
CD46 Genetic Variability and HIV-1 Infection Susceptibility

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Genotyping
2.3. Biostatistics and Bioinformatics Analysis
2.4. Ethics
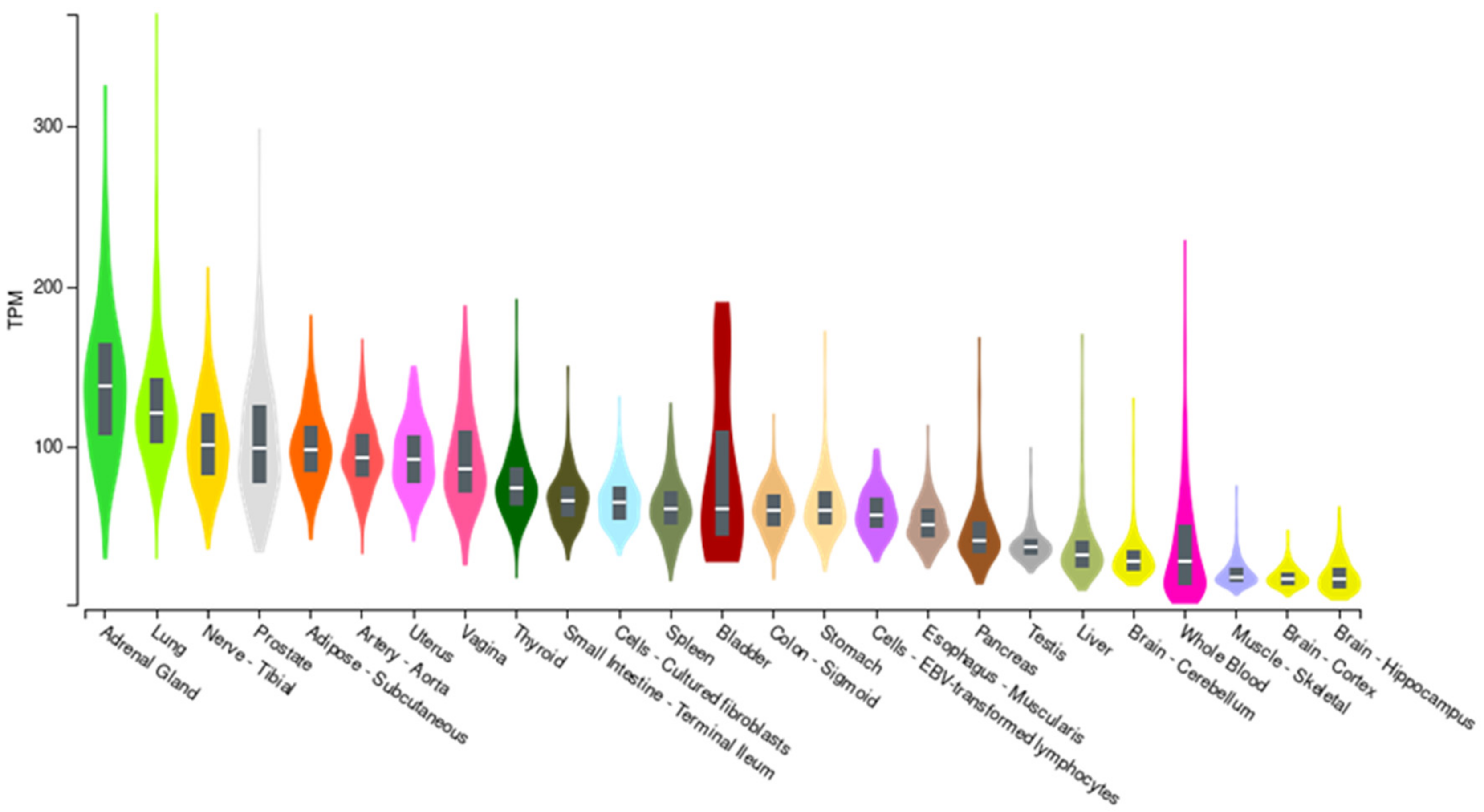
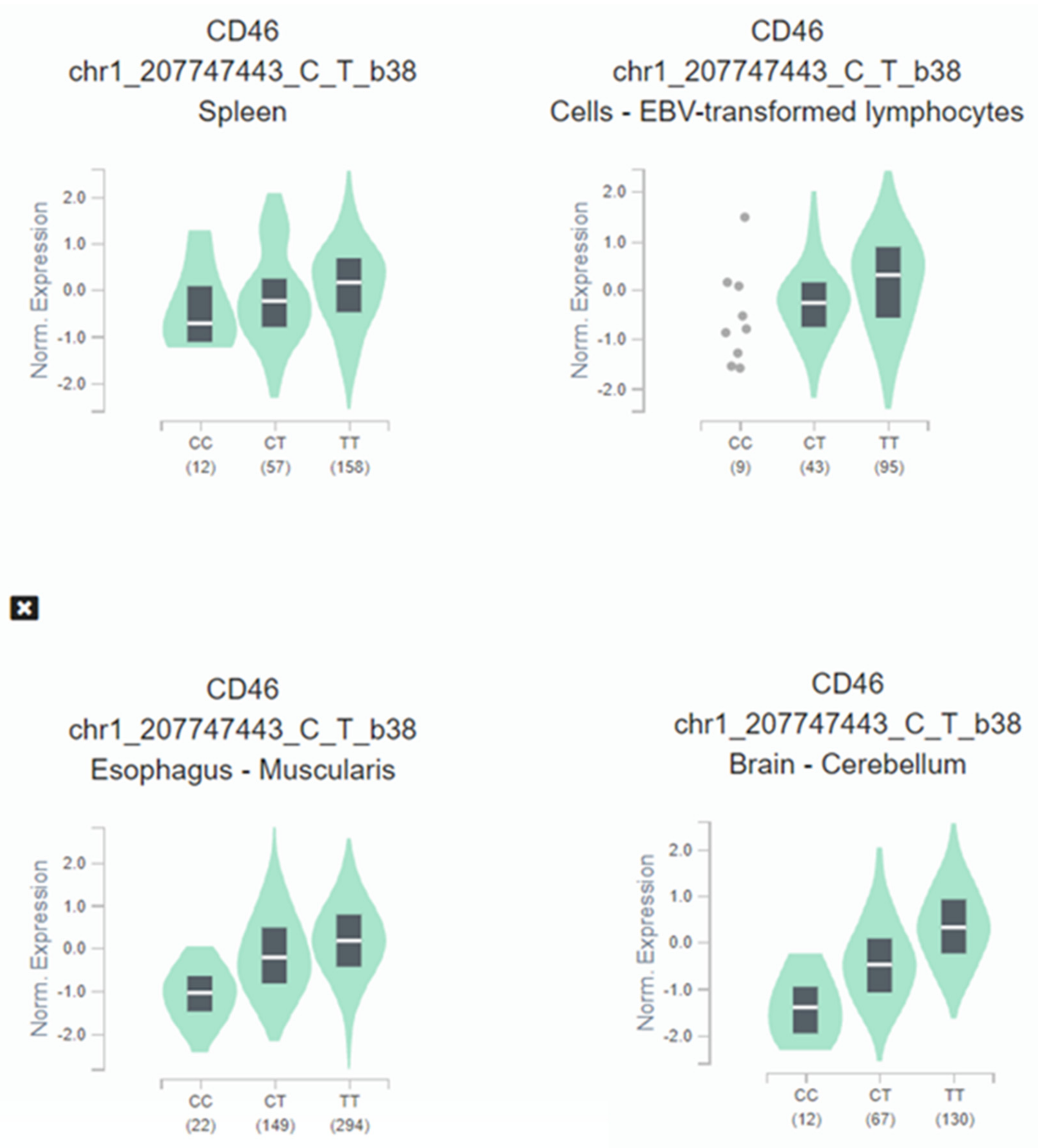
3. Results
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Merle, N.S.; Church, S.E.; Fremeaux-Bacchi, V.; Roumenina, L.T. Complement system part I—Molecular mechanisms of activation and regulation. Front. Immunol. 2015, 6, 262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merle, N.S.; Noe, R.; Halbwachs-Mecarelli, L.; Fremeaux-Bacchi, V.; Roumenina, L.T. Complement system part II: Role in immunity. Front. Immunol. 2015, 6, 257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, H.; Fara, A.; Dasgupta, P.; Kemper, C. CD46: The ‘multitasker’ of complement proteins. Int. J. Biochem. Cell Biol. 2013, 45, 2808–2820. [Google Scholar] [CrossRef] [PubMed]
- Stoiber, H.; Clivio, A.; Dierich, M.P. Role of complement in HIV infection. Annu. Rev. Immunol. 1997, 15, 649–674. [Google Scholar] [CrossRef]
- Bánki, Z.; Stoiber, H.; Dierich, M.P. HIV and human complement: Inefficient virolysis and effective adherence. Immunol. Lett. 2005, 97, 209–214. [Google Scholar] [CrossRef]
- Prohászka, Z.; Hidvégi, T.; Ujhelyi, E.; Stoiber, H.; Dierich, M.P.; Süsal, C.; Füst, G. Interaction of complement and specific antibodies with the external glycoprotein 120 of HIV-1. Immunology 1995, 85, 184–189. [Google Scholar]
- Ji, X.; Gewurz, H.; Spear, G.T. Mannose binding lectin (MBL) and HIV. Mol. Immunol. 2005, 42, 145–152. [Google Scholar] [CrossRef]
- Stoiber, H.; Kacani, L.; Speth, C.; Würzner, R.; Dierich, M.P. The supportive role of complement in HIV pathogenesis. Immunol. Rev. 2001, 180, 168–176. [Google Scholar] [CrossRef]
- Montefiori, D.C.; Cornell, R.J.; Zhou, J.Y.; Zhou, J.T.; Hirsch, V.M.; Johnson, P.R. Complement control proteins, CD46, CD55, and CD59, as common surface constituents of human and simian immunodeficiency viruses and possible targets for vaccine protection. Virology 1994, 205, 82–92. [Google Scholar] [CrossRef]
- Saifuddin, M.; Hedayati, T.; Atkinson, J.; Holguin, M.; Parker, C.; Spear, G. Human immunodeficiency virus type 1 incorporates both glycosyl phosphatidylinositol-anchored CD55 and CD59 and integral membrane CD46 at levels that protect from complement-mediated destruction. J. Gen. Virol. 1997, 78, 1907–1911. [Google Scholar] [CrossRef] [Green Version]
- Biswas, M.; Johnson, J.B.; Kumar, S.R.P.; Parks, G.D.; Subbiah, E. Incorporation of host complement regulatory proteins into newcastle disease virus enhances complement evasion. J. Virol. 2012, 86, 12708–12716. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Parks, G.D. Relative contribution of cellular complement inhibitors CD59, CD46, and CD55 to parainfluenza virus 5 inhibition of complement-mediated neutralization. Viruses 2018, 10, 219. [Google Scholar] [CrossRef] [Green Version]
- Ozog, S.; Chen, C.X.; Simpson, E.; Garijo, O.; Timberlake, N.D.; Minder, P.; Verhoeyen, E.; Torbett, B.E. CD46 null packaging cell line improves measles lentiviral vector production and gene delivery to hematopoietic stem and progenitor cells. Mol. Ther. Methods Clin. Dev. 2019, 13, 27–39. [Google Scholar] [CrossRef] [Green Version]
- Kathryn Liszewski, M.; Atkinson, J.P. Complement regulator CD46: Genetic variants and disease associations. Hum. Genom. 2015, 9, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Turek, I.; Irving, H. Moonlighting proteins shine new light on molecular signaling niches. Int. J. Mol. Sci. 2021, 22, 1367. [Google Scholar] [CrossRef]
- Anderson, D.; Abbott, A.; Jack, R. The role of complement component C3b and its receptors in sperm-oocyte interaction. Proc. Natl. Acad. Sci. USA 1993, 90, 10051–10055. [Google Scholar] [CrossRef] [Green Version]
- Díaz-Morán, S.; Palència, M.; Mont-Cardona, C.; Cañete, T.; Blázquez, G.; Martínez-Membrives, E.; López-Aumatell, R.; Sabariego, M.; Donaire, R.; Morón, I.; et al. Gene expression in hippocampus as a function of differential trait anxiety levels in genetically heterogeneous NIH-HS rats. Behav. Brain Res. 2013, 257, 129–139. [Google Scholar] [CrossRef]
- Le Friec, G.; Sheppard, D.; Whiteman, P.; Karsten, C.; Shamoun, S.; Laing, A.; Bugeon, L.; Dallman, M.; Melchionna, T.; Chillakuri, C.; et al. The CD46-Jagged1 interaction is critical for human TH1 immunity. Nat. Immunol. 2012, 13, 1213–1221. [Google Scholar] [CrossRef] [Green Version]
- Sakurai, F.; Nakashima, K.; Yamaguchi, T.; Ichinose, T.; Kawabata, K.; Hayakawa, T.; Mizuguchi, H. Adenovirus serotype 35 vector-induced innate immune responses in dendritic cells derived from wild-type and human CD46-transgenic mice: Comparison with a fiber-substituted Ad vector containing fiber proteins of Ad serotype 35. J. Control. Release 2010, 148, 212–218. [Google Scholar] [CrossRef]
- Hirano, A.; Kurita-Taniguchi, M.; KatayamaY, Y.; Matsumoto, M.; Wong, T.; Seya, T. Ligation of human CD46 with purified complement C3b or F(ab′)2 of monoclonal antibodies enhances isoform-specific interferon gamma-dependent nitric oxide production in macrophages. J. Biochem. 2002, 132, 83–91. [Google Scholar] [CrossRef]
- Kurita-Taniguchi, M.; Fukui, A.; Hazeki, K.; Hirano, A.; Tsuji, S.; Matsumoto, M.; Watanabe, M.; Ueda, S.; Seya, T. Functional modulation of human macrophages through CD46 (measles virus receptor): Production of IL-12 p40 and nitric oxide in association with recruitment of protein-tyrosine phosphatase SHP-1 to CD46. J. Immunol. 2000, 165, 5143–5152. [Google Scholar] [CrossRef] [Green Version]
- Oliaro, J.; Pasam, A.; Waterhouse, N.J.; Browne, K.; Ludford-Menting, M.; Trapani, J.; Russell, S. Ligation of the cell surface receptor, CD46, alters T cell polarity and response to antigen presentation. Proc. Natl. Acad. Sci. USA 2006, 103, 18685–18690. [Google Scholar] [CrossRef] [Green Version]
- Liszewski, M.K.; Kolev, M.; Le Friec, G.; Leung, M.; Bertram, P.G.; Fara, A.F.; Subias, M.; Pickering, M.C.; Drouet, C.; Meri, S.; et al. Intracellular complement activation sustains T cell homeostasis and mediates effector differentiation. Immunity 2013, 39, 1143–1157. [Google Scholar] [CrossRef] [Green Version]
- Arbore, G.; West, E.E.; Rahman, J.; Le Friec, G.; Niyonzima, N.; Pirooznia, M.; Tunc, I.; Pavlidis, P.; Powell, N.; Li, Y.; et al. Complement receptor CD46 co-stimulates optimal human CD8+ T cell effector function via fatty acid metabolism. Nat. Commun. 2018, 9, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Oliver, M.A.; Rojo, J.M.; Rodriguez de Cordoba, S.; Alberti, S. Binding of complement regulatory proteins to group A Streptococcus. Vaccine 2008, 26, I75–I78. [Google Scholar] [CrossRef] [Green Version]
- Gill, D.B.; Koomey, M.; Cannon, J.G.; Atkinson, J.P. Down-regulation of CD46 by Piliated Neisseria gonorrhoeae. J. Exp. Med. 2003, 198, 1313–1322. [Google Scholar] [CrossRef] [Green Version]
- Li, K.; Feito, M.; Sacks, S.; Sheerin, N. CD46 (membrane cofactor protein) acts as a human epithelial cell receptor for internalization of opsonized uropathogenic Escherichia coli. J. Immunol. 2006, 177, 2543–2551. [Google Scholar] [CrossRef] [Green Version]
- de Astorza, B.; Cortés, G.; Crespí, C.; Saus, C.; Rojo, J.; Albertí, S. C3 promotes clearance of Klebsiella pneumoniae by A549 epithelial cells. Infect. Immun. 2004, 72, 1767–1774. [Google Scholar] [CrossRef] [Green Version]
- Mahtout, H.; Chandad, F.; Rojo, J.M.; Grenier, D. Fusobacterium nucleatum Binding to complement regulatory protein CD46 modulates the expression and secretion of cytokines and matrix metalloproteinases by oral epithelial cells. J. Periodontol. 2011, 82, 311–319. [Google Scholar] [CrossRef]
- Dörig, R.; Marcil, A.; Chopra, A.; Richardson, C. The human CD46 molecule is a receptor for measles virus (Edmonston strain). Cell 1993, 75, 295–305. [Google Scholar] [CrossRef]
- Santoro, F.; Kennedy, P.E.; Locatelli, G.; Malnati, M.S.; Berger, E.A.; Lusso, P. CD46 is a cellular receptor for human herpesvirus 6. Cell 1999, 99, 817–827. [Google Scholar] [CrossRef] [Green Version]
- Stein, K.R.; Gardner, T.J.; Hernandez, R.E.; Kraus, T.A.; Duty, J.A.; Ubarretxena-Belandia, I.; Moran, T.M.; Tortorella, D. CD46 facilitates entry and dissemination of human cytomegalovirus. Nat. Commun. 2019, 10, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.F.; Lee, C.Y. Adenoviruses types, cell receptors and local innate cytokines in adenovirus infection. Int. Rev. Immunol. 2014, 33, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Liszewski, M.K.; Atkinson, J.P. Membrane cofactor protein (MCP.; CD46): Deficiency states and pathogen connections. Curr. Opin. Immunol. 2021, 72, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Byun, S.; Han, S.; Zheng, Y.; Planelles, V.; Lee, Y. The landscape of alternative splicing in HIV-1 infected CD4 T-cells. BMC Med. Genom. 2020, 13, 38. [Google Scholar] [CrossRef] [PubMed]
- Mishra, N.; Mohata, M.; Aggarwal, H.; Chaudhary, O.; Das, B.K.; Sinha, S.; Hazarika, A.; Luthra, K. Expression of complement receptor 3 (CR3) and regulatory protein CD46 on dendritic cells of antiretroviral naïve and treated HIV-1 infected individuals: Correlation with immune activation status. Mol. Immunol. 2018, 96, 83–87. [Google Scholar] [CrossRef] [PubMed]
- Aries, S.P.; Schaaf, B.; Hansen, F.; Weyrich, K.; Kurowski, V.; Dennin, R.; Dalhoff, K. Expression of complement receptors and regulatory proteins on alveolar CD4+ lymphocytes from human immunodeficiency virus-1 infected individuals. Eur. Respir. J. 1997, 10, 1736–1741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarvis, J.N.; Taylor, H.; Long, P.M.; Gutta, P.V.; Pousak, T.; Fine, N. Diminished expression of cell-surface complement regulatory proteins in HIV-infected children and with HIV infection of peripheral blood mononuclear cells in vitro. J. Acquir. Immune Defic. Syndr. Hum. Retrovirol. 1995, 9, 249–256. [Google Scholar] [CrossRef]
- Fenizia, C.; Rossignol, J.-F.; Clerici, M.; Biasin, M. Genetic and immune determinants of immune activation in HIV-exposed seronegative individuals and their role in protection against HIV infection. Infect. Genet. Evol. 2018, 66, 325–334. [Google Scholar] [CrossRef]
- Herrero, R.; Real, L.M.; Rivero-Juárez, A.; Pineda, J.A.; Camacho, Á.; Macías, J.; Laplana, M.; Konieczny, P.; Márquez, F.J.; Souto, J.C.; et al. Association of complement receptor 2 polymorphisms with innate resistance to HIV-1 infection. Genes Immun. 2015, 16, 134–141. [Google Scholar] [CrossRef]
- Meza, G.; Expósito, A.; Royo, J.L.; Ruiz-García, C.; Sánchez-Arcas, B.; Marquez, F.J.; Gómez-Vidal, M.A.; Omar, M.; Sinangil, F.; Higgins, K.; et al. Association of Complement C3d receptor 2 genotypes with the acquisition of HIV infection in a trial of recombinant glycoprotein 120 Vaccine. AIDS 2020, 34, 25–32. [Google Scholar] [CrossRef]
- Flynn, N.M.; Forthal, D.N.; Harro, C.D.; Judson, F.N.; Mayer, K.H.; Para, M.F. Placebo-controlled phase 3 trial of a recombinant glycoprotein 120 vaccine to prevent HIV-1 infection. J. Infect. Dis. 2005, 191, 654–665. [Google Scholar] [CrossRef]
- Gilbert, P.B.; Peterson, M.L.; Follmann, D.; Hudgens, M.G.; Francis, D.P.; Gurwith, M.; Heyward, W.L.; Jobes, D.V.; Popovic, V.; Self, S.G.; et al. Correlation between immunologic responses to a recombinant glycoprotein 120 vaccine and incidence of HIV-1 infection in a phase 3 HIV-1 preventive vaccine trial. J. Infect. Dis. 2005, 191, 666–677. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Knowles, D.; Humphrey, J.; Barbeira, A.; Dickinson, S.; Im, H.; Pritchard, J. Annotation-free quantification of RNA splicing using LeafCutter. Nat. Genet. 2018, 50, 151–158. [Google Scholar] [CrossRef]
- Sironi, M.; Biasin, M.; Gnudi, F.; Cagliani, R.; Saulle, I.; Forni, D.; Rainone, V.; Trabattoni, D.; Garziano, M.; Mazzotta, F.; et al. A regulatory polymorphism in HAVCR2 modulates susceptibility to HIV-1 infection. PLoS ONE 2014, 9, e106442. [Google Scholar] [CrossRef]
- Rallon, N.I.; Restrepo, C.; Naggie, S.; Lopez, M.; del Romero, J.; Goldstein, D.; McHutchison, J.; Soriano, V.; Benito, J.M. Interleukin-28B gene polymorphisms do not influence the susceptibility to HIV-infection or CD4 cell decline. AIDS 2011, 25, 269–271. [Google Scholar] [CrossRef]
- Biasin, M.; Sironi, M.; Saulle, I.; de Luca, M.; la Rosa, F.; Cagliani, R.; Forni, D.; Agliardi, C.; lo Caputo, S.; Mazzotta, F.; et al. Endoplasmic reticulum aminopeptidase 2 haplotypes play a role in modulating susceptibility to HIV infection. AIDS 2013, 27, 1697–1706. [Google Scholar] [CrossRef] [Green Version]
- Sironi, M.; Biasin, M.; Cagliani, R.; Gnudi, F.; Saulle, I.; Ibba, S.; Filippi, G.; Yahyaei, S.; Tresoldi, C.; Riva, S.; et al. Evolutionary analysis identifies an MX2 haplotype associated with natural resistance to HIV-1 infection. Mol. Biol. Evol. 2014, 31, 2402–2414. [Google Scholar] [CrossRef] [Green Version]
- Sironi, M.; Biasin, M.; Cagliani, R.; Forni, D.; De Luca, M.; Saulle, I.; Lo Caputo, S.; Mazzotta, F.; Macías, J.; Pineda, J.A.; et al. A common polymorphism in TLR3 confers natural resistance to HIV-1 infection. J. Immunol. 2012, 188, 818–823. [Google Scholar] [CrossRef] [Green Version]
- Herrero, R.; Pineda, J.A.; Rivero-Juarez, A.; Echbarthi, M.; Real, L.M.; Camacho, A.; Macias, J.; Fibla, J.; Rivero, A.; Caruz, A. Common haplotypes in CD209 promoter and susceptibility to HIV-1 infection intravenous drug users. Infect. Genet. Evol. 2016, 45, 20–25. [Google Scholar] [CrossRef]
- Jaimes-Bernal, C.; Rallón, N.; Benito, J.M.; Omar, M.; Gómez-Vidal, M.A.; Márquez, F.J.; Sánchez-Arcas, B.; Trujillo, M.; Royo, J.L.; Saulle, I.; et al. A knockout IFNL4 variant is associated with protection from sexually transmitted HIV-1 infection. J. Infect. Dis. 2018, 219, 772–776. [Google Scholar] [CrossRef]
- Feenstra, B.; Pasternak, B.; Geller, F.; Carstensen, L.; Wang, T.; Huang, F.; Eitson, J.L.; Hollegaard, M.V.; Svanström, H.; Vestergaard, M.; et al. Common variants associated with general and MMR vaccine-related febrile seizures. Nat. Genet. 2014, 46, 1274–1282. [Google Scholar] [CrossRef] [Green Version]
- Haralambieva, I.; Ovsyannikova, I.; Kennedy, R.; Larrabee, B.; Zimmermann, M.T.; Grill, D.; Schaid, D.; Poland, G. Genome-wide associations of CD46 and IFI44L genetic variants with neutralizing antibody response to measles vaccine. Hum. Genet. 2017, 136, 421–435. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Raffield, L.; Mousas, A.; Sakaue, S.; Huffman, J.; Moscati, A.; Trivedi, B.; Jiang, T.; Akbari, P.; Vuckovic, D.; et al. Trans-ethnic and ancestry-specific blood-cell genetics in 746,667 individuals from 5 global populations. Cell 2020, 182, 1198–1213. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
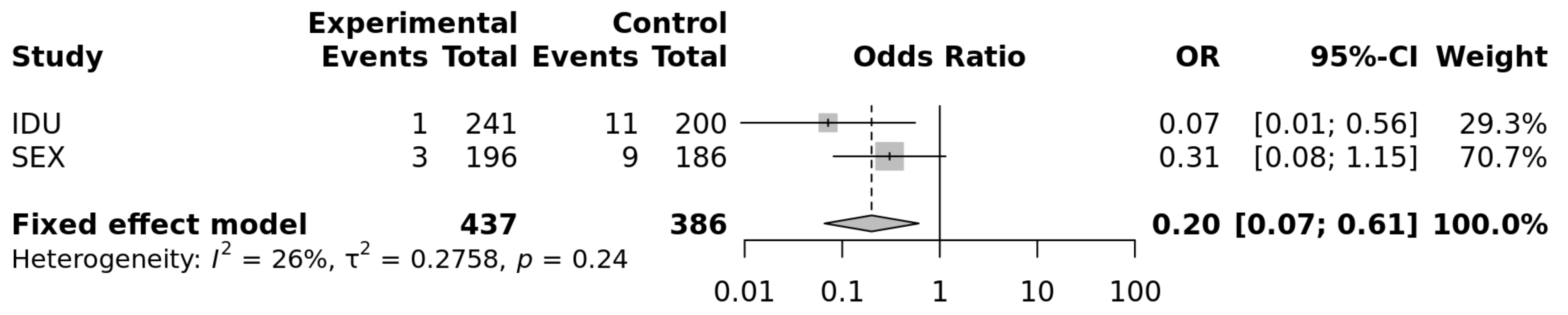
| IDU | Cohorts | Alleles a | Genotypes a | ||
|---|---|---|---|---|---|
| C | C/C | C/T | T/T | ||
| HIV-1, n (%) p c = 0.005 | 83 (17.2) | 1 (0.004) | 81 (34) | 159 (66) | |
| HESN, n (%) * p c = 0.3 | 83 (20.7) | 11 (6.0) | 65 (31) | 118 (64) | |
| Association test b | C vs. T | C/C vs. T/T | C/C vs. C/T | C/C vs. C/T + T/T | |
| OR (95% CI) | 0.83 (0.58–1.2) | 0.07 (0.009–0.5) | 0.06 (0.008–0.5) | 0.07 (0.009–0.5) | |
| p-value b | 0.18 | 0.001 | 0.001 | 0.001 | |
| Sex | Cohorts | Alleles a | Genotypes a | ||
| C | C/C | C/T | T/T | ||
| HIV-1, n (%) p c = 0.031 | 71 (18) | 3 (1.5) | 65 (33.1) | 128 (65.3) | |
| HESN, n (%) p c = 0.4 | 80 (22) | 9 (4.8) | 62 (3.3) | 108 (61) | |
| Association test b | C vs. T | C/C vs. T/T | C/C vs. C/T | C/C vs. C/T + T/T | |
| OR (95% CI) | 0.81 (0.5–1.2) | 0.32 (0.08–1.2) | 0.3 (0.07–1.2) | 0.31 (0.08–1.2) | |
| p-value b | 0.23 | 0.08 | 0.06 | 0.06 | |
| IDU + Sex | Cohorts | Alleles a | Genotypes a | ||
| C | C/C | C/T | T/T | ||
| HIV-1, n (%) p c = p = 0.001 | 154 (17.8) | 4 (0.01) | 146 (33) | 287 (65) | |
| HESN, n (%) p c = 0.3 | 163 (20.8) | 20 (5.1) | 123 (31) | 243 (62) | |
| Association test b | C vs. T | C/C vs. T/T | C/C vs. C/T | C/C vs. C/T + T/T | |
| OR (95% CI) | 0.8 (0.6–1.0) | 0.13 (0.03–0.04) | 0.13 (0.03–0.04) | 0.13 (0.03–0.04) | |
| p-value b | 0.07 | 0.0004 | 0.0003 | 0.0002 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Serrano-Rísquez, C.; Omar, M.; Gómez-Vidal, M.A.; Real, L.M.; Pineda, J.A.; Rivero, A.; Rivero-Juárez, A.; Forthal, D.; Márquez, F.J.; Lo Caputo, S.; et al. CD46 Genetic Variability and HIV-1 Infection Susceptibility. Cells 2021, 10, 3094. https://doi.org/10.3390/cells10113094
Serrano-Rísquez C, Omar M, Gómez-Vidal MA, Real LM, Pineda JA, Rivero A, Rivero-Juárez A, Forthal D, Márquez FJ, Lo Caputo S, et al. CD46 Genetic Variability and HIV-1 Infection Susceptibility. Cells. 2021; 10(11):3094. https://doi.org/10.3390/cells10113094
Chicago/Turabian StyleSerrano-Rísquez, Carmen, Mohamed Omar, María Amparo Gómez-Vidal, Luis Miguel Real, Juan Antonio Pineda, Antonio Rivero, Antonio Rivero-Juárez, Donald Forthal, Francisco J. Márquez, Sergio Lo Caputo, and et al. 2021. "CD46 Genetic Variability and HIV-1 Infection Susceptibility" Cells 10, no. 11: 3094. https://doi.org/10.3390/cells10113094