
Carbonic Anhydrase IV Selective Inhibitors Counteract the Development of Colitis-Associated Visceral Pain in Rats

 , , , , ,
, , , , ,  , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
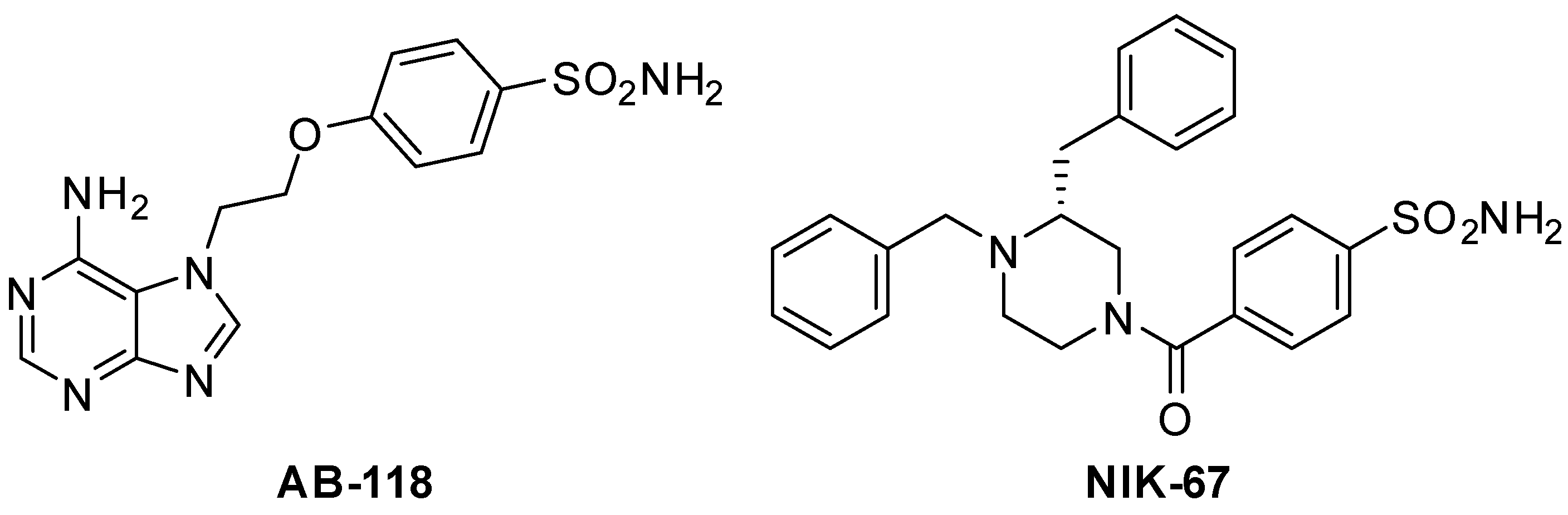
2.1. Chemistry
2.2. Synthesis of 4-(2-(6-Amino-7H-purin-7-yl)ethoxy)benzenesulfonamide AB-118
2.3. Animals
2.4. Induction of Colitis and Drug Administration
2.5. Assessment of Visceral Sensitivity by Viscero-Motor Response (VMR)
2.6. Assessment of Visceral Sensitivity by Abdominal Withdrawal Reflex (AWR)
2.7. Histological Analysis of Colonic Damage
2.8. Kinetic Determination of CA IV Activity in Colon Samples and AB-118 Inhibition Profile
2.9. Immunohistochemical Analysis of CA IV Expression
2.10. Statistics
3. Results
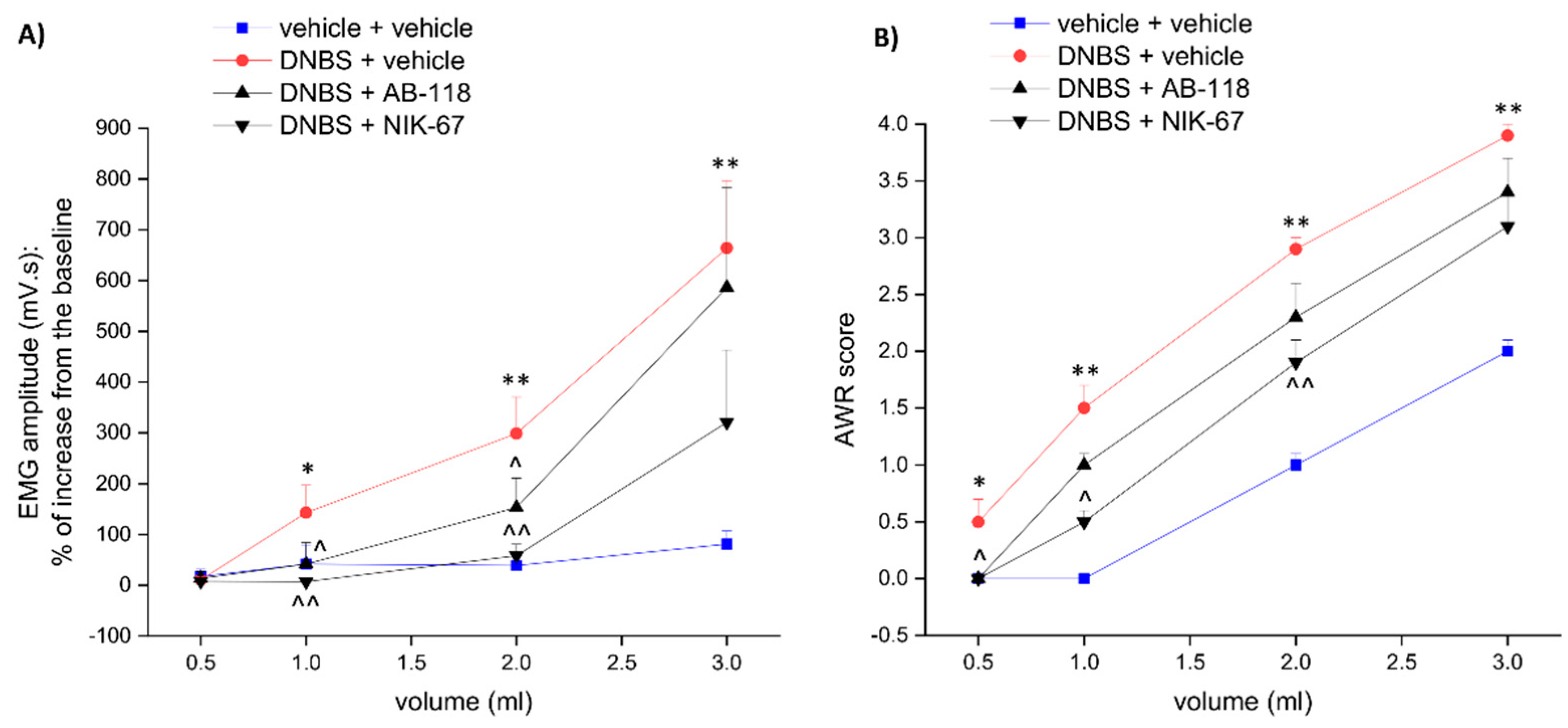
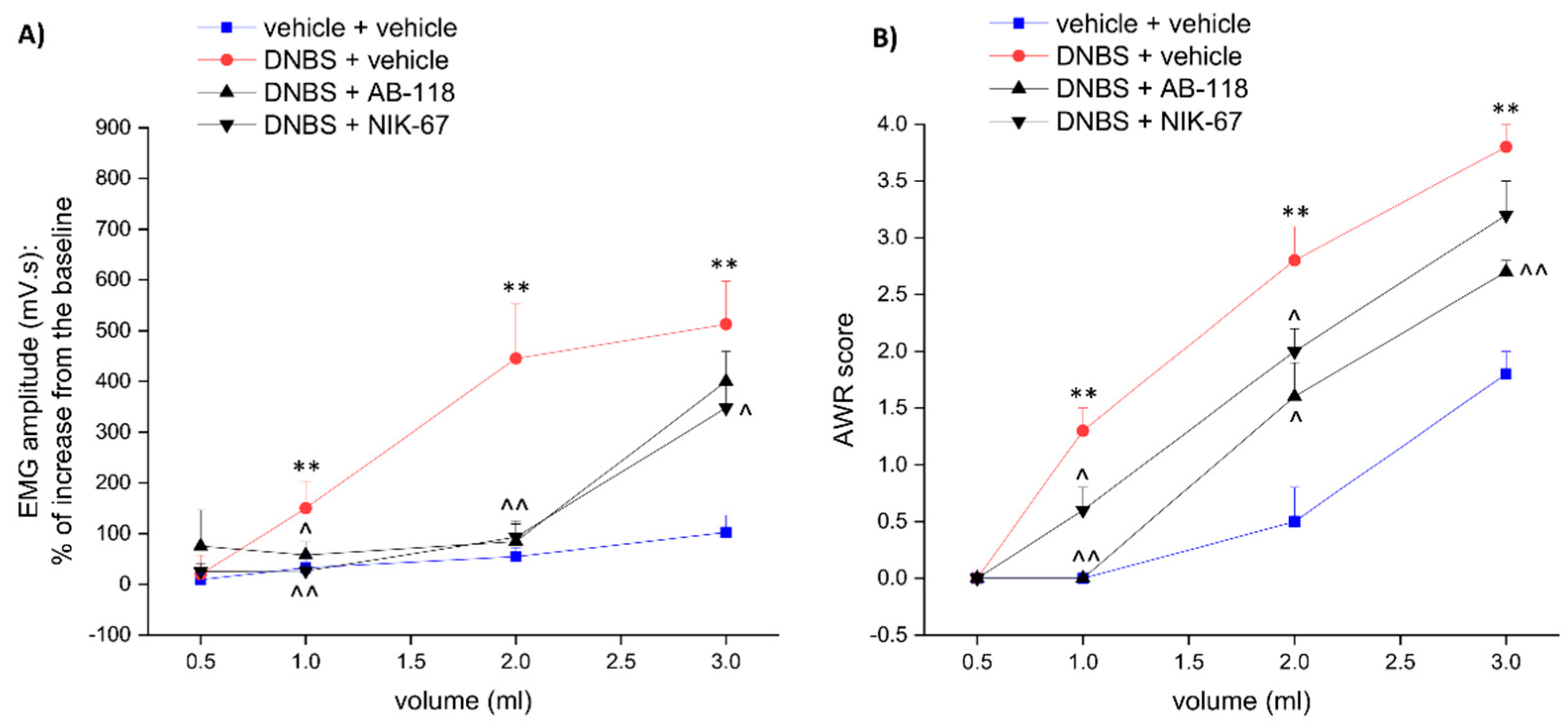
3.1. The CA IV Inhibitors AB-118 and NIK-67 Counteracted the Development of Visceral Pain Induced by Colitis in Rats
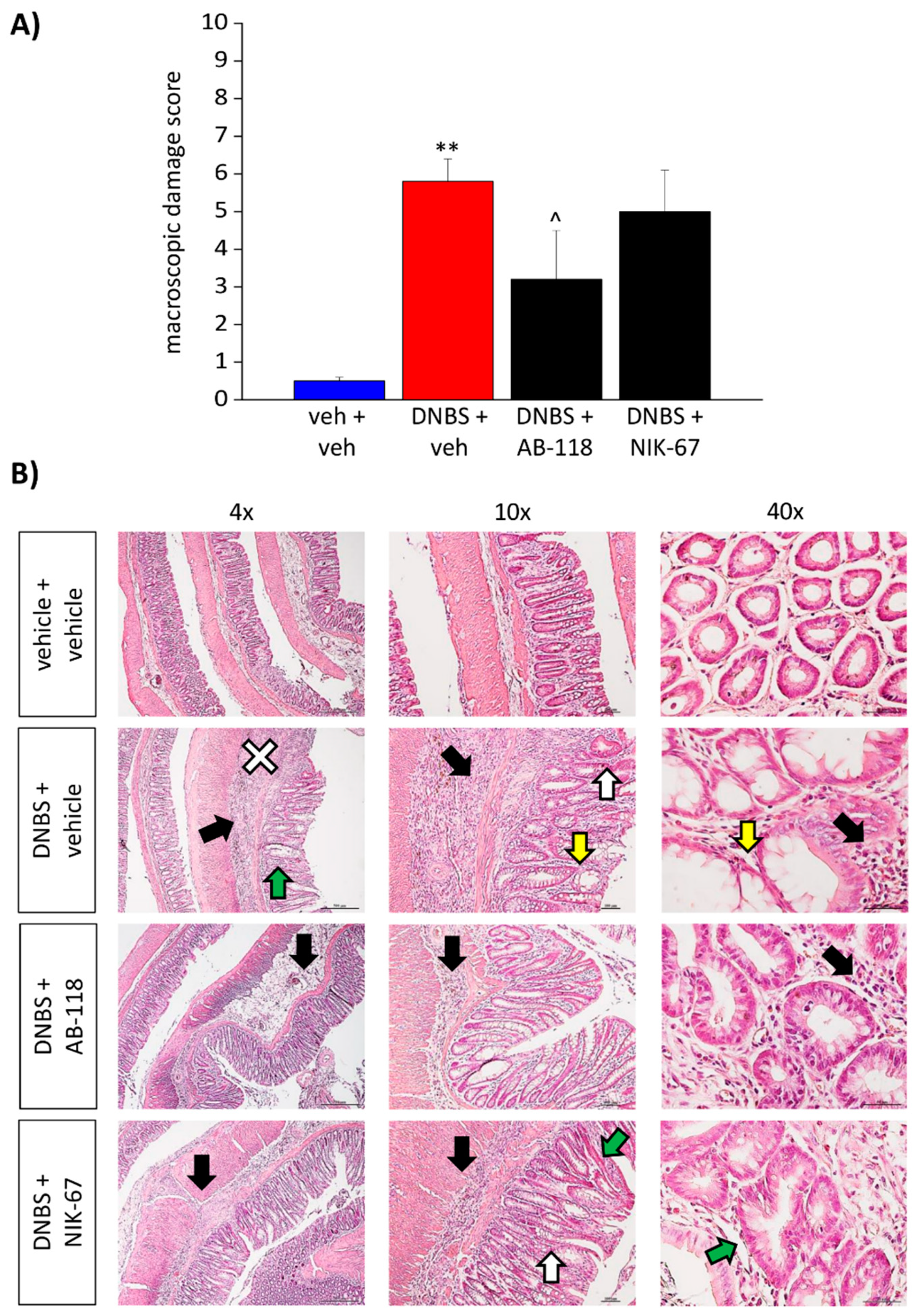
3.2. Effect of the Repeated Administration of AB2-118 and NIK-67 on Colon Damage
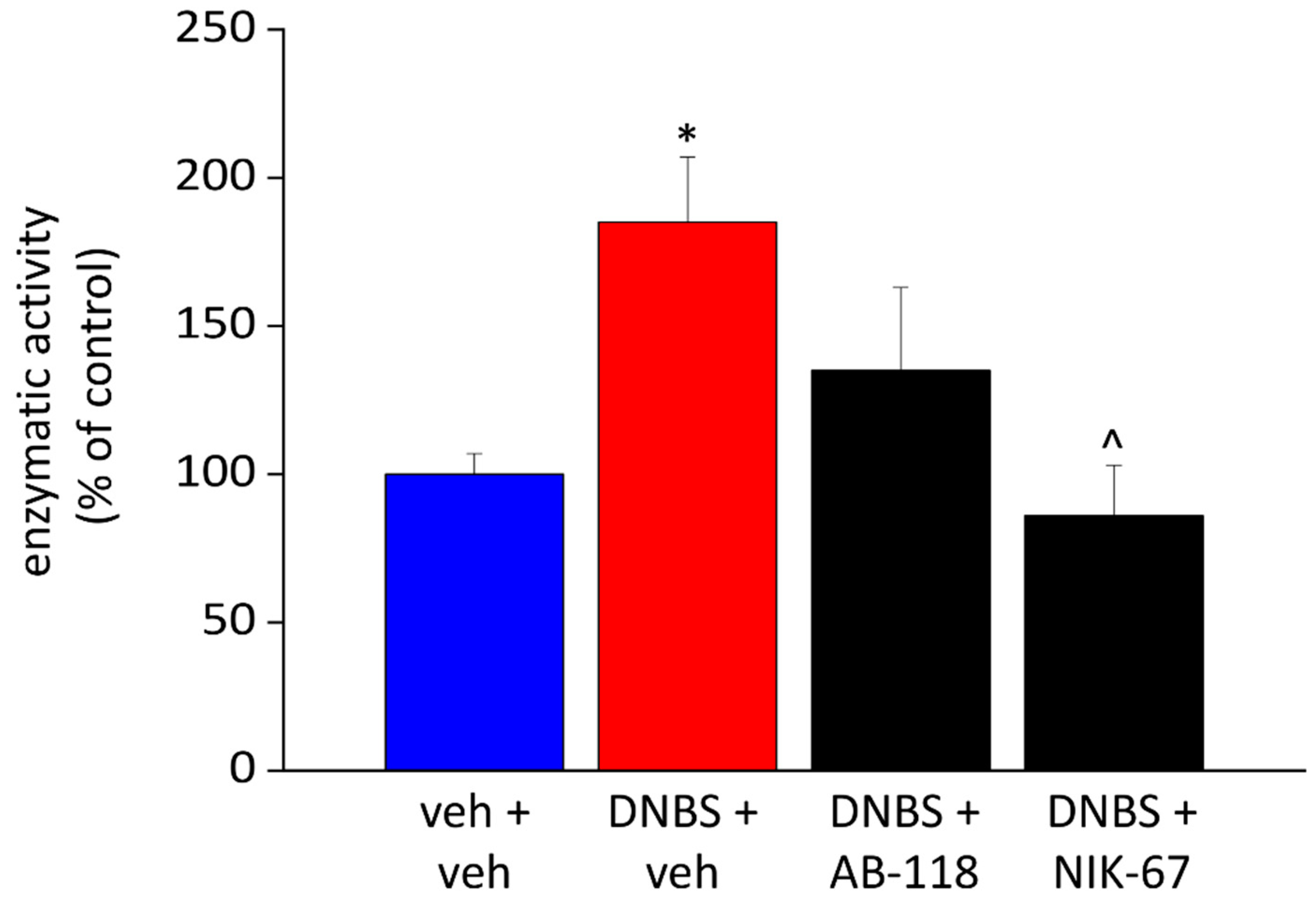
3.3. Effect of the Repeated Administration of AB2-118 and NIK-67 on Intestinal CA IV Activity
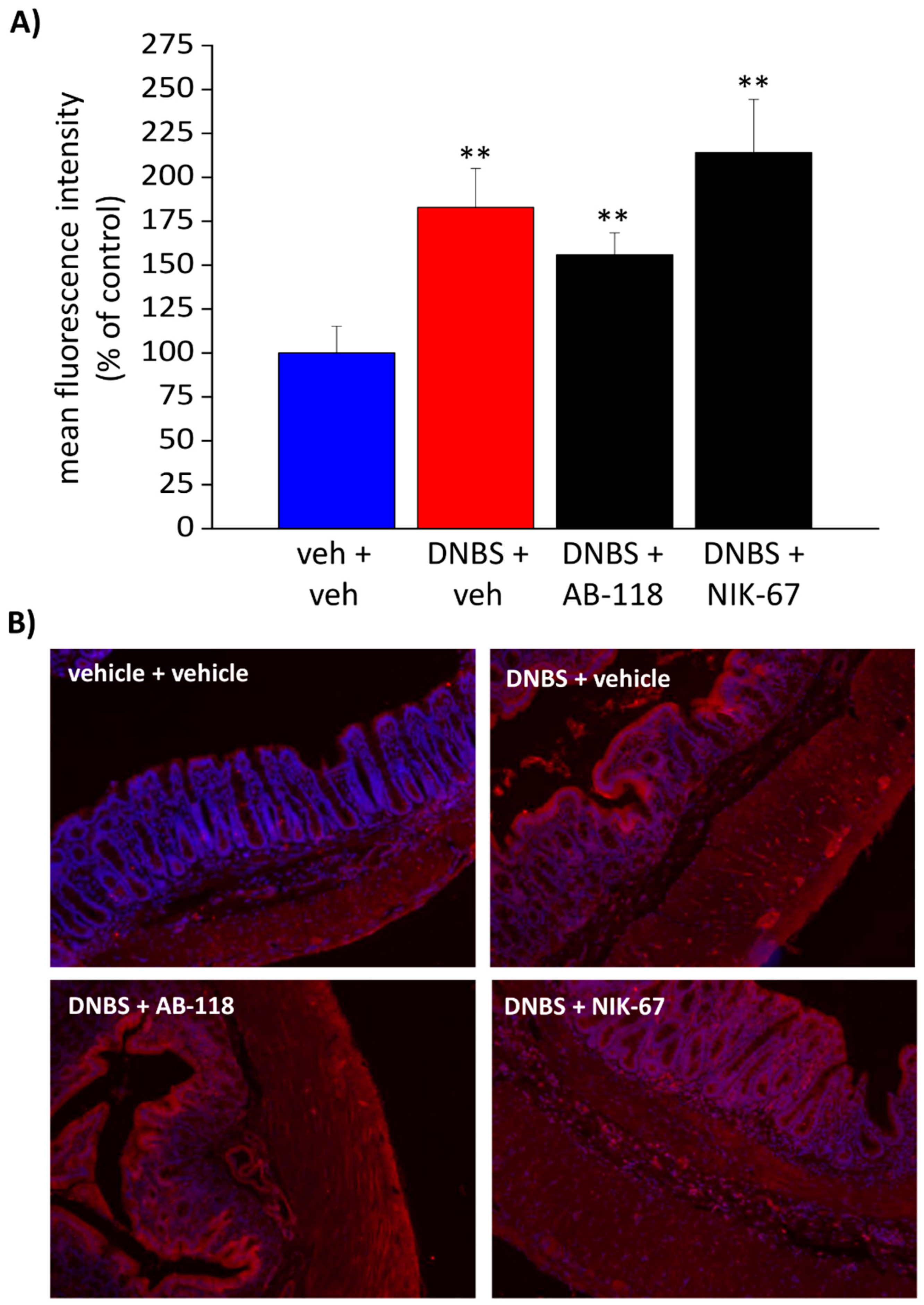
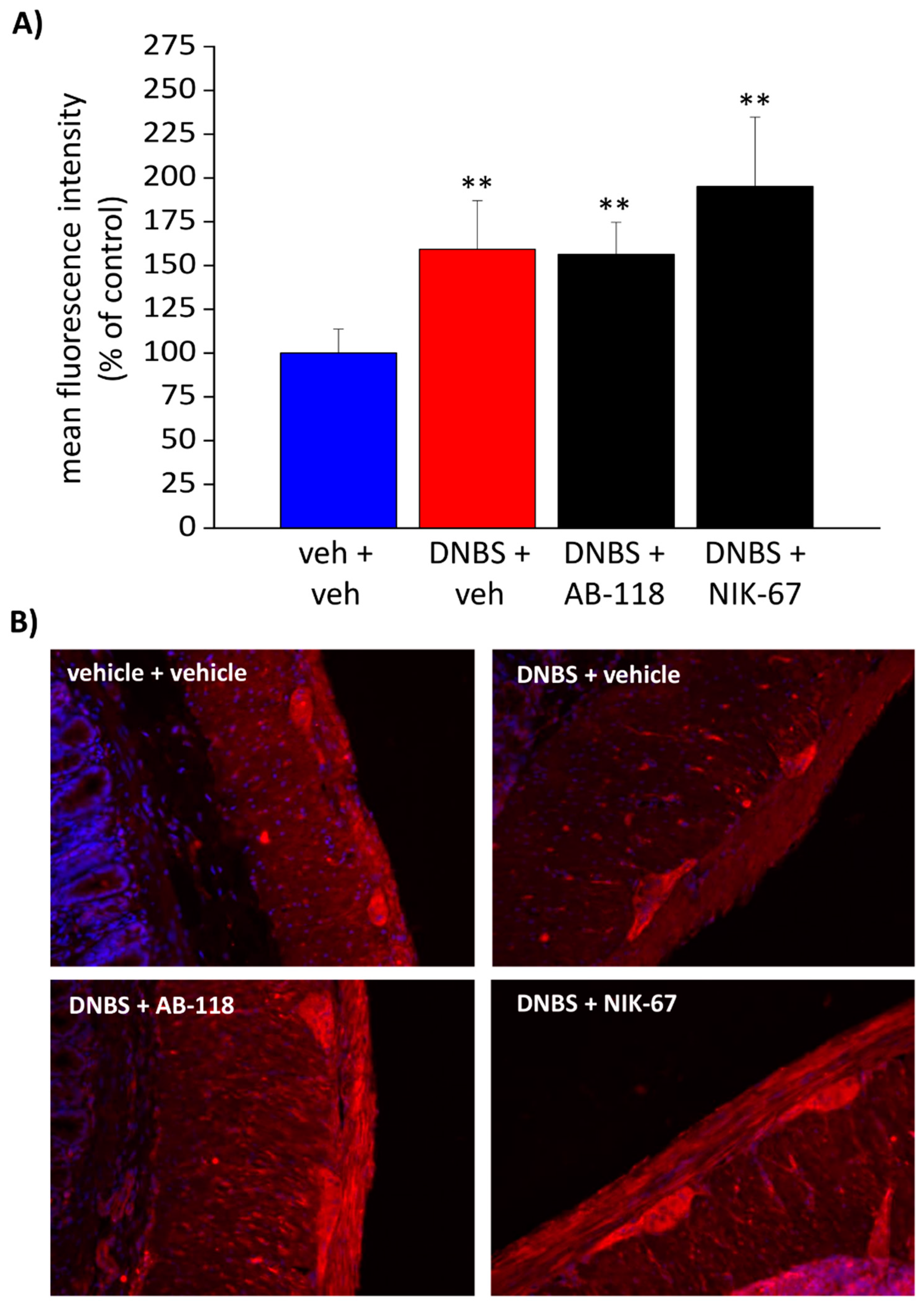
3.4. The Expression of CA IV Resulted Increased through the Colon Wall of DNBS-Treated Rats
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Srinath, A.I.; Walter, C.; Newara, M.C.; Szigethy, E.M. Pain management in patients with inflammatory bowel disease: Insights for the clinician. Ther. Adv. Gastroenterol. 2012, 5, 339–357. [Google Scholar] [CrossRef] [Green Version]
- Minderhoud, I.M.; Oldenburg, B.; Wismeijer, J.A.; Henegouwen, G.P.V.B.; Smout, A.J. IBS-like symptoms in patients with inflammatory bowel disease in remission; relationships with quality of life and coping behavior. Dig. Dis. Sci. 2004, 49, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Spiller, R.; Major, G. IBS and IBD—Separate entities or on a spectrum? Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 613–621. [Google Scholar] [CrossRef] [PubMed]
- Lucarini, E.; Parisio, C.; Branca, J.J.V.; Segnani, C.; Ippolito, C.; Pellegrini, C.; Antonioli, L.; Fornai, M.; Micheli, L.; Pacini, A.; et al. Deepening the Mechanisms of Visceral Pain Persistence: An Evaluation of the Gut-Spinal Cord Relationship. Cells 2020, 9, 1772. [Google Scholar] [CrossRef] [PubMed]
- Parisio, C.; Lucarini, E.; Micheli, L.; Toti, A.; Di Cesare Mannelli, L.; Antonini, G.; Panizzi, E.; Maidecchi, A.; Giovagnoni, E.; Lucci, J.; et al. Researching New Therapeutic Approaches for Abdominal Visceral Pain Treatment: Preclinical Effects of an Assembled System of Molecules of Vegetal Origin. Nutrients 2019, 12, 22. [Google Scholar] [CrossRef] [Green Version]
- Supuran, C.T. Carbonic anhydrase inhibition and the management of neuropathic pain. Expert Rev. Neurother. 2016, 16, 961–968. [Google Scholar] [CrossRef] [PubMed]
- Bua, S.; Lucarini, L.; Micheli, L.; Menicatti, M.; Bartolucci, G.; Selleri, S.; Di Cesare Mannelli, L.; Ghelardini, C.; Masini, E.; Carta, F.; et al. Bioisosteric Development of Multitarget Nonsteroidal Anti-Inflammatory Drug-Carbonic Anhydrases Inhibitor Hybrids for the Management of Rheumatoid Arthritis. J. Med. Chem. 2020, 63, 2325–2342. [Google Scholar] [CrossRef]
- Berrino, E.; Milazzo, L.; Micheli, L.; Vullo, D.; Angeli, A.; Bozdag, M.; Nocentini, A.; Menicatti, M.; Bartolucci, G.; di Cesare Mannelli, L.; et al. Synthesis and Evaluation of Carbonic Anhydrase Inhibitors with Carbon Monoxide Releasing Properties for the Management of Rheumatoid Arthritis. J. Med. Chem. 2019, 62, 7233–7249. [Google Scholar] [CrossRef]
- Micheli, L.; Bozdag, M.; Akgul, O.; Carta, F.; Guccione, C.; Bergonzi, M.C.; Bilia, A.R.; Cinci, L.; Lucarini, E.; Parisio, C.; et al. Pain Relieving Effect of-NSAIDs-CAIs Hybrid Molecules: Systemic and Intra-Articular Treatments against Rheumatoid Arthritis. Int. J. Mol. Sci. 2019, 20, 1923. [Google Scholar] [CrossRef]
- Asiedu, M.; Ossipov, M.H.; Kaila, K.; Price, T.J. Acetazolamide and midazolam act synergistically to inhibit neuropathic pain. Pain 2010, 148, 302–308. [Google Scholar] [CrossRef] [Green Version]
- Asiedu, M.N.; Mejia, G.L.; Hübner, C.A.; Kaila, K.; Price, T.J. Inhibition of carbonic anhydrase augments GABAA receptor-mediated analgesia via a spinal mechanism of action. J. Pain 2014, 15, 395–406. [Google Scholar] [CrossRef] [Green Version]
- Nocentini, A.; Alterio, V.; Bua, S.; Micheli, L.; Esposito, D.; Buonanno, M.; Bartolucci, G.; Osman, S.M.; ZA, A.L.; Cirilli, R.; et al. Phenyl(thio)phosphon(amid)ate Benzenesulfonamides as Potent and Selective Inhibitors of Human Carbonic Anhydrases II and VII Counteract Allodynia in a Mouse Model of Oxaliplatin-Induced Neuropathy. J. Med. Chem. 2020, 63, 5185–5200. [Google Scholar] [CrossRef]
- Angeli, A.; Di Cesare Mannelli, L.; Ghelardini, C.; Peat, T.S.; Bartolucci, G.; Menicatti, M.; Carta, F.; Supuran, C.T. Benzensulfonamides bearing spyrohydantoin moieties act as potent inhibitors of human carbonic anhydrases II and VII and show neuropathic pain attenuating effects. Eur. J. Med. Chem. 2019, 177, 188–197. [Google Scholar] [CrossRef]
- Angeli, A.; di Cesare Mannelli, L.; Lucarini, E.; Peat, T.S.; Ghelardini, C.; Supuran, C.T. Design, synthesis and X-ray crystallography of selenides bearing benzenesulfonamide moiety with neuropathic pain modulating effects. Eur. J. Med. Chem. 2018, 154, 210–219. [Google Scholar] [CrossRef] [PubMed]
- Carta, F.; Di Cesare Mannelli, L.; Pinard, M.; Ghelardini, C.; Scozzafava, A.; McKenna, R.; Supuran, C.T. A class of sulfonamide carbonic anhydrase inhibitors with neuropathic pain modulating effects. Bioorg. Med. Chem. 2015, 23, 1828–1840. [Google Scholar] [CrossRef]
- Henry, E.K.; Sy, C.B.; Inclan-Rico, J.M.; Espinosa, V.; Ghanny, S.S.; Dwyer, D.F.; Soteropoulos, P.; Rivera, A.; Siracusa, M.C. Carbonic anhydrase enzymes regulate mast cell-mediated inflammation. J. Exp. Med. 2016, 213, 1663–1673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Supuran, C.T. Carbonic anhydrases: Novel therapeutic applications for inhibitors and activators. Nat. Rev. Drug Discov. 2008, 7, 168–181. [Google Scholar] [CrossRef] [PubMed]
- Thiry, A.; Dogné, J.M.; Supuran, C.T.; Masereel, B. Carbonic anhydrase inhibitors as anticonvulsant agents. Curr. Top. Med. Chem. 2007, 7, 855–864. [Google Scholar] [CrossRef] [PubMed]
- Ruusuvuori, E.; Li, H.; Huttu, K.; Palva, J.M.; Smirnov, S.; Rivera, C.; Kaila, K.; Voipio, J. Carbonic anhydrase isoform VII acts as a molecular switch in the development of synchronous gamma-frequency firing of hippocampal CA1 pyramidal cells. J. Neurosci. 2004, 24, 2699–2707. [Google Scholar] [CrossRef]
- Radhakrishnan, R.; Sluka, K.A. Acetazolamide, a carbonic anhydrase inhibitor, reverses inflammation-induced thermal hyperalgesia in rats. J. Pharm. Exp. 2005, 313, 921–927. [Google Scholar] [CrossRef] [Green Version]
- Olausson, P.; Gerdle, B.; Ghafouri, N.; Larsson, B.; Ghafouri, B. Identification of proteins from interstitium of trapezius muscle in women with chronic myalgia using microdialysis in combination with proteomics. PLoS ONE 2012, 7, e52560. [Google Scholar] [CrossRef] [Green Version]
- Bala, I.; Bhatia, N.; Mishra, P.; Verma, G.R.; Kaman, L. Comparison of preoperative oral acetazolamide and intraperitoneal normal saline irrigation for reduction of postoperative pain after laparoscopic cholecystectomy. J. Laparoendosc. Adv. Surg. Tech. A 2015, 25, 285–290. [Google Scholar] [CrossRef]
- Sikandar, S.; Dickenson, A.H. Visceral pain: The ins and outs, the ups and downs. Curr. Opin. Support. Palliat Care 2012, 6, 17–26. [Google Scholar] [CrossRef]
- McGrath, J.C.; Lilley, E. Implementing guidelines on reporting research using animals (ARRIVE etc.): New requirements for publication in BJP. Br. J. Pharmacol. 2015, 172, 3189–3193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antonioli, L.; Fornai, M.; Colucci, R.; Ghisu, N.; Da Settimo, F.; Natale, G.; Kastsiuchenka, O.; Duranti, E.; Virdis, A.; Vassalle, C.; et al. Inhibition of adenosine deaminase attenuates inflammation in experimental colitis. J. Pharm. Exp. 2007, 322, 435–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Lin, C.; Tang, Y.; Chen, A.Q.; Liu, C.Y.; Lu, D.L. ZD 7288, an HCN channel blocker, attenuates chronic visceral pain in irritable bowel syndrome-like rats. World J. Gastroenterol. 2014, 20, 2091–2097. [Google Scholar] [CrossRef] [PubMed]
- Nocentini, A.; Ferraroni, M.; Carta, F.; Ceruso, M.; Gratteri, P.; Lanzi, C.; Masini, E.; Supuran, C.T. Benzenesulfonamides Incorporating Flexible Triazole Moieties Are Highly Effective Carbonic Anhydrase Inhibitors: Synthesis and Kinetic, Crystallographic, Computational, and Intraocular Pressure Lowering Investigations. J. Med. Chem. 2016, 59, 10692–10704. [Google Scholar] [CrossRef]
- Scozzafava, A.; Menabuoni, L.; Mincione, F.; Briganti, F.; Mincione, G.; Supuran, C.T. Carbonic anhydrase inhibitors. Synthesis of water-soluble, topically effective, intraocular pressure-lowering aromatic/heterocyclic sulfonamides containing cationic or anionic moieties: Is the tail more important than the ring? J. Med. Chem. 1999, 42, 2641–2650. [Google Scholar] [CrossRef] [PubMed]
- Wandernoth, P.M.; Raubuch, M.; Mannowetz, N.; Becker, H.M.; Deitmer, J.W.; Sly, W.S.; Wennemuth, G. Role of Carbonic Anhydrase IV in the Bicarbonate-Mediated Activation of Murine and Human Sperm. PLoS ONE 2010, 5, e15061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiaramonte, N.; Bua, S.; Ferraroni, M.; Nocentini, A.; Bonardi, A.; Bartolucci, G.; Durante, M.; Lucarini, L.; Chiapponi, D.; Dei, S.; et al. 2-Benzylpiperazine: A new scaffold for potent human carbonic anhydrase inhibitors. Synthesis, enzyme inhibition, enantioselectivity, computational and crystallographic studies and in vivo activity for a new class of intraocular pressure lowering agents. Eur. J. Med. Chem. 2018, 151, 363–375. [Google Scholar] [CrossRef]
- Khalifah, R.G. The carbon dioxide hydration activity of carbonic anhydrase. I. Stop-flow kinetic studies on the native human isoenzymes B and C. J. Biol. Chem. 1971, 246, 2561–2573. [Google Scholar] [CrossRef]
- Purkerson, J.M.; Schwartz, G.J. The role of carbonic anhydrases in renal physiology. Kidney Int. 2007, 71, 103–115. [Google Scholar] [CrossRef] [Green Version]
- Fleming, R.E.; Parkkila, S.; Parkkila, A.K.; Rajaniemi, H.; Waheed, A.; Sly, W.S. Carbonic anhydrase IV expression in rat and human gastrointestinal tract regional, cellular, and subcellular localization. J. Clin. Investig. 1995, 96, 2907–2913. [Google Scholar] [CrossRef] [Green Version]
- Vergnolle, N. The enteric nervous system in inflammation and pain: The role of proteinase-activated receptors. Can. J. Gastroenterol. 2003, 17, 589–592. [Google Scholar] [CrossRef]
- Drewes, A.M.; Olesen, A.E.; Farmer, A.D.; Szigethy, E.; Rebours, V.; Olesen, S.S. Gastrointestinal pain. Nat. Rev. Dis. Primers 2020, 6, 1. [Google Scholar] [CrossRef]
- Lardner, A. The effects of extracellular pH on immune function. J. Leukoc. Biol. 2001, 69, 522–530. [Google Scholar] [PubMed]
- Khan, I.; Khan, K. Uncoupling of Carbonic Anhydrase from Na-H exchanger-1 in Experimental Colitis: A Possible Mechanistic Link with Na-H Exchanger. Biomolecules 2019, 9, 700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hudalla, H.; Michael, Z.; Christodoulou, N.; Willis, G.R.; Fernandez-Gonzalez, A.; Filatava, E.J.; Dieffenbach, P.; Fredenburgh, L.E.; Stearman, R.S.; Geraci, M.W.; et al. Carbonic Anhydrase Inhibition Ameliorates Inflammation and Experimental Pulmonary Hypertension. Am. J. Respir. Cell Mol. Biol. 2019, 61, 512–524. [Google Scholar] [CrossRef] [PubMed]
- Mizoguchi, E.; Xavier, R.J.; Reinecker, H.C.; Uchino, H.; Bhan, A.K.; Podolsky, D.K.; Mizoguchi, A. Colonic epithelial functional phenotype varies with type and phase of experimental colitis. Gastroenterology 2003, 125, 148–161. [Google Scholar] [CrossRef]
- Potenzieri, A.; Riva, B.; Rigolio, R.; Chiorazzi, A.; Pozzi, E.; Ballarini, E.; Cavaletti, G.; Genazzani, A.A. Oxaliplatin-induced neuropathy occurs through impairment of haemoglobin proton buffering and is reversed by carbonic anhydrase inhibitors. Pain 2020, 161, 405–415. [Google Scholar] [CrossRef] [PubMed]
- Cho, C.H.; Pfeiffer, C.J. Study of the damaging effects of acetazolamide on gastric mucosa in rats. Acta Physiol. Hung. 1984, 64, 279–285. [Google Scholar] [PubMed]
- Johansson, M.E.; Sjövall, H.; Hansson, G.C. The gastrointestinal mucus system in health and disease. Nat. Rev. Gastroenterol. Hepatol 2013, 10, 352–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorofeyev, A.E.; Vasilenko, I.V.; Rassokhina, O.A.; Kondratiuk, R.B. Mucosal barrier in ulcerative colitis and Crohn’s disease. Gastroenterol Res. Pract. 2013, 2013, 431231. [Google Scholar] [CrossRef] [PubMed]
- Supuran, C.T.; Capasso, C. An Overview of the Bacterial Carbonic Anhydrases. Metabolites 2017, 7, 56. [Google Scholar] [CrossRef] [Green Version]
- Kivelä, A.-J.; Kivelä, J.; Saarnio, J.; Parkkila, S. Carbonic anhydrases in normal gastrointestinal tract and gastrointestinal tumours. World J. Gastroenterol. 2005, 11, 155–163. [Google Scholar] [CrossRef]
- Luczynski, P.; Tramullas, M.; Viola, M.; Shanahan, F.; Clarke, G.; O’Mahony, S.; Dinan, T.G.; Cryan, J.F. Microbiota regulates visceral pain in the mouse. Elife 2017, 6, e25887. [Google Scholar] [CrossRef]
- Esquerre, N.; Basso, L.; Defaye, M.; Vicentini, F.A.; Cluny, N.; Bihan, D.; Hirota, S.A.; Schick, A.; Jijon, H.B.; Lewis, I.A.; et al. Colitis-Induced Microbial Perturbation Promotes Postinflammatory Visceral Hypersensitivity. Cell Mol. Gastroenterol. Hepatol. 2020, 10, 225–244. [Google Scholar] [CrossRef]
- Sun, M.; Dahl, D.; Alkon, D.L. Heterosynaptic transformation of GABAergic gating in the hippocampus and effects of carbonic anhydrase inhibition. J. Pharm. Exp. 2001, 296, 811–817. [Google Scholar]
- Sun, M.K.; Zhao, W.Q.; Nelson, T.J.; Alkon, D.L. Theta rhythm of hippocampal CA1 neuron activity: Gating by GABAergic synaptic depolarization. J. Neurophysiol. 2001, 85, 269–279. [Google Scholar] [CrossRef]
- Loeza-Alcocer, E.; McPherson, T.P.; Gold, M.S. Peripheral GABA receptors regulate colonic afferent excitability and visceral nociception. J. Physiol. 2019, 597, 3425–3439. [Google Scholar] [CrossRef]
- Wang, B.; Samanani, N.; Roth, S.H.; Archer, D.P. Spinal carbonic anhydrase contributes to nociceptive reflex enhancement by midazolam, pentobarbital, and propofol. Anesthesiology 2003, 98, 921–927. [Google Scholar] [CrossRef]
- Lee, K.Y.; Prescott, S.A. Chloride dysregulation and inhibitory receptor blockade yield equivalent disinhibition of spinal neurons yet are differentially reversed by carbonic anhydrase blockade. Pain 2015, 156, 2431–2437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.Y.; Ratté, S.; Prescott, S.A. Excitatory neurons are more disinhibited than inhibitory neurons by chloride dysregulation in the spinal dorsal horn. Elife 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.; Qian, A.H.; Song, D.D.; Ben, Q.W.; Yao, W.Y.; Sun, J.; Li, W.G.; Xu, T.L.; Yuan, Y.Z. Role of the potassium chloride cotransporter isoform 2-mediated spinal chloride homeostasis in a rat model of visceral hypersensitivity. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 308, G767–G778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Yu, J.; Kannampalli, P.; Nie, L.; Meng, H.; Medda, B.K.; Shaker, R.; Sengupta, J.N.; Banerjee, B. MicroRNA-mediated downregulation of potassium-chloride-cotransporter and vesicular γ-aminobutyric acid transporter expression in spinal cord contributes to neonatal cystitis-induced visceral pain in rats. Pain 2017, 158, 2461–2474. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | KI (nM) [31]. | |||
|---|---|---|---|---|
| CA I | CA II | CA IV | CA IX | |
| NIK-67 [30] | 380.2 | 60.7 | 2.3 | 418.5 |
| AB-118 | 783.7 | 174.3 | 28.7 | 87.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lucarini, E.; Nocentini, A.; Bonardi, A.; Chiaramonte, N.; Parisio, C.; Micheli, L.; Toti, A.; Ferrara, V.; Carrino, D.; Pacini, A.; et al. Carbonic Anhydrase IV Selective Inhibitors Counteract the Development of Colitis-Associated Visceral Pain in Rats. Cells 2021, 10, 2540. https://doi.org/10.3390/cells10102540
Lucarini E, Nocentini A, Bonardi A, Chiaramonte N, Parisio C, Micheli L, Toti A, Ferrara V, Carrino D, Pacini A, et al. Carbonic Anhydrase IV Selective Inhibitors Counteract the Development of Colitis-Associated Visceral Pain in Rats. Cells. 2021; 10(10):2540. https://doi.org/10.3390/cells10102540
Chicago/Turabian StyleLucarini, Elena, Alessio Nocentini, Alessandro Bonardi, Niccolò Chiaramonte, Carmen Parisio, Laura Micheli, Alessandra Toti, Valentina Ferrara, Donatello Carrino, Alessandra Pacini, and et al. 2021. "Carbonic Anhydrase IV Selective Inhibitors Counteract the Development of Colitis-Associated Visceral Pain in Rats" Cells 10, no. 10: 2540. https://doi.org/10.3390/cells10102540