
Biomechanics of Ex Vivo-Generated Red Blood Cells Investigated by Optical Tweezers and Digital Holographic Microscopy

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
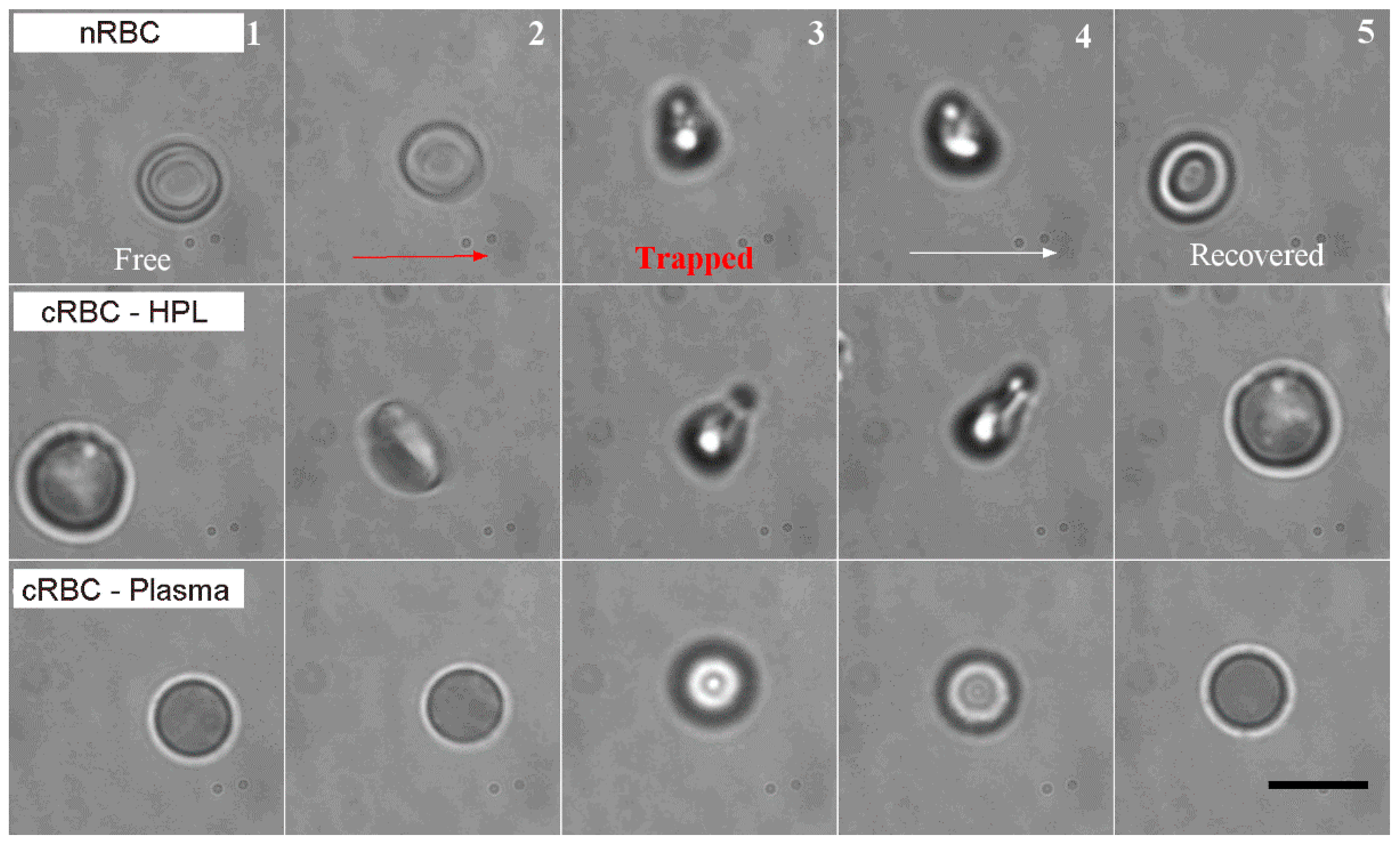
2.1. Cell Preparation
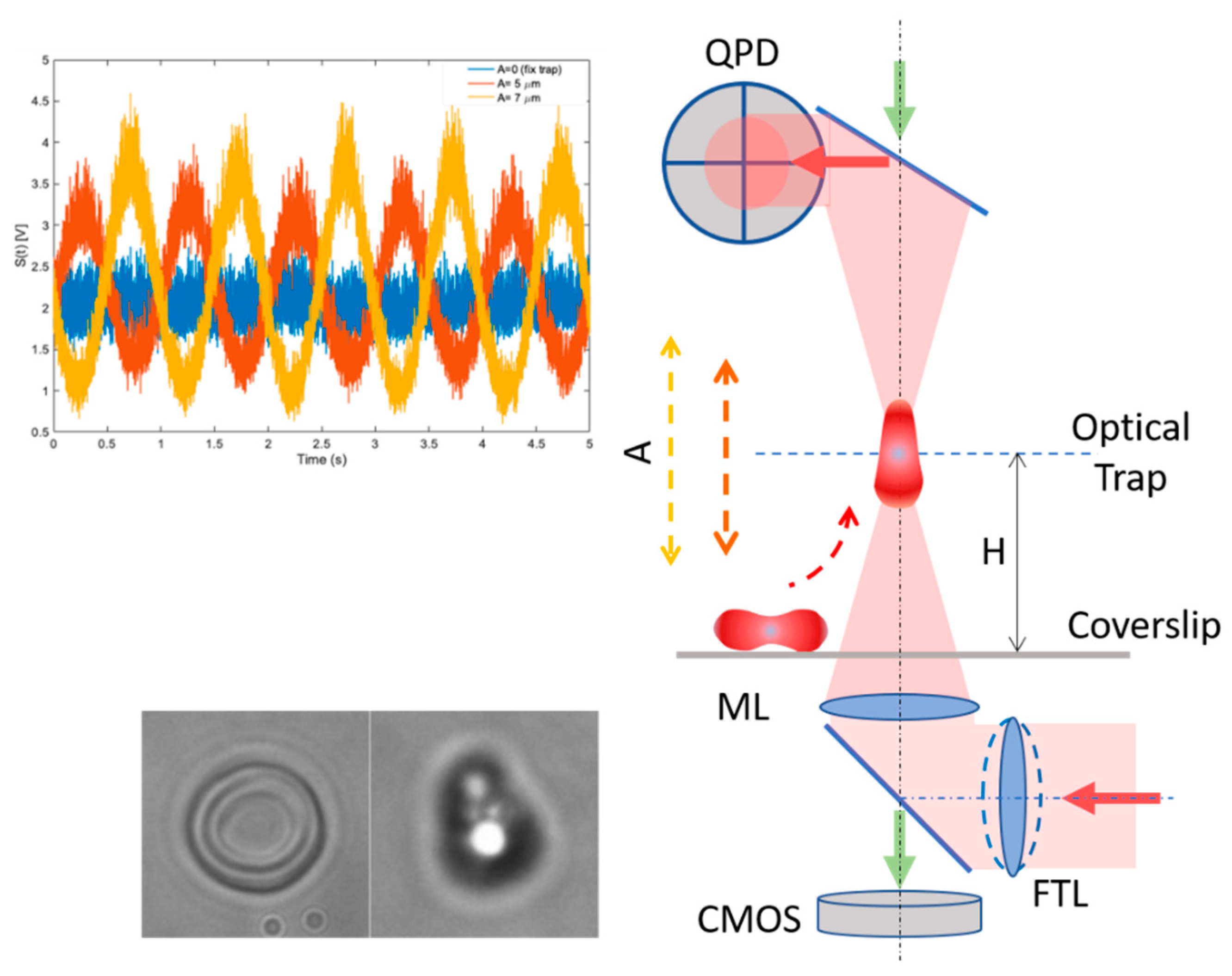
2.2. Cell Deformation and Cell Membrane Fluctuations in an Oscillatory Optical Trap
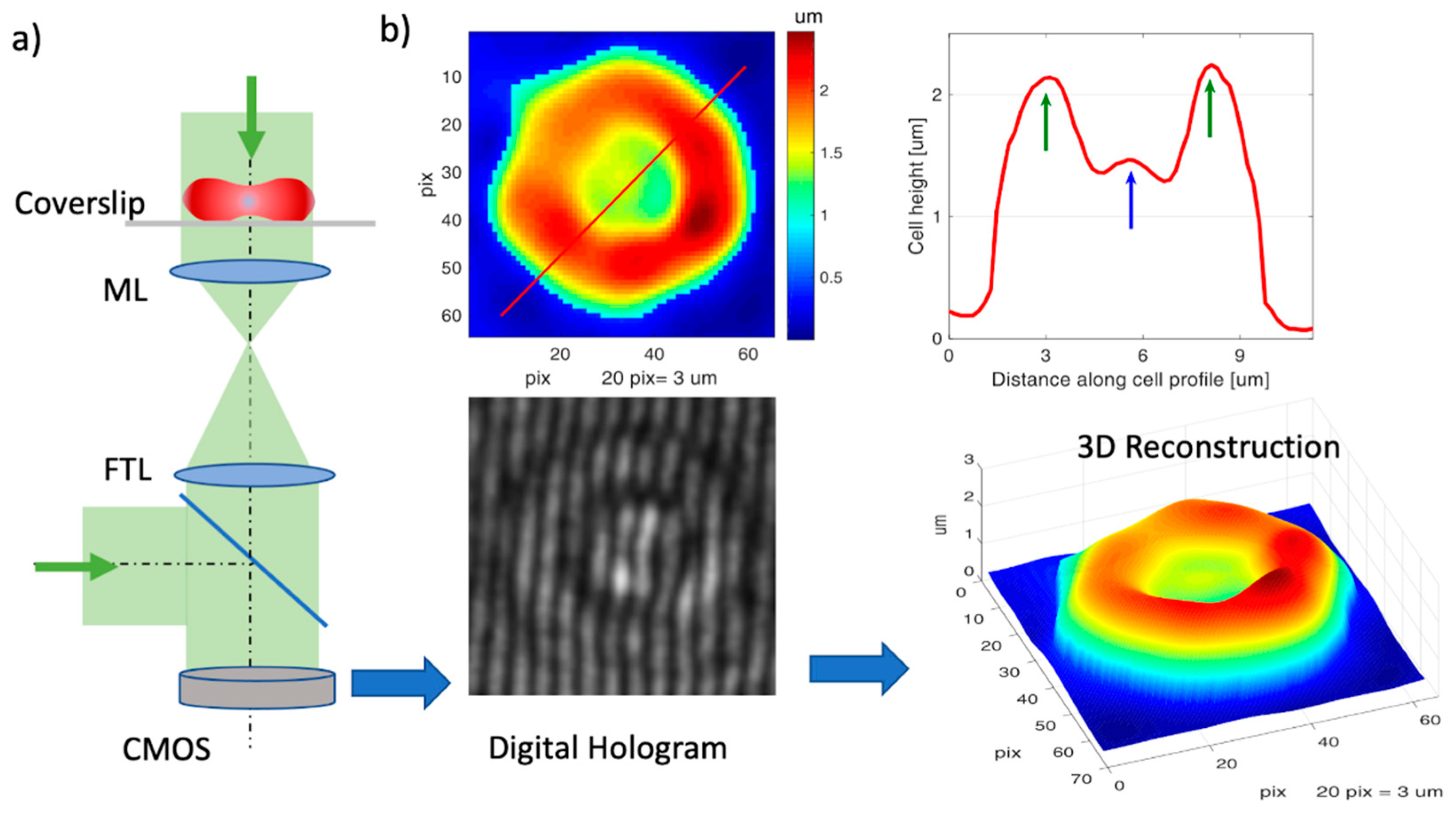
2.3. 3D Cell Imaging with Digital Holographic Microscopy
2.4. Calculation of the Cell Morphological Parameters and Cell Membrane Fluctuation
Cell Membrane Fluctuation (CMF)
3. Results
3.1. Deformation of Native vs. Ex Vivo-Generated RBC under Optical Forces
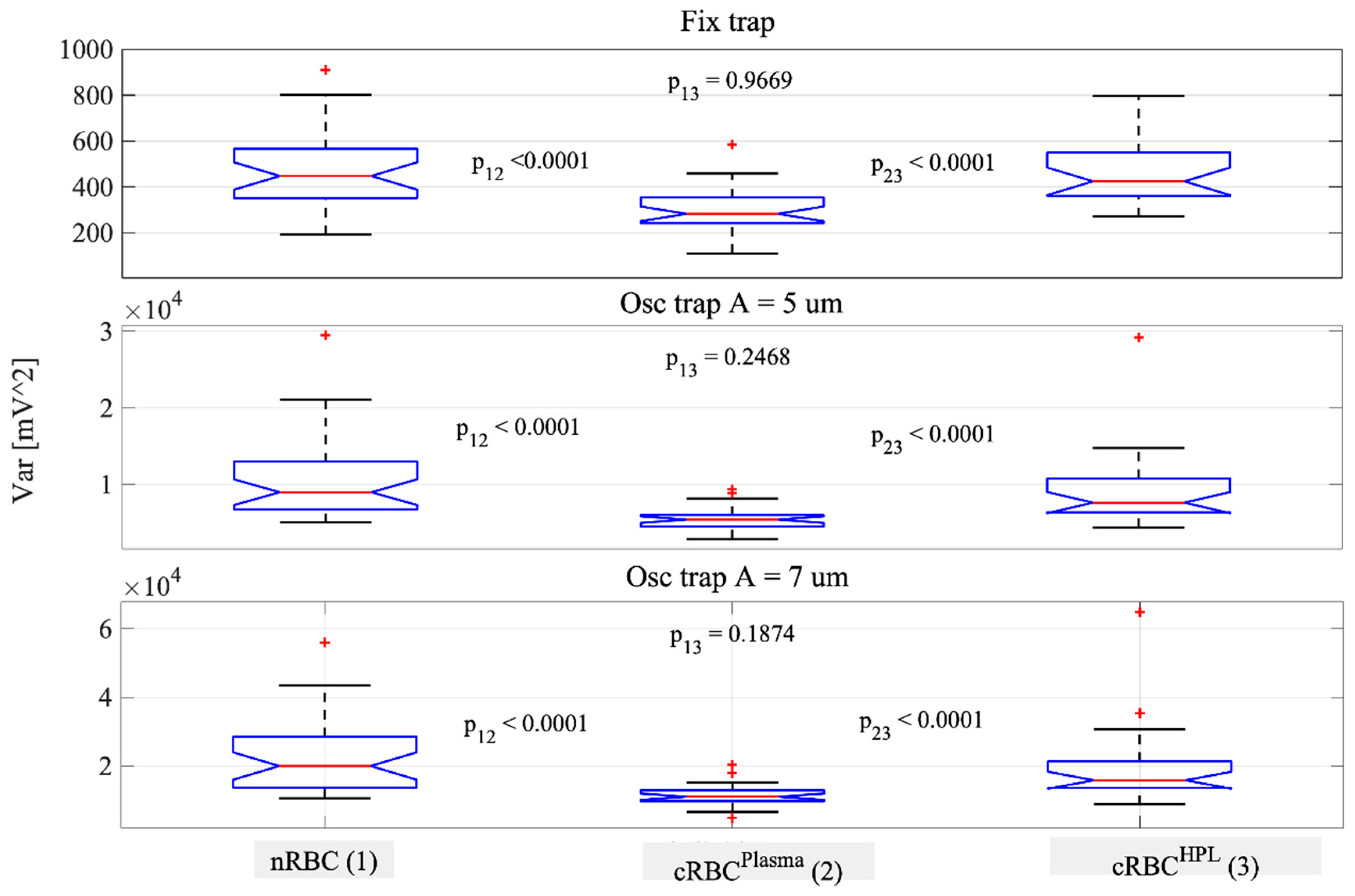
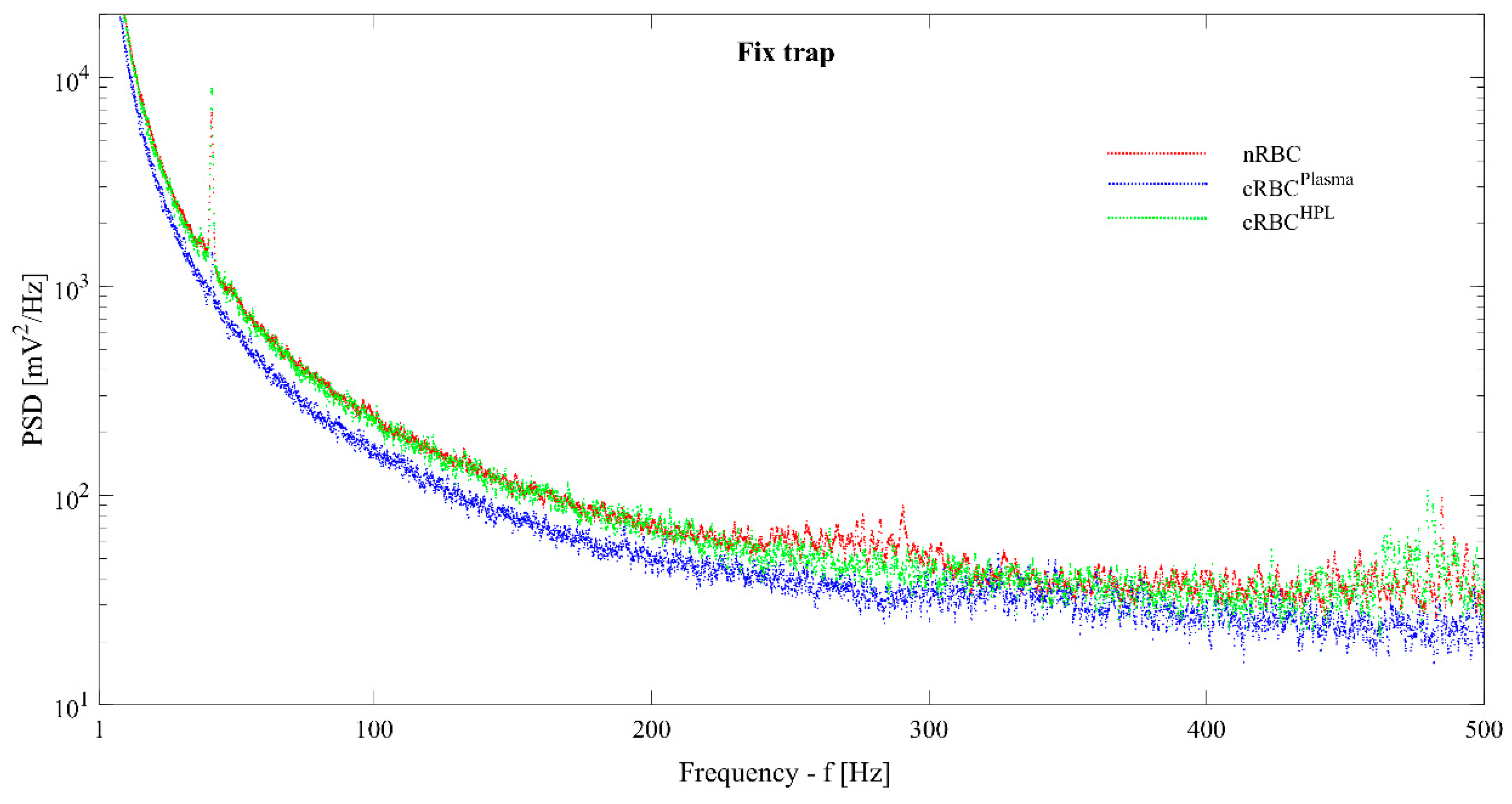
3.2. Cell Membrane Fluctuations of Native vs. Ex Vivo-Generated RBCs in OOTs
3.3. Morphological Parameters of Native vs. Ex Vivo-Generated RBCs Measured by DHM
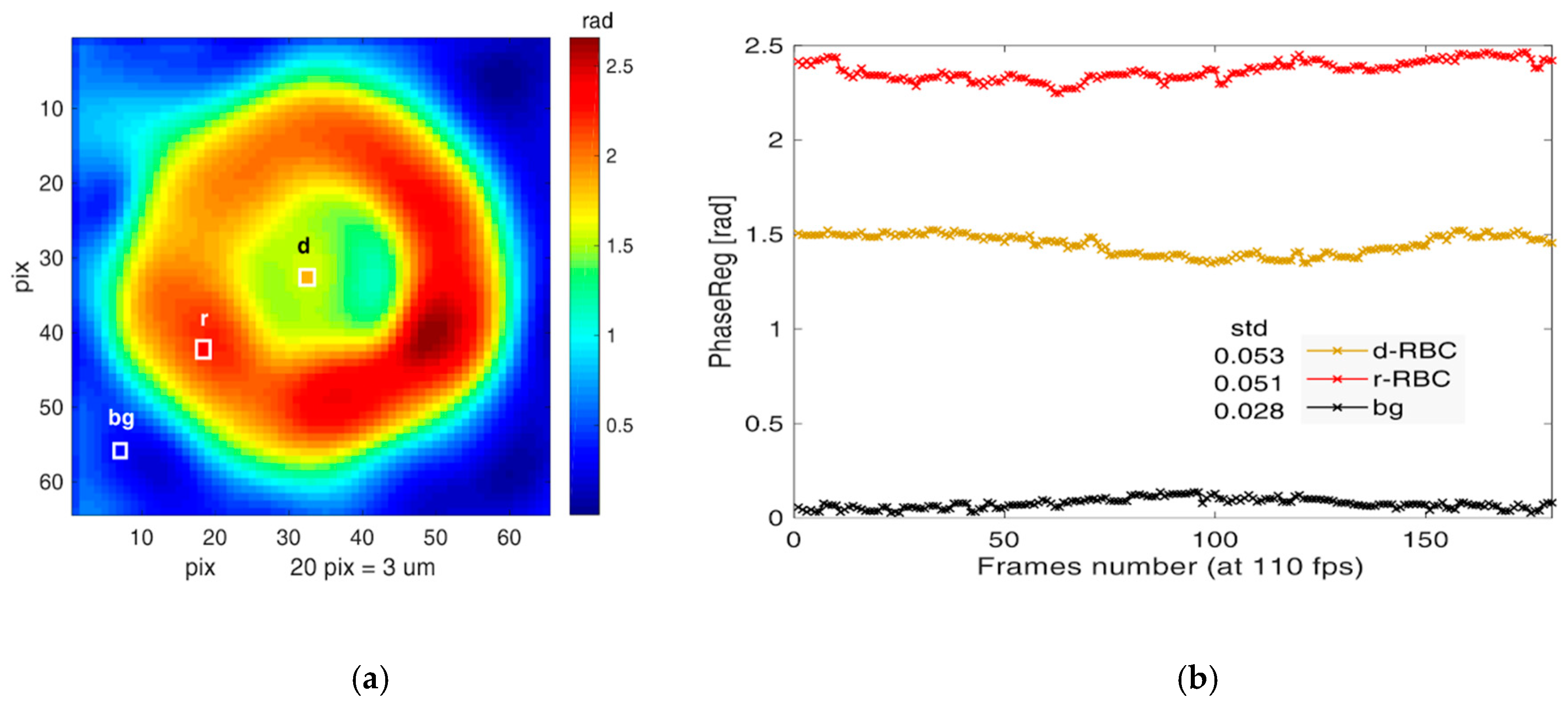
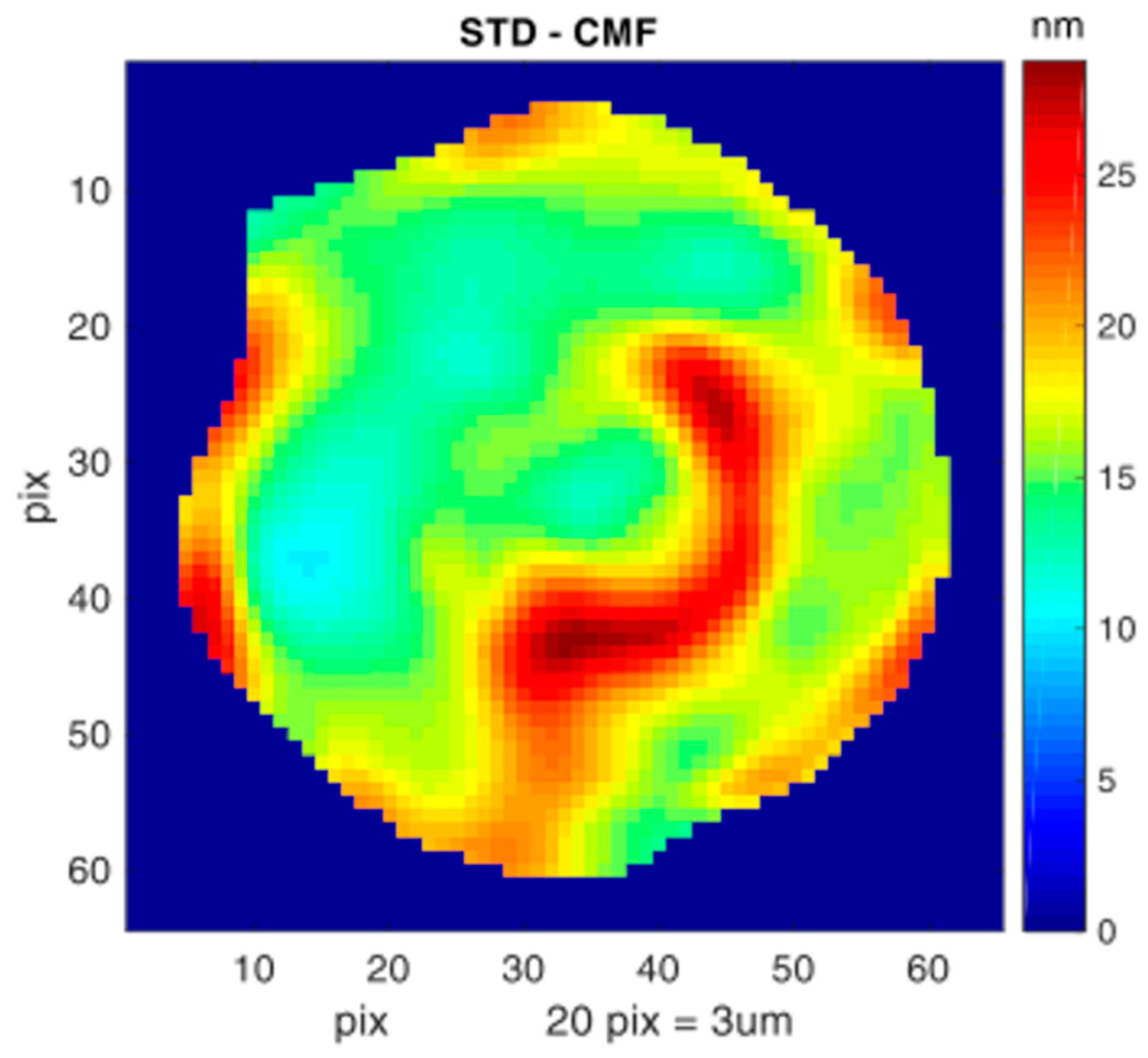
3.4. Cell Membrane Fluctuations of Native vs. Ex Vivo-Generated RBCs Measured by DHM
4. Discussion
4.1. OOTs Allow the Investigation of Cell Deformation under Forces of Similar Strength as Those on Cells in the Blood Stream
4.2. The Bending Modulus Can Be Estimated from the Cell Deformation and Membrane Fluctuations
4.3. DHM Provides Information Complementary to OOTs, Allowing a Faithful Identification of nRBCs vs. cRBCs
4.4. Conclusions about RBC Properties Based on DHM and OOT Data
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Minetti, G.; Achilli, C.; Perotti, C.; Ciana, A. Continuous Change in Membrane and Membrane-Skeleton Organization During Development From Proerythroblast to Senescent Red Blood Cell. Front. Physiol. 2018, 9, 286. [Google Scholar] [CrossRef] [Green Version]
- Bernecker, C.; Ackermann, M.; Lachmann, N.; Rohrhofer, L.; Zaehres, H.; Arauzo-Bravo, M.J.; van den Akker, E.; Schlenke, P.; Dorn, I. Enhanced Ex Vivo Generation of Erythroid Cells from Human Induced Pluripotent Stem Cells in a Simplified Cell Culture System with Low Cytokine Support. Stem Cells Dev. 2019, 28, 1540–1551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giarratana, M.C.; Kobari, L.; Lapillonne, H.; Chalmers, D.; Kiger, L.; Cynober, T.; Marden, M.C.; Wajcman, H.; Douay, L. Ex vivo generation of fully mature human red blood cells from hematopoietic stem cells. Nat. Biotechnol. 2005, 23, 69–74. [Google Scholar] [CrossRef]
- Trakarnsanga, K.; Griffiths, R.E.; Wilson, M.C.; Blair, A.; Satchwell, T.J.; Meinders, M.; Cogan, N.; Kupzig, S.; Kurita, R.; Nakamura, Y.; et al. An immortalized adult human erythroid line facilitates sustainable and scalable generation of functional red cells. Nat. Commun. 2017, 8, 14750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ovchynnikova, E.; Aglialoro, F.; von Lindern, M.; van den Akker, E. The Shape Shifting Story of Reticulocyte Maturation. Front. Physiol. 2018, 9, 829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pivkin, I.V.; Peng, Z.; Karniadakis, G.E.; Buffet, P.A.; Dao, M.; Suresh, S. Biomechanics of red blood cells in human spleen and consequences for physiology and disease. Proc. Natl. Acad. Sci. USA 2016, 113, 7804–7809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, X.; Lecomte, M.C.; Chasis, J.A.; Mohandas, N.; Gratzer, W. Shear-response of the spectrin dimer-tetramer equilibrium in the red blood cell membrane. J. Biol. Chem. 2002, 277, 31796–31800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huisjes, R.; Bogdanova, A.; van Solinge, W.W.; Schiffelers, R.M.; Kaestner, L.; van Wijk, R. Squeezing for Life—Properties of Red Blood Cell Deformability. Front. Physiol. 2018, 9, 656. [Google Scholar] [CrossRef] [PubMed]
- Lazari, D.; Freitas Leal, J.K.; Brock, R.; Bosman, G. The Relationship Between Aggregation and Deformability of Red Blood Cells in Health and Disease. Front. Physiol. 2020, 11, 288. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Lee, H.; Shin, S. Advances in the measurement of red blood cell deformability: A brief review. J. Cell. Biotechnol. 2015, 1, 63–79. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.; Kim, K.; Park, Y. Measurement Techniques for Red Blood Cell Deformability: Recent Advances. In Blood Cell—An Overview of Studies in Hematology; Moschandreou, T.E., Ed.; Intech Open: London, UK, 2012; pp. 167–194. [Google Scholar]
- Musielak, M. Red blood cell-deformability measurement: Review of techniques. Clin. Hemorheol. Microcirc. 2009, 42, 47–64. [Google Scholar] [CrossRef]
- Suresh, S. Mechanical response of human red blood cells in health and disease: Some structure-property-function relationships. J. Mater. Res. 2006, 21, 1871–1877. [Google Scholar] [CrossRef]
- Bronkhorst, P.J.; Streekstra, G.J.; Grimbergen, J.; Nijhof, E.J.; Sixma, J.J.; Brakenhoff, G.J. A new method to study shape recovery of red blood cells using multiple optical trapping. Biophys. J. 1995, 69, 1666–1673. [Google Scholar] [CrossRef] [Green Version]
- Guck, J.; Ananthakrishnan, R.; Mahmood, H.; Moon, T.J.; Cunningham, C.C.; Kas, J. The optical stretcher: A novel laser tool to micromanipulate cells. Biophys. J. 2001, 81, 767–784. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, A.; Sinha, S.; Dharmadhikari, J.A.; Roy, S.; Dharmadhikari, A.K.; Samuel, J.; Sharma, S.; Mathur, D. Euler buckling-induced folding and rotation of red blood cells in an optical trap. Phys. Biol. 2006, 3, 67–73. [Google Scholar] [CrossRef] [Green Version]
- Parthasarathi, P.; Nagesh, B.V.; Lakkegowda, Y.; Iyengar, S.S.; Ananthamurthy, S.; Bhattacharya, S. Orientational dynamics of human red blood cells in an optical trap. J. Biomed. Opt. 2013, 18, 25001. [Google Scholar] [CrossRef]
- Dao, M.; Lim, C.T.; Suresh, S. Mechanics of the human red blood cell deformed by optical tweezers. J. Mech. Phys. Solids 2005, 53, 2259–2280. [Google Scholar] [CrossRef]
- Henon, S.; Lenormand, G.; Richert, A.; Gallet, F. A new determination of the shear modulus of the human erythrocyte membrane using optical tweezers. Biophys. J. 1999, 76, 1145–1151. [Google Scholar] [CrossRef] [Green Version]
- Yoon, Y.Z.; Kotar, J.; Brown, A.T.; Cicuta, P. Red blood cell dynamics: From spontaneous fluctuations to non-linear response. Soft Matter 2011, 7, 2042–2051. [Google Scholar] [CrossRef]
- Turlier, H.; Fedosov, D.A.; Audoly, B.; Auth, T.; Gov, N.S.; Sykes, C.; Joanny, J.F.; Gompper, G.; Betz, T. Equilibrium physics breakdown reveals the active nature of red blood cell flickering. Nat. Phys. 2016, 12, 513–519. [Google Scholar] [CrossRef] [Green Version]
- Avsievich, T.; Zhu, R.; Popov, A.; Bykov, A.; Meglinski, I. The advancement of blood cell research by optical tweezers. Rev. Phys. 2020, 5, 100043. [Google Scholar] [CrossRef]
- Zhu, R.; Avsievich, T.; Popov, A.; Meglinski, I. Optical Tweezers in Studies of Red Blood Cells. Cells 2020, 9, 545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Betz, T.; Lenz, M.; Joanny, J.F.; Sykes, C. ATP-dependent mechanics of red blood cells. Proc. Natl. Acad. Sci. USA 2009, 106, 15320–15325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brochard, F.; Lennon, J.F. Frequency spectrum of the flicker phenomenon in erythrocytes. J. Phys. 1975, 36, 1035–1047. [Google Scholar] [CrossRef]
- Evans, J.; Gratzer, W.; Mohandas, N.; Parker, K.; Sleep, J. Fluctuations of the red blood cell membrane: Relation to mechanical properties and lack of ATP dependence. Biophys. J. 2008, 94, 4134–4144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, Y.Z.; Hong, H.; Brown, A.; Kim, D.C.; Kang, D.J.; Lew, V.L.; Cicuta, P. Flickering analysis of erythrocyte mechanical properties: Dependence on oxygenation level, cell shape, and hydration level. Biophys. J. 2009, 97, 1606–1615. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.; Depeursinge, C.; Popescu, G. Quantitative phase imaging in biomedicine. Nat. Photonics 2018, 12, 578–589. [Google Scholar] [CrossRef]
- Park, Y.; Best, C.A.; Auth, T.; Gov, N.S.; Safran, S.A.; Popescu, G.; Suresh, S.; Feld, M.S. Metabolic remodeling of the human red blood cell membrane. Proc. Natl. Acad. Sci. USA 2010, 107, 1289–1294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaferzadeh, K.; Moon, I.; Bardyn, M.; Prudent, M.; Tissot, J.D.; Rappaz, B.; Javidi, B.; Turcatti, G.; Marquet, P. Quantification of stored red blood cell fluctuations by time-lapse holographic cell imaging. Biomed. Opt. Express 2018, 9, 4714–4729. [Google Scholar] [CrossRef] [PubMed]
- Jaferzadeh, K.; Moon, I. Quantitative investigation of red blood cell three-dimensional geometric and chemical changes in the storage lesion using digital holographic microscopy. J. Biomed. Opt. 2015, 20, 111218. [Google Scholar] [CrossRef] [PubMed]
- Bernecker, C.; Kofeler, H.; Pabst, G.; Trotzmuller, M.; Kolb, D.; Strohmayer, K.; Trajanoski, S.; Holzapfel, G.A.; Schlenke, P.; Dorn, I. Cholesterol Deficiency Causes Impaired Osmotic Stability of Cultured Red Blood Cells. Front. Physiol. 2019, 10, 1529. [Google Scholar] [CrossRef]
- Falleroni, F.; Torre, V.; Cojoc, D. Cell Mechanotransduction With Piconewton Forces Applied by Optical Tweezers. Front. Cell Neurosci. 2018, 12, 130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yousafzai, M.S.; Ndoye, F.; Coceano, G.; Niemela, J.; Bonin, S.; Scoles, G.; Cojoc, D. Substrate-dependent cell elasticity measured by optical tweezers indentation. Opt. Lasers Eng. 2016, 76, 27–33. [Google Scholar] [CrossRef]
- Picazo-Bueno, J.A.; Cojoc, D.; Iseppon, F.; Torre, V.; Mico, V. Single-shot, dual-mode, water-immersion microscopy platform for biological applications. Appl. Opt. 2018, 57, A242–A249. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Liu, J.; Xue, F.; Halverson, G.; Reid, M.; Guo, A.; Chen, L.; Raza, A.; Galili, N.; Jaffray, J.; et al. Isolation and functional characterization of human erythroblasts at distinct stages: Implications for understanding of normal and disordered erythropoiesis in vivo. Blood 2013, 121, 3246–3253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashkin, A. Forces of a single-beam gradient laser trap on a dielectric sphere in the ray optics regime. Biophys. J. 1992, 61, 569–582. [Google Scholar] [CrossRef] [Green Version]
- Nebe, T.; Bentzien, F.; Bruegel, M.; Fiedler, G.M.; Gutensohn, K.; Heimpel, H.; Krebs, N.; Ossendorf, M.; Schuff-Werner, P.; Stamminger, G.; et al. Multizentrische Ermittlung von Referenzbereichen für Parameter des maschinellen Blutbildes/Multicentric Determination of Reference Ranges for Automated Blood Counts. J. Lab. Med. 2011, 35, 3–28. [Google Scholar] [CrossRef]
- Evans, E.A. Structure and deformation properties of red blood cells: Concepts and quantitative methods. Methods Enzymol. 1989, 173, 3–35. [Google Scholar] [CrossRef] [PubMed]
- Savin, T.; Bandi, M.M.; Mahadevan, L. Pressure-driven occlusive flow of a confined red blood cell. Soft Matter 2016, 12, 562–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Secomb, T.W.; Hsu, R.; Pries, A.R. Motion of red blood cells in a capillary with an endothelial surface layer: Effect of flow velocity. Am. J. Physiol. Heart Circ. Physiol. 2001, 281, H629–H636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGloin, D.; Garces-Chavez, V.; Dholakia, K. Interfering Bessel beams for optical micromanipulation. Opt. Lett. 2003, 28, 657–659. [Google Scholar] [CrossRef] [PubMed]
- Guzniczak, E.; Mohammad Zadeh, M.; Dempsey, F.; Jimenez, M.; Bock, H.; Whyte, G.; Willoughby, N.; Bridle, H. High-throughput assessment of mechanical properties of stem cell derived red blood cells, toward cellular downstream processing. Sci. Rep. 2017, 7, 14457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hategan, A.; Law, R.; Kahn, S.; Discher, D.E. Adhesively-tensed cell membranes: Lysis kinetics and atomic force microscopy probing. Biophys. J. 2003, 85, 2746–2759. [Google Scholar] [CrossRef] [Green Version]
- Mohandas, N.; Chasis, J.A. Red blood cell deformability, membrane material properties and shape: Regulation by transmembrane, skeletal and cytosolic proteins and lipids. Semin. Hematol. 1993, 30, 171–192. [Google Scholar]
- Peng, Z.; Li, X.; Pivkin, I.V.; Dao, M.; Karniadakis, G.E.; Suresh, S. Lipid bilayer and cytoskeletal interactions in a red blood cell. Proc. Natl. Acad. Sci. USA 2013, 110, 13356–13361. [Google Scholar] [CrossRef] [Green Version]
- Discher, D.E. New insights into erythrocyte membrane organization and microelasticity. Curr. Opin. Hematol. 2000, 7, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Eber, S.; Lux, S.E. Hereditary spherocytosis—Defects in proteins that connect the membrane skeleton to the lipid bilayer. Semin. Hematol. 2004, 41, 118–141. [Google Scholar] [CrossRef] [PubMed]
- Gaetani, M.; Mootien, S.; Harper, S.; Gallagher, P.G.; Speicher, D.W. Structural and functional effects of hereditary hemolytic anemia-associated point mutations in the alpha spectrin tetramer site. Blood 2008, 111, 5712–5720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iolascon, A.; Perrotta, S.; Stewart, G.W. Red blood cell membrane defects. Rev. Clin. Exp. Hematol. 2003, 7, 22–56. [Google Scholar]
- Mohandas, N.; Gallagher, P.G. Red cell membrane: Past, present, and future. Blood 2008, 112, 3939–3948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salomao, M.; Zhang, X.; Yang, Y.; Lee, S.; Hartwig, J.H.; Chasis, J.A.; Mohandas, N.; An, X. Protein 4.1R-dependent multiprotein complex: New insights into the structural organization of the red blood cell membrane. Proc. Natl. Acad. Sci. USA 2008, 105, 8026–8031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Betz, J.; Dorn, I.; Kouzel, I.U.; Bauwens, A.; Meisen, I.; Kemper, B.; Bielaszewska, M.; Mormann, M.; Weymann, L.; Sibrowski, W.; et al. Shiga toxin of enterohaemorrhagic Escherichia coli directly injures developing human erythrocytes. Cell Microbiol. 2016, 18, 1339–1348. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variance Mean/Std (mV2) | nRBC (n = 32) | cRBCPlasma (n = 24) | cRBCHPL (n = 30) |
| Fixed trap | 460/160 | 300/100 | 470/140 |
| Osc. A = 5 µm | 10,800/5500 | 5600/1500 | 9200/5100 |
| Osc. A = 7 µm | 22,600/10,800 | 11,300/3100 | 19,900/11,600 |
| Morphology | CA | CV | CS | MCH | hm | |
|---|---|---|---|---|---|---|
| mean ± std | mean ± std | mean ± std | mean ± std | mean | ||
| Cells | n | µm2 | µm3 (or fL) | - | Picogram (pg) | µm |
| nRBC | 25 | 55.42 ± 9.2 | 95.2 ± 16.6 | 0.57 ± 0.1 | 25.24 ± 5 | 1.72 ± 0.4 |
| cRBCPlasma | 24 | 41.05 ± 14.4 | 125.5 ± 43.3 | 1.04 ± 0.1 | 31.17 ± 11,7 | 3.06 ± 0.6 |
| cRBCHPL | 29 | 70 ± 21.7 | 107.1 ± 37.8 | 0.671 ± 0.4 | 28.1 ± 10.9 | 1.53 ± 0.3 |
| CMF | Cell Measured | Background | Height Normalization | Cell Corrected | |||
|---|---|---|---|---|---|---|---|
| mean | std | mean | std | c | mean | ||
| cells | n | nm | nm | nm | nm | - | nm |
| nRBC | 25 | 54.4 | 10.8 | 24.7 | 6.2 | 1.00 | 54.47 |
| cRBCPlasma | 24 | 53.8 | 21.4 | 25.3 | 9.1 | 1.78 | 30.27 |
| cRBCHPL | 29 | 37.9 | 14.3 | 19.4 | 7.9 | 0.89 | 42.61 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bernecker, C.; Lima, M.A.R.B.F.; Ciubotaru, C.D.; Schlenke, P.; Dorn, I.; Cojoc, D. Biomechanics of Ex Vivo-Generated Red Blood Cells Investigated by Optical Tweezers and Digital Holographic Microscopy. Cells 2021, 10, 552. https://doi.org/10.3390/cells10030552
Bernecker C, Lima MARBF, Ciubotaru CD, Schlenke P, Dorn I, Cojoc D. Biomechanics of Ex Vivo-Generated Red Blood Cells Investigated by Optical Tweezers and Digital Holographic Microscopy. Cells. 2021; 10(3):552. https://doi.org/10.3390/cells10030552
Chicago/Turabian StyleBernecker, Claudia, Maria Augusta R. B. F. Lima, Catalin D. Ciubotaru, Peter Schlenke, Isabel Dorn, and Dan Cojoc. 2021. "Biomechanics of Ex Vivo-Generated Red Blood Cells Investigated by Optical Tweezers and Digital Holographic Microscopy" Cells 10, no. 3: 552. https://doi.org/10.3390/cells10030552