and claudin-2 to promote intestinal calcium absorption; and TRPV5, klotho, and Npt2c to regulate renal calcium and phosphate reabsorption. | [55] | ||||
| SW480-ADH | 1,25(OH)2D3 activates the JMJD3 gene promoter and increases JMJD3 RNA in human cancer cells. JMJD3 knockdown or expression of an inactive mutant JMJD3 fragment decreased the induction by 1,25(OH)2D3 of several target genes and of an epithelial adhesive phenotype. It downregulated E-cadherin, claudin-1, and claudin-7. | [56] | |||
| calbindin-D9k−/− mutant mice | 1,25(OH)2D3 downregulates cadherin-17 and upregulates claudin-2 and claudin-12 in the intestine, suggesting that 1,25(OH)2D3 can route calcium through the paracellular path by regulating the epithelial cell junction proteins. | [57] | |||
| Caco-2 | VDR−/− mice | Claudin-2 and/or claudin-12-based TJs form paracellular Ca(2+) channels in intestinal epithelia. This study highlights a vitamin D-dependent mechanism in calcium homeostasis. | [1] | ||
| Barrier and mucosal immunity | Rat intestine | The most strongly affected gene in intestine was CYP24 with 97-fold increase at 6 h post-1,25(OH)2D3 treatment. Intestinal calcium absorption genes: TRPV5, TRPV6, calbindin D(9k), and Ca(2+) dependent ATPase all were upregulated in response to 1,25(OH)2D3, However, a 1,25-(OH)2D3 suppression of several intra- and intercellular matrix modeling proteins, such as sodium/potassium ATPase, claudin-3, aquaporin 8, cadherin 17, and RhoA, suggesting a vitamin D regulation of TJ permeability and paracellular calcium transport. Expression of several other genes related to the immune system and angiogenesis was changed in response to 1,25(OH)2D3. | [58] | ||
| Caco-2 | C57BL/6 mice | 1,25(OH)2D3 pre-treatment ameliorated the ethanol-induced barrier dysfunction, TJ disruption, phosphorylation level of MLC, and generation of ROS, compared with ethanol-exposed monolayers. Mice fed with vitamin D-sufficient diet had a higher plasma level of 25(OH)D3 and were more resistant to ethanol-induced acute intestinal barrier injury compared with the vitamin D-deficient group. | [59] | ||
| VDRΔIEC VDRΔCEC Mice TNBS- colitis model | Gut epithelial VDR deletion aggravates epithelial cell apoptosis, resulting in mucosal barrier permeability increases. | [60] | |||
| Caco-2 | DSS-colitis model | 1,25(OH)2D3 plays a protective role in mucosal barrier homeostasis by maintaining the integrity of junction complexes and in healing capacity of the colon epithelium. | [61] | ||
| Lung | E-cadherin β-catenin | A459 | Vitamin D pre-treatment reduced TGF-β and Wnt/β-catenin signaling by increasing p-VDR, protected from E-cadherin degradation and led to the regression of EMT (epithelial–mesenchymal transition)-mediated myofibroblast differentiation. | [62] | |
| 16HBE | Vitamin D is able to counteract the cigarette smoke extract-induced bronchial epithelial barrier disruption by TER (transepithelial electrical resistance) reduction inhibition, permeability increase, ERK phosphorylation increase, calpain-1 expression increase, and distribution anomalies and the cleavage of E-cadherin and β-catenin. | [63] | |||
| ZO-1 Occludin | VDR−/− WTMice | Vitamin D supplementation alleviated LPS-induced lung injury and preserved alveolar barrier function through maintenance of the pulmonary barrier by inducing expression of occludin and ZO-1 in whole lung homogenates. | [64] | ||
| Claudins | VDR−/− WT Mice | VDR−/− mice showed significantly decreased ZO-1, occludin, claudin-1, claudin-2, claudin-4, claudin-10, β-catenin, and VE-cadherin expression in the lungs tissue compared with WT mice. | [65] | ||
| Kidney | ZO-1 | Rat | Rat treated with 1,25(OH)2D3 is able to abrogate podocytes injury, detected as desmin expression and loss of nephrin and ZO-1. | [66] | |
| Claudins | HEK 293 OK | ICR mice | In kidney, 1,25(OH)2 VitD transcriptionally inhibits claudin-16 expression by a mechanism sensitive to CaSR and Mg2+. This renal effect of 1,25(OH)2 vitamin D may serve as an adaptive response to the 1,25(OH)2 vitamin D-induced increase in intestinal Mg2+ absorption. | [67] | |
| Cornea | ZO-1 Occludin | VDR−/− WT Mice | VDR−/− mice showed the decreased expression of ZO-1 and occludin, and changed ZO-distribution on corneas compared with WT mice. | [68] | |
| Corneal epithelium | Mouse, Rabbit, Human | Vitamin D enhances corneal epithelial barrier function. Cells showed increased TER, decreased IP, and increased occludin levels when cultured with 25(OH)D3 and 1,25(OH)2D3. | [69] | ||
| Skin | ZO-1 Claudins | Human Psoriatic and Normal Skin | Psoriatic skin reduced VDR, ZO-1, and claudin-1 expression compared to normal skin, and showed a significant correlation of downregulated VDR expression to claudin-1 and ZO-1. | [70] | |
| Brain | ZO-1 Occludin Claudins | End.3 | Following hypoxic injury, 1,25(OH)2D3 treatment prevented decreased barrier function, and expression of zonula occludin-1, claudin-5, and occludin. VDR mediated the protective effect of 1,25(OH)2D3 against ischemic injury-induced blood–brain barrier dysfunction in cerebral endothelial cells. | [71] | |
| Oral | ZO-1 E-cadherin β-catenin | HOK-16B | Vitamin D reinforces E-cadherin junctions by downregulating NF-κB signaling. In addition, vitamin D averts TNF-α-induced downregulation of the development of E-cadherin junctions in HGKs by decreasing the production of MMP-9, which was upregulated by TNF-α. | [72] | |
| Urinary bladder | Occludin Claudin-14 | Mouse, Human | During E. coli infection, vitamin D induced occludin and claudin-14 in mature superficial umbrella cells of the urinary bladder. Vitamin D increased cell–cell adhesion, thus consolidating the epithelial integrity during infection. | [73] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Sun, J.; Zhang, Y.-G. Vitamin D Receptor Influences Intestinal Barriers in Health and Disease. Cells 2022, 11, 1129. https://doi.org/10.3390/cells11071129
AMA Style
Sun J, Zhang Y-G. Vitamin D Receptor Influences Intestinal Barriers in Health and Disease. Cells. 2022; 11(7):1129. https://doi.org/10.3390/cells11071129
Chicago/Turabian StyleSun, Jun, and Yong-Guo Zhang. 2022. "Vitamin D Receptor Influences Intestinal Barriers in Health and Disease" Cells 11, no. 7: 1129. https://doi.org/10.3390/cells11071129
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.