
Mitochondrial Dysfunction in Cardiorenal Syndrome 3: Renocardiac Effect of Vitamin C

 ,
,  , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
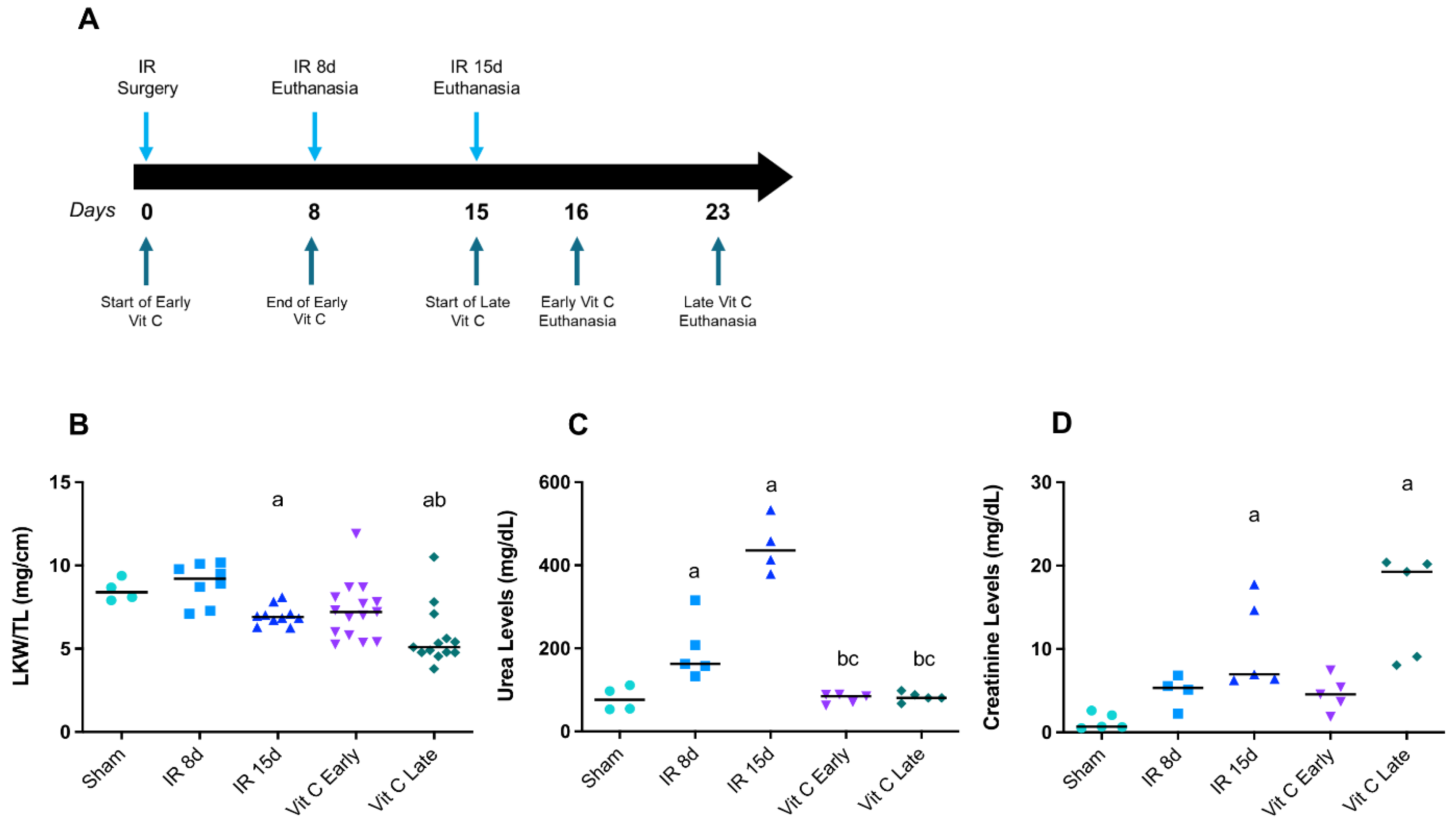
2.1. Experimental Animal Protocol
2.2. Biochemical Analyzes
2.3. RNA Isolation and Analysis of Gene Expression
2.4. Protein Extraction and Nitric Oxide Species Measurement
2.5. Evaluation of Mitochondrial Function
2.5.1. Preparation of Mitochondrial Fractions
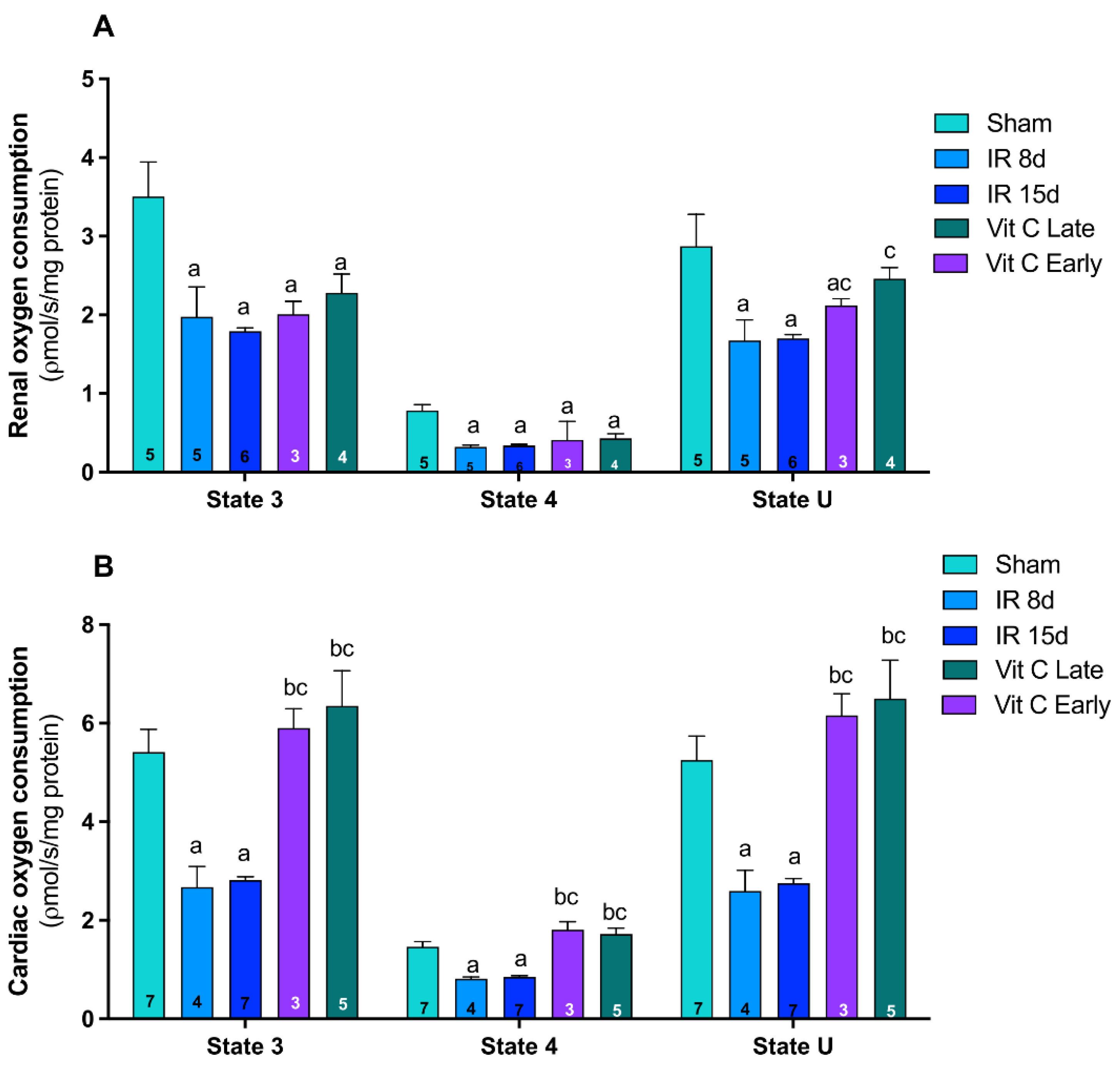
2.5.2. Mitochondrial Respiratory Parameters Determined by Oxygen Consumption
2.6. Statistical Analysis
3. Results
3.1. Vitamin C Preserved Renal Morphology and Function after IR Protocol
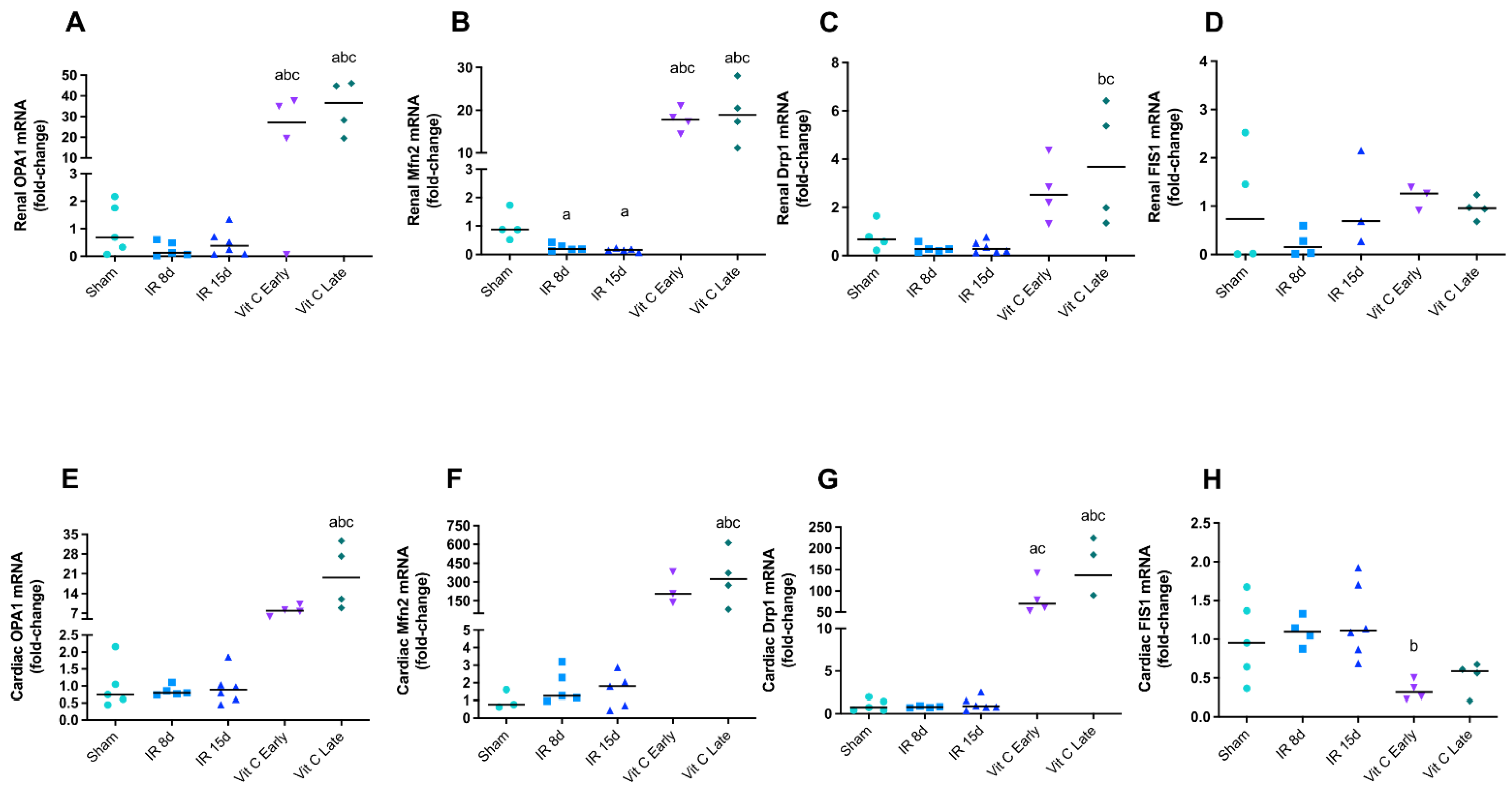
3.2. Vitamin C Modulated Mitochondrial Dynamics and Functioning after IR Protocol
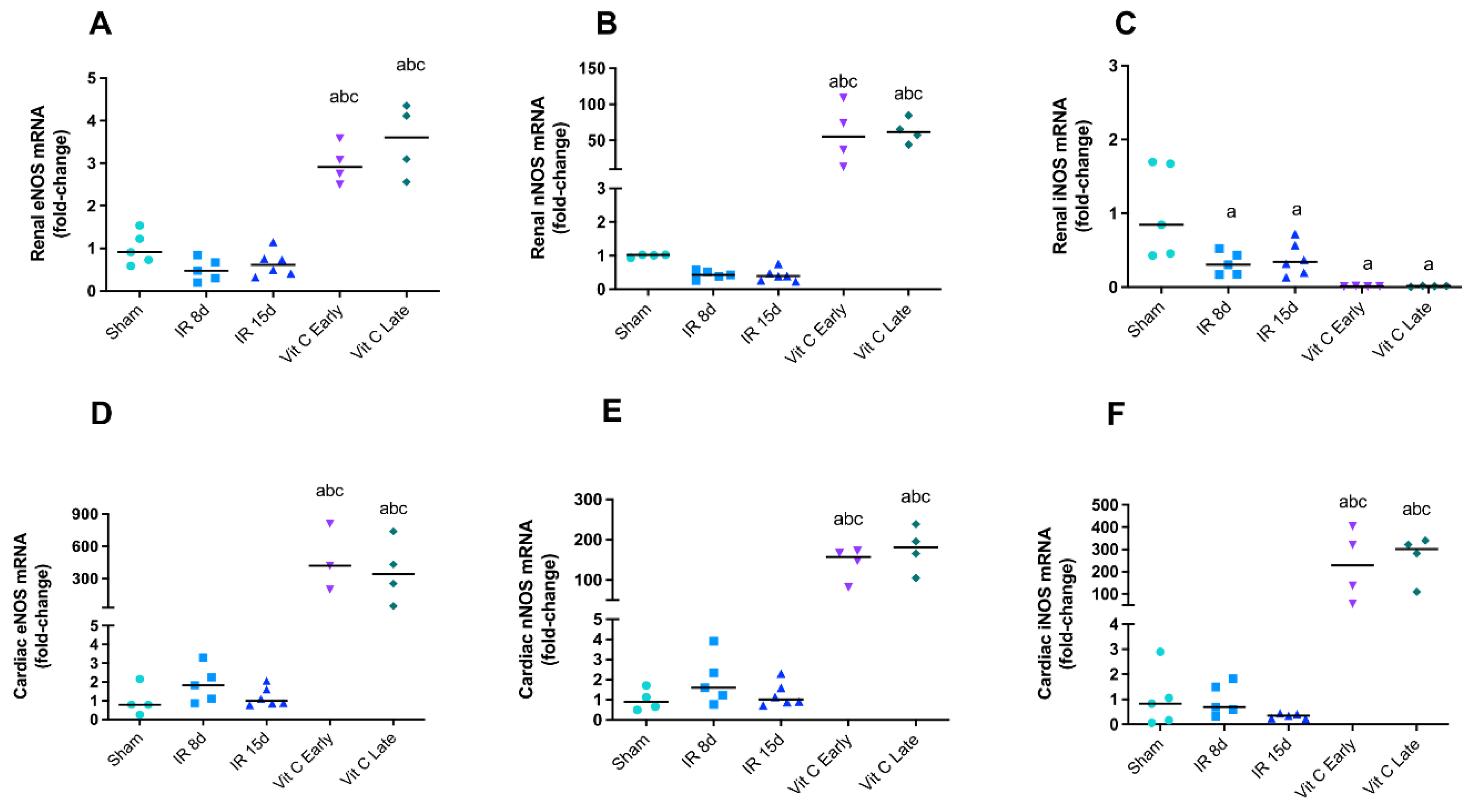
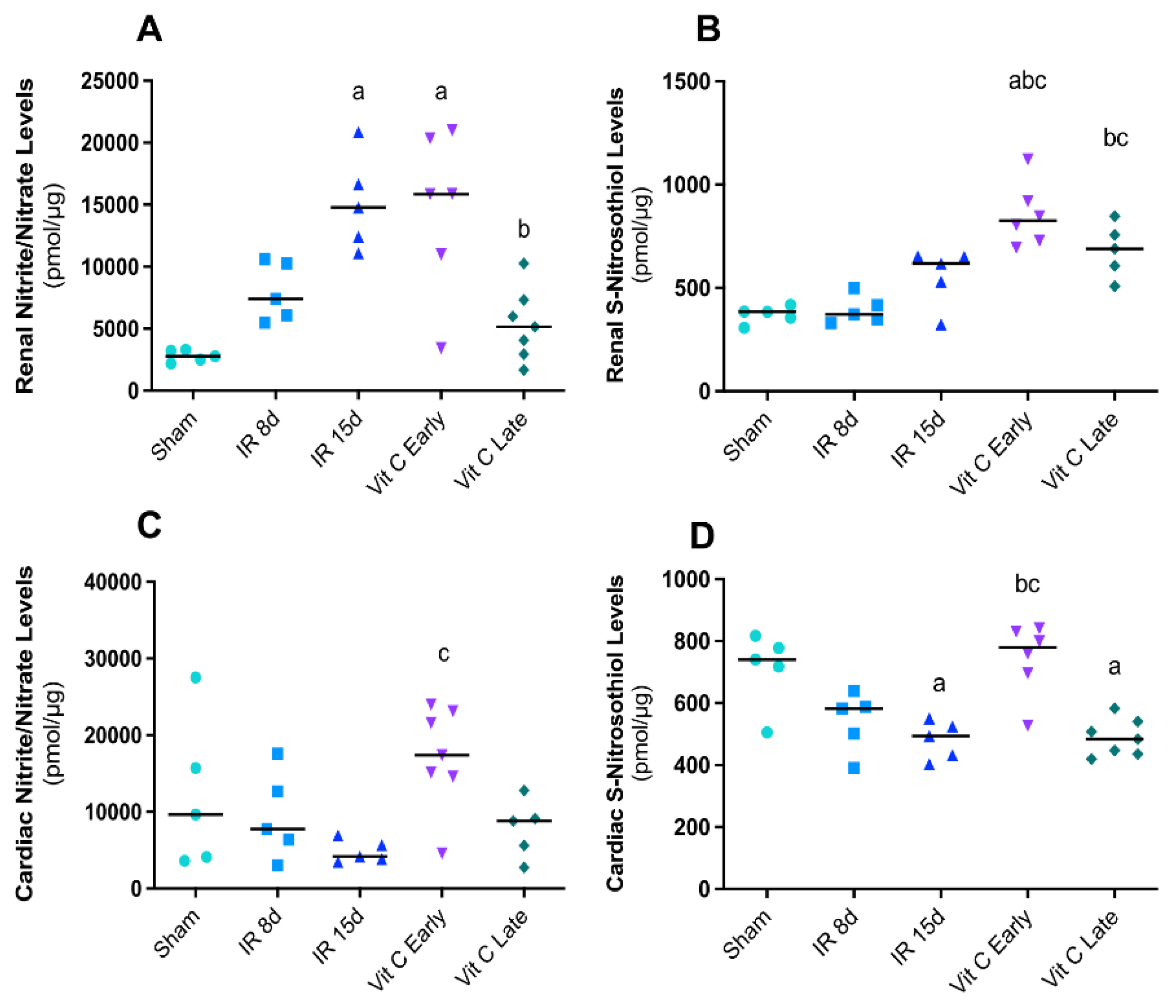
3.3. Vitamin C Modulated the Expression of Nitric Oxide Synthases Isoforms and Avoided Increase in Nitric Oxide Levels
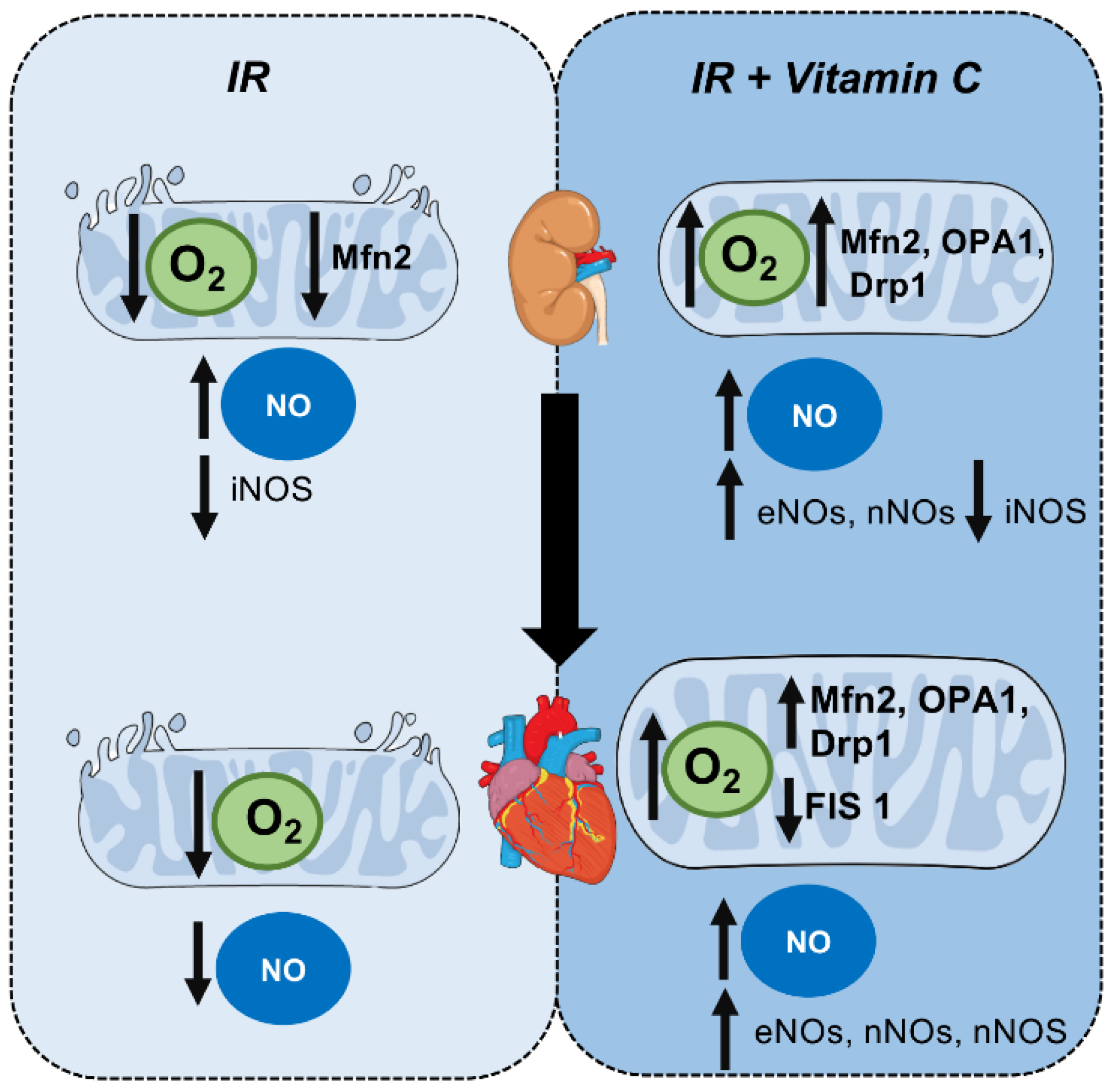
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Ronco, C.; McCullough, P.A.; Anker, S.D.; Anand, I.; Aspromonte, N.; Bagshaw, S.M.; Bellomo, R.; Berl, T.; Bobek, I.; Cruz, D.N.; et al. Cardiorenal syndromes: An executive summary from the consensus conference of the acute dialysis quality initiative (ADQI). Eur. Heart J. 2010, 31, 703–711. [Google Scholar] [CrossRef]
- Ronco, C.; House, A.A.; Haapio, M. Cardiorenal syndrome: Refining the definition of a complex symbosis gone wrong. Intensive Care Med. 2008, 34, 957–962. [Google Scholar] [CrossRef] [PubMed]
- Lullo, D.L.; Reeves, R.B.; Bellasi, A.; Ronco, C. Cardiorenal Syndrome in Acute Kidney Injury. Semin. Nephrol. 2019, 39, 31–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maulik, S.K.; Kumar, S. Oxidative stress and cardiac hypertrophy a review. Toxicol. Mech. Methods 2012, 22, 359–366. [Google Scholar] [CrossRef]
- Rocha, M.M.; Victor, V.M. Targeting antioxidants to mitochondria and cardiovascular diseases: The effects of mitoquinone. Med. Sci. Monit. 2007, 13, 132–145. [Google Scholar]
- Duann, P.; Lin, P.H. Mitochondria damage and kidney disease. Adv. Exp. Med. Biol. 2017, 982, 529–551. [Google Scholar] [CrossRef]
- Bhargava, P.; Schnellmann, R.G. Mitochondrial energetics in the kidney. Nat. Rev. Nephrol. 2017, 13, 629–646. [Google Scholar] [CrossRef] [PubMed]
- Emma, F.; Montini, G.; Parikh, S.M.; Salviati, L. Mitochondrial dysfunction in inherited renal disease and acute kidney injury. Nat. Rev. Nephrol. 2016, 12, 267–280. [Google Scholar] [CrossRef] [Green Version]
- Tang, C.; Cai, J.; Yin, X.; Weinberg, J.M.; Venkatachalam, M.A.; Dong, Z. Mitochondrial quality control in kidney injury and repair. Nat. Rev. Nephrol. 2021, 17, 299–318. [Google Scholar] [CrossRef]
- Palikaras, K.; Tavernarakis, N. Mitochondrial homeostasis: The interplay between mitophagy and mitochondrial biogenesis. Exp. Gerontol. 2014, 56, 182–188. [Google Scholar] [CrossRef]
- Pedro Bravo-San, J.M.; Kroemer, G.; Galluzzi, L. Autophagy and Mitophagy in Cardiovascular Disease. Circ. Res. 2017, 120, 1812–1824. [Google Scholar] [CrossRef] [PubMed]
- Zhan, M.; Brooks, C.; Liu, F.; Sun, L.; Dong, Z. Mitochondrial dynamics: Regulatory mechanisms and emerging role in renal pathophysiology. Int. Soc. Nephrol. 2013, 83, 568–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doi, K. Kidney–Heart Interactions in Acute Kidney Injury. Nephron 2016, 134, 141–144. [Google Scholar] [CrossRef]
- Doyle, M.P.; Hoekstra, J.W. Oxidation of nitrogen oxides by bound dioxygen in hemoproteins. J. Inorg. Biochem. 1984, 14, 351–358. [Google Scholar] [CrossRef]
- Hogg, N. The biochemistry and physiology of S-nitrosothiols. Annu. Rev. Pharmacol. Toxicol. 2002, 42, 585–600. [Google Scholar] [CrossRef]
- Stambler, J.S.; Jaraki, O.; Osborne, J.; Simon, D.I.; Keaney, J.; Vita, J.; Singel, D.; Valerei, C.R.; Loscalzo, J. Nitric oxide circulates in mammalian plasma primarily as an S-nitroso adduct of serum albumin. Proc. Natd. Acad. Sci. USA 1992, 89, 7674–7677. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, S.M.R.; Queiroz, J.H.; Pelúzo, M.C.G.; Costa, N.M.B.; Matta, S.L.P.; Queiroz, M.E.L.R. The formation and the effect of the reactive oxygen species in biological media. Biosci. J. 2015, 21, 133–149. [Google Scholar]
- Kim, H.K.; Nillius, B.; Nari, K.; Kyung, K.S.; Rhee, B.D.; Han, J. Cardiac response to oxidative stress induced by mitochondrial dysfunction. Rev. Physiol. Biochem. Pharmacol. 2016, 170, 101–127. [Google Scholar] [CrossRef]
- Ribeiro, D.A.; Buttros, J.B.; Oshima, C.T.F.; Bergamaschi, C.T.; Campos, R.R. Ascorbic acid prevents acute myocardial infarction induced by isoproterenol in rats: Role of inducible nitric oxide synthase production. J. Mol. Histol. 2009, 40, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Buttros, J.B.; Bergamaschi, C.T.; Ribeiro, D.A.; Fracalossi, A.C.C.; Campos, R.R. Cardioprotective Actions of Ascorbic Acid during Isoproterenol-Induced Acute Myocardial Infarction in Rats. Pharmacology 2009, 84, 29–37. [Google Scholar] [CrossRef]
- Dennis, J.M.; Witting, P.K. Protective Role for Antioxidants in Acute Kidney Disease. Nutrients 2017, 9, 718. [Google Scholar] [CrossRef] [Green Version]
- Du, J.; Cullena, J.I.; Buettner, G.R. Ascorbic acid: Chemistry, biology and the treatment of cancer. Biochim. Biophys. Acta 2012, 1826, 443–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caio-Silva, W.; Dias, D.S.; Junho, C.V.C.; Panico, P.; Neres-Santos, R.S.; Pelegrino, M.T.; Pieretti, J.C.; Seabra, A.B.; De Angelis, K.; Carneiro-Ramos, M.S. Characterization of the Oxidative Stress in Renal Ischemia/Reperfusion-Induced Cardiorenal Syndrome Type 3. Biomed. Res. Int. 2020, 2020, 1605358. [Google Scholar] [CrossRef] [PubMed]
- Trentin-Sonoda, M.; Silva, R.C.; Kmit, F.V.; Abrahão, M.V.; Cahli, G.M.; Brasil, G.V.; Muzi-Filho, H.; Silva, P.A.; Tovar-Moll, F.F.; Vieyra, A.; et al. Knockout of Toll-Like Receptors 2 and 4 Prevents Renal Ischemia-Reperfusion_Induced Cardiac Hypertrophy in Mice. PLoS ONE 2015, 10, e0139350. [Google Scholar] [CrossRef] [Green Version]
- Feitoza, C.Q. Inhibition of COX 1 and 2 prior to Renal Ischemia/Reperfusion Injury Decreases the Development of Fibrosis. Mol. Med. 2008, 724–730. [Google Scholar] [CrossRef]
- Trentin-Sonoda, M.; Fratoni, F.M.; Junho, C.V.C.; Silva, W.C.; Panico, K.; Carneiro-Ramos, M.S. Caspase-1 as Molecular Key in Cardiac Remodeling during Cardiorenal Syndrome Type 3 in the Murine Model. Curr. Mol. Med. 2019, 20, 72–78. [Google Scholar] [CrossRef]
- Zhu, Y.; Zhanga, Y.; Zhanga, J.; Zhanga, Y. Evaluation of Vitamin C Supplementation on Kidney Function and Vascular Reactivity Following Renal Ischemic Injury in Mice. Kidney Blood Press. Res. 2016, 41, 460–470. [Google Scholar] [CrossRef] [PubMed]
- Bell, J.P.; Mosfer, S.I.; Lang, D.; Donaldson, F.; Lewis, M.J. Vitamin C and quinapril abrogate LVH and endothelial dysfunction in aortic-banded guinea pigs. Am. J. Physiol.-Heart Circ. Physiol. 2001, 281, 1704–1710. [Google Scholar] [CrossRef]
- Koul, V.; Kaur, A.; Singh, A.P. Investigation of the role of nitric oxide/soluble guanylyl cyclase pathway in ascorbic acid-mediated protection against acute kidney injury in rats. Mol. Cell Biochem. 2015, 406, 1–7. [Google Scholar] [CrossRef]
- Haddad, P.S.; Santos, M.C.; Cassago, C.A.G.; Bernardes, J.S.; de Jesus, M.B.; Seabra, A.B. Synthesis, characterization, and cytotoxicity of glutathione-PEG-iron oxide magnetic nanoparticles. J. Nanopart. Res. 2016, 18, 369. [Google Scholar] [CrossRef]
- Zhang, X.; Cardosa, L.; Broderick, M.; Fein, H.; Davies, I.R. Novel Calibration Method for Nitric Oxide Microsensors by Stoichiometrical Generation of Nitric Oxide from SNAP. Electroanalysis 2000, 12, 425–428. [Google Scholar] [CrossRef]
- Mirandola, S.R.; Melo, D.R.; Saito, A.; Castilho, R.F. 3-Nitropropionic Acid-Induced Mitochondrial Permeability Transition: Omparative Study of Mitochondria from Different Tissues and Brain Regions. J. Neurosci. Res. 2010, 88, 630–639. [Google Scholar] [CrossRef] [PubMed]
- Gnaiger, E. Capacity of oxidative phosphorylation in human skeletal muscle: New perspectives of mitochondrial physiology. Int. J. Biochem. Cell Biol. 2009, 41, 1837–1845. [Google Scholar] [CrossRef] [PubMed]
- Chistiakov, D.A.; Shkurat, T.P.; Melnichenko, A.A.; Grechko, A.V.; Orekhov, A.N. The role of mitochondrial dysfunction in cardiovascular disease: A brief review. Ann. Med. 2017, 50, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Nourbakhsh, N.; Singh, S. Role of Renal Oxygenation and Mitochondrial Function in the Pathophysiology of Acute Kidney Injury. Nephron Clin. Pract. 2014, 127, 149–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, M.; Bai, M.; Lei, J.; Xie, Y.; Xu, S.; Jia, Z.; Zhang, A. Mitochondrial dysfunction and the AKI-to-CKD transition. Am. J. Physiol.-Ren. Physiol. 2020, 319, 1105–1116. [Google Scholar] [CrossRef] [PubMed]
- Doi, K.; Noiri, E. Mitochondrial Dysfunction in Cardiorenal Syndrome. Antioxid. Redox Signal. 2016, 25, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Cirino-Silva, R.; Kmit, F.V.; Trentin-Sonoda, M.; Nakama, K.K.; Panico, K.; Alvim, J.M.; Dreyer, T.R.; Martinho-Silva, H.; Carneiro-Ramos, M.S. Renal ischemia/reperfusion-induced cardiac hypertrophy in mice: Cardiac morphological and morphometric characterization. J. R. Soc. Med. Cardiovasc. Dis. 2017, 6, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strausser, S.A.; Nakano, D.; Souma, T. Acute kidney injury to chronic kidney disease transition: Insufficient cellular stress response. Curr. Opin. Nephrol. Hypertens. 2018, 27, 314–322. [Google Scholar] [CrossRef]
- Dong, Y.; Zhang, Q.; Wen, J.; Chen, T.; He, L.; Wang, Y.; Yin, J.; Wu, R.; Xue, R.; Li, S.; et al. Ischemic Duration and Frequency Determines AKI-to-CKD Progression Monitored by Dynamic Changes of Tubular Biomarkers in IRI Mice. Front. Physiol. 2019, 10, 153. [Google Scholar] [CrossRef]
- Shu, S.; Wang, Y.; Zheng, M.; Liu, Z.; Cai, J.; Tang, C.; Dong, Z. Hypoxia and Hypoxia-Inducible Factors in Kidney Injury and Repair. Cells 2019, 8, 207. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.P.; Singh, N.; Pathak, D.; Bedi, P.M.S. Estradiol attenuates ischemia reperfusion-induced acute kidney injury through PPAR-γ stimulated eNOS activation in rats. Mol. Cell. Biochem. 2019, 453, 1–9. [Google Scholar] [CrossRef]
- Hesketh, E.E.; Czopek, A.; Clay, M.; Borthwick, G.; Ferenbach, G.; Kluth, D.; Hughes, J. Renal ischaemia reperfusion injury: A mouse model of injury and regeneration. J. Vis. Exp. 2014, 88, e51816. [Google Scholar] [CrossRef] [Green Version]
- Sandhi, J.; Singh, J.P.; Kaur, T.; Ghuman, S.S.; Singh, A.P. Involvement of progesterone receptors in ascorbic acid mediated protection against ischemia-reperfusion induced acute kidney injury. J. Surg Res. 2014, 187, 278–288. [Google Scholar] [CrossRef] [PubMed]
- Duann, P.; Lianos, E.A.; Ma, J.; Lin, P. Autophagy, innate immunity and tissue repair in acute kidney injury. Int. J. Mol. Sci. 2016, 17, 662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, X.; Meng, X.; Xu, M.; Zhang, X.; Zhang, Y.; Ding, G.; Huang, S.; Zhang, A.; Jia, Z. Celastrol ameliorates cisplatin nephrotoxicity by inhibiting NF-κB and improving mitochondrial function. EBioMedicine 2018, 36, 266–280. [Google Scholar] [CrossRef] [Green Version]
- Long, Q.; Huang, L.; Huang, K.; Yang, Q. Assessing Mitochondrial Bioenergetics in Isolated Mitochondria from Mouse Heart Tissues Using Orobos 2k-Oxygraph. Methods Mol. Biol. 2019, 1966, 237–246. [Google Scholar] [CrossRef]
- Lindenmayer, G.E.; Sordahl, L.A.; Schwartz, A. Reevaluation of Oxidative Phosphorylation in Cardiac Mitochondria from Normal Animals and Animals in Heart Failure. Circ. Res. 1968, 23, 439–450. [Google Scholar] [CrossRef] [Green Version]
- Sordahl, L.A.; McCollum, W.B.; Wood, W.G.; Schwartz, A. Mitochondria and sarcoplasmic reticulum function in cardiac hypertrophy and failure. Am. J. Physiol. 1973, 224, 497–502. [Google Scholar] [CrossRef] [Green Version]
- Rosca, M.G.; Tandler, B.; Hoppe, C.L. Mitochondria in cardiac hypertrophy and heart failure. J. Mol. Cell. Cardiol. 2013, 55, 31–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowes, D.A.; Thottakam, B.M.V.; Webster, N.R.; Murphy, M.P.; Galley, H.F. The mitochondria-targeted antioxidant MitoQ protects against organ damage in a lipopolysaccharide–peptidoglycan model of sepsis. Free Radic. Biol. Med. 2008, 45, 1559–1565. [Google Scholar] [CrossRef] [PubMed]
- Popkov, V.A.; Silachev, D.N.; Zalevsky, A.O.; Zorov, D.B.; Plotnikov, E.V. Mitochondria as a Source and a Target for Uremic Toxins. Int. J. Mol. Sci. 2019, 20, 3094. [Google Scholar] [CrossRef] [Green Version]
- Bou-Teen, D.; Kaludercic, N.; Weissman, D.; Turan, B.; Maack, C.; Lisa, F.D.; Ruiz-Meana, M. Mitochondrial ROS and mitochondria-targeted antioxidants in the aged heart. Free Radic. Biol. Med. 2021, 167, 109–124. [Google Scholar] [CrossRef]
- Oyewole, A.O.; Birch-Machin, M.A. Mitochondria-targeted antioxidants. FASEB J. 2015, 29, 4766–4771. [Google Scholar] [CrossRef] [Green Version]
- Nan, J.; Zhu, W.; Rahman, M.S.; Liu, M.; Li, D.; Su, S.; Zhang, N.; Hu, X.; Yu, H.; Gupta, M.P.; et al. Molecular regulation of mitochondrial dynamics in cardiac disease. Biochim. Biophys. Acta 2017, 1864, 1260–1273. [Google Scholar] [CrossRef]
- Tan, Y.; Xia, F.; Li, L.; Peng, X.; Liu, W.; Zhang, Y.; Fang, H.; Zeng, Z.; Chen, Z. Novel Insights into the Molecular Features and Regulatory Mechanisms of Mitochondrial Dynamic Disorder in the Pathogenesis of Cardiovascular Disease. Oxidative Med. Cell. Longev. 2021, 2021, 6669075. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Hu, Y.; Quirós, P.M.; Wei, Q.; López-Otín, C.; Dong, Z. OMA1 mediates OPA1 proteolysis and mitochondrial fragmentation in experimental models of ischemic kidney injury. Am. J. Physiol.-Ren. Physiol. 2014, 306, 1318–1326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morigi, M.; Perico, L.; Rota, C.; Longaretti, L.; Conti, S.; Rottoli, D.; Novelli, T.; Remuzzi, G.; Benigni, A. Sirtuin 3–dependent mitochondrial dynamic improvements protect against acute kidney injury. J. Clin. Investig. 2015, 125, 715–726. [Google Scholar] [CrossRef] [Green Version]
- Long, R.; Peng, J.; Huang, L.; Jiang, G.; Liao, Y.; Sun, H.; Hu, Y.; Liao, X. Augmenter of Liver Regeneration Alleviates Renal Hypoxia-Reoxygenation Injury by Regulating Mitochondrial Dynamics in Renal Tubular Epithelial Cells. Mol. Cells 2019, 42, 893–905. [Google Scholar] [CrossRef]
- Noh, M.R.; Kim, K.Y.; Han, S.J.; Kim, J.I.; Kim, H.; Park, K.M. Methionine Sulfoxide Reductase A Deficiency Exacerbates Cisplatin-Induced Nephrotoxicity via Increased Mitochondrial Damage and Renal Cell Death. Antioxid. Redox Signal. 2017, 27, 727–741. [Google Scholar] [CrossRef]
- Wu, L.; Li, Q.; Liu, S.; An, X.; Huang, Z.; Zhang, B.; Yuan, Y.; Xing, C. Protective effect of hyperoside against renal ischemia-reperfusion injury via modulating mitochondrial fission, oxidative stress, and apoptosis. Free Radic. Res. 2019, 53, 727–736. [Google Scholar] [CrossRef]
- Perry, H.M.; Huang, L.; Wilson, R.J.; Bajwa, A.; Sesaki, H.; Yan, Z.; Rosin, D.L.; Kashatus, D.F.; Okusa, M.D. Dynamin-Related Protein 1 Deficiency Promotes Recovery from AKI. J. Am. Soc. Nephrol. 2017, 29, 194–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sumida, M.; Doi, K.; Ogasawara, E.; Yamashita, T.; Hamasaki, Y.; Kariya, T.; Takimoto, E.; Yahagi, N.; Nangaku, N.; Noiri, E. Regulation of Mitochondrial Dynamics by Dynamin_Related Protein-1 in Acute Cardiorenal Syndrome. J. Am. Soc. Nephrol. 2015, 26, 2378–2387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Funk, J.A.; Schnellmann, R.G. Persistent disruption of mitochondrial homeostasis after acute kidney injury. Am. J. Physiol.-Ren. Physiol. 2012, 302, 853–864. [Google Scholar] [CrossRef] [Green Version]
- Ortega-Domínguez, B.; Aparicio-Trejo, O.E.; García-Arroyo, F.E.; Leon-Contreras, J.C.; Tapia, E.; Molina-Jijon, E.; Hernandez-Pando, R.; Sanchez-Lozada, L.G.; Barrera-Oviedo, D.; Pedraza-Chaverri, J. Curcumin prevents cisplatin-induced renal alterations in mitochondrial bioenergetics and dynamic. Food Chem. Toxicol. 2017, 107, 373–385. [Google Scholar] [CrossRef]
- Rodríguez-Graciani, K.M.; Chapa-Dubocq, X.R.; MacMillan-Crow, L.A. Javadov, S. Association between L-OPA1 Cleavage and Cardiac Dysfunction. Cell. Physiol. Biochem. 2020, 54, 1101–1114. [Google Scholar] [CrossRef]
- Patten, D.; Harper, M.; Boardman, N. Harnessing the protective role of OPA1 in diabetic. Acta Physiol. 2020, 229, e13466. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Liu, Z.; Zhao, M.; Zhang, F.; Wang, K.; Feng, N.; Fu, F.; Li, J.; Li, J.; Liu, Y.; et al. κ-opioid receptor activation promotes mitochondrial fusion and enhances myocardial resistance to ischemia and reperfusion injury via STAT3-OPA1 pathway. Eur. J. Pharmacol. 2020, 874, 172987. [Google Scholar] [CrossRef]
- Knowlton, A.A.; Liu, T.T. Mitochondrial Dynamics and Heart Failure. Compr. Physiol. 2017, 6, 507–526. [Google Scholar] [CrossRef] [Green Version]
- Makino, A.; Suarez, J.; Gawlowski, T.; Han, W.; Wang, H.; Scott, B.T.; Dillmann, W.H. Regulation of mitochondrial morphology and function by O-GlcNAcylation in neonatal cardiac myocytes. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 300, 1296–1302. [Google Scholar] [CrossRef] [Green Version]
- Vásquez-Trincado, C.; García-Carvajal, I.; Pennanen, C.; Parra, V.; Hill, J.A.; Rothermel, B.A.; Lavandero, S. Mitochondrial dynamics, mitophagy and cardiovascular disease. J. Physiol. 2016, 594, 509–525. [Google Scholar] [CrossRef] [PubMed]
- Wai, T.; García-Pietro, J.; Baker, M.J.; Merkwirth, C.; Benit, P.; Rustin, P.; Rupérez, F.J.; Barbas, C.; Ibañez, B.; Langer, T. Imbalanced OPA1 processing and mitochondrial fragmentation cause heart failure in mice. Science 2015, 350, aad0116. [Google Scholar] [CrossRef] [PubMed]
- Liesa, M.; Borda-d’Água, B.; Medina-Gómez, G.; Lelliott, C.J.; Paz, J.C.; Rojo, M.; Palacín, M.; Vidal-Puig, A.; Zorzano, A. Mitochondrial Fusion Is Increased by the Nuclear Coactivator PGC-1b. PLoS ONE 2008, 10, e3613. [Google Scholar] [CrossRef] [Green Version]
- Haileselassie, B.; Mukherjee, R.; Joshi, A.U.; Napier, B.A.; Massis, L.M.; Ostberg, N.P.; Queliconi, B.B.; Monack, D.; Bernstein, D.; Mochly-Rosen, D. Drp1/Fis1 Interaction Mediates Mitochondrial Dysfunction in Septic Cardiomyopathy. J. Mol. Cell. Cardiol. 2019, 130, 160–169. [Google Scholar] [CrossRef]
- Aksu, U.; Demirci, C.; Ince, C. The Pathogenesis of Acute Kidney Injury and the Toxic Triangle of Oxygen, Reactive Oxygen Species and Nitric Oxide. Contrib. Nephrol. 2011, 174, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Bae, E.H.; Ma, S.K.; Kim, S.W. Altered Nitric Oxide System in Cardiovascular and Renal Diseases. Chonnam Med. J. 2016, 52, 81–90. [Google Scholar] [CrossRef] [Green Version]
- Kwon, O.; Hong, S.; Ramesh, G. Diminished NO generation by injured endothelium and loss of macula densa nNOS may contribute to sustained acute kidney injury after ischemia-reperfusion. Am. J. Physiol.-Ren. Physiol. 2009, 296, 25–33. [Google Scholar] [CrossRef] [Green Version]
- Ichinose, F.; Bloch, K.D.; Wu, J.C.; Hataishi, R.; Aretz, H.T.; Picard, M.H.; Scherrer-Crosbie, M. Pressure overload-induced LV hypertrophy and dysfunction in mice are exacerbated by congenital NOS3 deficiency. Am. J. Physiol.-Heart Circ. Physiol. 2004, 286, 1070–1075. [Google Scholar] [CrossRef]
- Yan, J.; Tie, G.; Messina, L.M. Tetrahydrobiopterin, L-arginine and vitamin C actsynergistically to decrease oxidative stress, increase nitricoxide and improve blood flow after induction of hindlimbischemia in the rat. Mol. Med. 2012, 18, 676–684. [Google Scholar] [CrossRef]
- Heller, R.; Unbehaun, A.; Schellenberg, S.; Mayer, B.; Werner-Felmayer, G.; Werner, E.R. L-Ascorbic Acid Potentiates Endothelial Nitric Oxide Synthesis via a Chemical Stabilization of Tetrahydrobiopterin. J. Biol. Chem. 2001, 276, 40–47. [Google Scholar] [CrossRef] [Green Version]
- Akolkar, G.; Dias, D.S.; Ayyappan, P.; Bagchi, A.K.; Jassal, D.S.; Salemi, V.M.C.; Irigoyen, M.C.; Angelis, K.; Singal, P.K. Vitamin C mitigates oxidative/nitrosative stress and inflammation in doxorubicin-induced cardiomyopathy. Am. J. Physiol.-Heart Circ. Physiol. 2017, 313, 795–809. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sense | Antisense |
|---|---|---|
| α-actin | GGCAAGATGAGAGTGCACAA | CGGAGAATGATGGTCCAGAT |
| ANF | ATCTGCCCTCTTGAAAAGCA | ACACACCACAAGGGCTTAGG |
| Cyclophilin A | AGCATACAGGTCCTGGCATC | AGCTGTCCACAGTCGGAAAT |
| nNOS | TCGATGCCAAGGCTATGTCC | CCTTGTAGCTCTTCCTCTCCTC |
| iNOS | GCTCTAGTGAAGCAAAGCCC | GGATTCTGGAACATTCTGTGCT |
| eNOS | CCCAGCCTCTCCAGCAC | GCCCATCCTGCTGAGCC |
| Mfn2 | AGGTTGAGGTGACAGCGTTC | CTCCACCTGTCCAAGCTTCA |
| OPA1 | TTAGAAAAGCCCTGCCCAGC | AGGTGAACCTGCAGTGAAGA |
| Drp1 | GCCTCAGATCGTCGTAGTGG | TCCATGTGGCAGGGTCATTT |
| FIS1 | CAGTGTTGCGTGTTAAGGGATG | TTCAAAATTCCTTGCAGCTTCGT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Neres-Santos, R.S.; Junho, C.V.C.; Panico, K.; Caio-Silva, W.; Pieretti, J.C.; Tamashiro, J.A.; Seabra, A.B.; Ribeiro, C.A.J.; Carneiro-Ramos, M.S. Mitochondrial Dysfunction in Cardiorenal Syndrome 3: Renocardiac Effect of Vitamin C. Cells 2021, 10, 3029. https://doi.org/10.3390/cells10113029
Neres-Santos RS, Junho CVC, Panico K, Caio-Silva W, Pieretti JC, Tamashiro JA, Seabra AB, Ribeiro CAJ, Carneiro-Ramos MS. Mitochondrial Dysfunction in Cardiorenal Syndrome 3: Renocardiac Effect of Vitamin C. Cells. 2021; 10(11):3029. https://doi.org/10.3390/cells10113029
Chicago/Turabian StyleNeres-Santos, Raquel Silva, Carolina Victoria Cruz Junho, Karine Panico, Wellington Caio-Silva, Joana Claudio Pieretti, Juliana Almeida Tamashiro, Amedea Barozzi Seabra, César Augusto João Ribeiro, and Marcela Sorelli Carneiro-Ramos. 2021. "Mitochondrial Dysfunction in Cardiorenal Syndrome 3: Renocardiac Effect of Vitamin C" Cells 10, no. 11: 3029. https://doi.org/10.3390/cells10113029