
Leukocyte Telomere Length Correlates with Extended Female Fertility

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design and Participants
2.2. Leukocyte Genomic DNA Preparation
2.3. Telomere Length Measurements
2.4. Statistical Analysis
3. Results
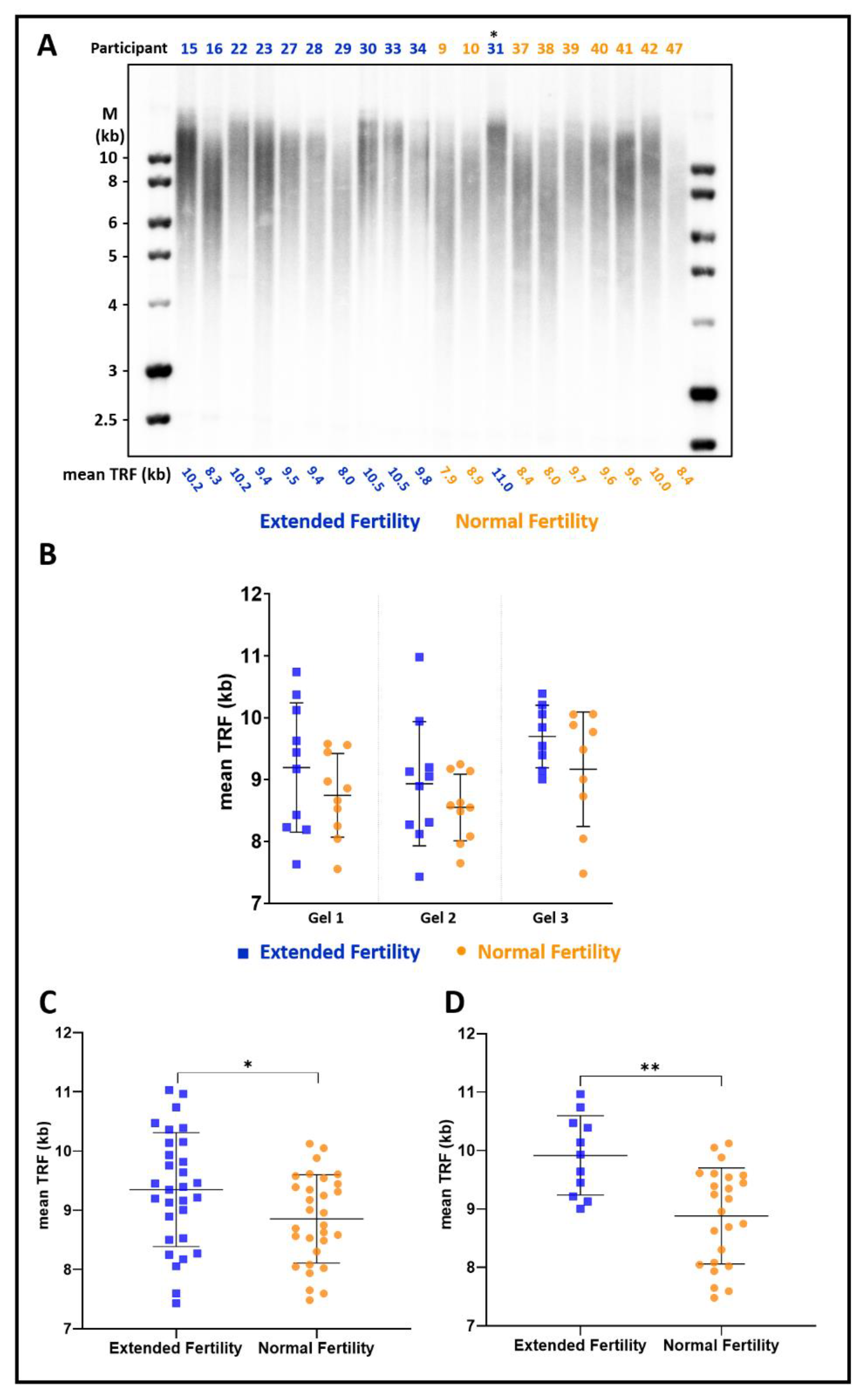
3.1. Women with Extended Fertility Have Longer Telomeres
3.2. The Difference in Telomere Length Is More Pronounced in Women with Up to Eight Children
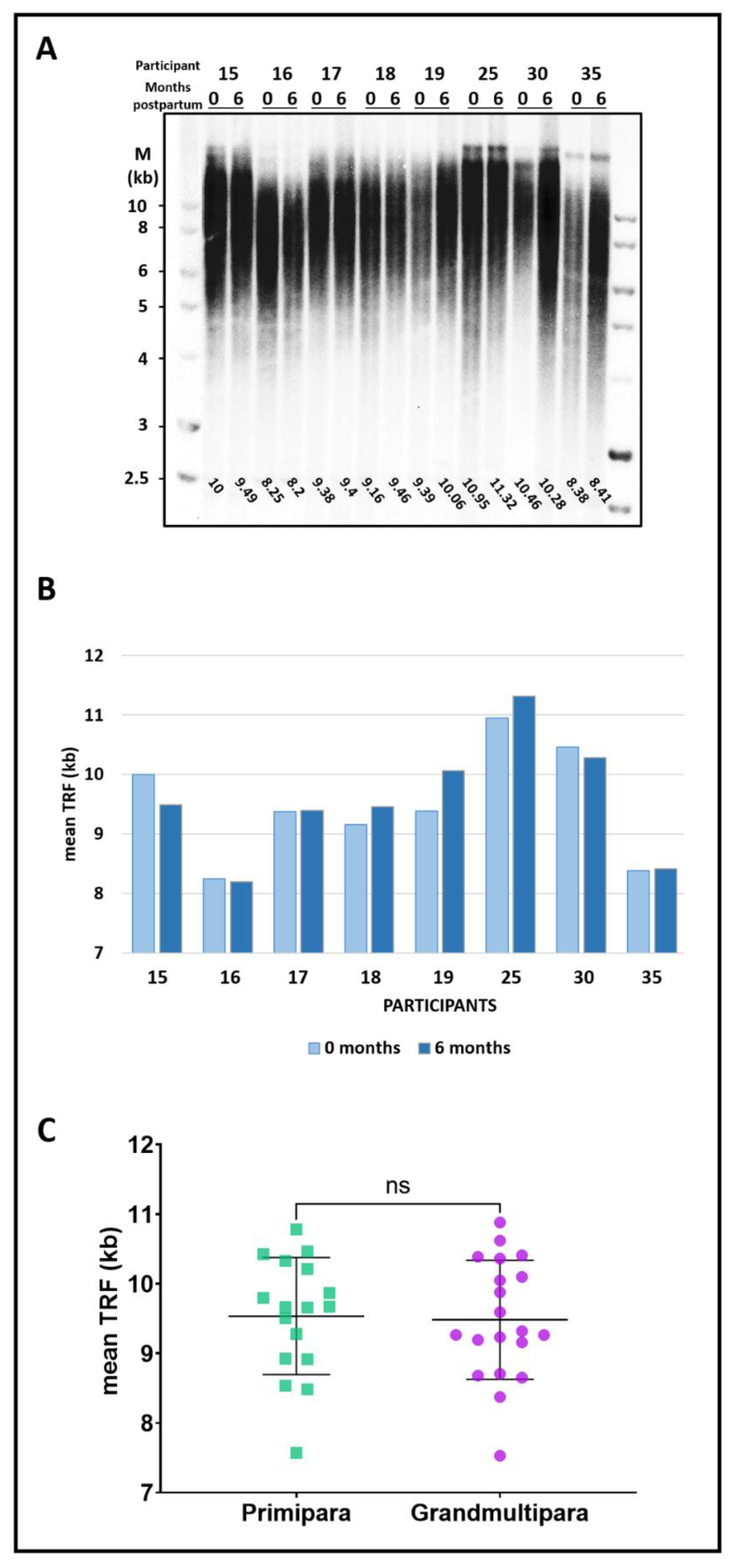
3.3. Pregnancy, Delivery, and Parity Do Not Affect Telomere Length
4. Discussion
4.1. Women with Unusual Extended Fertility Have Longer Telomeres
4.2. Parity and Delivery Do Not Affect Average Telomere Length
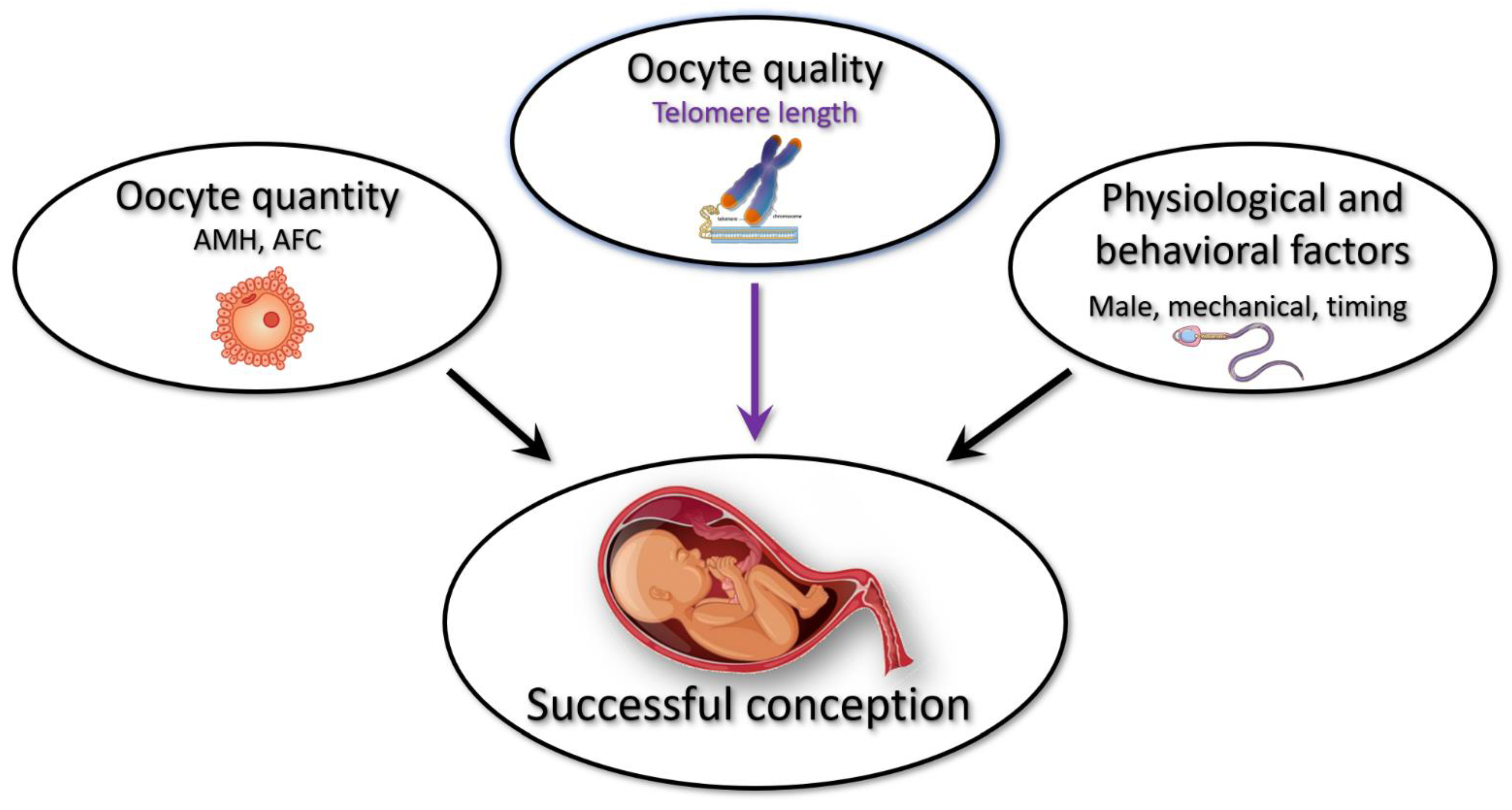
4.3. Telomere Length as a Biomarker for Oocyte Quality
4.4. A Working Model for Extended Fertility
4.5. Do Telomeres Play a Direct Role in Extended Fertility?
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Practice Committee of the American Society for Reproductive Medicine in collaboration with the Society for Reproductive Endocrinology and Infertility. Optimizing natural fertility: A committee opinion. Fertil. Steril. 2017, 107, 52–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- American College of Obstetricians and Gynecologists. ACOG Committee Opinion. Female Age-Related Fertility Decline. Fertil. Steril. 2014, 101, 633–634. [Google Scholar] [CrossRef] [PubMed]
- Szewczuk, E. Age-related infertility: A tale of two technologies. Sociol. Health Illn. 2012, 34, 429–443. [Google Scholar] [CrossRef]
- Battaglia, D.E.; Goodwin, P.; Klein, N.A.; Soules, M.R. Fertilization and early embryology: Influence of maternal age on meiotic spindle assembly oocytes from naturally cycling women. Hum. Reprod. 1996, 11, 2217–2222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steiner, A.Z.; Pritchard, D.; Stanczyk, F.Z.; Kesner, J.S.; Meadows, J.W.; Herring, A.H.; Baird, D.D. Association Between Biomarkers of Ovarian Reserve and Infertility Among Older Women of Reproductive Age. JAMA 2017, 318, 1367–1376. [Google Scholar] [CrossRef] [PubMed]
- McArdle, P.F.; Pollin, T.I.; O’Connell, J.R.; Sorkin, J.D.; Agarwala, R.; Schäffer, A.A.; Streeten, E.A.; King, T.M.; Shuldiner, A.R.; Mitchell, B.D. Does having children extend life span? A genealogical study of parity and longevity in the Amish. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2006, 61, 190–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaffe, D.; Kogan, L.; Manor, O.; Gielchinsky, Y.; Dior, U.; Laufer, N. Influence of late-age births on maternal longevity. Ann. Epidemiol. 2015, 25, 387–391. [Google Scholar] [CrossRef]
- Smith, K.R.; Gagnon, A.; Cawthon, R.M.; Mineau, G.P.; Mazan, R.; Desjardins, B. Familial aggregation of survival and late female reproduction. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2009, 64, 740–744. [Google Scholar] [CrossRef]
- Gagnon, A. Natural fertility and longevity. Fertil. Steril. 2015, 103, 1109–1116. [Google Scholar] [CrossRef]
- Müller, H.-G.; Chiou, J.-M.; Carey, J.R.; Wang, J.-L. Fertility and life span: Late children enhance female longevity. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2002, 57, B202–B206. [Google Scholar] [CrossRef] [Green Version]
- Blackburn, E.H.; Epel, E.S.; Lin, J. Human telomere biology: A contributory and interactive factor in aging, disease risks, and protection. Science 2015, 350, 1193–1198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Codd, V.; Wang, Q.; Allara, E.; Musicha, C.; Kaptoge, S.; Stoma, S.; Jiang, T.; Hamby, S.E.; Braund, P.S.; Bountziouka, V.; et al. Polygenic basis and biomedical consequences of telomere length variation. Nat. Genet. 2021, 53, 1425–1433. [Google Scholar] [CrossRef]
- Kimura, M.; Cherkas, L.F.; Kato, B.S.; Demissie, S.; Hjelmborg, J.B.; Brimacombe, M.; Cupples, A.; Hunkin, J.L.; Gardner, J.P.; Lu, X.; et al. Offspring’s leukocyte telomere length, paternal age, and telomere elongation in sperm. PLoS Genet. 2008, 4, e37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keefe, D.L.; Marquard, K.; Liu, L. The telomere theory of reproductive senescence in women. Curr. Opin. Obstet. Gynecol. 2006, 18, 280–285. [Google Scholar] [CrossRef]
- Anifandis, G.; Samara, M.; Simopoulou, M.; Messini, C.I.; Chatzimeletiou, K.; Thodou, E.; Daponte, A.; Georgiou, I. Insights into the Role of Telomeres in Human Embryological Parameters. Opinions Regarding IVF. J. Dev. Biol. 2021, 9, 49. [Google Scholar] [CrossRef]
- Awad, A.; Glousker, G.; Lamm, N.; Tawil, S.; Hourvitz, N.; Smoom, R.; Revy, P.; Tzfati, Y. Full length RTEL1 is required for the elongation of the single-stranded telomeric overhang by telomerase. Nucleic Acids Res. 2020, 48, 7239–7251. [Google Scholar] [CrossRef] [PubMed]
- Göhring, J.; Fulcher, N.; Jacak, J.; Riha, K. TeloTool: A new tool for telomere length measurement from terminal restriction fragment analysis with improved probe intensity correction. Nucleic Acids Res. 2014, 42, e21. [Google Scholar] [CrossRef] [Green Version]
- Pollack, A.Z.; Rivers, K.; Ahrens, K.A. Parity associated with telomere length among US reproductive age women. Hum. Reprod. 2018, 33, 736–744. [Google Scholar] [CrossRef] [PubMed]
- Gagnon, A.; Smith, K.R.; Tremblay, M.; Vézina, H.; Paré, P.-P.; Desjardins, B. Is there a trade-off between fertility and longevity? A comparative study of women from three large historical databases accounting for mortality selection. Am. J. Hum. Biol. 2009, 21, 533–540. [Google Scholar] [CrossRef]
- Committee of the American Society for Reproductive Medicine. Testing and interpreting measures of ovarian reserve: A committee opinion. Fertil. Steril. 2020, 114, 1151–1157. [Google Scholar] [CrossRef]
- Iliodromiti, S.; Anderson, R.A.; Nelson, S.M. Technical and performance characteristics of anti-Müllerian hormone and antral follicle count as biomarkers of ovarian response. Hum. Reprod. Update 2015, 21, 698–710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morin, S.J.; Patounakis, G.; Juneau, C.R.; Neal, S.A.; Scott, R.T.; Seli, E. Diminished ovarian reserve and poor response to stimulation in patients <38 years old: A quantitative but not qualitative reduction in performance. Hum. Reprod. 2018, 33, 1489–1498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cobo, A.; García-Velasco, J.A.; Coello, A.; Domingo, J.; Pellicer, A.; Remohí, J. Oocyte vitrification as an efficient option for elective fertility preservation. Fertil. Steril. 2016, 105, 755–764.e8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalmbach, K.H.; Fontes Antunes, D.M.; Dracxler, R.C.; Knier, T.W.; Seth-Smith, M.L.; Wang, F.; Liu, L.; Keefe, D.L. Telomeres and human reproduction. Fertil. Steril. 2013, 99, 23–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reig-Viader, R.; Garcia-Caldés, M.; Ruiz-Herrera, A. Telomere homeostasis in mammalian germ cells: A review. Chromosoma 2016, 125, 337–351. [Google Scholar] [CrossRef]
- Ge, J.; Li, C.; Li, C.; Huang, Z.; Zeng, J.; Han, L.; Wang, Q. SIRT6 participates in the quality control of aged oocytes via modulating telomere function. Aging 2019, 11, 1965–1976. [Google Scholar] [CrossRef]
- Ghosh, S.; Feingold, E.; Chakraborty, S.; Dey, S.K. Telomere length is associated with types of chromosome 21 nondisjunction: A new insight into the maternal age effect on Down syndrome birth. Hum. Genet. 2010, 127, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Gruhn, J.R.; Zielinska, A.P.; Shukla, V.; Blanshard, R.; Capalbo, A.; Cimadomo, D.; Nikiforov, D.; Chan, A.C.H.; Newnham, L.J.; Vogel, I.; et al. Chromosome errors in human eggs shape natural fertility over reproductive life span. Science 2019, 365, 1466–1469. [Google Scholar] [CrossRef] [Green Version]
- Rizzo, M.; Ducheyne, K.D.; Deelen, C.; Beitsma, M.; Cristarella, S.; Quartuccio, M.; Stout, T.A.E.; Ruijter-Villani, M. Advanced mare age impairs the ability of in vitro-matured oocytes to correctly align chromosomes on the metaphase plate. Equine Vet. J. 2019, 51, 252–257. [Google Scholar] [CrossRef]
- Daniali, L.; Benetos, A.; Susser, E.; Kark, J.D.; Labat, C.; Kimura, M.; Desai, K.; Granick, M.; Aviv, A. Telomeres shorten at equivalent rates in somatic tissues of adults. Nat. Commun. 2013, 4, 1597. [Google Scholar] [CrossRef]
- Demanelis, K.; Jasmine, F.; Chen, L.S.; Chernoff, M.; Tong, L.; Delgado, D.; Zhang, C.; Shinkle, J.; Sabarinathan, M.; Lin, H.; et al. Determinants of telomere length across human tissues. Science 2020, 369, eaaz6876. [Google Scholar] [CrossRef] [PubMed]
- Goglin, S.E.; Farzaneh-Far, R.; Epel, E.S.; Lin, J.; Blackburn, E.H.; Whooley, M.A. Change in Leukocyte Telomere Length Predicts Mortality in Patients with Stable Coronary Heart Disease from the Heart and Soul Study. PLoS ONE 2016, 11, e0160748. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Case (n = 30) | Control (n = 30) | |
|---|---|---|
| Age (years), mean ± SD | 44.23 ± 1.40 | 44.9 ± 1.35 |
| Gravida, mean ± SD | 11.63 ± 5.03 | 7.5 ± 4.09 |
| Para, mean ± SD | 9.37 ± 3.83 | 5.7 ± 3.27 |
| Miscarriages, mean ± SD | 2.27 ± 1.93 | 1.77 ± 1.84 |
| Living children, mean ± SD | 9.4 ± 3.89 | 5.83 ± 3.25 |
| Menarche (years), mean ± SD | 13.14 ± 1.68 | 13.28 ± 1.41 |
| Mother’s age at last child (years), mean ± SD | 37.19 ± 5.56 | 36.93 ± 5.58 |
| Sister’s age at last child (years), mean ± SD * | 41.18 ± 2.32 | 38.87 ± 4.14 |
| Primipara | Grandmultipara | |
|---|---|---|
| (n = 17) | (n = 20) | |
| Age (years), mean ± SD | 31.47 ± 1.33 | 32.95 ± 1.24 |
| Gravida, mean ± SD | 1.29 ± 0.75 | 8.30 ± 1.55 |
| Para, mean ± SD | 1 | 7.45 ± 1.32 |
| Miscarriages, mean ± SD | 0.29 ± 0.75 | 0.85 ± 1.11 |
| Living children, mean ± SD | 1 | 7.40 ± 1.36 |
| Menarche (years), mean ± SD | 13.71 ± 1.52 | 12.90 ± 0.85 |
| Mother’s age at last child (years), mean ± SD | 34.69 ± 5.11 | 39.15 ± 4.38 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Michaeli, J.; Smoom, R.; Serruya, N.; El Ayoubi, H.; Rotshenker-Olshinka, K.; Srebnik, N.; Michaeli, O.; Eldar-Geva, T.; Tzfati, Y. Leukocyte Telomere Length Correlates with Extended Female Fertility. Cells 2022, 11, 513. https://doi.org/10.3390/cells11030513
Michaeli J, Smoom R, Serruya N, El Ayoubi H, Rotshenker-Olshinka K, Srebnik N, Michaeli O, Eldar-Geva T, Tzfati Y. Leukocyte Telomere Length Correlates with Extended Female Fertility. Cells. 2022; 11(3):513. https://doi.org/10.3390/cells11030513
Chicago/Turabian StyleMichaeli, Jennia, Riham Smoom, Noa Serruya, Hosniyah El Ayoubi, Keren Rotshenker-Olshinka, Naama Srebnik, Ofir Michaeli, Talia Eldar-Geva, and Yehuda Tzfati. 2022. "Leukocyte Telomere Length Correlates with Extended Female Fertility" Cells 11, no. 3: 513. https://doi.org/10.3390/cells11030513