
Impaired Glucocorticoid Receptor Signaling Aggravates Lung Injury after Hemorrhagic Shock


Abstract
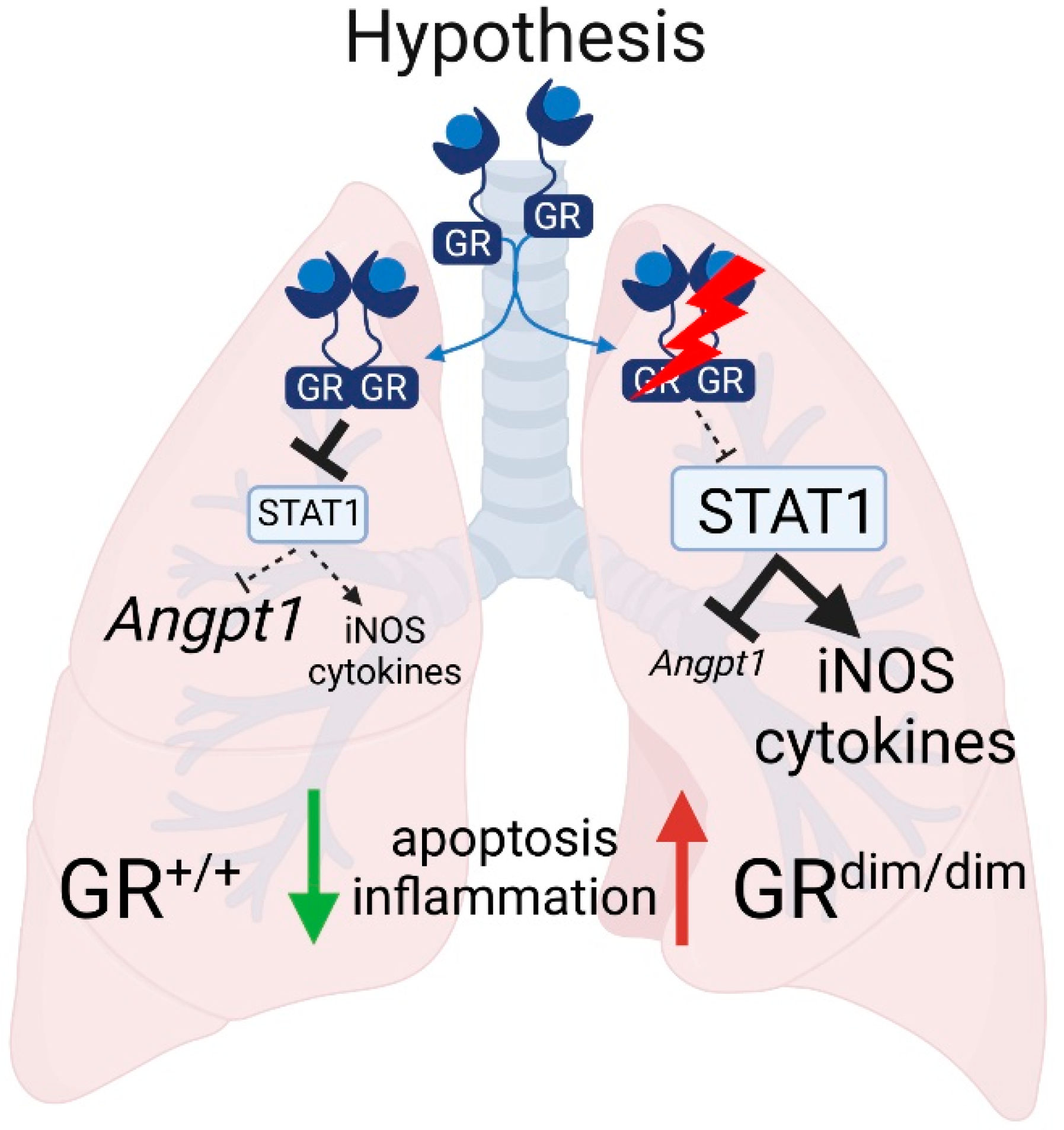
:1. Introduction
2. Materials and Methods
2.1. Animal Housing
2.2. Mouse Intensive Care Unit (MICU)
2.3. Hemorrhagic Shock (HS) Model
2.4. Western Blot
2.5. Bulk RNA-Sequencing of Lung Tissue
2.6. Histology
2.7. Immunohistochemical (IHC) Detection of Lung Albumin Level as Indicator for Vascular Permeability
2.8. Assessment of Relative mRNA Expression with Quantitative real Time Polymerase Chain Reaction (qRT-PCR)
2.9. TUNEL Assay for Apoptosis Measurement
2.10. Bio-Plex Based Measurement of Cytokines and Chemokines
2.11. Statistics and Graph Visualization
3. Results
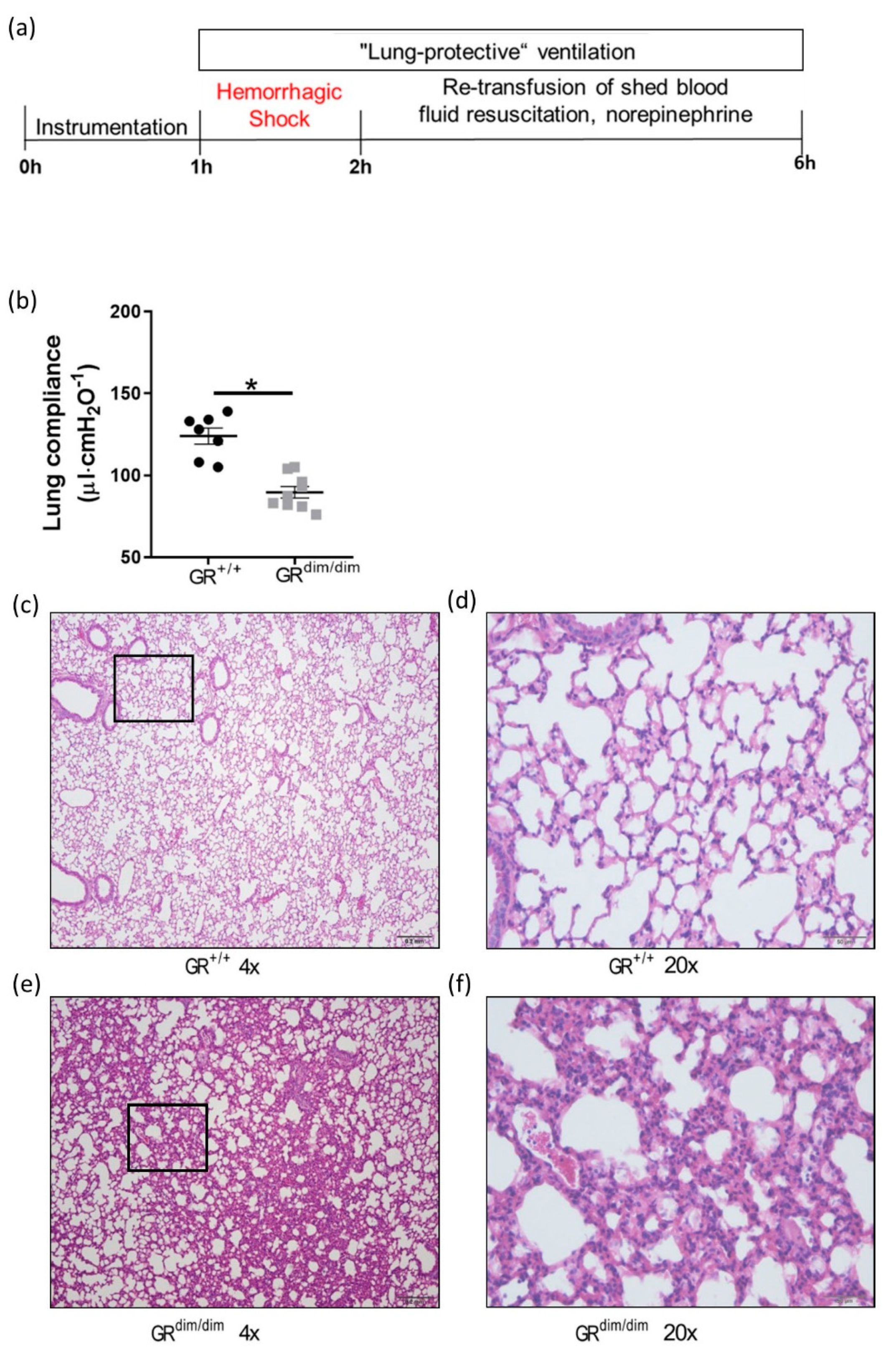
3.1. Functional GR Is Crucial for Lung Mechanics after Hemorrhagic Shock
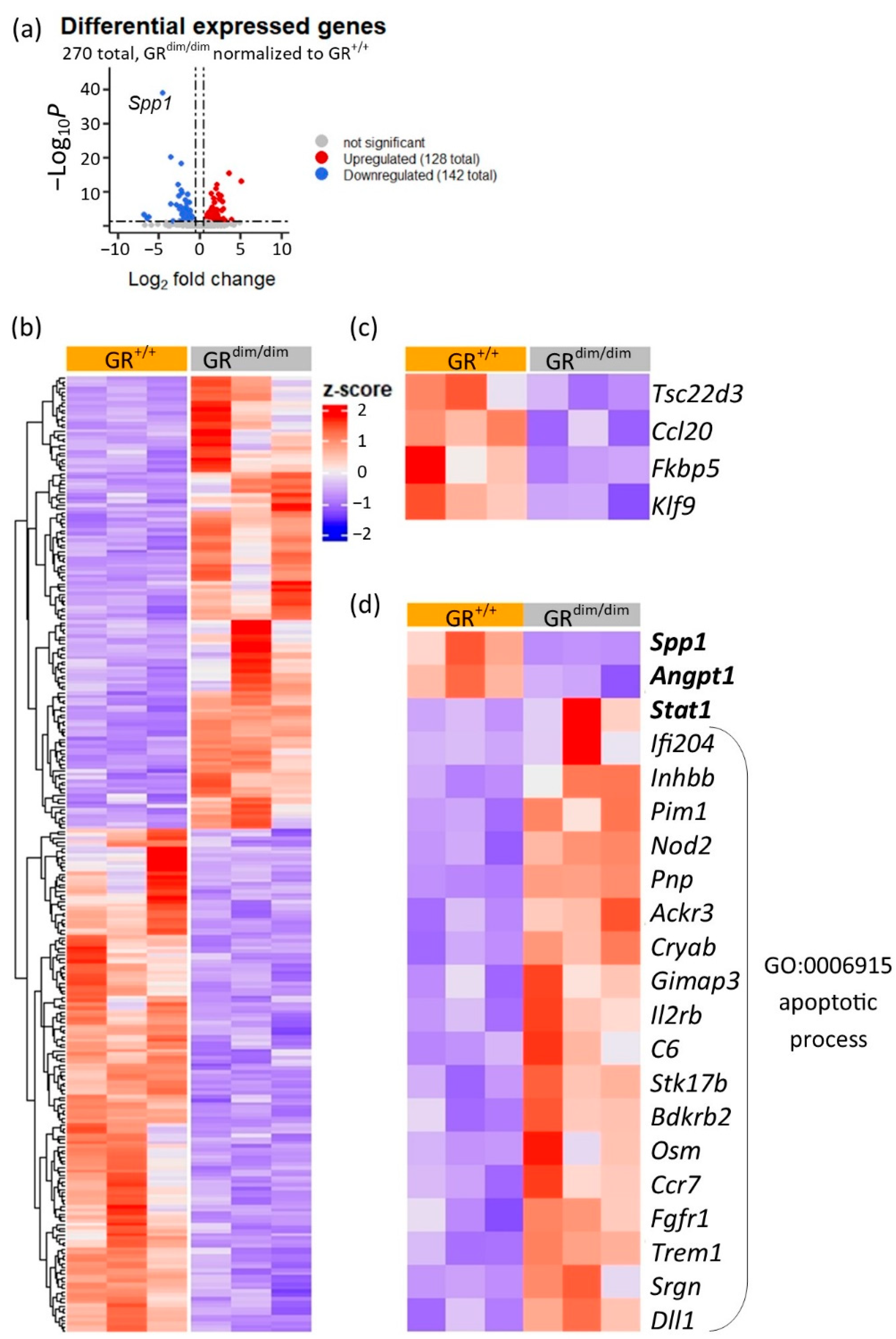
3.2. RNA-Sequencing of Lung Tissue Samples from GRdim/dim and GR+/+ Mice
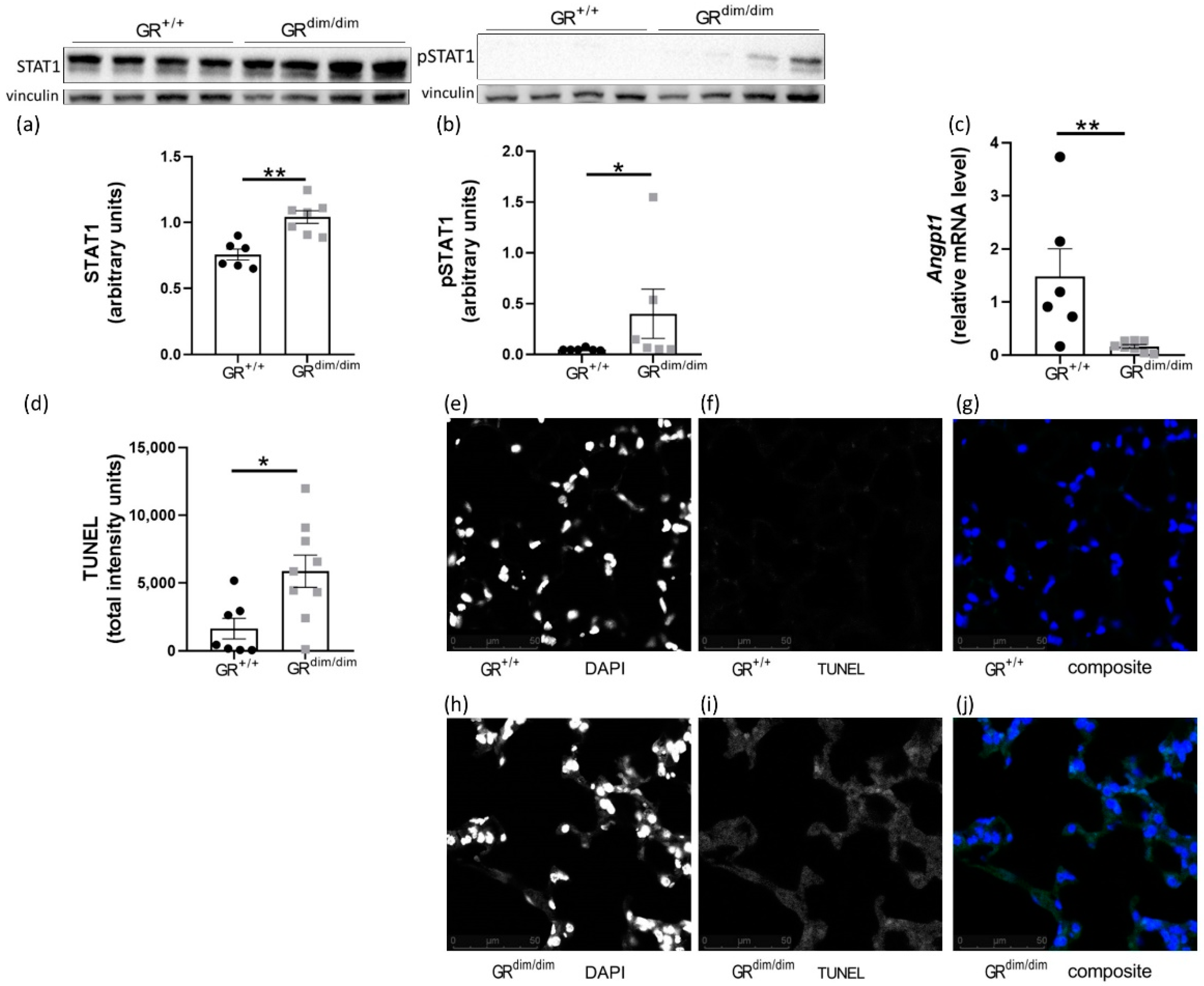
3.3. Enhanced Apoptotic Processes in GRdim/dim after HS
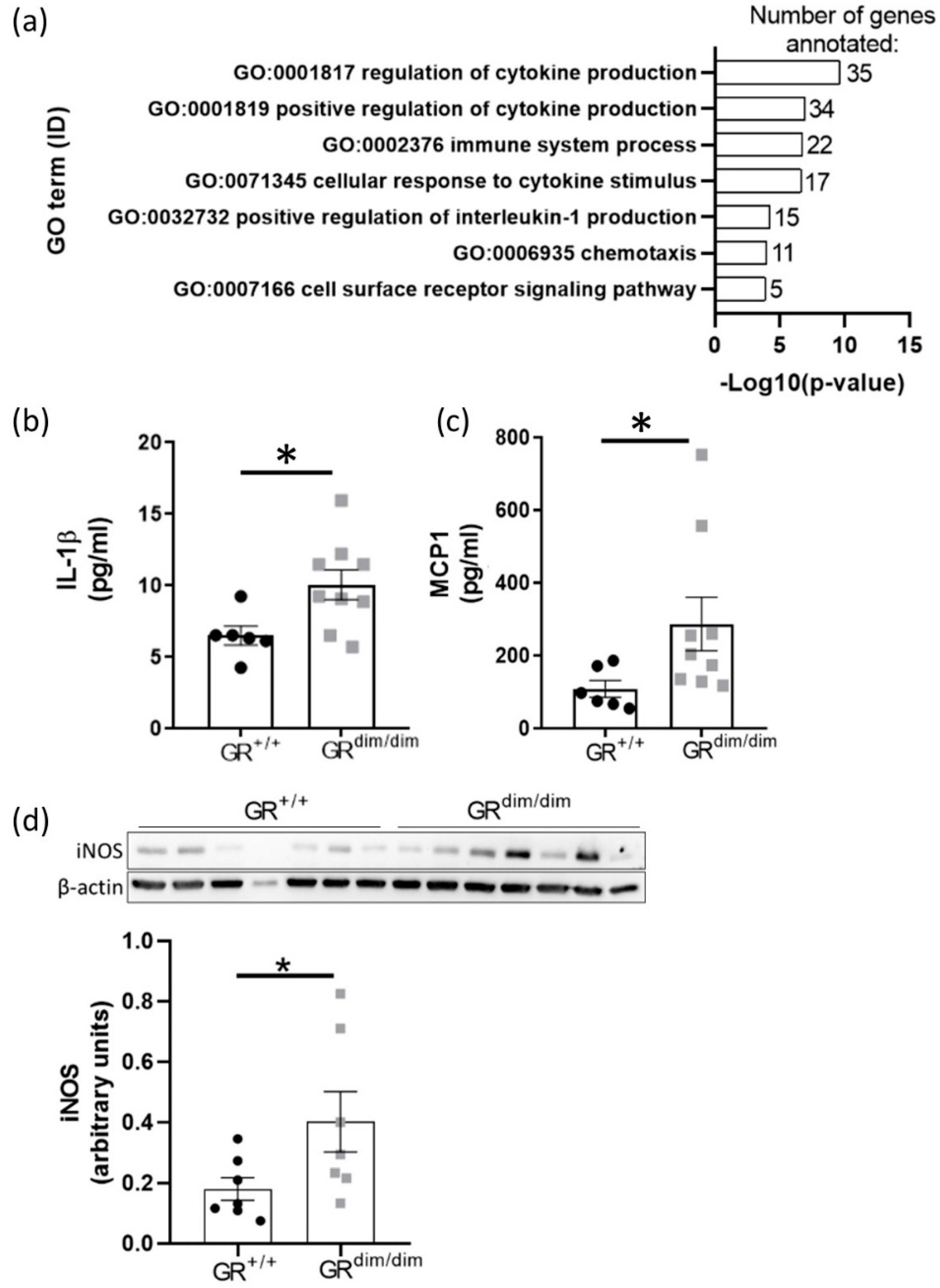
3.4. Enhanced Lung Inflammation in GRdim/dim Mice after HS
4. Discussion
4.1. Functional GR Is Crucial for Lung Health in Resuscitated Shock
4.2. Full Funtional GR Is Necessary for Repression of HS-Induced STAT1/pSTAT1
4.3. Increased Apoptosis in Lungs of GRdim/dim Mice after HS
4.4. Impaired GR Enhances HS-Induced Lung Inflammation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Escoter-Torres, L.; Caratti, G.; Mechtidou, A.; Tuckermann, J.; Uhlenhaut, N.H.; Vettorazzi, S. Fighting the Fire: Mechanisms of Inflammatory Gene Regulation by the Glucocorticoid Receptor. Front. Immunol. 2019, 10, 1859. [Google Scholar] [CrossRef]
- Vettorazzi, S.; Nalbantoglu, D.; Gebhardt, J.C.M.; Tuckermann, J. A guide to changing paradigms of glucocorticoid receptor function-a model system for genome regulation and physiology. FEBS J. 2021. online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Vandevyver, S.; Dejager, L.; Tuckermann, J.; Libert, C. New insights into the anti-inflammatory mechanisms of glucocorticoids: An emerging role for glucocorticoid-receptor-mediated transactivation. Endocrinology 2013, 154, 993–1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.C.; Derynck, M.K.; Nonaka, D.F.; Khodabakhsh, D.B.; Haqq, C.; Yamamoto, K.R. Chromatin immunoprecipitation (ChIP) scanning identifies primary glucocorticoid receptor target genes. Proc. Natl. Acad. Sci. USA 2004, 101, 15603–15608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandevyver, S.; Dejager, L.; Van Bogaert, T.; Kleyman, A.; Liu, Y.; Tuckermann, J.; Libert, C. Glucocorticoid receptor dimerization induces MKP1 to protect against TNF-induced inflammation. J. Clin. Investig. 2012, 122, 2130–2140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reichardt, H.M.; Tuckermann, J.P.; Göttlicher, M.; Vujic, M.; Weih, F.; Angel, P.; Herrlich, P.; Schütz, G. Repression of inflammatory responses in the absence of DNA binding by the glucocorticoid receptor. EMBO J. 2001, 20, 7168–7173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuckermann, J.P.; Reichardt, H.M.; Arribas, R.; Richter, K.H.; Schütz, G.; Angel, P. The DNA binding-independent function of the glucocorticoid receptor mediates repression of AP-1-dependent genes in skin. J. Cell Biol. 1999, 147, 1365–1370. [Google Scholar] [CrossRef] [PubMed]
- Vettorazzi, S.; Bode, C.; Dejager, L. Glucocorticoids limit acute lung inflammation in concert with inflammatory stimuli by induction of SphK1. Nat. Commun. 2015, 6, 7796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reichardt, H.M.; Kaestner, K.H.; Tuckermann, J. DNA binding of the glucocorticoid receptor is not essential for survival. Cell 1998, 93, 531–541. [Google Scholar] [CrossRef] [Green Version]
- Wepler, M.; Preuss, J.M.; Merz, T. Impaired Glucocorticoid Receptor Dimerization Aggravates LPS-Induced Circulatory and Pulmonary Dysfunction. Front. Immunol. 2020, 10, 3152. [Google Scholar] [CrossRef] [PubMed]
- Datzmann, T.; Hoffmann, A.; McCook, O. Effects of sodium thiosulfate (Na2S2O3) during resuscitation from hemorrhagic shock in swine with preexisting atherosclerosis. Pharm. Res. 2020, 151, 104536. [Google Scholar] [CrossRef]
- Childs, E.W.; Tharakan, B.; Byrge, N.; Tinsley, J.H.; Hunter, F.A.; Smythe, W.R. Angiopoietin-1 inhibits intrinsic apoptotic signaling and vascular hyperpermeability following hemorrhagic shock. Am. J. Physiol. Heart Circ. Physiol. 2008, 294, 2285–2295. [Google Scholar] [CrossRef]
- Wagner, K.; Gröger, M.; McCook, O.; Scheuerle, A.; Asfar, P.; Stahl, B.; Huber-Lang, M.; Ignatius, A.; Jung, B.; Duechs, M.; et al. Blunt chest trauma in mice after cigarette smoke-exposure: Effects of mechanical ventilation with 100% O2. PLoS ONE 2015, 10, e0132810. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Marini, F.; Binder, H. pcaExplorer: An R/Bioconductor package for interacting with RNA-seq principal components. BMC Bioinform. 2019, 20, 331. [Google Scholar] [CrossRef] [Green Version]
- Gu, Z.; Eils, R.; Schlesner, M. Complex heatmaps reveal patterns and correlations in multidimensional genomic data. Bioinformatics 2016, 32, 2847–2849. [Google Scholar] [CrossRef] [Green Version]
- Mi, H.; Ebert, D.; Muruganujan, A. PANTHER version 16: A revised family classification, tree-based classification tool, enhancer regions and extensive API. Nucleic Acids Res. 2021, 49, 394–403. [Google Scholar] [CrossRef]
- Hartmann, C.; Gröger, M.; Noirhomme, J.P. In-Depth Characterization of the Effects of Cigarette Smoke Exposure on the Acute Trauma Response and Hemorrhage in Mice. Shock 2019, 51, 68–77. [Google Scholar] [CrossRef]
- McQuin, C.; Goodman, A.; Chernyshev, V. CellProfiler 3.0: Next-generation image processing for biology. PLoS Biol. 2018, 16, e2005970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Preuss, J.M.; Burret, U.; Vettorazzi, S. Multiplex Fluorescent Bead-Based Immunoassay for the Detection of Cytokines, Chemokines, and Growth Factors. Proteomic Profiling 2021, 2261, 247–262. [Google Scholar]
- Cannon, J.W. Hemorrhagic shock. N. Engl. J. Med. 2018, 378, 370–379. [Google Scholar] [CrossRef]
- Asada, M.; Rauch, A.; Shimizu, H. DNA binding-dependent glucocorticoid receptor activity promotes adipogenesis via Krüppel-like factor 15 gene expression. Lab. Investig. 2011, 91, 203–215. [Google Scholar] [CrossRef] [Green Version]
- Bernasconi, A.R.; Yancoski, J.; Villa, M. Increased STAT1 Amounts Correlate with the Phospho-STAT1 Level in STAT1 Gain-of-function Defects. J. Clin. Immunol. 2018, 38, 745–747. [Google Scholar] [CrossRef]
- Rychli, K.; Kaun, C.; Hohensinner, P.; Rega, G.; Pfaffenberger, S.; Vyskocil, E.; Breuss, J.; Furnkranz, A.; Uhrin, P.; Zaujec, J.; et al. The inflammatory mediator oncostatin M induces angiopoietin 2 expression in endothelial cells in vitro and in vivo. J. Thromb. Haemost. 2010, 8, 596–604. [Google Scholar] [CrossRef]
- Papapetropoulos, A.; Fulton, D.; Mahboubi, K.; Kalb, R.G.; O’Connor, D.S.; Li, F.; Altieri, D.C.; Sessa, W.C. Angiopoietin-1 inhibits endothelial cell apoptosis via the Akt/survivin pathway. J. Biol. Chem. 2000, 275, 9102–9105. [Google Scholar] [CrossRef] [Green Version]
- Pan, Q.; Mathison, J.; Fearns, C. MDP-induced interleukin-1beta processing requires Nod2 and CIAS1/NALP3. J. Leukoc. Biol. 2007, 82, 177–183. [Google Scholar] [CrossRef]
- Yi, Y.S.; Jian, J.; Gonzalez-Gugel, E. p204 is Required for Canonical Lipopolysaccharide-induced TLR4 Signaling in Mice. EBioMedicine 2018, 29, 78–91. [Google Scholar] [CrossRef] [Green Version]
- Barsness, K.A.; Arcaroli, J.; Harken, A.H. Hemorrhage-induced acute lung injury is TLR-4 dependent. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2014, 287, 592–599. [Google Scholar] [CrossRef] [Green Version]
- Kleiman, A.; Hübner, S.; Parkitna, J.M.R. Glucocorticoid receptor dimerization is required for survival in septic shock via suppression of interleukin-1 in macrophages. FASEB J. 2012, 26, 722–729. [Google Scholar] [CrossRef]
- Wepler, M.; Preuss, J.M.; Merz, T.; McCook, O.; Radermacher, P.; Tuckermann, J.P.; Vettorazzi, S. Impact of downstream effects of glucocorticoid receptor dysfunction on organ function in critical illness-associated systemic inflammation. Intensive Care Med. Exp. 2020, 8, 37. [Google Scholar] [CrossRef]
- Guillon, A.; Preau, S.; Aboab, J.; Azabou, E.; Jung, B.; Silva, S.; Textoris, J.; Uhel, F.; Vodovar, D.; Zafrani, L.; et al. Preclinical septic shock research: Why we need an animal ICU. Ann. Intensive Care 2019, 9, 66. [Google Scholar] [CrossRef] [Green Version]
- Song, Z.; Zhao, X.; Gao, Y.; Liu, M.; Hou, M.; Jin, H.; Cui, Y. Recombinant human brain natriuretic peptide ameliorates trauma-induced acute lung injury via inhibiting JAK/STAT signaling pathway in rats. J. Trauma Acute Care Surg. 2015, 78, 980–987. [Google Scholar] [CrossRef]
- Ballegeer, M.; Van Looveren, K.; Timmermans, S. Glucocorticoid receptor dimers control intestinal STAT1 and TNF-induced inflammation in mice. J. Clin. Investig. 2018, 128, 3265–3279. [Google Scholar] [CrossRef]
- Surjit, M.; Ganti, K.P.; Mukherji, A.; Ye, T.; Hua, G.; Metzger, D.; Li, M.; Chambon, P. Widespread negative response elements mediate direct repression by agonist-liganded glucocorticoid receptor. Cell 2011, 145, 224–241. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Xie, W.; Wang, Y.; Chen, S.; Han, J.; Wang, L.; Gui, P.; Wu, Q. JAK2/STAT1-mediated HMGB1 translocation increases inflammation and cell death in a ventilator-induced lung injury model. Lab. Investig. 2019, 99, 1810–1821. [Google Scholar] [CrossRef]
- Mei, S.H.J.; McCarter, S.D.; Deng, Y.; Parker, C.H.; Liles, W.C.; Stewart, D.J. Prevention of LPS-induced acute lung injury in mice by mesenchymal stem cells overexpressing angiopoietin 1. PLoS Med. 2007, 4, 269. [Google Scholar] [CrossRef]
- Witzenbichler, B.; Westermann, D.; Knueppel, S.; Schultheiss, H.P.; Tschope, C. Protective role of angiopoietin-1 in endotoxic shock. Circulation 2005, 111, 97–105. [Google Scholar] [CrossRef] [Green Version]
- Kumada, Y.; Takahashi, T.; Shimizu, H.; Nakamura, R.; Omori, E.; Inoue, K.; Morimatsu, H. Therapeutic effect of carbon monoxide-releasing molecule-3 on acute lung injury after hemorrhagic shock and resuscitation. Exp. Med. 2019, 17, 3429–3440. [Google Scholar] [CrossRef] [Green Version]
- Tang, M.; Cai, J.; Wang, Y.; Huan, Z.; Yao, H.; Xu, C.; Ge, X.; Song, S. Tyrosine kinase nonreceptor 1 (TNK1) knockdown ameliorates hemorrhage shock-induced kidney injury via inhibiting macrophage M1 polarization. 3 Biotech 2021, 11, 501. [Google Scholar] [CrossRef]
- Tsung, Y.C.; Chung, C.Y.; Wan, H.C.; Chang, Y.Y.; Shih, P.C.; Hsu, H.S.; Kao, M.C.; Huang, C.J. Dimethyl Sulfoxide Attenuates Acute Lung Injury Induced by Hemorrhagic Shock/Resuscitation in Rats. Inflammation 2017, 40, 555–565. [Google Scholar] [CrossRef]
- Baron, R.M.; Carvajal, I.M.; Fredenburgh, L.E.; Liu, X.; Porrata, Y.; Cullivan, M.L.; Haley, K.J.; Sonna, L.A.; De Sanctis, G.T.; Ingenito, E.P.; et al. Nitric oxide synthase-2 down-regulates surfactant protein-B expression and enhances endotoxin-induced lung injury in mice. FASEB J. 2004, 18, 1276–1278. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | GR+/+ | GRdim/dim | p-Value |
|---|---|---|---|
| Alveolar membrane thickening | 1.0 (0) | 1.3 (0.15) | 0.0799 |
| Dystelectasis | 10 (2.5) | 6.4 (2.1) | 0.3100 |
| Emphysema | 1.3 (0.18) | 1.8 (0.22) | 0.2290 |
| Lymphocytes | 1.0 (0) | 1.1 (0.16) | 0.8981 |
| Macrophages | 8.5 (0.96) | 7.5 (0.53) | 0.3583 |
| Inflammatory alteration | 1.4 (0.18) | 1.7 (0.24) | 0.3245 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Preuss, J.M.; Burret, U.; Gröger, M.; Kress, S.; Scheuerle, A.; Möller, P.; Tuckermann, J.P.; Wepler, M.; Vettorazzi, S. Impaired Glucocorticoid Receptor Signaling Aggravates Lung Injury after Hemorrhagic Shock. Cells 2022, 11, 112. https://doi.org/10.3390/cells11010112
Preuss JM, Burret U, Gröger M, Kress S, Scheuerle A, Möller P, Tuckermann JP, Wepler M, Vettorazzi S. Impaired Glucocorticoid Receptor Signaling Aggravates Lung Injury after Hemorrhagic Shock. Cells. 2022; 11(1):112. https://doi.org/10.3390/cells11010112
Chicago/Turabian StylePreuss, Jonathan M., Ute Burret, Michael Gröger, Sandra Kress, Angelika Scheuerle, Peter Möller, Jan P. Tuckermann, Martin Wepler, and Sabine Vettorazzi. 2022. "Impaired Glucocorticoid Receptor Signaling Aggravates Lung Injury after Hemorrhagic Shock" Cells 11, no. 1: 112. https://doi.org/10.3390/cells11010112