
Differential Effects of 25-Hydroxyvitamin D3 versus 1α 25-Dihydroxyvitamin D3 on Adipose Tissue Browning in CKD-Associated Cachexia

Abstract
:1. Introduction
2. Materials and Methods
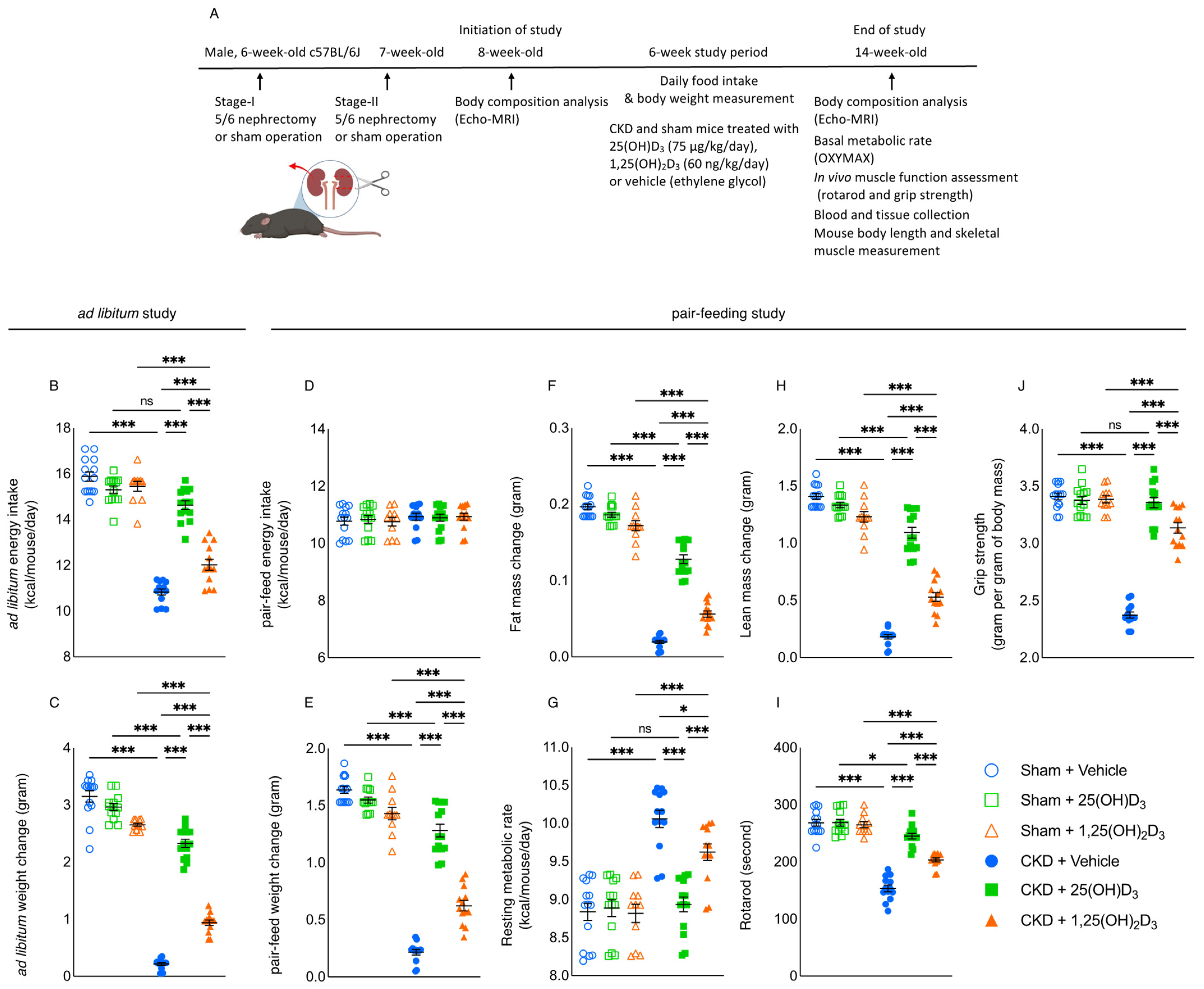
2.1. Study Design
2.2. Body Composition Analysis
2.3. Resting Metabolic Rate
2.4. Mouse Muscle Function
2.5. Serum and Blood Chemistry
2.6. Protein Assay for Muscle and Adipose Tissue
2.7. Fiber Size and Fatty Infiltration of Gastrocnemoius
2.8. RT2 Profiler PCR Array for Muscle Fibrosis
2.9. Muscle RNAseq Analysis
2.10. Quantative Real-Time PCR
2.11. Statistics
3. Results
3.1. Supplementation of Vitamin D Replenishes Serum Vitamin D Levels in CKD Mice
3.2. Repletion of 25-Hydroxyvitamin D3 Normalizes Caloric Intake and Improves Weight Gain in CKD Mice
3.3. Repletion of 25-Hydroxyvitamin D3 Improves Energy Homeostasis in CKD Mice
3.4. Repletion of 25-Hydroxyvitamin D3 Attenuates Adipose Tissue and Skeletal Muscle Energy Homeostasis in CKD Mice
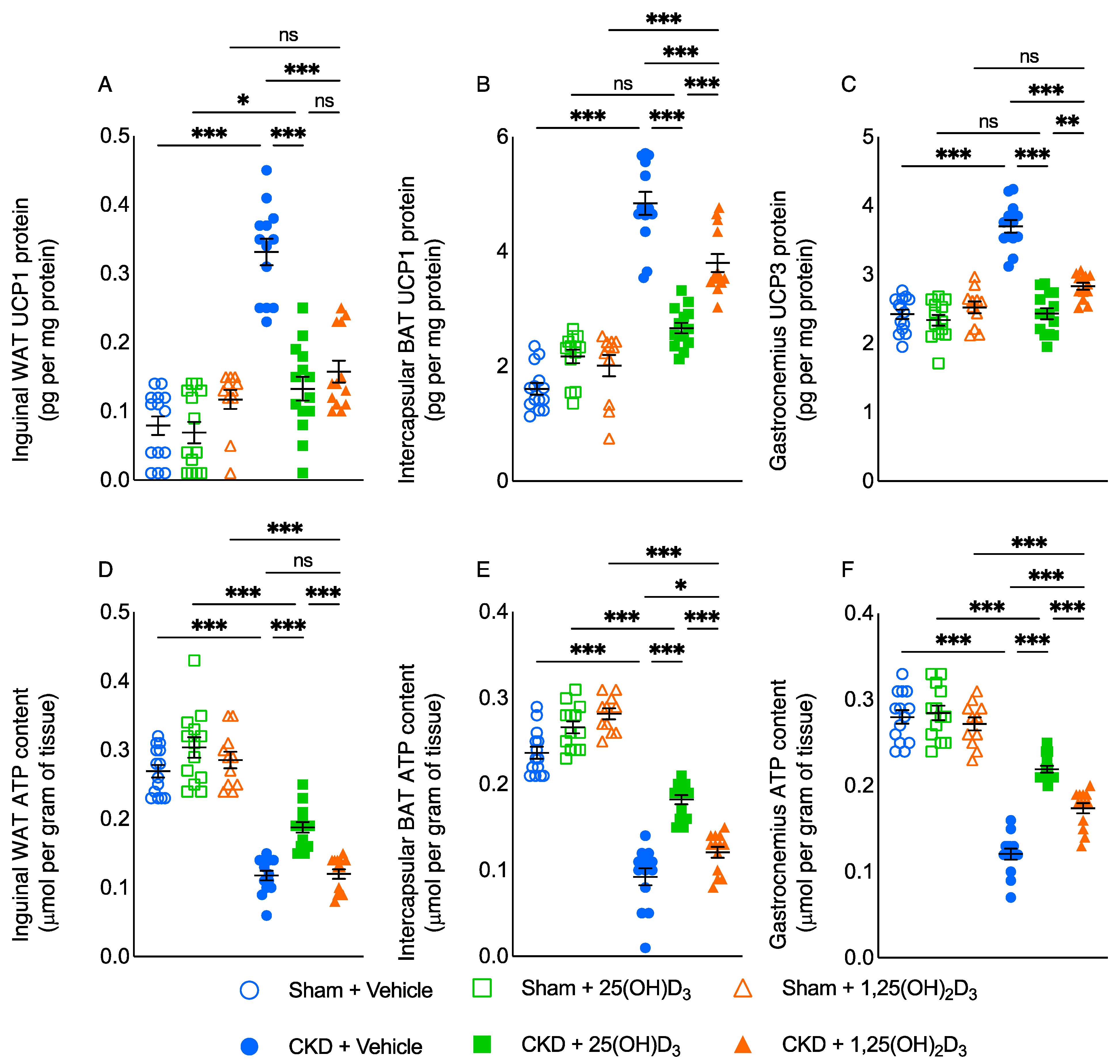
3.5. Repletion of 25-Hydroxyvitamin D3 Attenuates Browning of White Adipose Tissue in CKD Mice
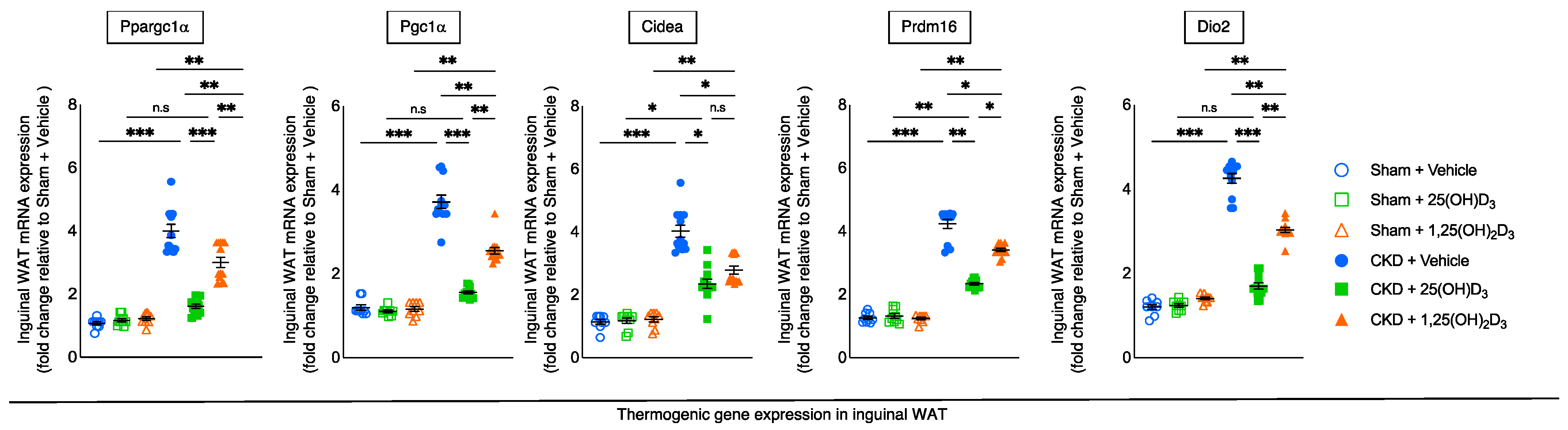
3.6. Repletion of 25-Hydroxyvitamin D3 Attenuates WAT Thermogenic Gene Expression in CKD Mice
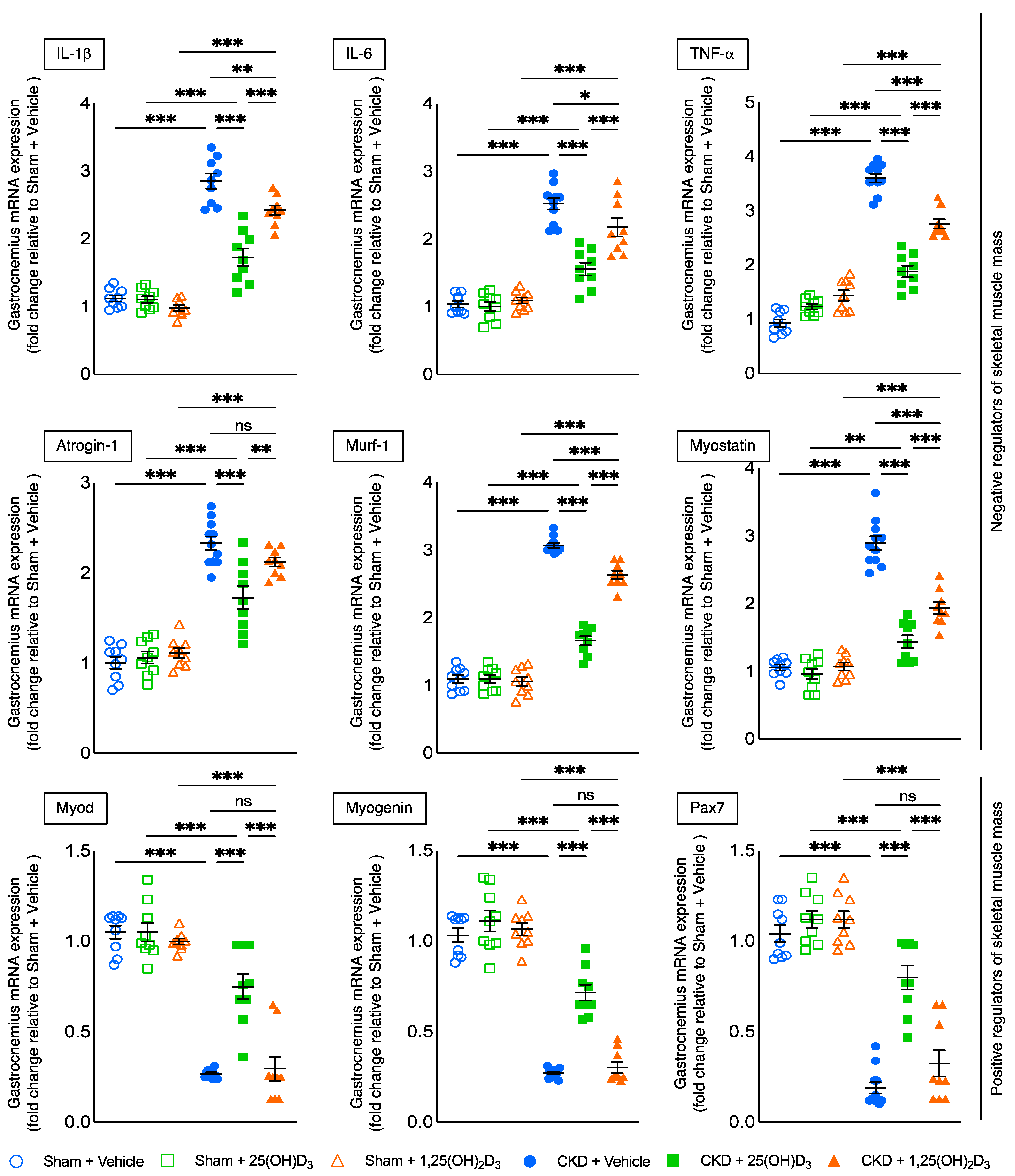
3.7. Repletion of 25-Hydroxyvitamin D3 Attenuates Muscle Wasting Signaling Pathways in CKD Mice
3.8. Repletion of 25-Hydroxyvitamin D3 Increases Muscle Fiber Size in CKD Mice
3.9. Repletion of 25-Hydroxyvitamin D3 Normalizes Muscle Fat Infiltration in CKD Mice
3.10. Repletion of 25-Hydroxyvitamin D3 Attenuates Aberrant Muscle Fibrotic Gene Expression in CKD Mice
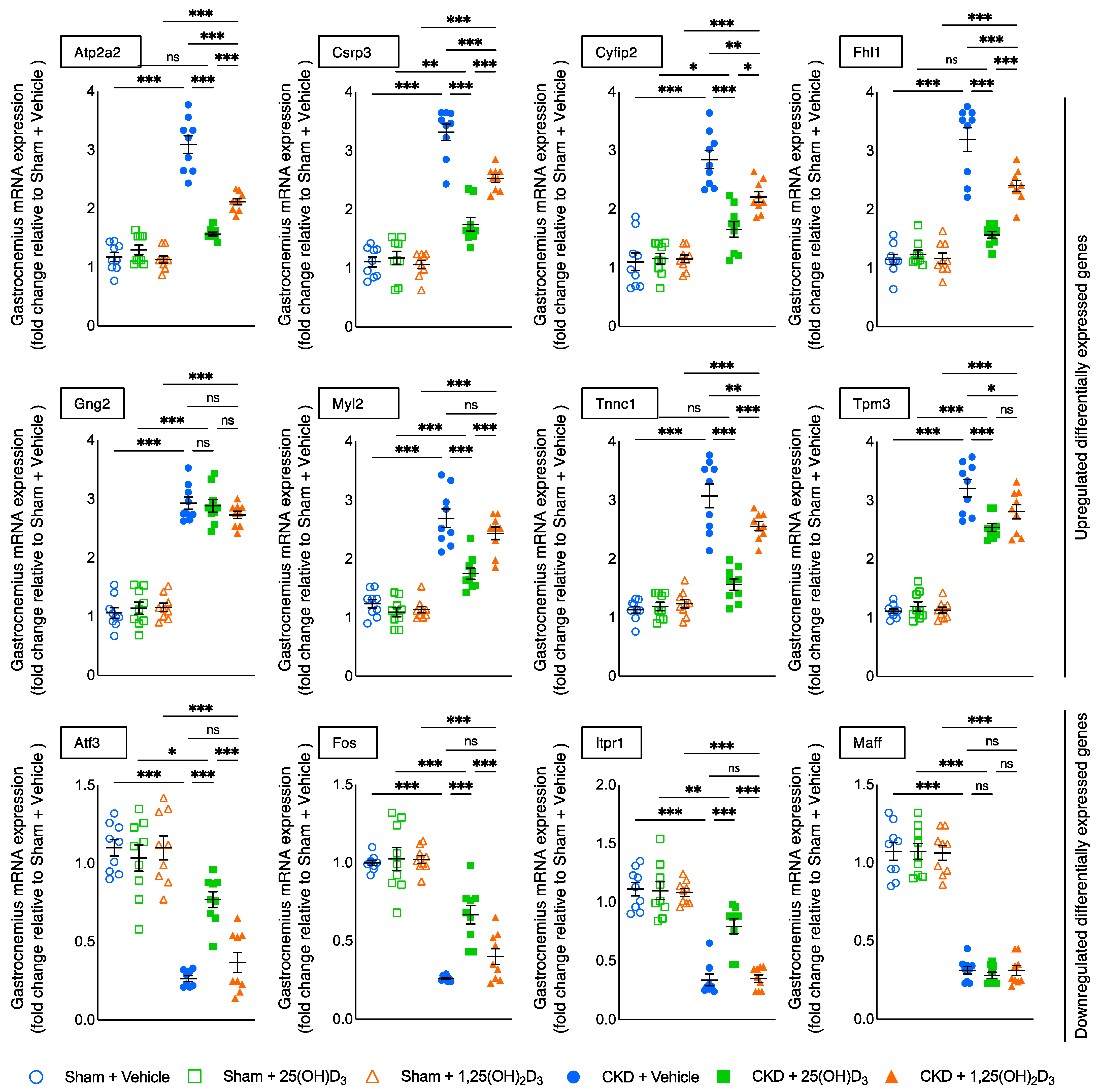
3.11. Molecular Mechanism of 25-Hydroxyvitamin D3 Repletion by RNAseq Analysis
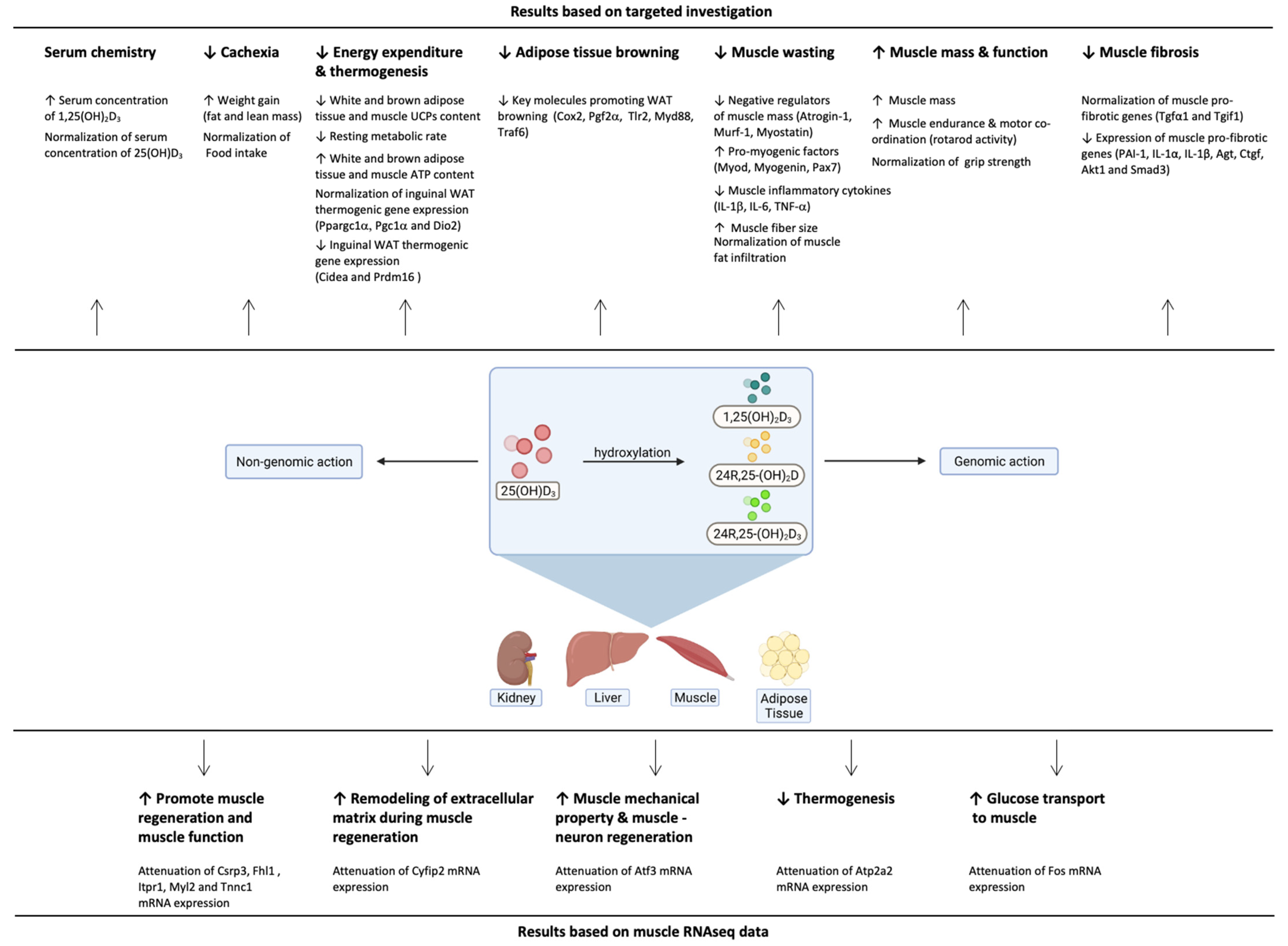
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Workeneh, B.T.; Mitch, W.E. Review of muscle wasting associated with chronic kidney disease. Am. J. Clin. Nutr. 2010, 91 (Suppl. S4), 1128S–1132S. [Google Scholar] [CrossRef] [Green Version]
- Mak, R.H.; Ikizler, A.T.; Kovesdy, C.P.; Raj, D.S.; Stenvinkel, P.; Kalantar-Zadeh, K. Wasting in chronic kidney disease. J. Cachexia Sarcopenia Muscle 2011, 2, 9–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sidossis, L.; Kajimura, S. Brown and beige fat in humans: Thermogenic adipocytes that control energy and glucose homeostasis. J. Clin. Investig. 2015, 125, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Bostrom, P.; Sparks, L.M.; Ye, L.; Choi, J.H.; Giang, A.-H.; Khandekar, M.; Virtanen, K.A.; Nuutila, P.; Schaart, G.; et al. Beige adipocytes are a distinct type of thermogenic fat cell in mouse and human. Cell 2012, 150, 366–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petruzzelli, M.; Schweiger, M.; Schreiber, R.; Campos-Olivas, R.; Tsoli, M.; Allen, J.; Swarbrick, M.; Rose-John, S.; Rincon, M.; Robertson, G.; et al. A switch from white to brown fat increases energy expenditure in cancer-associated cachexia. Cell Metab. 2014, 20, 433–447. [Google Scholar] [CrossRef] [Green Version]
- Vaitkus, J.A.; Celi, F.S. The role of adipose tissue in cancer-associated cachexia. Exp. Biol. Med. 2016, 242, 473–481. [Google Scholar] [CrossRef]
- Elattar, S.; Dimri, M.; Satyanarayana, A. The tumor secretory factor ZAG promotes white adipose tissue browning and energy wasting. FASEB J. 2018, 32, 4727–4743. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Liu, R.-X.; Zhu, M.-T.; Shen, W.-B.; Xie, J.; Zhang, Z.-Y.; Chen, N.; Shan, C.; Guo, X.-Z.; Lu, Y.-D.; et al. The browning of white adipose tissue and body weight loss in primary hyperparathyroidism. EBioMedicine 2019, 40, 56–66. [Google Scholar] [CrossRef] [Green Version]
- Querfeld, U.; Mak, R.H. Vitamin D deficiency and toxicity in chronic kidney disease: In search of the therapeutic window. Pediatr. Nephrol. 2010, 25, 2413–2430. [Google Scholar] [CrossRef]
- Dusso, A.S. Kidney disease and vitamin D levels: 25-hydroxyvitamin D, 1,25-dihydroxyvitamin D, and VDR activation. Kidney Int. Suppl. 2011, 1, 136–141. [Google Scholar] [CrossRef] [Green Version]
- Holden, R.M.; Morton, A.R.; Garland, J.S.; Pavlov, A.; Day, A.G.; Booth, S.L. Vitamins K and D status in stages 3-5 chronic kidney disease. Clin. J. Am. Soc. Nephrol. 2010, 5, 590–597. [Google Scholar] [CrossRef] [PubMed]
- Cheung, W.W.; Ding, W.; Hoffman, H.M.; Wang, Z.; Hao, S.; Zheng, R.; Gonzalez, A.; Zhan, J.Y.; Zhou, P.; Li, S.P.; et al. Vitamin D ameliorates adipose browning in chronic kidney disease cachexia. Sci. Rep. 2020, 10, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Lou, Y.-R.; Laaksi, I.; Syvala, A.; Blauer, M.; Tammela, T.L.J.; YLikomi, T.; Tuohimaa, P. 25-hydroxyvitamin D3 is an active hormone in human primary prostatic stromal cells. FASEB J. 2004, 18, 332–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, X.; Hawthorne, M.; Vaishnav, A.; St-Arnaud, R.; Mehta, R.G. 25-Hydroxyvitamin D3 is a natural chemopreventive agent against carcinogen induced precancerous lesions in mouse mammary gland organ culture. Breast Cancer Res. Treat. 2009, 113, 31–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lou, Y.-R.; Molnar, F.; Perakyla, M.; Qiao, S.; Kalueff, A.V.; St-Arnaud, R.; Carlberg, C.; Tuohimaa, P. 25-Hydroxyvitamin D(3) is an agonistic vitamin D receptor ligand. J. Steroid Biochem. Mol. Biol. 2010, 118, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Ritter, C.S.; Armbrecht, H.J.; Slatopolsky, E.; Brown, A.J. 25-Hydroxyvitamin D(3) suppresses PTH synthesis and secretion by bovine parathyroid cells. Kidney Int. 2006, 70, 654–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.L.; Ding, X.F.; Tong, J.; Li, B.Y. Partial rescue of the phenotype in 1alpha-hydroxylase gene knockout mice by vitamin D3 injection. Endocr. Res. 2011, 36, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Tuohimaa, P.; Wang, J.-H.; Khan, S.; Kuuslahti, M.; Qian, K.; Manninen, T.; Auvinen, P.; Vihinen, M.; Lou, Y.-R. Gene expression profiles in human and mouse primary cells provide new insights into the differential actions of vitamin D3 metabolites. PLoS ONE 2013, 8, e75338. [Google Scholar] [CrossRef]
- Gonzalez, A.; Cheung, W.W.; Perens, E.A.; Oliveira, E.A.; Gertler, A.; Mak, R.H. A leptin receptor antagonist attenuates adipose tissue browning and muscle wasting in infantile nephrotic cystinosis-associated cachexia. Cells 2021, 10, 1954. [Google Scholar] [CrossRef] [PubMed]
- Young, S.; Struys, E.; Wood, T. Quantification of creatine and guanidinoacetate using GC-MS and LC-MS/MS for the detection of cerebral creatine deficiency syndromes. Curr. Protoc. Hum. Genet. 2007, 54, 17.3.1–17.3.18. [Google Scholar] [CrossRef] [PubMed]
- Dubowitz, V.; Sewry, C.A.; Oldfors, A.; Lane, R.J.M. Muscle Biopsy: A Practical Approach; Fourth Edition Elsevier Health Sciences: Amsterdam, The Netherlands, 2013; 572p. [Google Scholar]
- Mehlem, A.; Hagberg, C.; Muhl, L.; Eriksson, U.; Falkevall, A. Imaging of neutral lipids by oil red O for analyzing the metabolic status in health and disease. Nat. Protoc. 2013, 8, 1149–1154. [Google Scholar] [CrossRef] [Green Version]
- Periasamy, M.; Herrera, J.L.; Reis, F.C.G. Skeletal muscle thermogenesis and its role in whole body energy metabolism. Diabetes Metab. J. 2017, 41, 327–336. [Google Scholar] [CrossRef]
- Vegiopoulos, A.; Müller-Decker, K.; Strzoda, D.; Schmitt, I.; Chichelnitskiy, E.; Ostertag, A.; Diaz, M.B.; Rozman, J.; de Angelis, M.H.; Nüsing, R.M.; et al. Cyclooxygenase-2 controls energy homeostasis in mice by de novo recruitment of brown adipocytes. Science 2010, 328, 1158–1161. [Google Scholar] [CrossRef]
- Ikeda, K.; Kang, Q.; Yoneshiro, T.; Camporez, J.P.; Maki, H.; Homma, M.; Shinoda, K.; Chen, Y.; Lu, X.; Maretich, P.; et al. UCP1-independent signaling involving SERCA2b-mediated calcium cycling regulates beige fat thermogenesis and systemic glucose homeostasis. Nat. Med. 2017, 23, 1454–1465. [Google Scholar] [CrossRef] [Green Version]
- Vafiadaki, E.; Arvanitis, D.A.; Papalouka, V.; Terzis, G.; Roumeliotis, T.I.; Spengos, K.; Garbos, S.D.; Manta, P.; Kranias, E.G.; Sanoudou, D. Muscle lim protein isoform negatively regulates striated muscle actin dynamic and differentiation. FEBS J. 2014, 281, 3261–3279. [Google Scholar] [CrossRef] [Green Version]
- Cui, C.; Han, S.; Tang, S.; He, H.; Shen, X.; Zhao, J.; Chen, Y.; Wei, Y.; Wang, Y.; Zhu, Q.; et al. The autophagy regulatory molecule CSRP3 interacts with LC3 and protects against muscular dystrophy. Int. J. Mol. Sci. 2020, 21, 749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Llano-Diez, M.; Gustafson, A.-M.; Olsson, C.; Göransson, H.; Larsson, L. Muscle wasting and the temporal gene expression pattern in a novel rat intensive care unit model. BMC Genom. 2011, 12, 602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saller, E.; Tom, E.; Brunori, M.; Otter, M.; Estreicher, A.; Mack, D.H.; Iggo, R. Increased apoptosis induction by 121F mutant p53. EMBO J. 1999, 18, 4424–4437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.Y.; Lori, D.; Wells, D.J.; Kemp, P.R. FHL1 activates myostatin signaling in skeletal muscle and promotes atrophy. FEBS Open Bio 2015, 5, 753–762. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Campagna, J.; John, V.; Damoiseaux, R.; Mokhonova, E.; Becerra, D.; Meng, H.; McNally, E.M.; Pyle, A.D.; Kramerova, I.; et al. A small-molceule approach to restore a slow-oxidative phenotype and defective CaMKIIβ signaling in limb girdle muscle dystrophy. Cell Rep. Med. 2020, 1, 100122. [Google Scholar] [CrossRef]
- Weterman, M.A.J.; Barth, P.G.; Van Spaendonck-Zwarts, K.Y.; Aronica, E.; Poll-The, B.-T.; Brouwer, O.F.; Van Tintelen, J.P.; Qahar, Z.; Bradley, E.J.; De Wissel, M.; et al. Recessive MYL2 mutations cause infantile type I muscle fibre disease and cardiomyopathy. Brain 2013, 136, 282–293. [Google Scholar] [CrossRef] [Green Version]
- Sheikh, F.; Lyon, R.C.; Chen, J. Functions of myosin light chain-2 in cardiac muscle and disease. Gene 2015, 569, 14–20. [Google Scholar] [CrossRef] [Green Version]
- Hui, X.; Zhu, W.; Wang, Y.; Lam, K.S.L.; Zhang, J.; Wu, D.; Kraegen, E.W.; Li, Y.; Xu, A. Major urinary protein-1 increases energy expenditure and improves glucose intolerance through enhancing mitochondrial function in skeletal muscle of diabetic mice. J. Biol. Chem. 2009, 284, 14050–14057. [Google Scholar] [CrossRef] [Green Version]
- Linda, H.; Skold, M.K.; Ochsmann, T. Activating transcription factor 3, a useful marker for regenerative response after nerve root injury. Front. Neurol. 2011, 2, 30. [Google Scholar] [CrossRef] [Green Version]
- Trouche, D.; Grigoriev, M.; Lenormand, J.-L.; Robin, P.; Leibovitch, S.A.; Sassone-Corsi, P.; Harel-Bellan, A.; Grigoriev, M. Repression of c-fos promoter by MyoD on muscle cell differentiation. Nat. Cell Biol. 1993, 363, 79–82. [Google Scholar] [CrossRef]
- Low, B.C.; Ross, I.K.; Grigor, M.R. Angiotension II stimulates glucose transport activity in cultures vascular smooth muscle cells. J. Biol. Chem. 1992, 267, 20740–20745. [Google Scholar] [CrossRef]
- Suman, M.; Sharpe, J.A.; Bentham, R.B.; Kotiadis, V.N.; Menegollo, M.; Pignataro, V.; Molgo, J.; Muntoni, F.; Duchen, M.R.; Pegoraro, E.; et al. Inositol trisphosphate receptor-mediated Ca2+ signalling stimulates mitochondrial function and gene expression in core myopathy patients. Hum. Mol. Genet. 2018, 27, 2367–2382. [Google Scholar] [CrossRef] [PubMed]
- Hassan-Smith, Z.; Jenkinson, C.; Smith, D.J.; Hernadez, I.; Morgan, S.A.; Crabtree, N.J.; Gittoes, N.J.; Keevil, B.G.; Stewart, P.M.; Hewison, M. 25-hydroxyvitamin D3 and 1,25-dihydroxyvitamin D3 exert distinct effects on human skeletal muscle function and gene expression. PLoS ONE 2017, 12, e0170665. [Google Scholar] [CrossRef] [Green Version]
- Van Driel, M.; Koedam, M.; Buurman, C.J.; Hewison, M.; Chiba, H.; Uitterlinden, A.G.; Pols, H.A.P.; van Leeuwen, J.P.T.M. Evidence for auto/paracrine actions of vitamin D in bone: 1alpha-hydroxylase expression and activity in human bone cells. FASEB J. 2006, 20, 2417–2419. [Google Scholar] [CrossRef]
- Atkins, G.J.; Anderson, P.H.; Findlay, D.M.; Welldon, K.J.; Vincent, C.; Zannettino, A.C.W.; O’Loughlin, P.D.; Morris, H.A. Metabolism of vitamin D3 in human osteoblasts: Evidence for autocrine and paracrine activities of 1 alpha,25-dihydroxyvitamin D3. Bone 2007, 40, 1517–1528. [Google Scholar] [CrossRef] [PubMed]
- Boullata, J.I. Vitamin D supplementation: A pharmacologic perspective. Curr. Opin. Clin. Nutr. Metab. Care 2010, 13, 677–684. [Google Scholar] [CrossRef] [PubMed]
- St-Arnaud, R. CYP24A1-deficient mice as a tool to uncover a biological activity for vitamin D metabolites hydroxylated at position 24. J. Steroid Biochem. Mol. Biol. 2010, 121, 254–256. [Google Scholar] [CrossRef]
- Askeno, A.; Saikatsu, S.; Kawane, T.; Horiuchi, N. Mouse vitamin D-24-hydroxylase: Molecular cloning, tissue distribution, and transcriptional regulation by 1alpha,25-dihydroxyvitamin D3. Endocrinology 1997, 138, 2233–2240. [Google Scholar] [CrossRef]
- Munetsuna, E.; Kawanami, R.; Nishikawa, M.; Ikeda, S.; Nakabayashi, S.; Yasuda, K.; Ohta, M.; Kamakura, M.; Ikushiro, S.; Sakaki, T. Anti-proliferative activity of 25-hydroxyvitamin D3 in human prostate cells. Mol. Cell. Endocrinol. 2014, 382, 960–970. [Google Scholar] [CrossRef] [PubMed]
- Horst, R.L.; Wovkulich, P.M.; Baggiolini, E.G.; Uskoković, M.R.; Engstrom, G.W.; Napoli, J.L. (23S)-1,23,25-Trihydroxyvitamin D3: Its biologic activity and role in 1 alpha,25-dihydroxyvitamin D3 26,23-lactone biosynthesis. Biochemistry 1984, 23, 3973–3979. [Google Scholar] [CrossRef] [PubMed]
- Kalousova, M.; Dusilova-Sulkova, S.; Zakiyanov, O.; Kostirova, M.; Safranek, R.; Tesar, V.; Zima, T. Vitamin D binding protein is not involved in Vitamin D deficiency in patients with chronic kidney disease. BioMed Res. Int. 2015, 2015, 492365. [Google Scholar] [CrossRef]
- Kir, S.; White, J.P.; Kleiner, S.; Kazak, L.; Cohen, P.; Baracos, V.E.; Spiegelman, B.M. Tumour-derived PTH-related protein triggers adipose tissue browning and cancer cachexia. Nat. Cell Biol. 2014, 513, 100–104. [Google Scholar] [CrossRef]
- Kir, S.; Komaba, H.; Garcia, A.P.; Economopoulos, K.P.; Liu, W.; Lanske, B.; Hodin, R.A.; Spiegelman, B.M. PTH/PTHrP receptor mediates cachexia in models of kidney failure and cancer. Cell Metab. 2016, 23, 315–323. [Google Scholar] [CrossRef] [Green Version]
- Ikizler, T.A.; Wingard, R.L.; Sun, M.; Harvell, J.; Parker, R.A.; Hakim, R.M. Increased energy expenditure in hemodialysis patients. J. Am. Soc. Nepohrol. 1996, 7, 2646–2653. [Google Scholar] [CrossRef]
- Wang, A.Y.-M.; Sea, M.M.-M.; Tang, N.; Sanderson, J.E.; Lui, S.-F.; Li, P.K.-T.; Woo, J. Resting energy expenditure and subsequent mortality risk in peritoneal dialysis patients. J. Am. Soc. Nephrol. 2004, 15, 3134–3143. [Google Scholar] [CrossRef] [Green Version]
- Rousset, S.; Alves-Guerra, M.-C.; Mozo, J.; Miroux, B.; Cassard-Doulcier, A.-M.; Bouillaud, F.; Ricquier, D. The biology of mitochondrial uncoupling proteins. Diabetes 2004, 53 (Suppl. S1), S130–S135. [Google Scholar] [CrossRef] [Green Version]
- Bing, C.; Brown, M.; King, P.; Collins, P.; Tisdsale, M.J.; Williams, G. Increased gene expression of brown fat uncoupling protein (UCP)1 and skeletal muscle UCP2 and UCP3 in MAC16-induced cancer cachexia. Cancer Res. 2000, 60, 2405–2410. [Google Scholar]
- Wong, K.E.; Szeto, F.L.; Zhang, W.; Ye, H.; Kong, J.; Zhang, Z.; Sun, X.J.; Li, Y.C. Involvement of the vitamin D receptor in energy metabolism: Regulation of uncoupling proteins. Am. J. Physiol. Endocrinol. Metab. 2009, 296, E820–E828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamei, Y.; Kawada, T.; Kazuki, R.; Ono, T.; Kato, S.; Sugimoto, E. Vitamin D receptor gene expression is up-regulated by 1, 25-dihydroxyvitamin D3 in 3T3-L1 preadipocytes. Biochem. Biophys. Res. Commun. 1993, 193, 948–955. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.-Q.; Pan, D.-X.; Hu, C.-Q.; Zhu, Y.-L.; Liu, X.-J. Vitamin D–vitamin D receptor system down-regulates expression of uncoupling proteins in brown adipocyte through interaction with hairless protein. Biosci. Rep. 2020, 40, BSR20194294. [Google Scholar] [CrossRef] [PubMed]
- Jonas, M.I.; Kurylowicz, A.; Bartoszewicz, Z.; Lisik, W.; Jonas, M.; Kozniewski, K.; Puzianowska-Kuznicka, M. Vitamin D receptor gene expression in adipose tissue of obese individuals is regulated by miRNA and correlates with the pro-inflammatory cytokine level. Int. J. Mol. Sci. 2019, 20, 5272. [Google Scholar] [CrossRef] [Green Version]
- Nimitphong, H.; Holick, M.F.; Fried, S.K.; Lee, M.-J. 25-hydroxyvitamin D3 and 1,25-dihydroxyvitamin D3 promote the differentiation of human subcutaneous preadipocytes. PLoS ONE 2012, 7, e52171. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.; Futawaka, K.; Komaya, R.; Fukuda, Y.; Hayashi, M.; Imamoto, M.; Miyawaki, T.; Kasahara, M.; Tagami, T.; Moriyama, K. Vitamin D3/VDR resists diet-induced obesity by modulating UCP3 expression in muscle. J. Biomed. Sci. 2016, 23, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Mierzejewski, B.; Archacka, K.; Grabowska, I.; Florkowska, A.; Ciemerych, M.A.; Brzoska, E. Human and mouse skeletal muscle stem and progenitor cells in health and disease. Semin. Cell Dev. Biol. 2020, 104, 93–104. [Google Scholar] [CrossRef]
- Dengjie, Y.; Cai, Z.; Li, D.; Zhang, Y.; He, M.; Yang, Y.; Liu, D.; Xie, W.; Li, Y.; Xiao, W. Myogenic differentiation of stem cells for skeletal muscle regeneration. Stem Cells Int. 2021, 2021, 8884283. [Google Scholar]
- Srikuea, R.; Hirunsai, M. Effects of intramuscular administration of 1α,25(OH)2D3 during skeletal muscle regeneration on regenerative capacity, muscle fibrosis, and angiogenesis. J. Appl. Physiol. 2016, 120, 1381–1393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgerton, V.R.; Smith, J.L.; Simpson, D.R. Muscle fibre type populations of human leg muscles. Histochem. J. 1975, 7, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Mann, C.J.; Perdiguero, E.; Kharraz, Y.; Aguilar, S.; Pessina, P.; Serrano, A.L.; Munoz-Canoves, P. Aberrant repair and fibrosis development in skeletal muscle. Skelet. Muscle 2011, 1, 1–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sham + Vehicle (n = 6) | CKD + Vehicle (n = 8) | CKD + 25(OH)D3 (n = 8) | CKD + 1,25(OH)2D3 (n = 8) | |
|---|---|---|---|---|
| BUN (mg/dL) | 26.7 ± 3.2 | 54.7 ± 16.1 a | 64.3 ± 14.1 a | 62.1 ± 5.9 a |
| Creatinine (mg/dL) | 0.12 ± 0.04 | 0.23 ± 0.05 a | 0.18 ± 0.02 a | 0.21 ± 0.06 a |
| Bicarbonate (mmol/L) | 27.6 ± 3.2 | 28.1 ± 2.5 | 26.3 ± 2.8 | 27.4 ± 1.6 |
| 25(OH)D3 (ng/mL) | 110.3 ± 28.4 | 43.5 ± 4.1 a | 69.7 ± 8.4 a,b | 57.6 ± 10.5 a |
| 1,25(OH)2D3 (pg/mL) | 282.1 ± 56.6 | 124.3 ± 24.1 a | 146.7 ± 25.1 a | 176.1 ± 27.9 a,b |
| Sham + Vehicle (n = 6) | CKD + Vehicle (n = 6) | CKD + 25(OH)D3 (n = 6) | CKD + 1,25(OH)2D3 (n = 6) | |
|---|---|---|---|---|
| BUN (mg/dL) | 30.1± 3.8 | 72.3 ± 12.1 a | 83.1 ± 16.8 a | 58.6 ± 13.7 a |
| Creatinine (mg/dL) | 0.09 ± 0.02 | 0.28 ± 0.08 a | 0.27 ± 0.03 a | 0.26 ± 0.06 a |
| Bicarbonate (mmol/L) | 27.3 ± 1.4 | 27.5 ± 0.9 | 27.2 ± 0.8 | 27.8 ± 1.4 |
| 25(OH)D3 (ng/mL) | 121.5 ± 22.1 | 50.7 ± 8.7 a | 97.6 ± 7.6 a,b | 60.6 ± 7.4 a |
| 1,25(OH)2D3 (pg/mL) | 267.8 ± 33.6 | 136.4 ± 16.1 a | 176.8 ± 27.3 a | 231.1 ± 25.4 a,b |
| Sham + Vehicle (n = 4) | CKD + Vehicle (n = 6) | CKD + 25(OH)D3 (n = 8) | CKD + 1,25(OH)2D3 (n = 8) | |
|---|---|---|---|---|
| BUN (mg/dL) | 27.5 ± 6.4 | 65.8 ± 11.4 a | 67.8 ± 5.1 a | 75.4 ± 9.3 a |
| Creatinine (mg/dL) | 0.11 ± 0.02 | 0.18 ± 0.02 a | 0.21 ± 0.03 a | 0.23 ± 0.07 a |
| Bicarbonate (mmol/L) | 27.6 ± 2.2 | 27.8 ± 2.3 | 27.3 ± 2.1 | 27.3 ± 1.6 |
| 25(OH)D3 (ng/mL) | 113.5 ± 11.9 | 50.7 ± 9.4 a | 115.3 ± 7.6 b | 67.4 ± 11.5 a |
| 1,25(OH)2D3 (pg/mL) | 267.3 ± 23.1 | 116.4 ± 17.2 a | 213.4 ± 21.3 a,b | 257.1 ± 25.2 b |
| Sham + Vehicle (n = 14) | Sham + 25(OH)D3 (n = 13) | Sham + 1,25(OH)2D3 (n = 11) | CKD + Vehicle (n = 13) | CKD + 25(OH)D3 (n = 14) | CKD + 1,25(OH)2D3 (n = 13) | |
|---|---|---|---|---|---|---|
| BUN (mg/dL) | 26.5 ± 5.6 | 32.4 ± 3.1 | 27.3 ± 4.3 | 57.5 ± 6.5 a | 68.7 ± 6.9 a | 58.7 ± 6.4 a |
| Creatinine (mg/dL) | 0.09 ± 0.03 | 0.12 ± 0.04 | 0.09 ± 0.02 | 0.21 ± 0.04 a | 0.17 ± 0.04 a | 0.19 ± 0.05 a |
| Bicarbonate (mmol/L) | 27.5 ± 1.2 | 27.8 ± 1.4 | 27.9 ± 1.2 | 26.8 ± 1.5 | 26.5 ± 1.7 | 27.9 ± 2.3 |
| 25(OH)D3 (ng/mL) | 109.5 ± 14.4 | 112.4 ± 16.5 | 124.3 ± 15.6 | 54.7 ± 11.6 b | 105.4 ± 13.8 c,d | 53.8 ± 9.8 b |
| 1,25(OH)2D3 (pg/mL) | 256.7 ± 21.3 | 243.5 ± 24.3 | 265.4 ± 19.5 | 136.5 ± 15.3 b | 185.7 ± 15.5 b,c | 246.9 ± 21.4 c,d |
| Sham + Vehicle (n = 11) | Sham + 25(OH)D3 (n = 11) | Sham + 1,25(OH)2D3 (n = 11) | CKD + Vehicle (n = 14) | CKD + 25(OH)D3 (n = 12) | CKD + 1,25(OH)2D3 (n = 13) | |
|---|---|---|---|---|---|---|
| BUN (mg/dL) | 27.4 ± 4.6 | 29.5 ± 3.7 | 28.5 ± 5.6 | 64.6 ± 6.6 a | 72.5 ± 8.7 a | 68.7 ± 9.9 a |
| Ca (mg/dL) | 11.2 ± 0.4 | 11.9± 0.5 | 11.4 ± 0.7 | 9.7 ± 0.4 b | 8.9 ± 0.7 b | 9.4 ± 0.5 b |
| Creatinine (mg/dL) | 0.06 ± 0.04 | 0.09 ± 0.03 | 0.07 ± 0.02 | 0.17± 0.05 a | 0.18 ± 0.07 a | 0.21 ± 0.07 a |
| Bicarbonate (mmol/L) | 28.5 ± 2.3 | 28.3 ± 2.2 | 28.3 ± 2.1 | 27.8 ± 2.5 | 26.8 ± 2.4 | 26.7 ± 2.1 |
| 25(OH)D3 (ng/mL) | 115.5 ± 18.7 | 107.8 ± 12.6 | 123.2 ± 21.5 | 47.8 ± 9.8 b | 124.6 ± 11.5 c,d | 50.1 ± 11.4 b |
| 1,25(OH)2D3 (pg/mL) | 268.7 ± 17.4 | 277.4 ± 31.7 | 247.8 ± 23.6 | 109.6 ± 21.5 b | 207.8 ± 21.5 c | 276.5 ± 33.7 c,d |
| Pi (mg/dL) | 7.5 ± 0.3 | 7.3 ± 0.6 | 7.4 ± 0.6 | 9.7 ± 0.5 a | 9.8 ± 0.6 a | 9.6 ± 0.4 a |
| PTH (pg/mL) | 118.3 ± 15.7 | 105.7 ± 17.8 | 109.5 ± 19.8 | 326.7 ± 23.6 a | 283 ± 32.5 a | 275.5 ± 27.3 a |
| VDBP (ug/mL) | 398.5 ± 27.5 | 408.4 ± 26.7 | 387.5 ± 27.6 | 591.5 ± 25.6 a | 581.8 ± 23.1 a | 625.8 ± 25.7 a |
| Upregulated DEG | Functional Significance and References |
|---|---|
| Atp2a2 | associated with UCP-1 independent beige thermogenesis [25] |
| Csrp3 | associated with skeletal muscle dystrophy [26,27] |
| Cyfip2 | associated with muscle atrophy [28,29] |
| Fhl1 | activates myostatin signaling and promotes atrophy in skeletal muscle [30] |
| Myl2 | associated with muscle cycling kinetics [31,32,33] |
| Tnnc1 | biomarker for muscle depolarization [34] |
| Downregulated DEG | Functional Significance and References |
| Atf3 | Biomarker for neural injury, associated with reduced regeneration of neurons [35] |
| Fos | associated with decreased skeletal muscle regeneration [36] |
| Itpr1 | impairs glucose transport in muscle [37] associated with decreased muscle regeneration and mitochondrial dysfunction in myopathy patients [38] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mak, R.H.; Querfeld, U.; Gonzalez, A.; Gunta, S.; Cheung, W.W. Differential Effects of 25-Hydroxyvitamin D3 versus 1α 25-Dihydroxyvitamin D3 on Adipose Tissue Browning in CKD-Associated Cachexia. Cells 2021, 10, 3382. https://doi.org/10.3390/cells10123382
Mak RH, Querfeld U, Gonzalez A, Gunta S, Cheung WW. Differential Effects of 25-Hydroxyvitamin D3 versus 1α 25-Dihydroxyvitamin D3 on Adipose Tissue Browning in CKD-Associated Cachexia. Cells. 2021; 10(12):3382. https://doi.org/10.3390/cells10123382
Chicago/Turabian StyleMak, Robert H., Uwe Querfeld, Alex Gonzalez, Sujana Gunta, and Wai W. Cheung. 2021. "Differential Effects of 25-Hydroxyvitamin D3 versus 1α 25-Dihydroxyvitamin D3 on Adipose Tissue Browning in CKD-Associated Cachexia" Cells 10, no. 12: 3382. https://doi.org/10.3390/cells10123382