
Spatio-Temporal Multiscale Analysis of Western Diet-Fed Mice Reveals a Translationally Relevant Sequence of Events during NAFLD Progression

 , , , , , , , , , , , , , and
, , , , , , , , , , , , , and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Mice and Feeding Style
2.2. Induction of Acute-on-Chronic Liver Injury by Acetaminophen
2.3. Induction of Acute-on-Chronic Liver Injury by LPS
2.4. Intravital Imaging
2.5. Magnetic Resonance Imaging (MRI)
2.5.1. Tumor Detection
2.5.2. Estimation of the Fat Fraction
2.5.3. Assessment of Hepatocyte Uptake Capacity
2.6. Sample Collection
2.7. Liver Enzyme Assay
2.8. Histopathology, Immunohistochemistry, and TUNEL Staining
2.9. RNA-Seq Analysis
2.10. Bioinformatics
2.11. Western Blot Analysis and Quantification
2.12. Quantification of Plasma Metabolites
2.13. Image Analysis
2.14. Patients
2.15. Statistical Analysis
3. Results
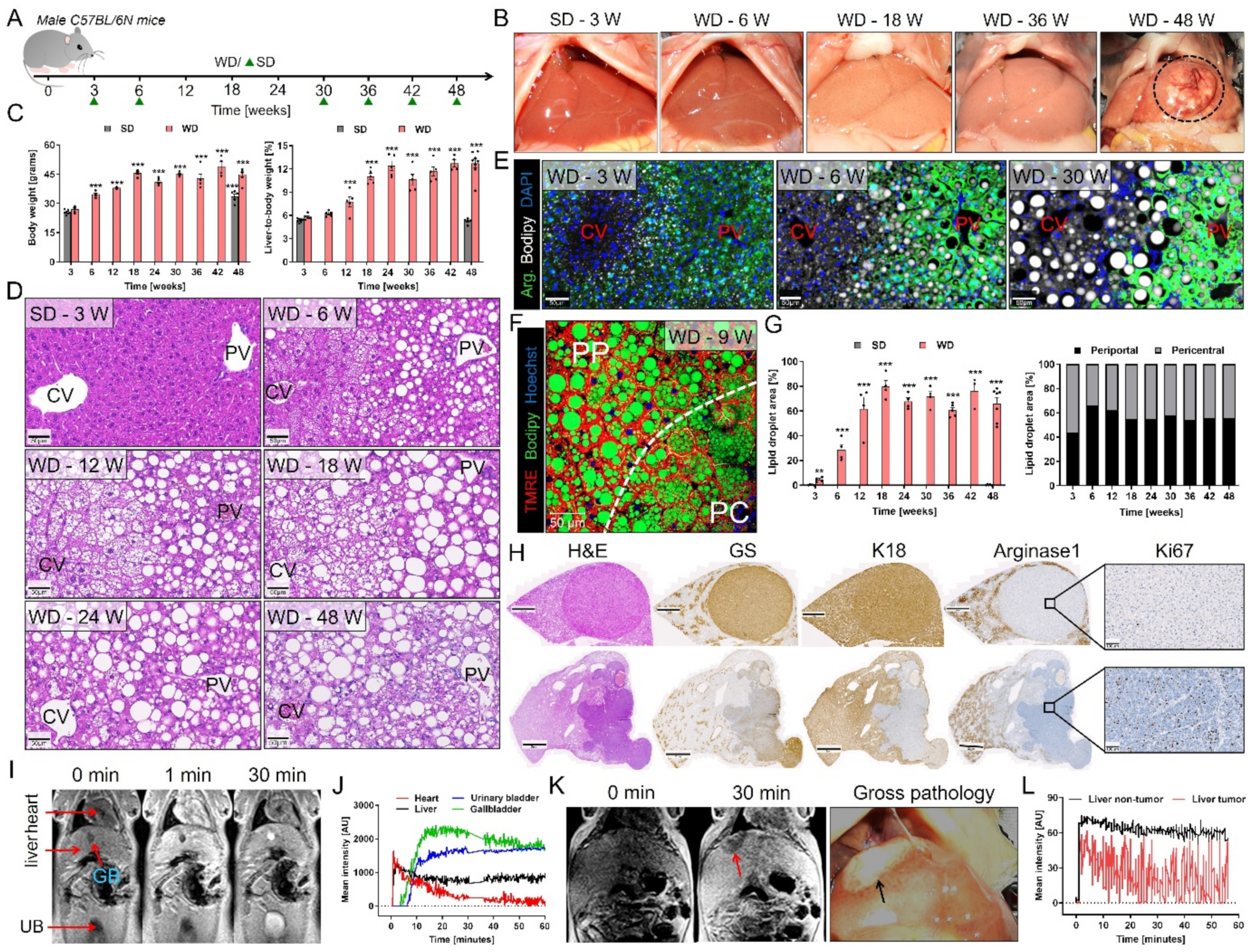
3.1. Spatio-Temporal Accumulation of Lipid Droplets and Tumor Development after Western-Style Diet Feeding
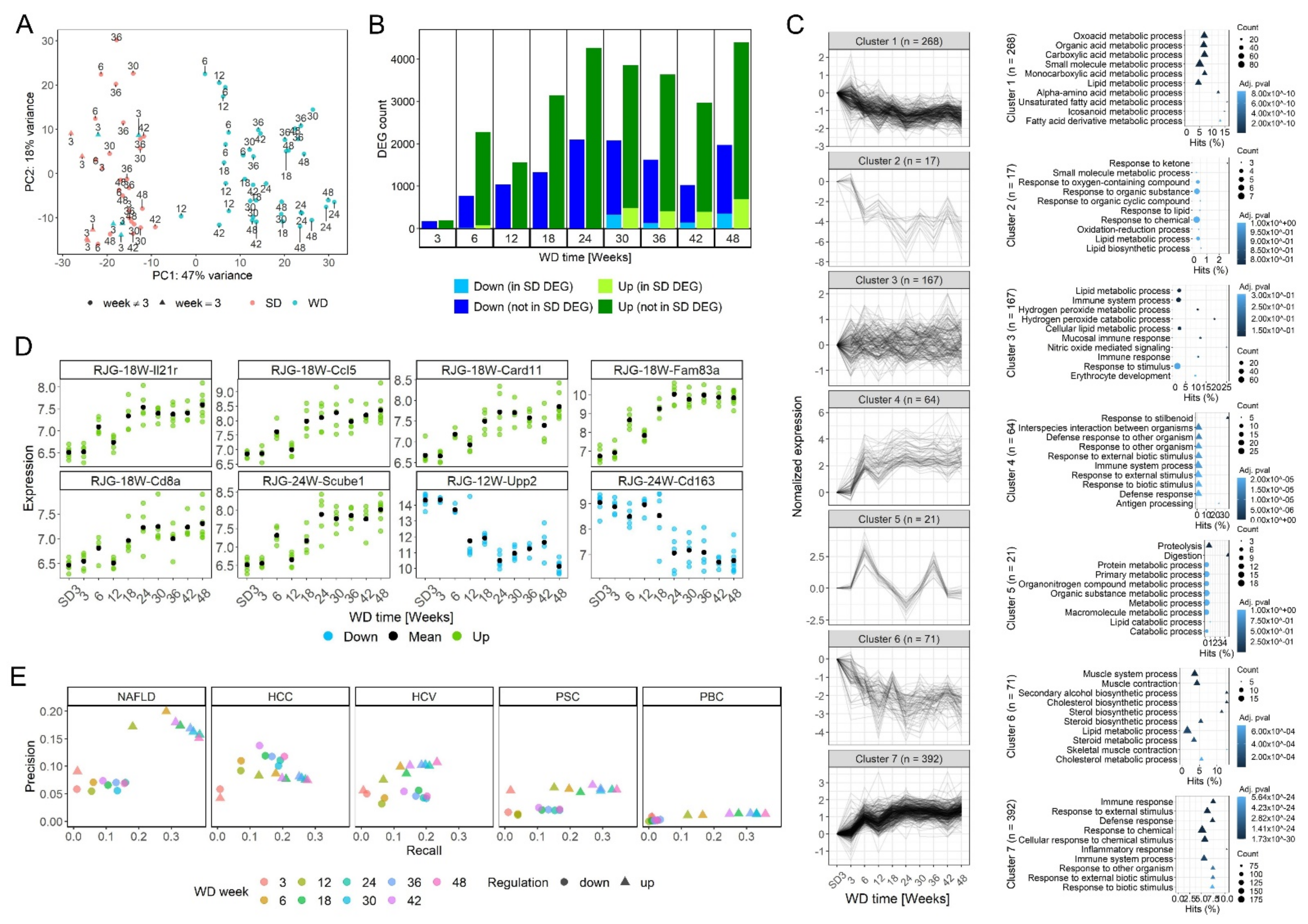
3.2. Time-Resolved Genome-Wide Expression Analysis
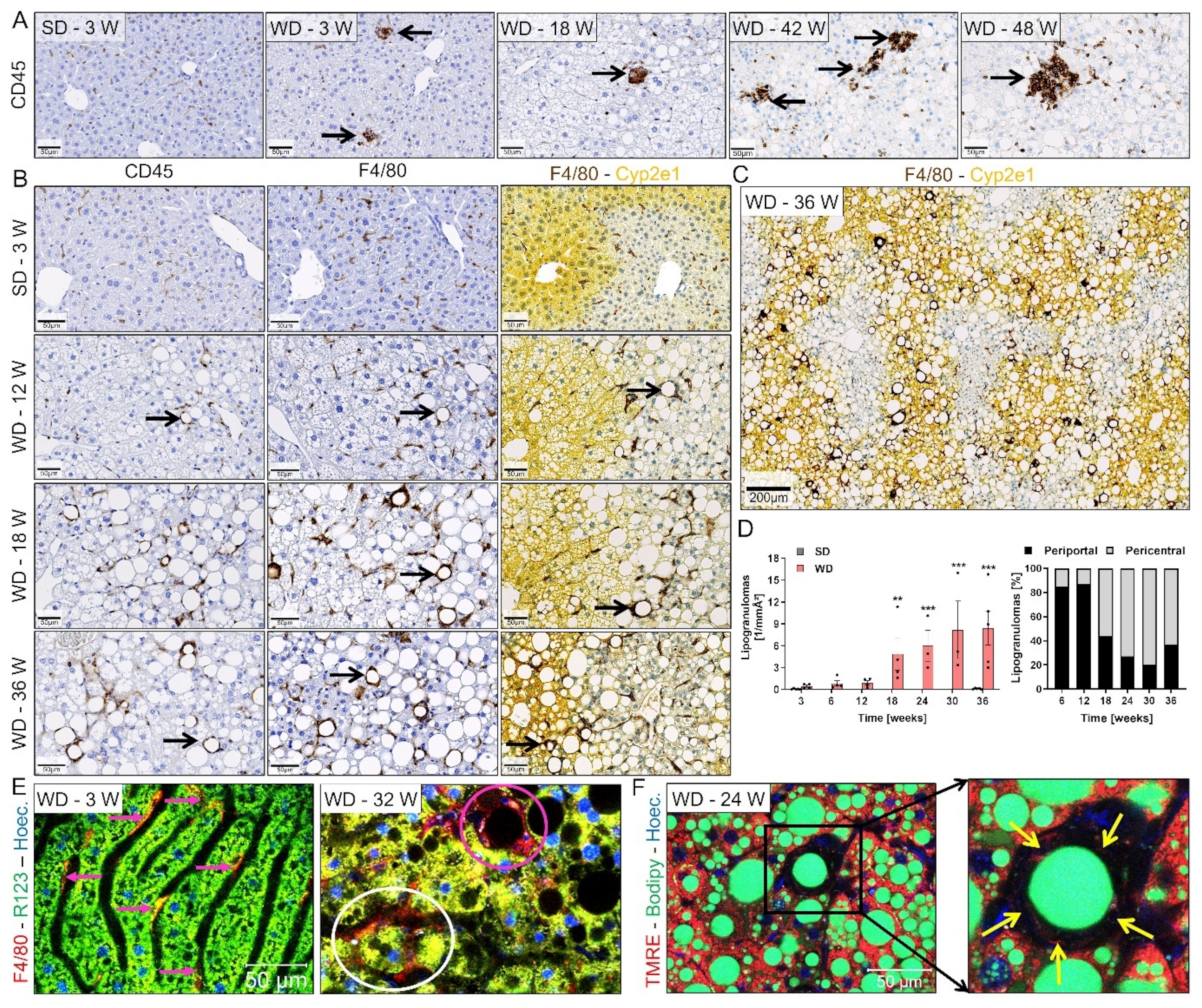
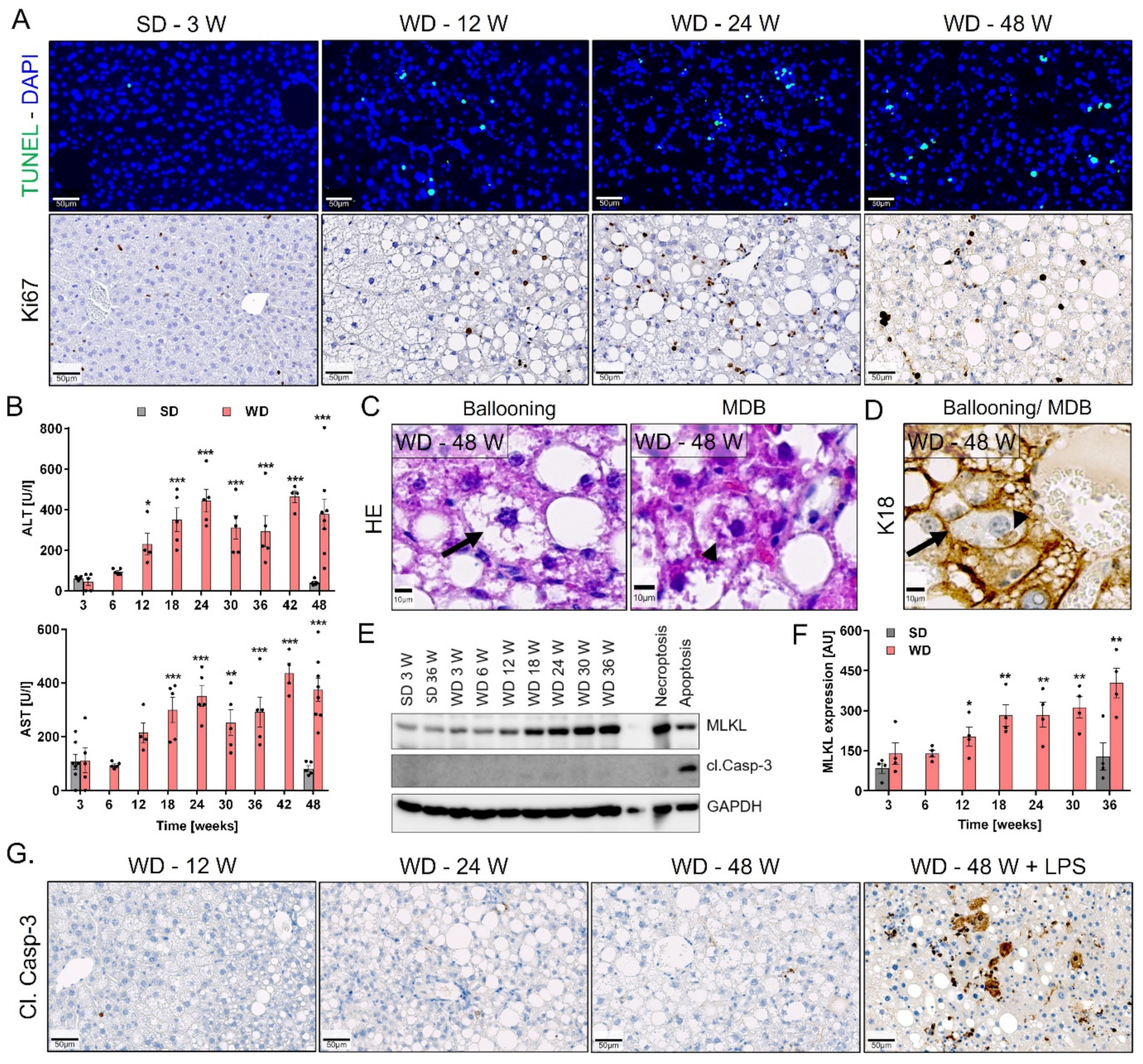
3.3. Progression from Simple Steatosis to Steatohepatitis
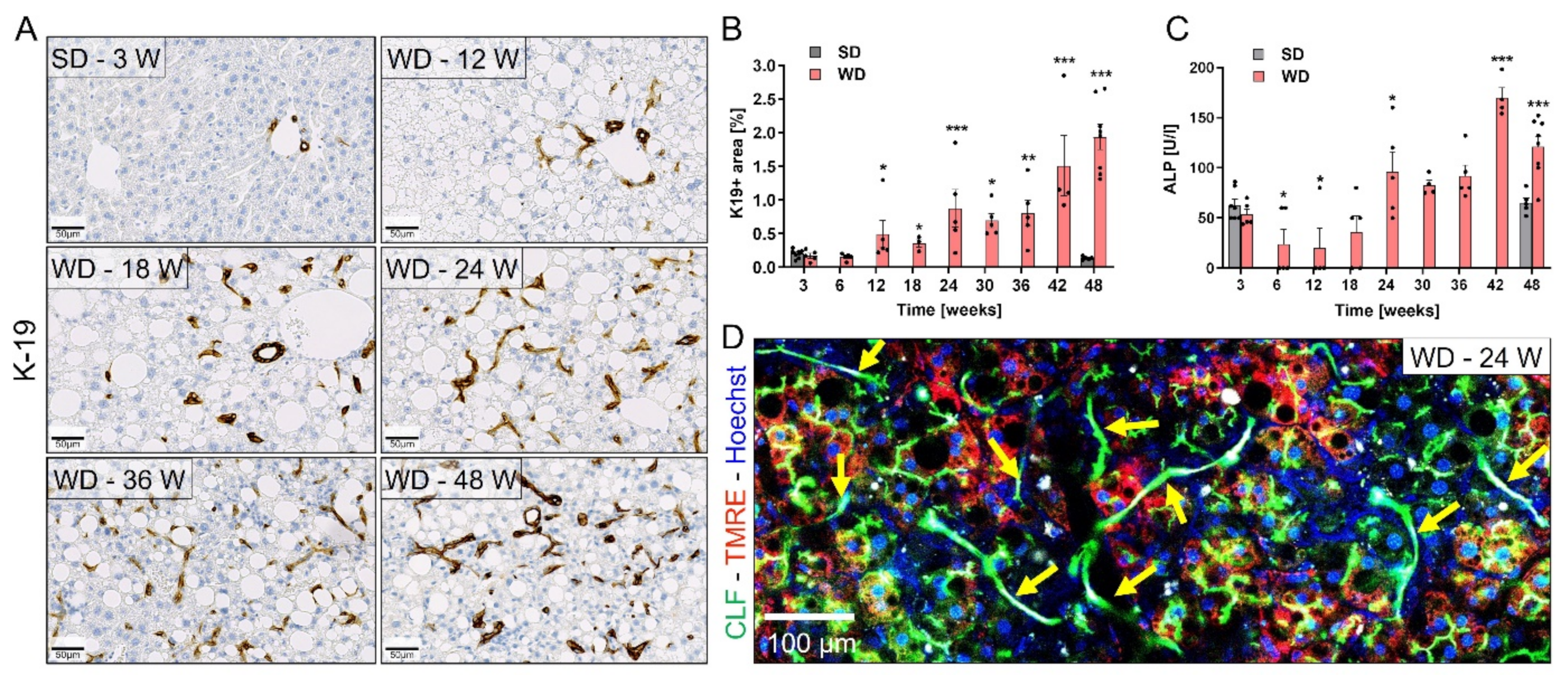
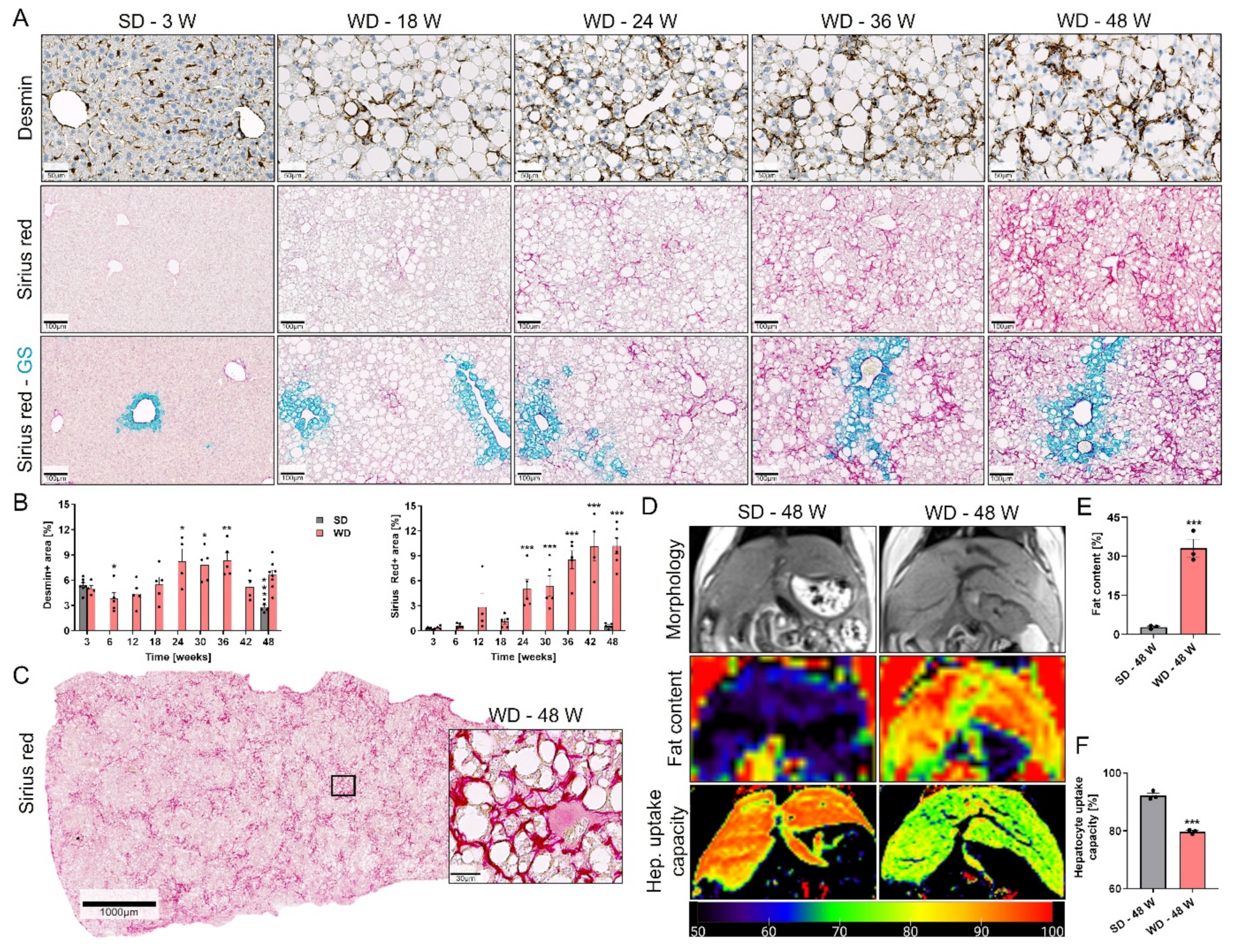
3.4. Ductular Reaction (DR) and Fibrosis Progression
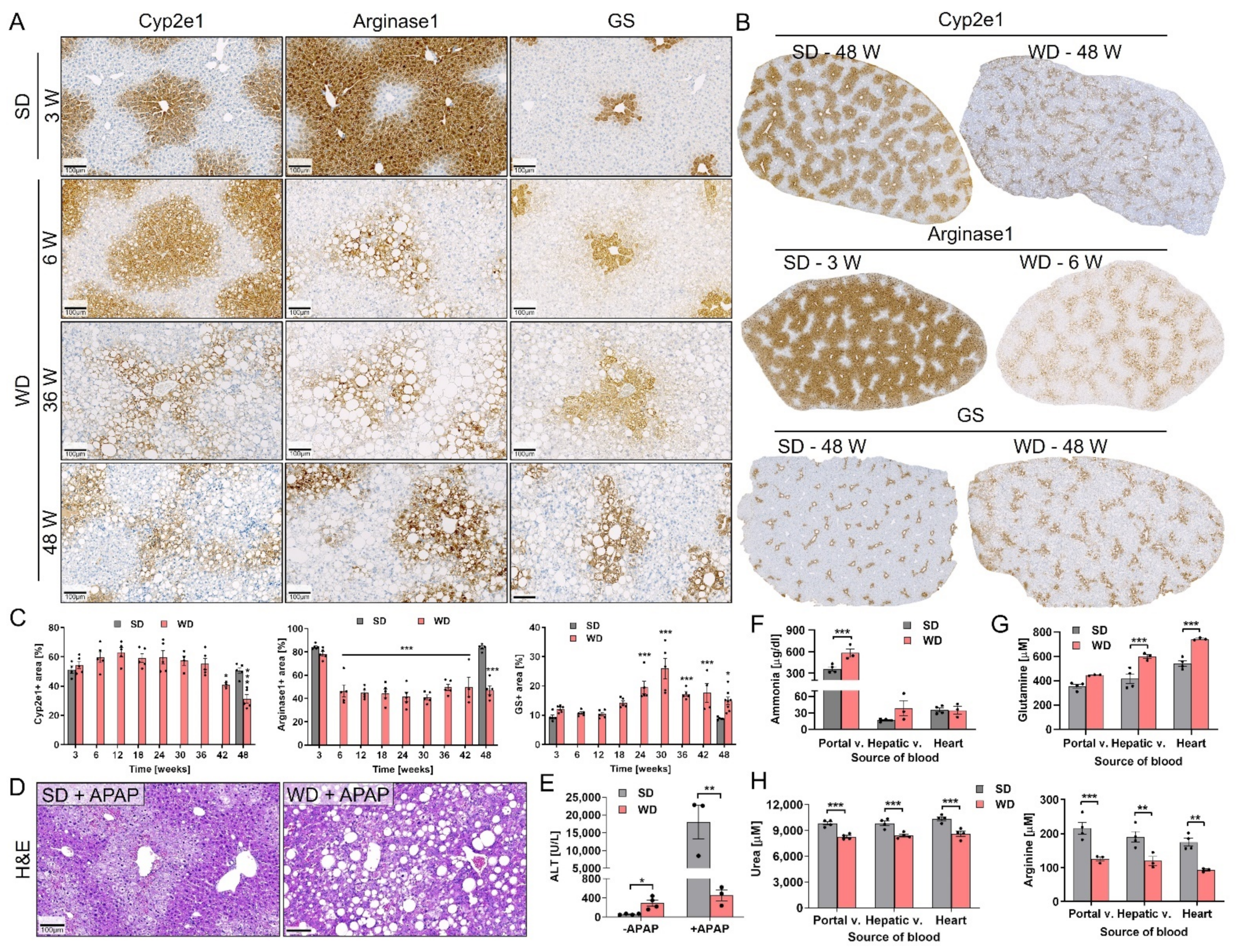
3.5. Reorganization of Zonally Expressed Enzymes and Functional Consequences
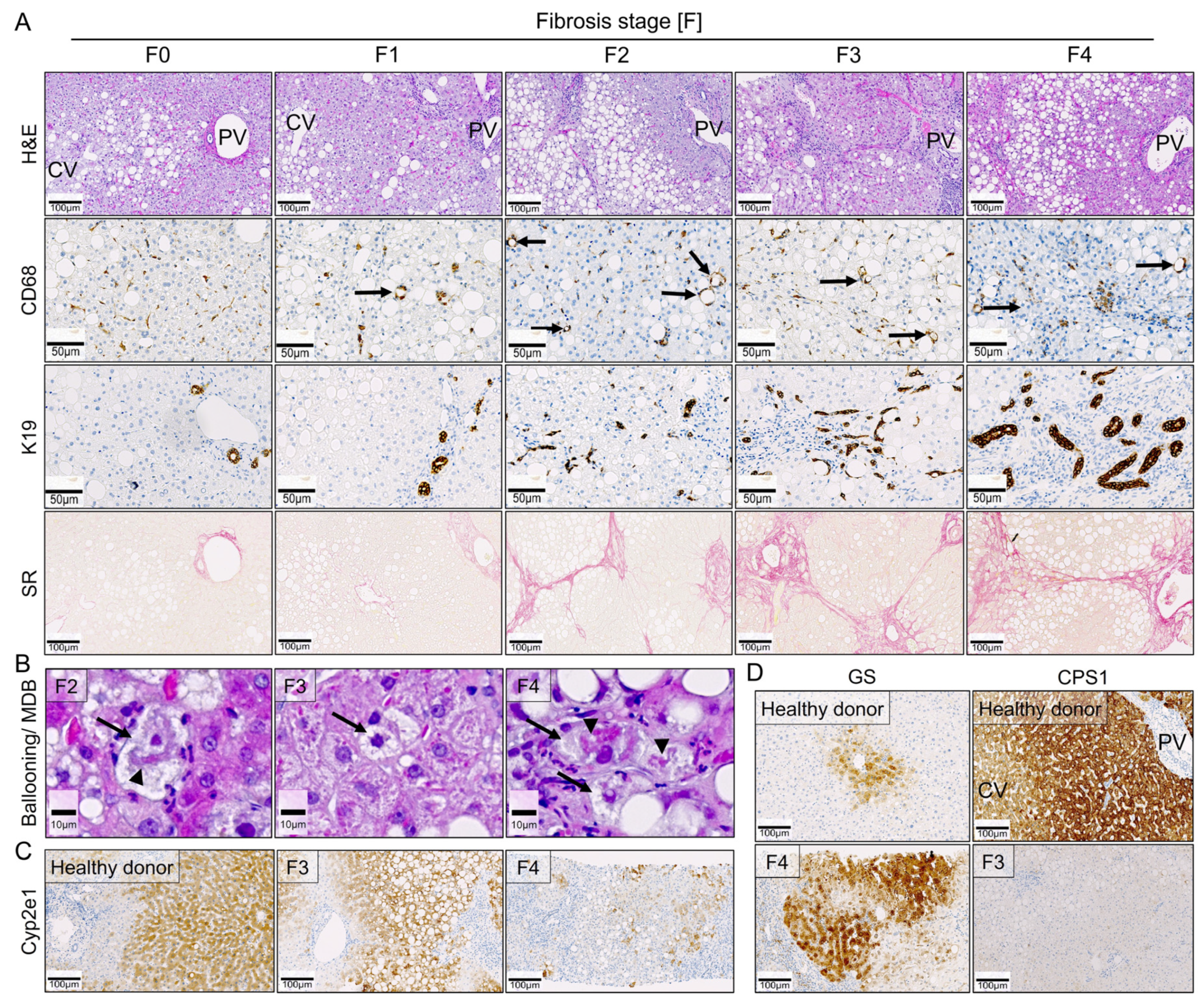
3.6. Comparison of Key Histological Features of WD-Fed Mice to NAFLD Patients
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Estes, C.; Anstee, Q.M.; Arias-Loste, M.T.; Bantel, H.; Bellentani, S.; Caballeria, J.; Colombo, M.; Craxi, A.; Crespo, J.; Day, C.P.; et al. Modeling NAFLD disease burden in China, France, Germany, Italy, Japan, Spain, United Kingdom, and United States for the period 2016–2030. J. Hepatol. 2018, 69, 896–904. [Google Scholar] [CrossRef]
- Lazarus, J.V.; Ekstedt, M.; Marchesini, G.; Mullen, J.; Novak, K.; Pericas, J.M.; Roel, E.; Romero-Gómez, M.; Ratziu, V.; Tacke, F.; et al. A cross-sectional study of the public health response to non-alcoholic fatty liver disease in Europe. J. Hepatol. 2019, 72, 14–24. [Google Scholar] [CrossRef]
- Ramadori, P.; Weiskirchen, R.; Trebicka, J.; Streetz, K. Mouse models of metabolic liver injury. Lab. Anim. 2015, 49, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Schuppan, D.; Schattenberg, J.M. Non-alcoholic steatohepatitis: Pathogenesis and novel therapeutic approaches. J. Gastroenterol. Hepatol. 2013, 28, 68–76. [Google Scholar] [CrossRef]
- Friedman, S.L.; Neuschwander-Tetri, B.A.; Rinella, M.; Sanyal, A.J. Mechanisms of NAFLD development and therapeutic strategies. Nat. Med. 2018, 24, 908–922. [Google Scholar] [CrossRef]
- Rinella, M.E.; Tacke, F.; Sanyal, A.J.; Anstee, Q.M. Report on the AASLD/EASL joint workshop on clinical trial endpoints in NAFLD. J. Hepatol. 2019, 71, 823–833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Younossi, Z.; Anstee, Q.M.; Marietti, M.; Hardy, T.; Henry, L.; Eslam, M.; George, J.; Bugianesi, E. Global burden of NAFLD and NASH: Trends, predictions, risk factors and prevention. Nat. Rev. Gastroenterol. Hepatol. 2017, 15, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Jahn, D.; Dorbath, D.; Schilling, A.-K.; Gildein, L.; Meier, C.; Vuille-Dit-Bille, R.; Schmitt, J.; Kraus, D.; Fleet, J.C.; Hermanns, H.M.; et al. Intestinal vitamin D receptor modulates lipid metabolism, adipose tissue inflammation and liver steatosis in obese mice. Biochim. Biophys. Acta (BBA)—Mol. Basis Dis. 2019, 1865, 1567–1578. [Google Scholar] [CrossRef]
- Tsuchida, T.; Lee, Y.A.; Fujiwara, N.; Ybanez, M.; Allen, B.; Martins, S.; Fiel, M.I.; Goossens, N.; Chou, H.-I.; Hoshida, Y.; et al. A simple diet- and chemical-induced murine NASH model with rapid progression of steatohepatitis, fibrosis and liver cancer. J. Hepatol. 2018, 69, 385–395. [Google Scholar] [CrossRef]
- Drescher, H.; Weiskirchen, R.; Fülöp, A.; Hopf, C.; Román, E.G.D.S.; Huesgen, P.F.; De Bruin, A.; Bongiovanni, L.; Christ, A.; Tolba, R.; et al. The Influence of Different Fat Sources on Steatohepatitis and Fibrosis Development in the Western Diet Mouse Model of Non-alcoholic Steatohepatitis (NASH). Front. Physiol. 2019, 10. [Google Scholar] [CrossRef]
- Farrell, G.; Schattenberg, J.M.; Leclercq, I.; Yeh, M.M.; Goldin, R.; Teoh, N.; Schuppan, D. Mouse Models of Nonalcoholic Steatohepatitis: Toward Optimization of Their Relevance to Human Nonalcoholic Steatohepatitis. Hepatology 2018, 69, 2241–2257. [Google Scholar] [CrossRef] [Green Version]
- Machado, M.V.; Michelotti, G.A.; Xie, G.; De Almeida, T.P.; Boursier, J.; Bohnic, B.; Guy, C.D.; Diehl, A.M. Mouse Models of Diet-Induced Nonalcoholic Steatohepatitis Reproduce the Heterogeneity of the Human Disease. PLoS ONE 2015, 10, e0127991. [Google Scholar] [CrossRef] [Green Version]
- Hansen, H.H.; Feigh, M.; Veidal, S.S.; Rigbolt, K.T.; Vrang, N.; Fosgerau, K. Mouse models of nonalcoholic steatohepatitis in preclinical drug development. Drug Discov. Today 2017, 22, 1707–1718. [Google Scholar] [CrossRef]
- Asgharpour, A.; Cazanave, S.C.; Pacana, T.; Seneshaw, M.; Vincent, R.; Banini, B.A.; Kumar, D.P.; Daita, K.; Min, H.-K.; Mirshahi, F.; et al. A diet-induced animal model of non-alcoholic fatty liver disease and hepatocellular cancer. J. Hepatol. 2016, 65, 579–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stephenson, K.; Kennedy, L.; Hargrove, L.; Demieville, J.; Thomson, J.; Alpini, G.; Francis, H. Updates on Dietary Models of Nonalcoholic Fatty Liver Disease: Current Studies and Insights. Gene. Expr. 2018, 18, 5–17. [Google Scholar] [CrossRef] [PubMed]
- Schuran, F.A.; Lommetz, C.; Steudter, A.; Ghallab, A.; Wieschendorf, B.; Schwinge, D.; Zuehlke, S.; Reinders, J.; Heeren, J.; Lohse, A.W.; et al. Aryl Hydrocarbon Receptor Activity in Hepatocytes Sensitizes to Hyperacute Acetaminophen-Induced Hepatotoxicity in Mice. Cell. Mol. Gastroenterol. Hepatol. 2021, 11, 371–388. [Google Scholar] [CrossRef]
- Schneider, K.M.; Elfers, C.; Ghallab, A.; Schneider, C.V.; Galvez, E.J.; Mohs, A.; Gui, W.; Candels, L.S.; Wirtz, T.H.; Zuehlke, S.; et al. Intestinal Dysbiosis Amplifies Acetaminophen-Induced Acute Liver Injury. Cell. Mol. Gastroenterol. Hepatol. 2021, 11, 909–933. [Google Scholar] [CrossRef]
- Campos, G.; Schmidt-Heck, W.; De Smedt, J.; Widera, A.; Ghallab, A.; Pütter, L.; González, D.; Edlund, K.; Cadenas, C.; Marchan, R.; et al. Inflammation-associated suppression of metabolic gene networks in acute and chronic liver disease. Arch. Toxicol. 2020, 94, 205–217. [Google Scholar] [CrossRef] [PubMed]
- Koeppert, S.; Ghallab, A.; Peglow, S.; Winkler, C.F.; Graeber, S.; Büscher, A.; Hengstler, J.G.; Jahnen-Dechent, W. Live Imaging of Calciprotein Particle Clearance and Receptor Mediated Uptake: Role of Calciprotein Monomers. Front. Cell Dev. Biol. 2021, 9. [Google Scholar] [CrossRef]
- Reif, R.; Ghallab, A.; Beattie, L.; Günther, G.; Kuepfer, L.; Kaye, P.M.; Hengstler, J.G. In vivo imaging of systemic transport and elimination of xenobiotics and endogenous molecules in mice. Arch. Toxicol. 2016, 91, 1335–1352. [Google Scholar] [CrossRef] [Green Version]
- Ghallab, A.; Hassan, R.; Myllys, M.; Albrecht, W.; Friebel, A.; Hoehme, S.; Hofmann, U.; Seddek, A.-L.; Braeuning, A.; Kuepfer, L.; et al. Subcellular spatio-temporal intravital kinetics of aflatoxin B1 and ochratoxin A in liver and kidney. Arch. Toxicol. 2021, 95, 2163–2177. [Google Scholar] [CrossRef]
- Zhong, X.; Nickel, M.D.; Kannengiesser, S.A.; Dale, B.M.; Kiefer, B.; Bashir, M.R. Liver fat quantification using a multi-step adaptive fitting approach with multi-echo GRE imaging. Magn. Reson. Med. 2013, 72, 1353–1365. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.H.; Lee, J.M.; Kang, H.-J.; Ahn, S.J.; Yang, H.; Kim, E.; Okuaki, T.; Han, J.K. Quantitative Assessment of Liver Function by Using Gadoxetic Acid–enhanced MRI: Hepatocyte Uptake Ratio. Radiology 2019, 290, 125–133. [Google Scholar] [CrossRef] [Green Version]
- Ghallab, A.; Cellière, G.; Henkel, S.G.; Driesch, D.; Hoehme, S.; Hofmann, U.; Zellmer, S.; Godoy, P.; Sachinidis, A.; Blaszkewicz, M.; et al. Model-guided identification of a therapeutic strategy to reduce hyperammonemia in liver diseases. J. Hepatol. 2015, 64, 860–871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schenk, A.; Ghallab, A.; Hofmann, U.; Hassan, R.; Schwarz, M.; Schuppert, A.; Schwen, L.O.; Braeuning, A.; Teutonico, D.; Hengstler, J.; et al. Physiologically-based modelling in mice suggests an aggravated loss of clearance capacity after toxic liver damage. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Holland, C.H.; Ramirez Flores, R.O.; Myllys, M.; Hassan, R.; Edlund, K.; Hofmann, U.; Marchan, R.; Cadenas, C.; Reinders, J.; Hoehme, S.; et al. Transcriptomic cross-species analysis of chronic liver disease reveals consistent regulation between humans and mice. Hepatol. Commun. 2021. Accepted. [Google Scholar] [CrossRef]
- Ghallab, A.; Myllys, M.; Holland, C.H.; Zaza, A.; Murad, W.; Hassan, R.; Ahmed, Y.A.; Abbas, T.; Abdelrahim, E.A.; Schneider, K.M.; et al. Influence of Liver Fibrosis on Lobular Zonation. Cells 2019, 8, 1556. [Google Scholar] [CrossRef] [Green Version]
- Patro, R.; Duggal, G.; Love, M.I.; Irizarry, M.I.L.R.A.; Kingsford, C. Salmon provides fast and bias-aware quantification of transcript expression. Nat. Methods 2017, 14, 417–419. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Soneson, C.; Hickey, P.F.; Johnson, L.K.; Pierce, N.T.; Shepherd, L.; Morgan, M.; Patro, R. Tximeta: Reference sequence checksums for provenance identification in RNA-seq. PLoS Comput. Biol. 2020, 16, e1007664. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome. Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome. Biol. 2010, 11, R12–R14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Itzel, T.; Neubauer, M.; Ebert, M.; Evert, M.; Teufel, A. Hepamine—A Liver Disease Microarray Database, Visualization Platform and Data-Mining Resource. Sci. Rep. 2020, 10, 4760–4767. [Google Scholar] [CrossRef] [PubMed]
- Lian, Q.; Wang, S.; Zhang, G.; Wang, D.; Luo, G.; Tang, J.; Chen, L.; Gu, J. HCCDB: A Database of Hepatocellular Carcinoma Expression Atlas. Genom. Proteom. Bioinform. 2018, 16, 269–275. [Google Scholar] [CrossRef]
- Schneider, A.T.; Gautheron, J.; Feoktistova, M.; Roderburg, C.; Loosen, S.H.; Roy, S.; Benz, F.; Schemmer, P.; Büchler, M.W.; Nachbur, U.; et al. RIPK1 Suppresses a TRAF2-Dependent Pathway to Liver Cancer. Cancer Cell 2016, 31, 94–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hofmann, U.; Maier, K.; Niebel, A.; Vacun, G.; Reuss, M.; Mauch, K. Identification of metabolic fluxes in hepatic cells from transient13C-labeling experiments: Part I. Experimental observations. Biotechnol. Bioeng. 2007, 100, 344–354. [Google Scholar] [CrossRef]
- Maier, K.; Hofmann, U.; Reuss, M.; Mauch, K. Dynamics and Control of the Central Carbon Metabolism in Hepatoma Cells. BMC Syst. Biol. 2010, 4, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bankhead, P.; Loughrey, M.B.; Fernández, J.A.; Dombrowski, Y.; McArt, D.; Dunne, P.D.; McQuaid, S.; Gray, R.T.; Murray, L.J.; Coleman, H.G.; et al. QuPath: Open source software for digital pathology image analysis. Sci. Rep. 2017, 7, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Berg, S.; Kutra, D.; Kroeger, T.; Straehle, C.N.; Kausler, B.X.; Haubold, C.; Schiegg, M.; Ales, J.; Beier, T.; Rudy, M.; et al. ilastik: Interactive machine learning for (bio)image analysis. Nat. Methods. 2019, 16, 1226–1232. [Google Scholar] [CrossRef] [PubMed]
- Rimola, J.; Forner, A.; Sapena, V.; Llarch, N.; Darnell, A.; Díaz, A.; García-Criado, A.; Bianchi, L.; Vilana, R.; Díaz-González, A.; et al. Performance of gadoxetic acid MRI and diffusion-weighted imaging for the diagnosis of early recurrence of hepatocellular carcinoma. Eur. Radiol. 2019, 30, 186–194. [Google Scholar] [CrossRef]
- Sutti, S.; Albano, E. Adaptive immunity: An emerging player in the progression of NAFLD. Nat. Rev. Gastroenterol. Hepatol. 2019, 17, 81–92. [Google Scholar] [CrossRef]
- Kleiner, D.E.; Brunt, E.M.; Van Natta, M.; Behling, C.; Contos, M.J.; Cummings, O.W.; Ferrell, L.D.; Liu, Y.-C.; Torbenson, M.S.; Unalp-Arida, A.; et al. Design and validation of a histological scoring system for nonalcoholic fatty liver disease. Hepatology 2005, 41, 1313–1321. [Google Scholar] [CrossRef] [PubMed]
- Gadd, V.L.; Skoien, R.; Powell, E.; Fagan, K.J.; Winterford, C.; Horsfall, L.; Irvine, K.; Clouston, A.D. The portal inflammatory infiltrate and ductular reaction in human nonalcoholic fatty liver disease. Hepatology 2014, 59, 1393–1405. [Google Scholar] [CrossRef] [PubMed]
- Schuppan, D.; Surabattula, R.; Wang, X. Determinants of fibrosis progression and regression in NASH. J. Hepatol. 2018, 68, 238–250. [Google Scholar] [CrossRef]
- Sezgin, S.; Hassan, R.; Zühlke, S.; Kuepfer, L.; Hengstler, J.G.; Spiteller, M.; Ghallab, A. Spatio-temporal visualization of the distribution of acetaminophen as well as its metabolites and adducts in mouse livers by MALDI MSI. Arch. Toxicol. 2018, 92, 2963–2977. [Google Scholar] [CrossRef]
- De Chiara, F.; Heebøll, S.; Marrone, G.; Montoliu, C.; Hamilton-Dutoit, S.; Ferrandez, A.; Andreola, F.; Rombouts, K.; Grønbæk, H.; Felipo, V.; et al. Urea cycle dysregulation in non-alcoholic fatty liver disease. J. Hepatol. 2018, 69, 905–915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartl, M.; Pfaff, M.; Ghallab, A.; Driesch, D.; Henkel, S.G.; Hengstler, J.G.; Schuster, S.; Kaleta, C.; Gebhardt, R.; Zellmer, S.; et al. Optimality in the zonation of ammonia detoxification in rodent liver. Arch. Toxicol. 2015, 89, 2069–2078. [Google Scholar] [CrossRef]
- Schliess, F.; Hoehme, S.; Henkel, S.G.; Ghallab, A.; Driesch, D.; Böttger, J.; Guthke, R.; Pfaff, M.; Hengstler, J.G.; Gebhardt, R.; et al. Integrated metabolic spatial-temporal model for the prediction of ammonia detoxification during liver damage and regeneration. Hepatology 2014, 60, 2040–2051. [Google Scholar] [CrossRef]
- Häussinger, D. Ammonia, urea production and pH regulation. In Hepatology: From Basic Science to Clinical Practice; Rodes, J.B.J., Blei, A., Reichen, J., Rizzetto, M., Eds.; Blackwell Publishing: Hoboken, NJ, USA, 2007; pp. 181–192. [Google Scholar]
- Hawkins, R.A.; Jessy, J.; Mans, A.M.; Chedid, A.; DeJoseph, M.R. Neomycin Reduces the Intestinal Production of Ammonia from Glutamine. Adv. Exp. Med. Biol. 1994, 368, 125–134. [Google Scholar] [CrossRef]
- Lefere, S.; Tacke, F. Macrophages in obesity and non-alcoholic fatty liver disease: Crosstalk with metabolism. JHEP Rep. 2019, 1, 30–43. [Google Scholar] [CrossRef] [Green Version]
- Schwabe, R.F.; Luedde, T. Apoptosis and necroptosis in the liver: A matter of life and death. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 738–752. [Google Scholar] [CrossRef] [PubMed]
- Manco, R.; Clerbaux, L.-A.; Verhulst, S.; Nader, M.B.; Sempoux, C.; Ambroise, J.; Bearzatto, B.; Gala, J.L.; Horsmans, Y.; van Grunsven, L.; et al. Reactive cholangiocytes differentiate into proliferative hepatocytes with efficient DNA repair in mice with chronic liver injury. J. Hepatol. 2019, 70, 1180–1191. [Google Scholar] [CrossRef]
- Raven, A.; Lu, W.-Y.; Man, T.Y.; Ferreira-Gonzalez, S.; O’Duibhir, E.; Dwyer, B.; Thomson, J.P.; Meehan, R.; Bogorad, R.; Koteliansky, V.; et al. Cholangiocytes act as facultative liver stem cells during impaired hepatocyte regeneration. Nature 2017, 547, 350–354. [Google Scholar] [CrossRef]
- Jörs, S.; Jeliazkova, P.; Ringelhan, M.; Thalhammer, J.; Dürl, S.; Ferrer, J.; Sander, M.; Heikenwalder, M.; Schmid, R.M.; Siveke, J.T.; et al. Lineage fate of ductular reactions in liver injury and carcinogenesis. J. Clin. Investig. 2015, 125, 2445–2457. [Google Scholar] [CrossRef] [Green Version]
- Kamimoto, K.; Nakano, Y.; Kaneko, K.; Miyajima, A.; Itoh, T. Multidimensional imaging of liver injury repair in mice reveals fundamental role of the ductular reaction. Commun. Biol. 2020, 3, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ghallab, A.; Hofmann, U.; Sezgin, S.; Vartak, N.; Hassan, R.; Zaza, A.; Godoy, P.; Schneider, K.M.; Guenther, G.; Ahmed, Y.; et al. Bile Microinfarcts in Cholestasis Are Initiated by Rupture of the Apical Hepatocyte Membrane and Cause Shunting of Bile to Sinusoidal Blood. Hepatology 2018, 69, 666–683. [Google Scholar] [CrossRef]
- Schneider, K.M.; Candels, L.S.; Hov, J.R.; Myllys, M.; Hassan, R.; Schneider, C.V.; Wahlström, A.; Mohs, A.; Zühlke, S.; Liao, L.; et al. Gut microbiota depletion exacerbates cholestatic liver injury via loss of FXR signaling. Nat. Metab. 2021, 3, 1228–1241. [Google Scholar]
- Bastati, N.; Feier, D.; Wibmer, A.; Traussnigg, S.; Balassy, C.; Tamandl, D.; Einspieler, H.; Wrba, F.; Trauner, M.; Herold, C.; et al. Noninvasive Differentiation of Simple Steatosis and Steatohepatitis by Using Gadoxetic Acid–enhanced MR Imaging in Patients with Nonalcoholic Fatty Liver Disease: A Proof-of-Concept Study. Radiology 2014, 271, 739–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hailfinger, S.; Jaworski, M.; Braeuning, A.; Buchmann, A.; Schwarz, M. Zonal gene expression in murine liver: Lessons from tumors. Hepatology 2006, 43, 407–414. [Google Scholar] [CrossRef]
- Pesce, J.; Kaviratne, M.; Ramalingam, T.R.; Thompson, R.W.; Urban, J.; Cheever, A.W.; Young, D.A.; Collins, M.; Grusby, M.J.; Wynn, T.A. The IL-21 receptor augments Th2 effector function and alternative macrophage activation. J. Clin. Investig. 2006, 116, 2044–2055. [Google Scholar] [CrossRef]
- Seo, W.; Shimizu, K.; Kojo, S.; Okeke, A.; Kohwi-Shigematsu, T.; Fujii, S.-I.; Taniuchi, I. Runx-mediated regulation of CCL5 via antagonizing two enhancers influences immune cell function and anti-tumor immunity. Nat. Commun. 2020, 11, 1–16. [Google Scholar] [CrossRef]
- Bedsaul, J.R.; Carter, N.; Deibel, K.E.; Hutcherson, S.M.; Jones, T.A.; Wang, Z.; Yang, C.; Yang, Y.-K.; Pomerantz, J.L. Mechanisms of Regulated and Dysregulated CARD11 Signaling in Adaptive Immunity and Disease. Front. Immunol. 2018, 9, 2105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, F.; Wang, X.; Liu, F.; Meng, Q.; Yu, Y. FAM83A drives PD-L1 expression via ERK signaling and FAM83A/PD-L1 co-expression correlates with poor prognosis in lung adenocarcinoma. Int. J. Clin. Oncol. 2020, 25, 1612–1623. [Google Scholar] [CrossRef]
- Breuer, D.A.; Pacheco, M.C.; Washington, M.K.; Montgomery, S.A.; Hasty, A.H.; Kennedy, A. CD8+ T cells regulate liver injury in obesity-related nonalcoholic fatty liver disease. Am. J. Physiol. Liver Physiol. 2020, 318, G211–G224. [Google Scholar] [CrossRef] [PubMed]
- Ostroumov, D.; Duong, S.; Wingerath, J.; Woller, N.; Manns, M.P.; Timrott, K.; Kleine, M.; Ramackers, W.; Roessler, S.; Nahnsen, S.; et al. Transcriptome Profiling Identifies TIGIT as a Marker of T-Cell Exhaustion in Liver Cancer. Hepatology 2020, 73, 1399–1418. [Google Scholar] [CrossRef] [PubMed]
- Liao, W.-J.; Wu, M.-Y.; Peng, C.-C.; Tung, Y.-C.; Yang, R.-B. Epidermal growth factor-like repeats of SCUBE1 derived from platelets are critical for thrombus formation. Cardiovasc. Res. 2019, 116, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Van Herck, M.A.; Weyler, J.; Kwanten, W.J.; Dirinck, E.L.; De Winter, B.Y.; Francque, S.M.; Vonghia, L. The Differential Roles of T Cells in Non-alcoholic Fatty Liver Disease and Obesity. Front. Immunol. 2019, 10, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hudert, C.A.; Selinski, S.; Rudolph, B.; Bläker, H.; Loddenkemper, C.; Thielhorn, R.; Berndt, N.; Golka, K.; Cadenas, C.; Reinders, J.; et al. Genetic determinants of steatosis and fibrosis progression in paediatric non-alcoholic fatty liver disease. Liver Int. 2018, 39, 540–556. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reagent or Resource | Source | Identifier |
|---|---|---|
| Antibodies, Reagents, and Dyes Used for Immunohistochemistry | ||
| Anti-liver arginase1 antibody, rabbit | Abcam, Cambridge, UK | ab203490 |
| Anti-arginase1 antibody, goat | Novus Biologicals, Littleton, USA | NB100-59740 |
| Bodipy 495/503 | Thermo Fisher Scientific, Waltham, USA | D3922 |
| Anti-mouse CD45 antibody, rat | BD Bioscience, Heidelberg, Germany | 550539 |
| Anti-human CD68 monoclonal antibody, mouse | DakoCytomation A/S, Glostrup, Denmark | M0876 |
| Anti-K18 polyclonal antibody, rabbit | Proteintech, Manchester, UK | 10830-1-AP |
| Recombinant anti-K19 antibody, rabbit | Abcam, Cambridge, UK | ab52625 |
| Recombinant anti-CPS1 monoclonal antibody, rabbit | Abcam, Cambridge, UK | ab129076 |
| Anti-Cyp2e1 antibody, rabbit | Sigma-Aldrich, St. Louis, USA | HPA009128 |
| Anti-mouse desmin antibody, rabbit | Thermo Fisher Scientific, Waltham, USA | RB -9014-P0 |
| Anti-mouse F4/80 monoclonal antibody, rat | Bio-Rad, Hercules, USA | MCA497 |
| Anti-GS polyclonal antibody, rabbit | Sigma, St. Louis, USA | G2781 |
| Anti-GS polyclonal antibody, rabbit | Sigma, St. Louis, USA | G2781 |
| Anti-Ki67 antibody, rabbit | Cell Signaling Technology, Danvers, USA | D3B5 |
| Anti- cl. Caspase 3 (rabbit) monoclonal | Cell Signaling Technology, Danvers, USA | 9661S |
| Fluorescent Markers/Dyes Used for Intravital Imaging | ||
| Hoechst 33258 | Thermo Fisher Scientific, Waltham, USA | H21491 |
| Tetramethylrhodamine ethyl ester (TMRE) | Thermo Fisher Scientific, Waltham, USA | T669 |
| Cholyl-lysyl-fluorescein (CLF) | Corning | 451041 |
| PE-F4/80 antibody | Thermo Scientific (eBioscience) , Waltham, USA | 12-4801-82 |
| Rhodamine 123 | Thermo Fisher Scientific, Waltham, USA | R302 |
| Bodipy 493/503 | Thermo Fisher Scientific, Waltham, USA | D3922 |
| Mouse Diets | ||
| ssniff R/M-H, 10 mm standard diet | Ssniff, Soest, Germany | V1534-000 |
| Western-style diet | Research Diets, New Brunswick, USA | D09100301 |
| Drugs/Contrast Agents/Toxins | ||
| Acetaminophen | Sigma-Aldrich, St. Louis, USA | A7085-500G |
| LPS | Sigma-Aldrich, St. Louis, USA | 297-473-0 |
| Gadoxetic acid (Primovist) 0.25 mmol/mL | Bayer, Wuppertal, Germany | KT07561 |
| Commercial Kits/Reagents | ||
| DeadEnd™ Fluorometric TUNEL System | Promega, Walldorf, Germany | G3250 |
| Bluing Reagent | Roche, Mannheim, Germany | 05 266 769 001 |
| Discovery yellow Kit (RUO) | Roche, Mannheim, Germany | 07 698 445 001 |
| Discovery Teal HRP Kit (RUO) | Roche, Mannheim, Germany | 8254338001 |
| Chromo Map DAB | Roche, Mannheim, Germany | 05 266 645 001 |
| Piccolo general chemistry 13 | Hitado, Möhnesee, Germany | AB-114-400-0029 |
| Picrosirius Red Stain Kit | Polysciences Polysciences Inc., Warrington, USA | 24901 |
| RNeasy Mini Kit | Qiagen, Hilden, Germany | 74116 |
| RNase-Free DNase Set | Qiagen, Hilden, Germany | 79254 |
| RNA BR Assay Kit | Thermo Fisher Scientific, Waltham, USA | Q10210 |
| RNA 6000 Nano Kit | Agilent Technologies, CA, USA | 5067-1511 |
| Qubit 1X dsDNA HS Assay Kit | Thermo Fisher Scientific, Waltham, USA | Q33230 |
| DNA 1000 Kit | Agilent Technologies, CA, USA | 5067-1504 |
| Antibodies Used for Western Blotting | ||
| Anti- MLKL | Biorbyt LLC, Cambridge, UK | orb32399 |
| Anti- cleaved-Caspase-3 | Cell Signaling Technology, Danvers, USA | 9661S |
| Anti- GAPDH | AbD Serotec, Hercules, USA | MCA 4739 |
| Software and Algorithms | ||
| GraphPad Prism 9.1 Software | GraphPad, San Diego, USA | NA |
| Zen | Carl-Zeiss, Jena, Germany | NA |
| ImageJ 1.8.0_172 | https://imagej.nih.gov; 22 April 2021 | NA |
| Instruments | ||
| LSM MP7 two-photon microscope | Zeiss, Jena, Germany | NA |
| Axio Scan.Z1 | Zeiss, Jena, Germany | N/A |
| Confocal Laser Scanning Microscope FLUOVIEW FV1000 | Olympus, Hamburg, Germany | N/A |
| DISCOVERY ULTRA Automated Slide Preparation System | Roche, Mannheim, Germany | N/A |
| Piccolo Xpress® chemistry analyzer | Abaxis, Union City, USA | N/A |
| PocketChem BA PA-4140 ammonia meter | Arkray.inc, Amstelveen, The Netherlands | N/A |
| 3Tesla MRI scanner | Prisma, Siemens Healthineers, Erlangen, Germany | |
| Ingredient | Grams | Kcal | % |
|---|---|---|---|
| Casein, 80 mesh | 200 | 800 | 22.12 |
| l-cystine | 3 | 12 | 0.33 |
| Maltodextrin 10 | 100 | 400 | 11.06 |
| Fructose | 200 | 800 | 22.12 |
| Sucrose | 96 | 384 | 10.62 |
| Cellulose, BW200 | 50 | 0 | 5.53 |
| Soybean oil | 25 | 225 | 2.77 |
| Primex shortening | 135 | 1215 | 14.93 |
| Lard | 20 | 180 | 2.21 |
| Mineral Mix S10026 | 10 | 0 | 1.11 |
| Dicalcium phosphate | 13 | 0 | 1.44 |
| Calcium carbonate | 5.5 | 0 | 0.61 |
| Potassium citrate, 1 H2O | 16.5 | 0 | 1.83 |
| Vitamin Mix V10001 | 10 | 40 | 1.11 |
| Choline bitartrate | 2 | 0 | 0.22 |
| Cholesterol | 18 | 0 | 1.99 |
| FD&C Yellow dye | 0.05 | 0 | 0.006 |
| Total | 904.05 | 4056 | 100 |
| Total protein | 20 Kcal % | 22 | |
| Total carbohydrate | 40 Kcal % | 45 | |
| Total fat | 40 Kcal % | 20 |
| Diet | Feeding Time (Weeks) | Number of Mice Analyzed | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Liver/Body Weight, RNA-seq, Histology, IHC | HCC | MRI | Intravital Imaging | Nitrogen Metabolism | APAP Experiment | Necroptosis Analysis | LPS | |||
| + | − | |||||||||
| Western Diet | 3 | 5 | 0 | 5 | 3 | 4 | ||||
| 6 | 5 | 0 | 5 | 4 | ||||||
| 9 | 3 | - | ||||||||
| 12 | 5 | 0 | 5 | 4 | ||||||
| 18 | 5 | 0 | 5 | 4 | ||||||
| 24 | 5 | 0 | 5 | 6 | 4 | |||||
| 30 | 5 | 4 | 1 | 4 | ||||||
| 32 | 3 | |||||||||
| 36 | 5 | 1 | 4 | 4 | ||||||
| 42 | 4 | 5 | 4 | 3 | ||||||
| 48 | 8 | 6 | 2 | 4 | 3 | |||||
| ~50 | 7 | |||||||||
| Standard Diet | 3 | 7 | 0 | 7 | 3 | 4 | ||||
| 6 | 5 | 0 | 5 | |||||||
| 30 | 5 | 0 | 5 | |||||||
| 36 | 7 | 0 | 7 | 4 | ||||||
| 42 | 3 | 0 | 3 | 4 | ||||||
| 48 | 5 | 0 | 5 | 4 | ||||||
| ~50 | 7 | |||||||||
| Dye/Marker | Marker for | Dose (mg/kg) | Vehicle | Two-Photon Excitation Range (nm) |
|---|---|---|---|---|
| Hoechst 33258 | Nuclei | 5 | PBS | 700–800 |
| TMRE | Lobular zonation; mitochondrial membrane potential | 0.96 | Methanol/PBS (1:1) | 780–820 |
| Rhodamine123 | 0.8 | Methanol/PBS (1:1) | 720–820 | |
| Cholyl-lysyl-fluorescein | Bile acid analogue | 1 | PBS | 740–820 |
| Bodipy 493/503 | Lipids | 0.004 | DMSO | 900–940 |
| PE-F4/80 antibody | Macrophages | 0.06 | PBS | 720–760 |
| Target | Primary Antibodies | Secondary Antibodies | ||
|---|---|---|---|---|
| Antibody | Dilution | Antibody | Dilution | |
| Lipids | Bodipy 495/503 | 2 µg/mL | - | - |
| Arginase1 | Anti-arginase1 antibody, goat | 1:100 | Cy™5-conjugated AffiniPure donkey anti-goat IgG (H + L) | 1:200 |
| Anti-liver arginase1 antibody, rabbit | 1:2000 | Ultra-Map anti-rabbit HRP | Automatic Discovery Ready to use | |
| Ultra-Map anti-rabbit alkaline phosphatase | ||||
| Leukocyte common antigen | Anti-mouse CD45 antibody, rat | 1:400 | Ultra-Map anti-rat HRP | |
| Macrophages, human | Anti-human CD68 monoclonal antibody, mouse | 1:500 | Ultra-Map anti-mouse HRP | |
| Cytoskeleton | Anti-K18 polyclonal antibody, rabbit | 1:400 | Ultra-Map anti-rabbit HRP | |
| Cholangiocyte, mouse | Recombinant anti-K19 antibody, rabbit | 1:500 | Ultra-Map anti-rabbit HRP | |
| Cholangiocyte, human | Recombinant anti-K19 antibody, rabbit | 1:2000 | Ultra-Map anti-rabbit HRP | |
| Carbamoyl-Phosphate Synthase1 | Recombinant anti-CPS1 monoclonal antibody, rabbit | 1:500 | Ultra-Map anti-rabbit HRP | |
| Cyp2e1 | Anti-Cyp2e1 antibody, rabbit | 1:100 | Ultra-Map anti-rabbit HRP | |
| Ultra-Map anti-rabbit alkaline phosphatase | ||||
| Hepatic stellate cells | Anti-mouse desmin antibody, rabbit | 1:400 | Ultra-Map anti-rabbit HRP | |
| Macrophages, mouse | Anti-mouse F4/80 monoclonal antibody, rat | 1:50 | Ultra-Map anti-rat HRP | |
| Glutamine synthetase, mouse | Anti-GS polyclonal antibody, rabbit | 1:15,000 | Ultra-Map anti-rabbit HRP | |
| Apoptosis | Anti- cl. Caspase 3 monoclonal antibody, rabbit | 1:500 | Ultra-Map anti-rabbit HRP | |
| Glutamine synthetase, human | Anti-GS polyclonal antibody, rabbit | 1:5000 | Ultra-Map anti-rabbit HRP | |
| Cell proliferation antigen | Anti-Ki-67 antibody, rabbit | 1:100 | Ultra-Map anti-rabbit HRP | |
| Similarities | Differences |
|---|---|
|
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghallab, A.; Myllys, M.; Friebel, A.; Duda, J.; Edlund, K.; Halilbasic, E.; Vucur, M.; Hobloss, Z.; Brackhagen, L.; Begher-Tibbe, B.; et al. Spatio-Temporal Multiscale Analysis of Western Diet-Fed Mice Reveals a Translationally Relevant Sequence of Events during NAFLD Progression. Cells 2021, 10, 2516. https://doi.org/10.3390/cells10102516
Ghallab A, Myllys M, Friebel A, Duda J, Edlund K, Halilbasic E, Vucur M, Hobloss Z, Brackhagen L, Begher-Tibbe B, et al. Spatio-Temporal Multiscale Analysis of Western Diet-Fed Mice Reveals a Translationally Relevant Sequence of Events during NAFLD Progression. Cells. 2021; 10(10):2516. https://doi.org/10.3390/cells10102516
Chicago/Turabian StyleGhallab, Ahmed, Maiju Myllys, Adrian Friebel, Julia Duda, Karolina Edlund, Emina Halilbasic, Mihael Vucur, Zaynab Hobloss, Lisa Brackhagen, Brigitte Begher-Tibbe, and et al. 2021. "Spatio-Temporal Multiscale Analysis of Western Diet-Fed Mice Reveals a Translationally Relevant Sequence of Events during NAFLD Progression" Cells 10, no. 10: 2516. https://doi.org/10.3390/cells10102516