
Long-Term Maintenance of Viable Adult Rat Sertoli Cells Able to Establish Testis Barrier Components and Function in Response to Androgens

 ,
,
Abstract
:1. Introduction
2. Material and Methods
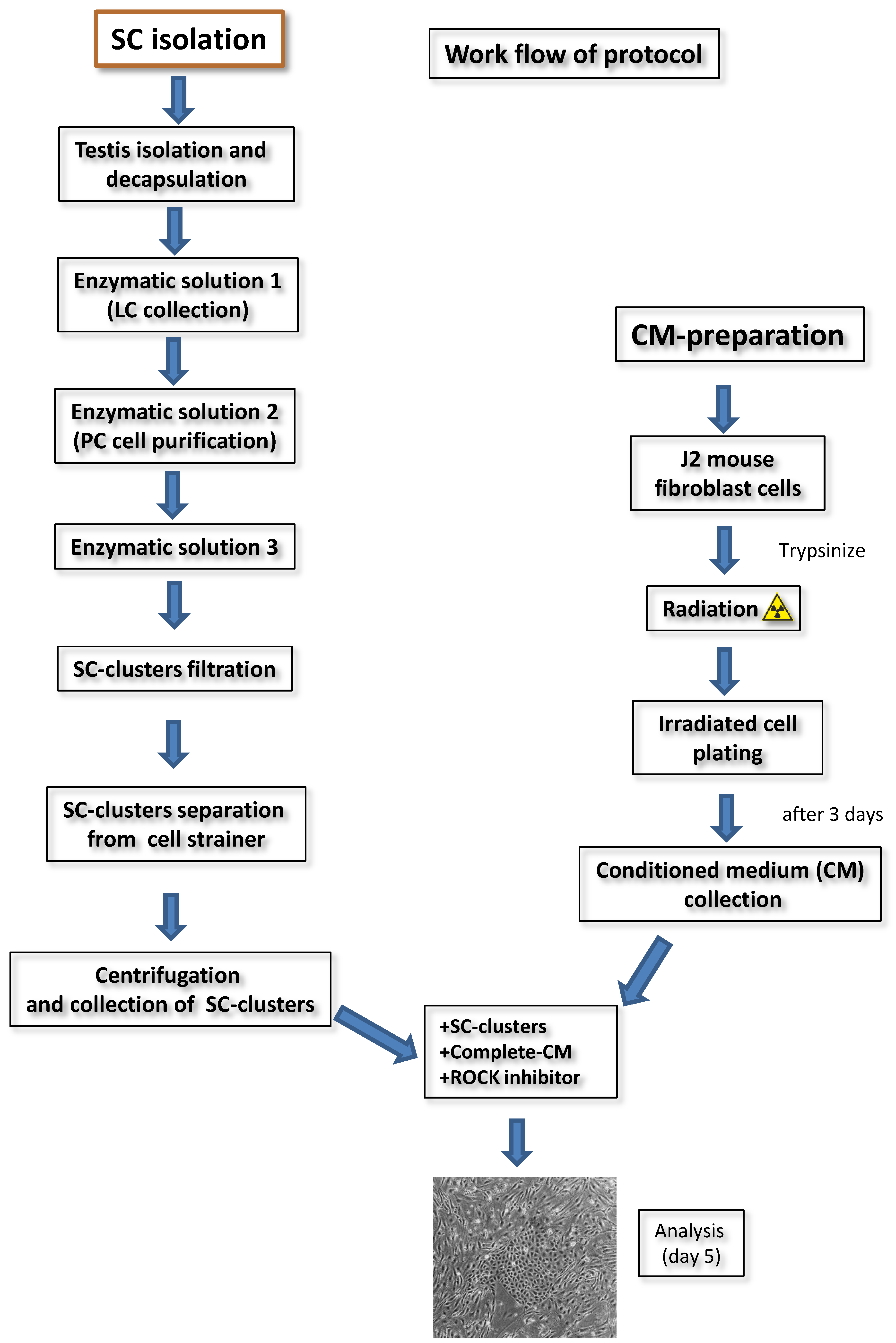
2.1. Isolation of Adult Rat SC
2.2. Irradiation of Feeder Cells and Preparation of Conditioned Medium (CM)
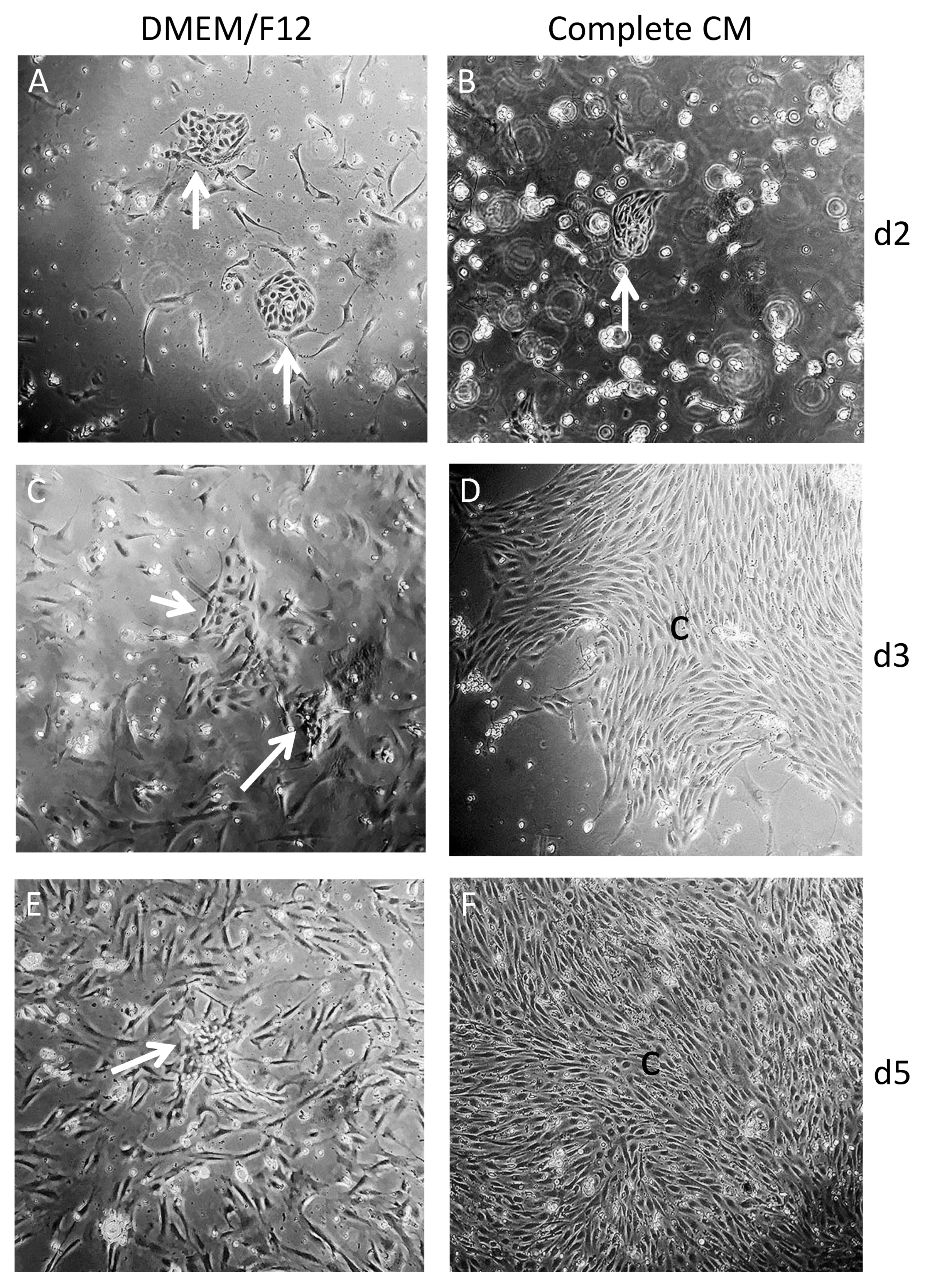
2.3. Culture of Freshly Isolated SC Clusters with CM
2.4. Immunofluorescence (IF)
2.5. RNA Extraction, RT-PCR, and Quantitative Real Time PCR
2.6. Measurement of Transepithelial Resistance (TER)
2.7. Tracer Diffusion Assay (TDA)
2.8. Statistical Analysis
3. Results
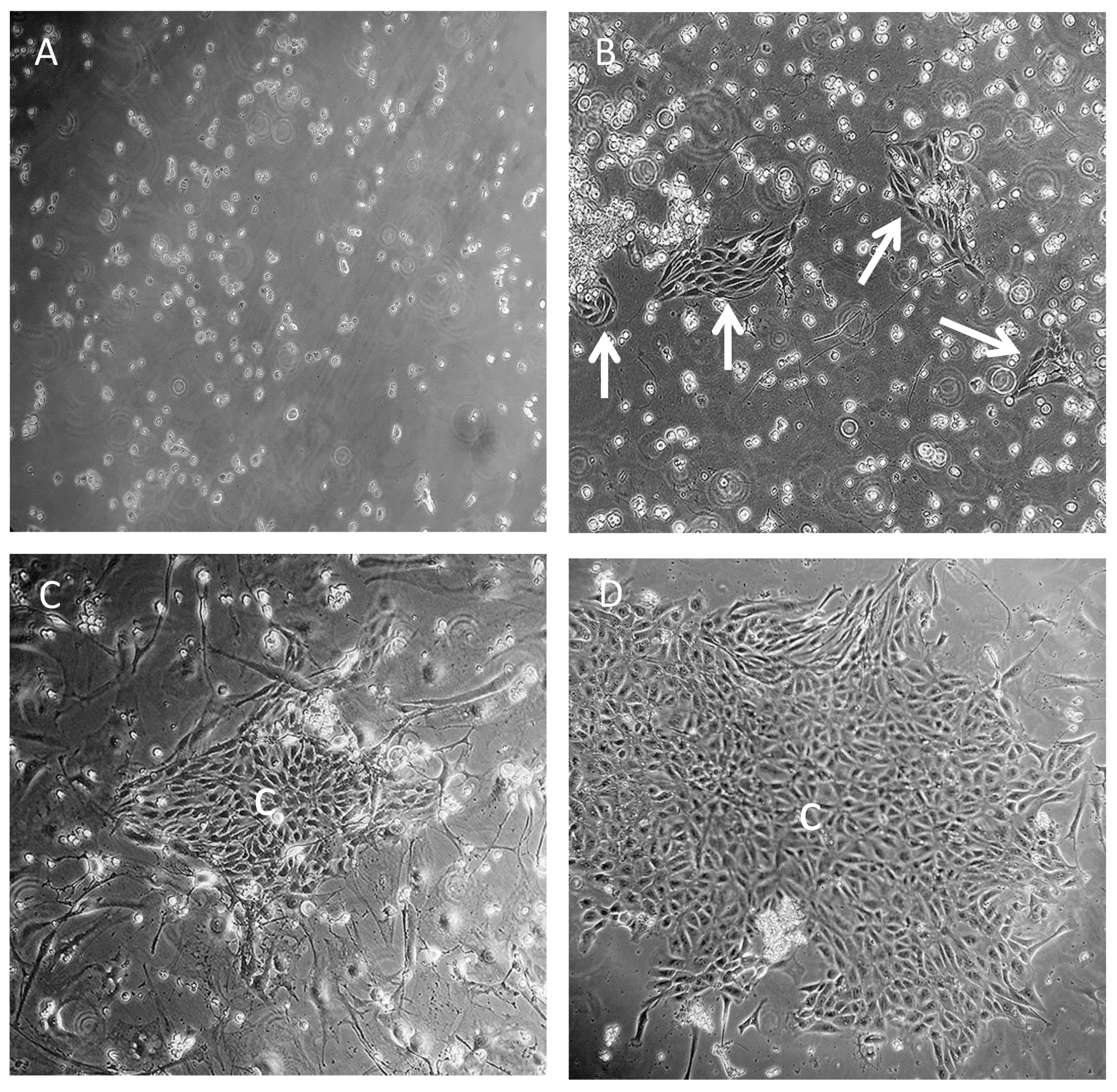
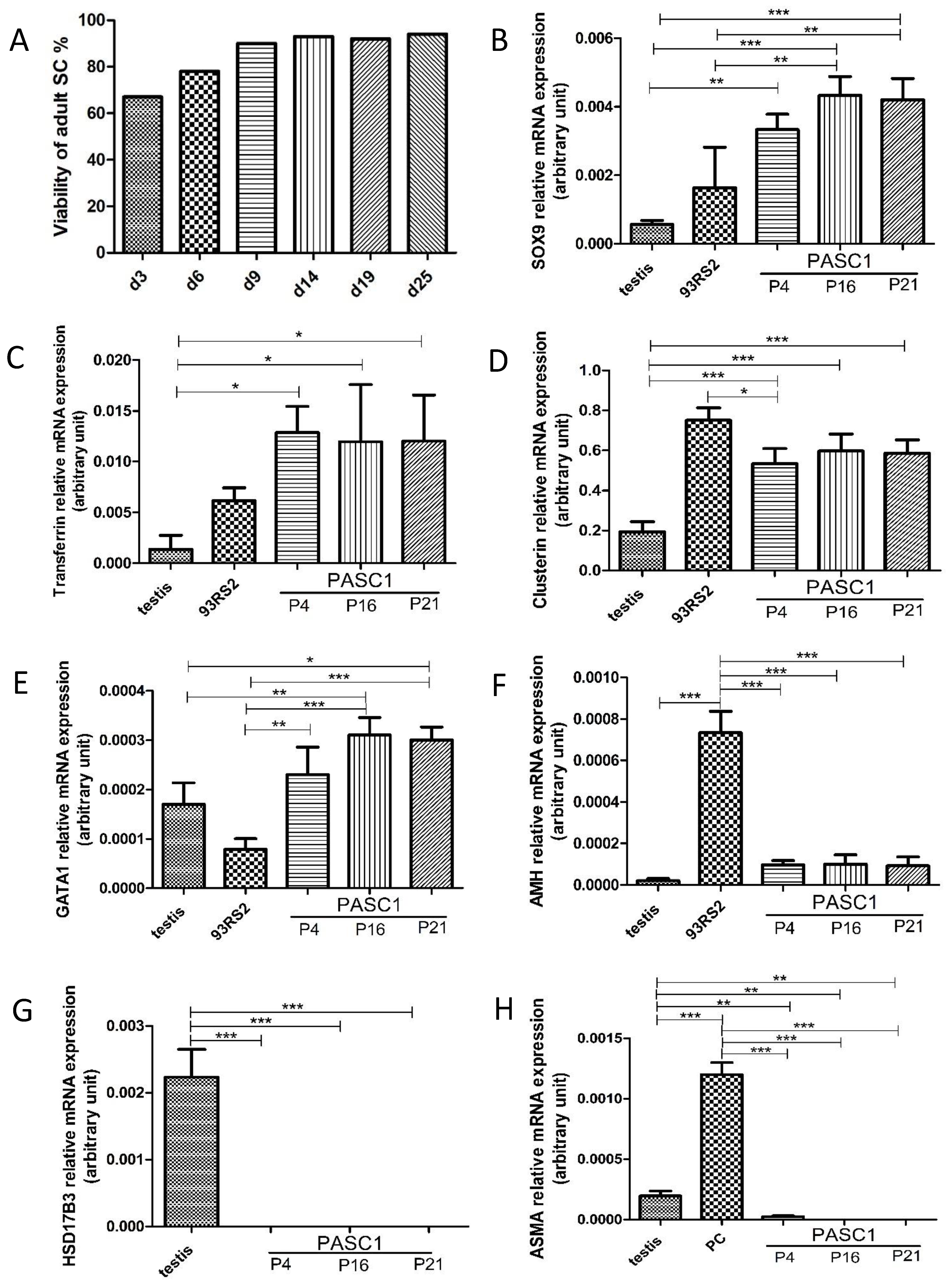
3.1. Isolation of Adult Rat SCs
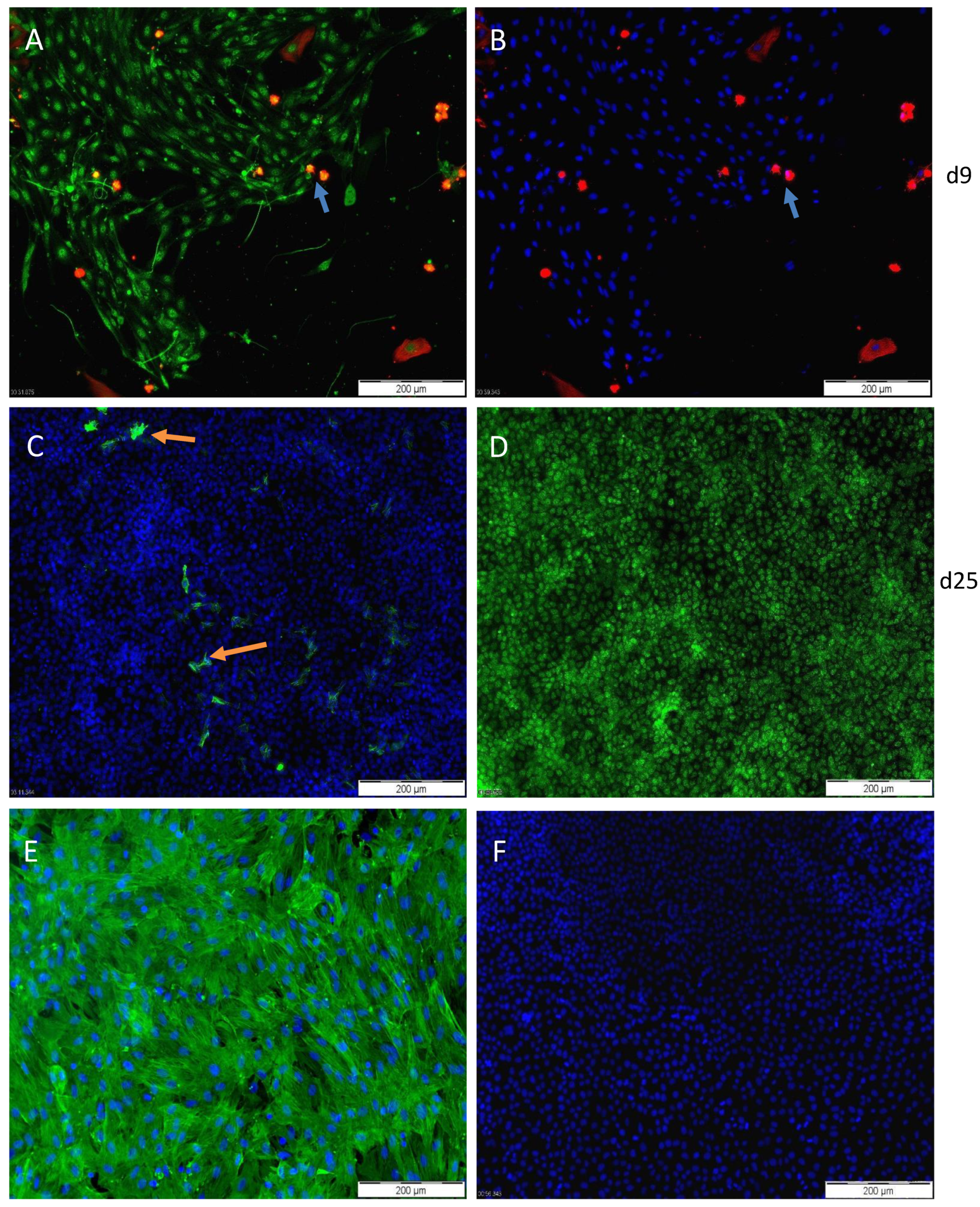
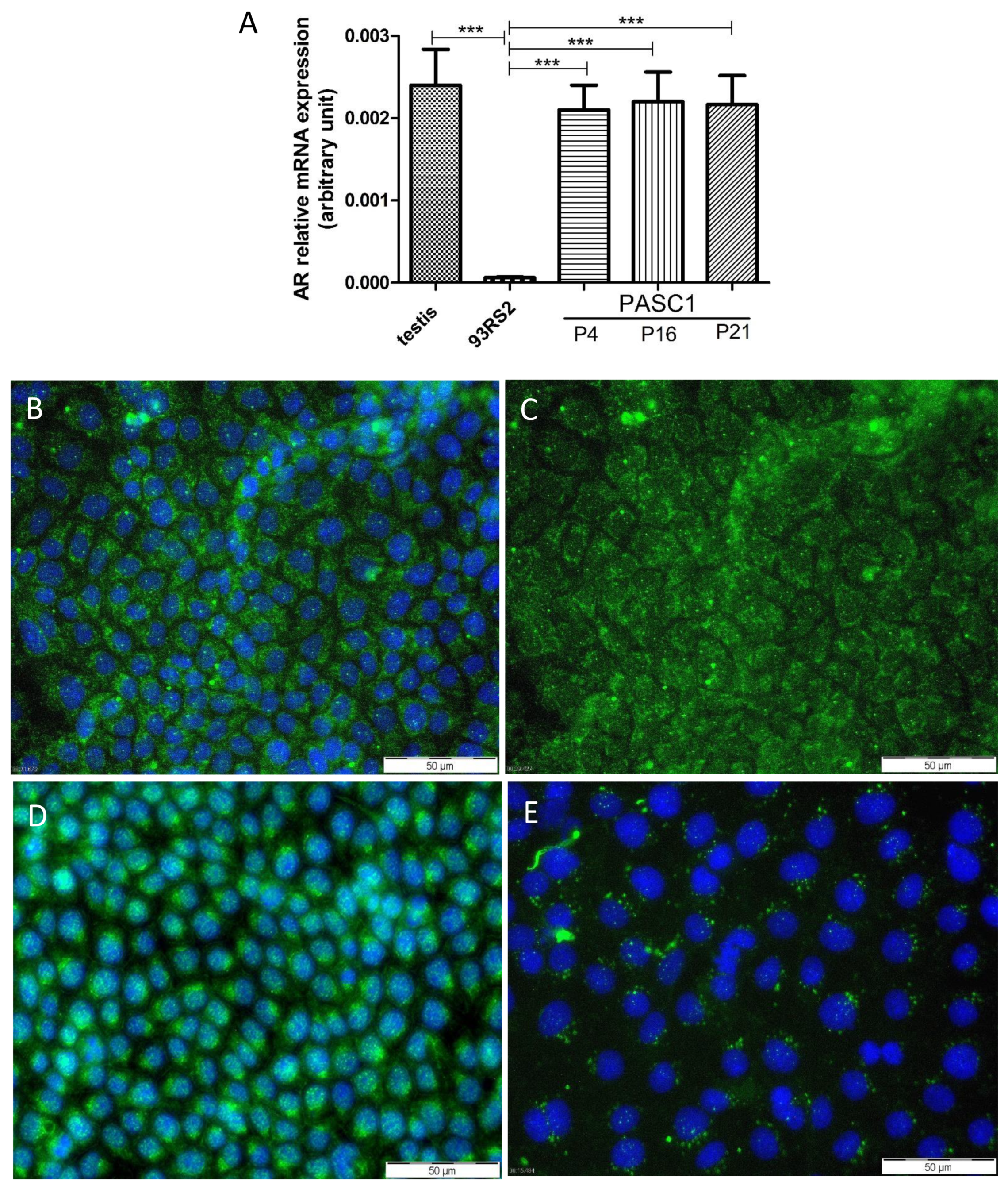
3.2. Characterization of PASC1—Expression of Testicular Markers
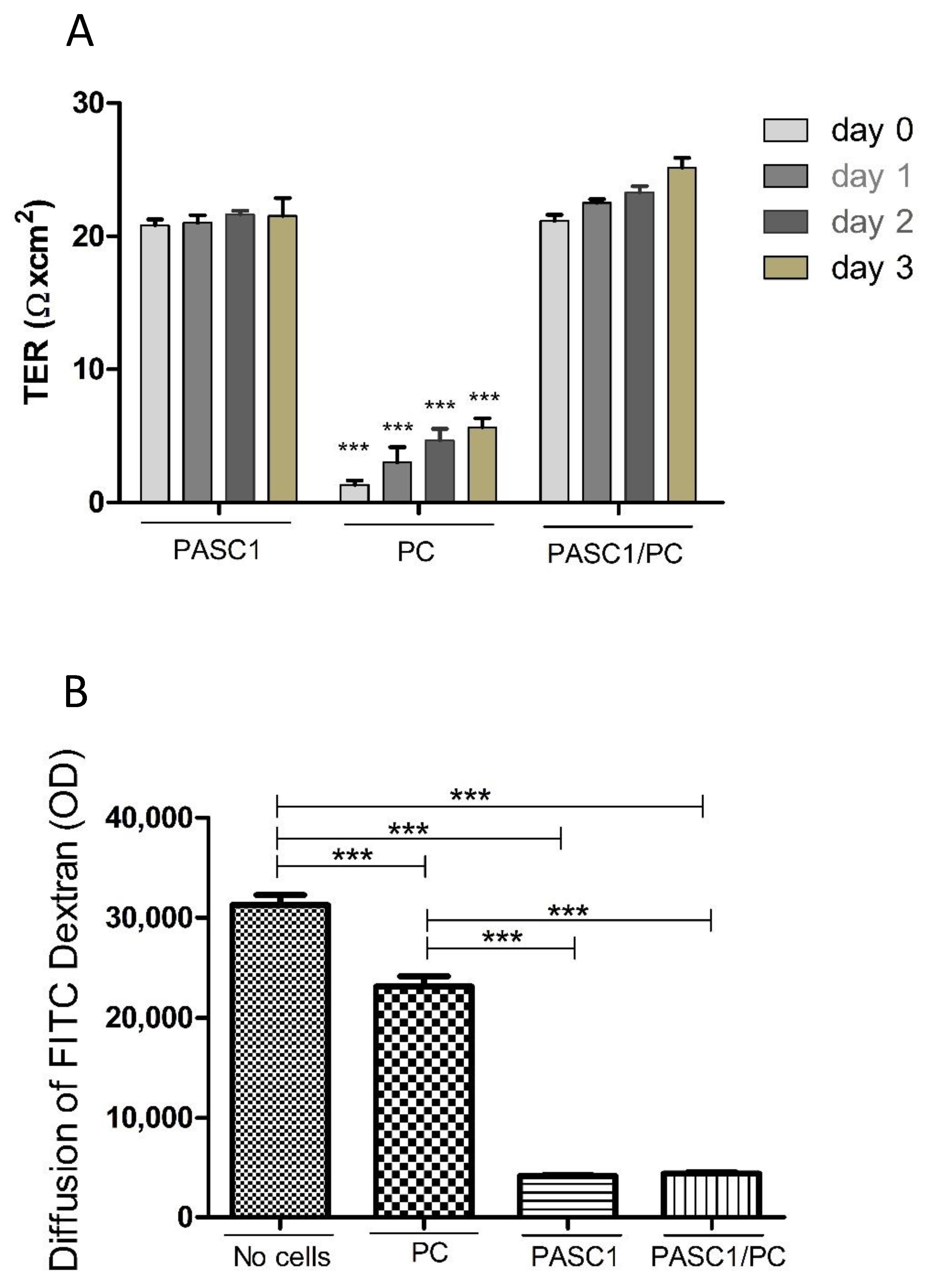
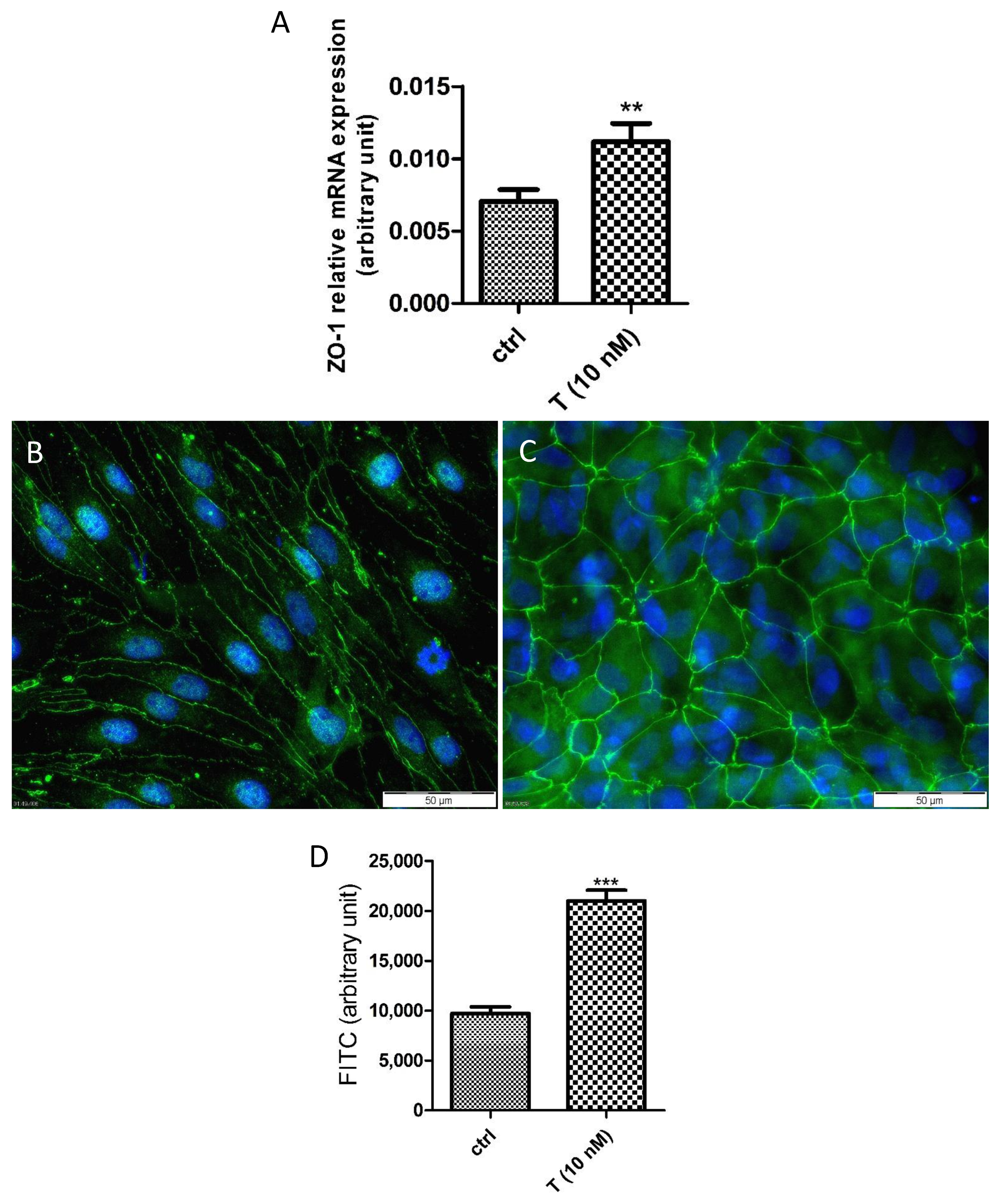
3.3. Functional Characterization of PASC1—Formation of the SC Barrier
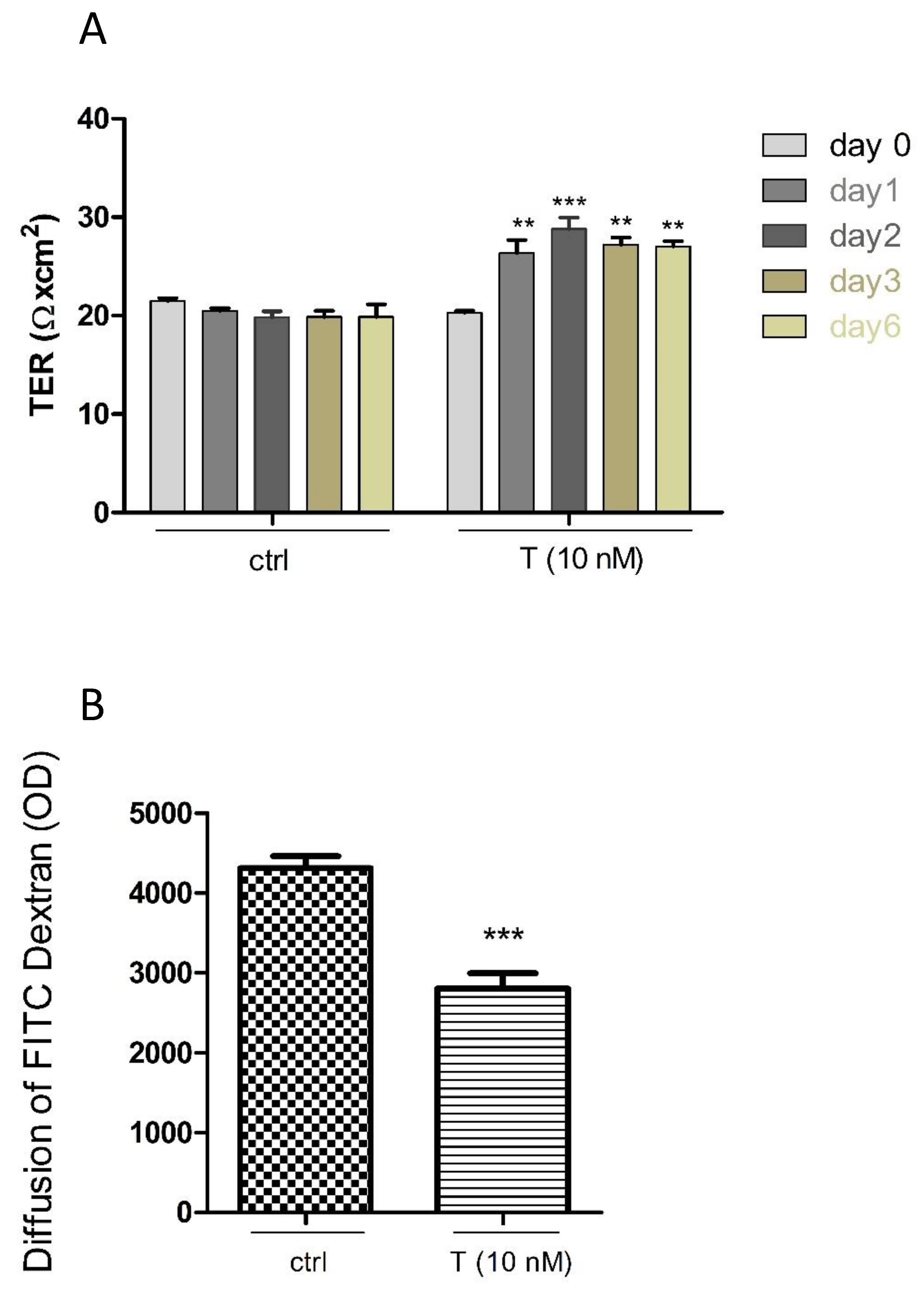
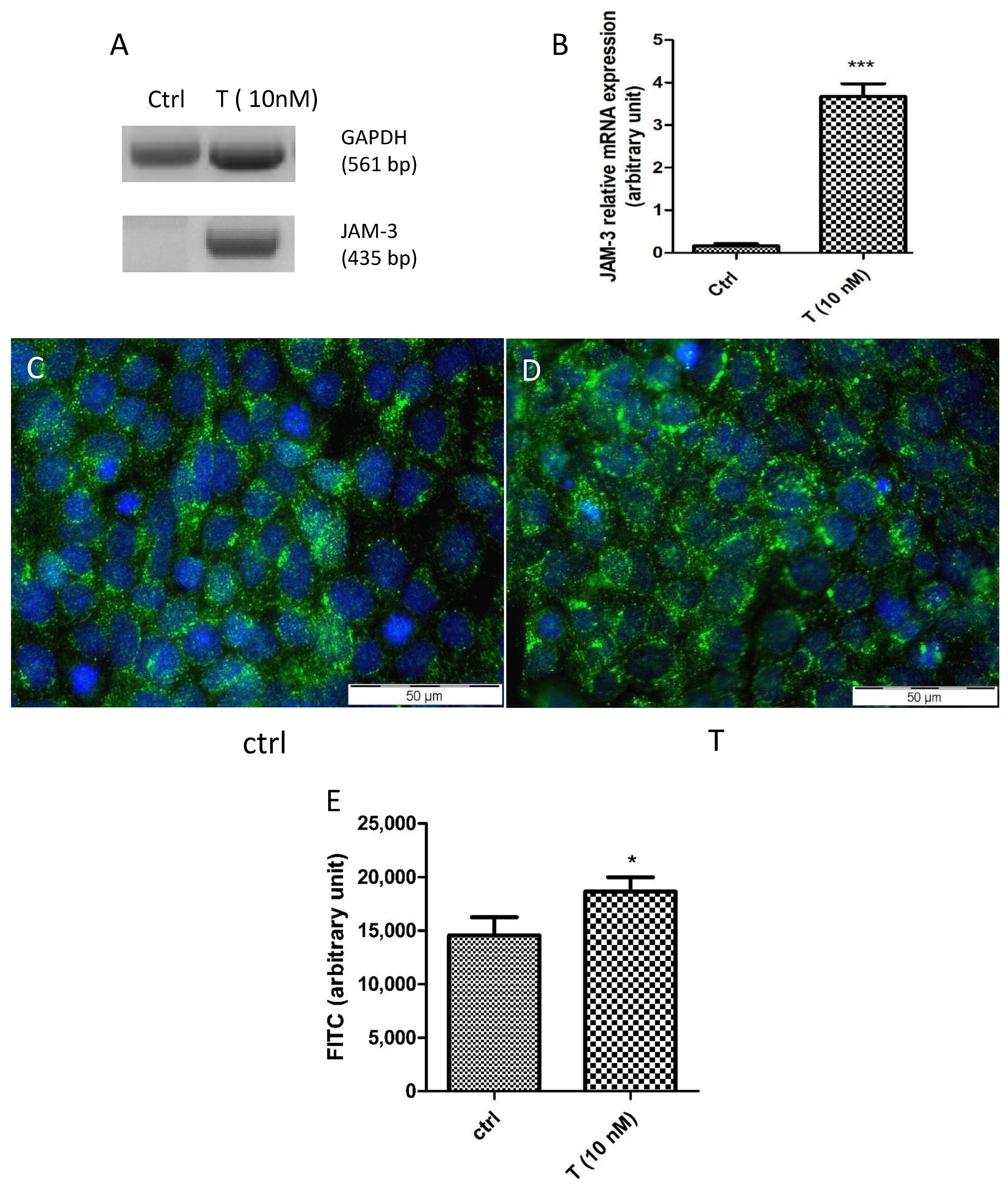
3.4. Effects of T on the PASC1 Barrier Integrity
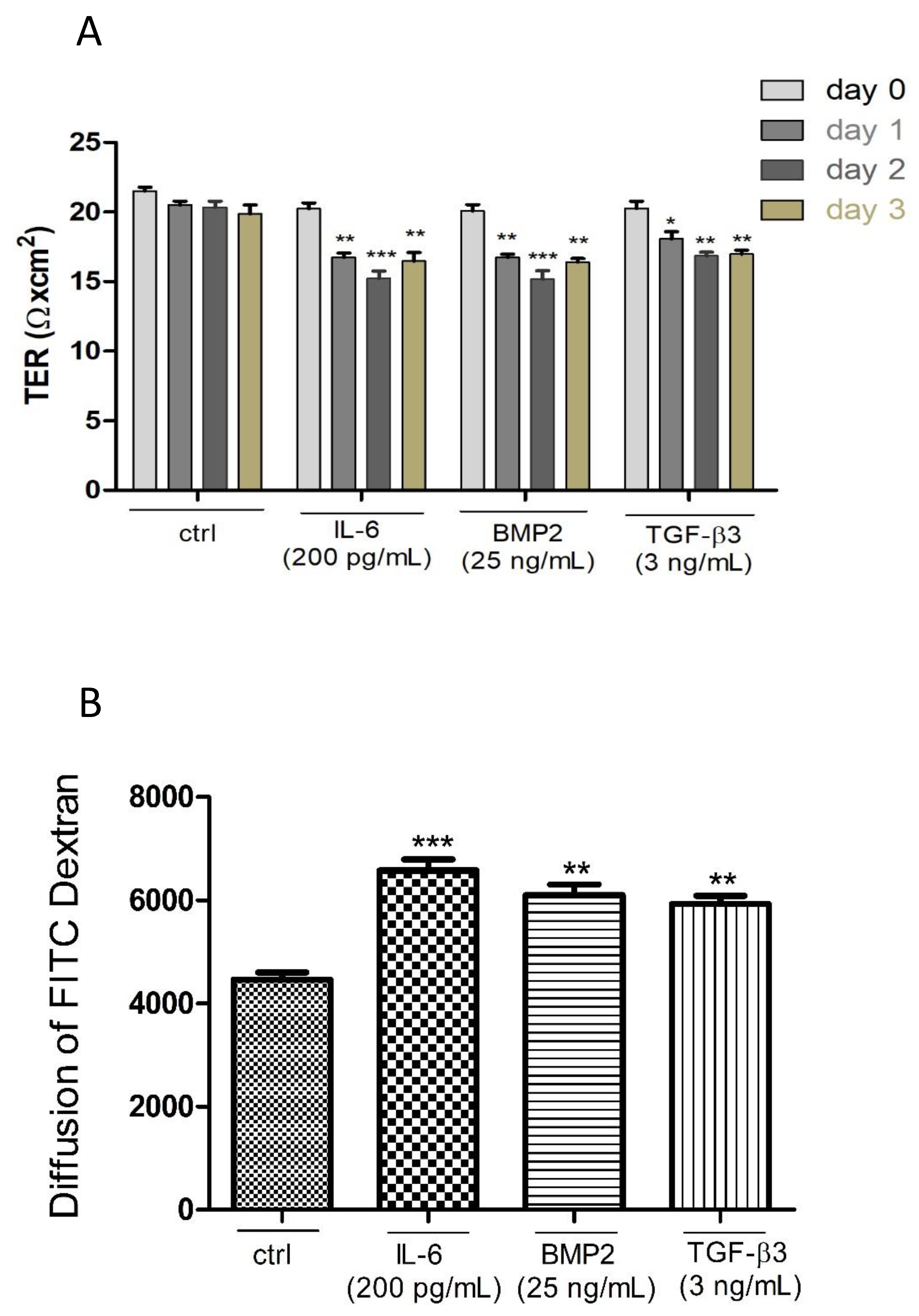
3.5. Effects of Different Cytokines on the TJ Barrier of PASC1
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zomer, H.D.; Reddi, P.P. Characterization of rodent Sertoli cell primary cultures. Mol. Reprod. Dev. 2020, 87, 857–870. [Google Scholar] [CrossRef]
- Karzai, A.W.; Wright, W.W. Regulation of the synthesis and secretion of transferrin and cyclic protein-2/cathepsin L by mature rat Sertoli cells in culture. Biol. Reprod. 1992, 47, 823–831. [Google Scholar] [CrossRef] [Green Version]
- Mays-Hoopes, L.L.; Bolen, J.; Riggs, A.D.; Singer-Sam, J. Preparation of spermatogonia, spermatocytes, and round spermatids for analysis of gene expression using fluorescence-activated cell sorting. Biol. Reprod. 1995, 53, 1003–1011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lakpour, M.R.; Aghajanpour, S.; Koruji, M.; Shahverdi, A.; Sadighi-Gilani, M.A.; Sabbaghian, M.; Aflatoonian, R.; Rajabian-Naghandar, M. Isolation, culture and characterization of human Sertoli cells by flow cytometry: Development of procedure. J. Reprod. Infertil. 2017, 18, 213–217. [Google Scholar] [PubMed]
- Kubota, H.; Avarbock, M.R.; Brinster, R.L. Spermatogonial stem cells share some, but not all, phenotypic and functional characteristics with other stem cells. Proc. Natl. Acad. Sci. USA 2003, 100, 6487–6492. [Google Scholar] [CrossRef] [Green Version]
- Hoeben, E.; Deboel, L.; Rombauts, L.; Heyns, W.; Verhoeven, G. Different cells and cell lines produce factors that modulate Sertoli cell function. Mol. Cell. Endocrinol. 1994, 101, 263–275. [Google Scholar] [CrossRef]
- Anway, M.D.; Folmer, J.; Wright, W.W.; Zirkin, B.R. Isolation of Sertoli cells from adult rat testes: An approach to ex vivo studies of Sertoli cell function. Biol. Reprod. 2003, 68, 996–1002. [Google Scholar] [CrossRef] [Green Version]
- Sato, Y.; Yoshida, K.; Nozawa, S.; Yoshiike, M.; Arai, M.; Otoi, T.; Iwamoto, T. Establishment of adult mouse Sertoli cell lines by using the starvation method. Reproduction 2013, 145, 505–516. [Google Scholar] [CrossRef] [Green Version]
- Bhushan, S.; Aslani, F.; Zhang, Z.; Sebastian, T.; Elsässer, H.P.; Klug, J. Isolation of Sertoli cells and peritubular cells from rat testes. J. Vis. Exp. 2016, 108, e53389. [Google Scholar] [CrossRef] [Green Version]
- Ali, S.H.; DeCaprio, J.A. Cellular transformation by SV40 large T antigen: Interaction with host proteins. Semin. Cancer Biol. 2001, 11, 15–23. [Google Scholar] [CrossRef]
- Konrad, L.; Munir Keilani, M.; Cordes, A.; Völck-Badouin, E.; Laible, L.; Albrecht, M.; Renneberg, H.; Aumüller, G. Rat Sertoli cells express epithelial but also mesenchymal genes after immortalization with SV40. Biochim. Biophys. Acta 2005, 1722, 6–14. [Google Scholar] [CrossRef]
- Chapman, S.; Liu, X.; Meyers, C.; Schlegel, R.; McBride, A.A. Human keratinocytes are efficiently immortalized by a Rho kinase inhibitor. J. Clin. Investig. 2010, 120, 2619–2626. [Google Scholar] [CrossRef] [Green Version]
- Suprynowicz, F.A.; Upadhyay, G.; Krawczyk, E.; Kramer, S.C.; Hebert, J.D.; Liu, X.; Yuan, H.; Cheluvaraju, C.; Clapp, P.W.; Boucher, R.C., Jr.; et al. Conditionally reprogrammed cells represent a stem-like state of adult epithelial cells. Proc. Natl. Acad. Sci. USA 2012, 109, 20035–20040. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Ory, V.; Chapman, S.; Yuan, H.; Albanese, C.; Kallakury, B.; Timofeeva, O.; Nealon, C.; Dakic, A.; Simic, V.; et al. ROCK inhibitor and feeder cells induce the conditional reprogramming of epithelial cells. Am. J. Pathol. 2012, 180, 599–607. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Krawczyk, E.; Suprynowicz, F.; Palechor-Ceron, N.; Yuan, H.; Dakic, A.; Simic, V.; Zheng, Y.L.; Sripadhan, P.; Chen, C.; et al. Conditional reprogramming and long-term expansion of normal and tumor cells from human biospecimens. Nat. Protoc. 2017, 12, 439–451. [Google Scholar] [CrossRef]
- Terunuma, A.; Limgala, R.P.; Park, C.J.; Choudhary, I.; Vogel, J.C. Efficient procurement of epithelial stem cells from human tissue specimens using a Rho-associated protein kinase inhibitor Y-27632. Tissue Eng. Part A 2010, 16, 1363–1368. [Google Scholar] [CrossRef] [Green Version]
- Rheinwald, J.G.; Green, H. Serial cultivation of strains of human epidermal keratinocytes: The formation of keratinizing colonies from single cells. Cell 1975, 6, 331–343. [Google Scholar] [CrossRef]
- Yue, J.; Shukla, R.; Accardi, R.; Zanella-Cleon, I.; Siouda, M.; Cros, M.P.; Krutovskikh, V.; Hussain, I.; Niu, Y.; Hu, S.; et al. Cutaneous human papillomavirus type 38 E7 regulates actin cytoskeleton structure for increasing cell proliferation through CK2 and the eukaryotic elongation factor 1A. J. Virol. 2011, 85, 8477–8494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mruk, D.D.; Cheng, C.J. The mammalian blood-testis barrier: Its biology and regulation. Endocr. Rev. 2015, 36, 564–591. [Google Scholar] [CrossRef] [PubMed]
- Ni, F.D.; Hao, S.L.; Yang, W.X. Molecular insights into hormone regulation via signaling pathways in Sertoli cells: With discussion on infertility and testicular tumor. Gene 2020, 753, 144812. [Google Scholar] [CrossRef] [PubMed]
- Linn, E.; Ghanem, L.; Bhakta, H.; Greer, C.; Avella, M. Genes regulating spermatogenesis and sperm function associated with rare disorders. Front. Cell Dev. Biol. 2021, 9, 634536. [Google Scholar] [CrossRef]
- Washburn, R.L.; Hibler, T.; Thompson, L.A.; Kaur, G.; Dufour, J.M. Therapeutic application of Sertoli cells for treatment of various diseases. Semin. Cell Dev. Biol. 2021, 25, S1084-9521(21)00080-X. [Google Scholar]
- Bulldan, A.; Dietze, R.; Shihan, M.; Scheiner-Bobis, G. Non-classical testosterone signaling mediated through ZIP9 stimulates claudin expression and tight junction formation in Sertoli cells. Cell. Signal. 2016, 28, 1075–1085. [Google Scholar] [CrossRef]
- Oduwole, O.O.; Peltoketo, H.; Poliandri, A.; Vengadabady, L.; Chrusciel, M.; Doroszko, M.; Samanta, L.; Owen, L.; Keevil, B.; Rahman, N.A.; et al. Constitutively active follicle-stimulating hormone receptor enables androgen-independent spermatogenesis. J. Clin. Investig. 2018, 128, 1787–1792. [Google Scholar] [CrossRef] [Green Version]
- Karl, A.F.; Griswold, M.D. Sertoli cells of the testis: Preparation of cell cultures and effects of retinoids. Methods Enzymol. 1990, 190, 71–75. [Google Scholar] [PubMed]
- Sneddon, S.F.; Walther, N.; Saunders, P.T.K. Expression of androgen and estrogen receptors in Sertoli cells: Studies using the SK11 cell line. Endocrinology 2005, 146, 5304–5312. [Google Scholar] [CrossRef] [Green Version]
- Jiang, C.; Hall, S.H.; Boekelheide, K. Development and characterization of a prepubertal rat Sertoli cell line, 93RS2. J. Androl. 1997, 18, 393–399. [Google Scholar] [PubMed]
- Lee, H.J.; Lee, S.M.; Moon, Y.G.; Jung, Y.S.; Lee, J.H.; Saralamma, V.V.G.; Kim, Y.S.; Pak, J.E.; Lee, H.J.; Kim, G.S.; et al. Membrane-free stem cell components inhibit interleukin-1α-stimulated inflammation and cartilage degradation in vitro and in vivo: A rat model of osteoarthritis. Int. J. Mol. Sci. 2019, 20, 4869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stammler, A.; Müller, D.; Tabuchi, Y.; Konrad, L.; Middendorff, R. TGFβs modulate paracellular permeability in a new in vitro model of the blood epididymal barrier. PLoS ONE 2013, 8, e80611. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.F.; Lee-Chang, J.S.; Panneerdoss, S.; MacLean, J.A., II; Rao, M.K. Isolation of Sertoli, Leydig, and spermatogenic cells from the mouse testis. Biotechniques 2011, 51, 341–342, 344. [Google Scholar] [CrossRef] [Green Version]
- Skinner, M.K.; Griswold, M.D. Sertoli cells synthesize and secrete transferrin-like protein. J. Biol. Chem. 1980, 255, 9523–9525. [Google Scholar] [CrossRef]
- Bailey, R.; Griswold, M.D. Clusterin in the male reproductive system: Localization and possible function. Mol. Cell. Endocrinol. 1995, 151, 17–23. [Google Scholar] [CrossRef] [Green Version]
- Fröjdman, K.; Harley, V.; Pelliniemi, L. Sox9 protein in rat Sertoli cells is age and stage dependent. Histochemistry 2000, 113, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Jégou, B.; Sharpe, R.M. Paracrine mechanisms in testicular control. In Molecular Biology of the Male Reproductive System; De Kretser, D.M., Ed.; Academic Press: New York, NY, USA, 1993; pp. 271–310. [Google Scholar]
- Yomogida, K.; Ohtani, H.; Harigae, H.; Ito, E.; Nishimune, Y.; Engel, J.D.; Yamamoto, M. Developmental stage- and spermatogenic cycle-specific expression of transcription factor GATA-1 in mouse Sertoli cells. Development 1994, 120, 1759–1766. [Google Scholar] [CrossRef]
- Beau, C.; Rauch, M.; Joulin, V.; Jégou, B.; Guerrier, D. GATA-1 is a potential repressor of anti-Müllerian hormone expression during the establishment of puberty in the mouse. Mol. Reprod. Dev. 2000, 56, 124–138. [Google Scholar] [CrossRef]
- Chakraborty, P.; Buaas, F.W.; Sharma, M.; Smith, B.E.; Greenlee, A.R.; Eacker, S.M.; Braun, R.E. Androgen-dependent Sertoli cell tight junction remodeling is mediated by multiple tight junction components. Mol. Endocrinol. 2014, 29, 1055–1072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Regadera, J.; Martínez-García, F.; González-Peramato, P.; Serrano, A.; Nistal, M.; Suárez-Quian, C. Androgen receptor expression in Sertoli cells as a function of seminiferous tubule maturation in the human cryptorchid testis. J. Clin. Endocrinol. Metab. 2001, 86, 413–421. [Google Scholar] [CrossRef]
- Bulldan, A.; Malviya, V.N.; Upmanyu, N.; Konrad, L.; Scheiner-Bobis, G. Testosterone/bicalutamide antagonism at the predicted extracellular androgen binding site of ZIP9. Biochim. Biophys. Acta-Mol. Cell Res. 2017, 1864, 2402–2414. [Google Scholar] [CrossRef]
- Bulldan, A.; Bartsch, J.W.; Konrad, L.; Scheiner-Bobis, G. ZIP9 but not the androgen receptor mediates testosterone-induced migratory activity of metastatic prostate cancer cells. Biochim. Biophys. Acta-Mol. Cell Res. 2018, 1865, 1857–1868. [Google Scholar] [CrossRef] [PubMed]
- Gliki, G.; Ebnet, K.; Aurrand-Lions, M.; Imhof, B.A.; Adams, R.H. Spermatid differentiation requires the assembly of a cell polarity complex downstream of junctional adhesion molecule-C. Nature 2004, 431, 320–324. [Google Scholar] [CrossRef]
- Willems, A.; Batlouni, S.R.; Esnal, A.; Swinnen, J.V.; Saunders, P.T.K.; Sharpe, R.M.; Franca, L.R.; De Gendt, K.; Verhoeven, G. Selective ablation of the androgen receptor in mouse Sertoli cells affects Sertoli cell maturation, barrier formation and cytoskeletal development. PLoS ONE 2010, 5, e14168. [Google Scholar] [CrossRef] [Green Version]
- Lui, W.Y.; Cheng, C.Y. Transcriptional regulation of cell adhesion at the blood-testis barrier and spermatogenesis in the testis. Adv. Exp. Med. Biol. 2012, 763, 281–294. [Google Scholar]
- De Gendt, K.; Verhoeven, G.; Amieux, P.S.; Wilkinson, M.F. Genome-wide identification of AR-regulated genes translated in Sertoli cells in vivo using the RiboTag approach. Mol. Endocrinol. 2014, 28, 575–591. [Google Scholar] [CrossRef] [Green Version]
- Xia, W.; Wong, E.W.; Mruk, D.D.; Cheng, C.Y. TGF-beta3 and TNFα perturb blood-testis barrier (BTB) dynamics by accelerating the clathrin-mediated endocytosis of integral membrane proteins: A new concept of BTB regulation during spermatogenesis. Dev. Biol. 2009, 327, 48–61. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Yin, Y.; Wang, G.; Liu, Z.; Liu, L.; Sun, F. Interleukin-6 disrupts blood-testis barrier through inhibiting protein degradation or activating phosphorylated ERK in Sertoli cells. Sci. Rep. 2014, 4, 4260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lui, W.Y.; Lee, W.M.; Cheng, C.Y. Transforming growth factor beta3 regulates the dynamics of Sertoli cell tight junctions via the p38 mitogen-activated protein kinase pathway. Biol. Reprod. 2003, 68, 1597–1612. [Google Scholar] [CrossRef]
- Lui, W.Y.; Lee, W.M.; Cheng, C.Y. Transforming growth factor-β3 perturbs the inter-Sertoli tight junction permeability barrier in vitro possibly mediated via its effects on occludin, zonula occludens-1, and claudin-11. Endocrinology 2001, 142, 1865–1877. [Google Scholar] [CrossRef] [PubMed]
- Ciller, I.M.; Palanisamy, S.K.; Ciller, U.A.; McFarlane, J.R. Postnatal expression of bone morphogenetic proteins and their receptors in the mouse testis. Physiol. Res. 2016, 65, 673–682. [Google Scholar] [CrossRef] [PubMed]
- Gautam, M.; Bhattacharya, I.; Devi, Y.S.; Arya, S.P.; Majumdar, S.S. Hormone responsiveness of cultured Sertoli cells obtained from adult rats after their rapid isolation under less harsh conditions. Andrology 2016, 4, 509–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saenz, F.R.; Ory, V.; AlOtaiby, M.; Rosenfield, S.; Furlong, M.; Cavalli, L.R.; Johnson, M.D.; Liu, X.; Schlegel, R.; Wellstein, A.; et al. Conditionally reprogrammed normal and transformed mouse mammary epithelial cells display a progenitor-cell-like phenotype. PLoS ONE 2014, 9, e97666. [Google Scholar] [CrossRef]
- Hynds, R.E.; Vladimirou, E.; Janes, S.M. The secret lives of cancer cell lines. Dis. Model Mech. 2018, 11, dmm037366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shay, J.W.; Wright, W.E. Senescence and immortalization: Role of telomeres and telomerase. Carcinogenesis 2005, 26, 867–874. [Google Scholar] [CrossRef] [PubMed]
- Ailenberg, M.; Tung, P.S.; Pelletier, M.; Fritz, I.B. Modulation of Sertoli cell functions in the two-chamber assembly by peritubular cells and extracellular matrix. Endocrinology 1988, 122, 2604–2612. [Google Scholar] [CrossRef]
- Legendre, A.; Froment, P.; Desmots, S.; Lecomte, A.; Habert, R.; Lemazurier, E. An engineered 3D blood-testis barrier model for the assessment of reproductive toxicity potential. Biomaterials 2010, 31, 4492–4505. [Google Scholar] [CrossRef]
- Su, L.; Wang, Z.; Xie, S.; Hu, D.; Cheng, Y.C.; Mruk, D.D.; Guan, Y. Testin regulates the blood-testis barrier via disturbing occludin/ZO-1 association and actin organization. J. Cell. Physiol. 2020, 235, 6127–6138. [Google Scholar] [CrossRef]
- Ghafouri-Fard, S.; Shoorei, H.; Mohaqiq, M.; Raza, S.H.A.; Taheri, M. The role of different compounds on the integrity of blood-testis barrier: A concise review based on in vitro and in vivo studies. Gene 2021, 780, 145531. [Google Scholar] [CrossRef]
- Puglisi, R.; Montanari, M.; Chiarella, P.; Stefanini, M.; Boitani, C. Regulatory role of BMP2 and BMP7 in spermatogonia and Sertoli cell proliferation in the immature mouse. Eur. J. Endocrinol. 2004, 151, 511–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Itman, C.; Loveland, K.L. SMAD expression in the testis: An insight into BMP regulation of spermatogenesis. Dev. Dyn. 2008, 237, 97–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, K.; Xie, W.; Luo, B.; Xiao, W.; Teitelbaum, D.H.; Yang, H.; Zhang, K.; Zhang, C. Intestinal mucosal barrier is injured by BMP2/4 via activation of NF-κB signals after ischemic reperfusion. Mediators Inflamm. 2014, 2014, 901530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helbing, T.; Herold, E.M.; Hornstein, A.; Wintrich, S.; Heinke, J.; Grundmann, S.; Patterson, C.; Bode, C.; Moser, M. Inhibition of BMP activity protects epithelial barrier function in lung injury. J. Pathol. 2013, 231, 105–116. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes (Species) and Acc. No. | Sequence (5′ -> 3′) | AT | Size (bp) |
|---|---|---|---|
| GAPDH (rat) XM_039107008.1 | GACCCCTTCATTGACCTCAAC fwd GATGACCTTGCCCACAGCCTT rev | 59 °C | 561 |
| ASMA (rat) NM_031004.2 | GAAGAGGAAGACAGCACAGC fwd TTGGTGATGATGCCGTGTTC rev | 58 °C | 233 |
| SOX9 (rat) [28] NM_080403.2 | CATCAAGACGGAGCAACTGAG fwd GTGGTCGGTGTAGTCATACTGC rev | 59 °C | 148 |
| Transferrin [11] NM_001013110.1 | TATTGGCCCAGCAAAATGTG fwd CCGGAACAAACAGAAATTGC rev | 59 °C | 370 |
| Clusterin (rat) [11] XM_039092999.1 | AGGAGCTAAACGACTCGCT fwd GCTTTTCCTGCGGTATTCC rev | 59 °C | 362 |
| GATA1 (rat) NM_012764.2 | ATAGCAAGACGGCGCTCTAC fwd CACTCTCTGGCCTCACAAGG rev | 59 °C | 319 |
| AMH (rat) NM_012902.1 | AACTGACCAATACCAGGGGC fwd GGCTCCCATATCACTTCAGCC rev | 59 °C | 334 |
| AR (rat) [11] NM_012502.2 | GCCAGTGGCTGAGGATGAG fwd GGTGAGCTGGTAGAAGCGC rev | 59 °C | 236 |
| JAM-3 (rat) NM_001004269.1 | CTTCTTCCTGCTGCTGCTCT fwd TCTTGGCATTGCAGTGTTGC rev | 59 °C | 435 |
| HSD17B3 (rat) NM_054007 | GGAAGCCGTGTGAAGGTT fwd GACACTCTGGCTCTCACC rev | 58 °C | 171 |
| ZO-1 (rat) XM_0391053461 | CTTGCCACACTGTGACCCTA fwdGGGGCATGCTCACTAACCTT rev | 58 °C | 262 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kabbesh, H.; Riaz, M.A.; Jensen, A.D.; Scheiner-Bobis, G.; Konrad, L. Long-Term Maintenance of Viable Adult Rat Sertoli Cells Able to Establish Testis Barrier Components and Function in Response to Androgens. Cells 2021, 10, 2405. https://doi.org/10.3390/cells10092405
Kabbesh H, Riaz MA, Jensen AD, Scheiner-Bobis G, Konrad L. Long-Term Maintenance of Viable Adult Rat Sertoli Cells Able to Establish Testis Barrier Components and Function in Response to Androgens. Cells. 2021; 10(9):2405. https://doi.org/10.3390/cells10092405
Chicago/Turabian StyleKabbesh, Hassan, Muhammad A. Riaz, Alexandra D. Jensen, Georgios Scheiner-Bobis, and Lutz Konrad. 2021. "Long-Term Maintenance of Viable Adult Rat Sertoli Cells Able to Establish Testis Barrier Components and Function in Response to Androgens" Cells 10, no. 9: 2405. https://doi.org/10.3390/cells10092405