Flux Enforcement for Fermentative Production of 5-Aminovalerate and Glutarate by Corynebacterium glutamicum



1
Genetics of Prokaryotes, Faculty of Biology and CeBiTec, Bielefeld University, Universitätsstr. 25, 33615 Bielefeld, Germany
2
TBI, Université de Toulouse, CNRS, INRAE, INSA, 31077 Toulouse, France
3
Department of Biotechnology and Food Science, Norwegian University of Science and Technology (NTNU), 7034 Trondheim, Norway
*
Author to whom correspondence should be addressed.
Catalysts 2020, 10(9), 1065; https://doi.org/10.3390/catal10091065
Submission received: 28 August 2020
/
Revised: 8 September 2020
/
Accepted: 14 September 2020
/
Published: 16 September 2020
(This article belongs to the Special Issue Multi-Step Syntheses in Biology & Chemistry)
Abstract
:Bio-based plastics represent an increasing percentage of the plastics economy. The fermentative production of bioplastic monomer 5-aminovalerate (5AVA), which can be converted to polyamide 5 (PA 5), has been established in Corynebacterium glutamicum via two metabolic pathways. l-lysine can be converted to 5AVA by either oxidative decarboxylation and subsequent oxidative deamination or by decarboxylation to cadaverine followed by transamination and oxidation. Here, a new three-step pathway was established by using the monooxygenase putrescine oxidase (Puo), which catalyzes the oxidative deamination of cadaverine, instead of cadaverine transaminase. When the conversion of 5AVA to glutarate was eliminated and oxygen supply improved, a 5AVA titer of 3.7 ± 0.4 g/L was reached in microcultivation that was lower than when cadaverine transaminase was used. The elongation of the new pathway by 5AVA transamination by GABA/5AVA aminotransferase (GabT) and oxidation by succinate/glutarate semialdehyde dehydrogenase (GabD) allowed for glutarate production. Flux enforcement by the disruption of the l-glutamic acid dehydrogenase-encoding gene gdh rendered a single transaminase (GabT) in glutarate production via the new pathway responsible for nitrogen assimilation, which increased the glutarate titer to 7.7 ± 0.7 g/L, i.e., 40% higher than with two transaminases operating in glutarate biosynthesis. Flux enforcement was more effective with one coupling site, thus highlighting requirements regarding the modularity and stoichiometry of pathway-specific flux enforcement for microbial production.
1. Introduction
Plastics are primarily synthesized chemically from petroleum and natural gas. However, the annual market volume of bioplastics is predicted to increase from 2.11 million tons in 2019 to 2.43 million tons in 2024 [1]. Among bioplastics, the biopolyamides find many applications ranging from the production of parachutes during World War II to filaments for 3D-printing at present [2,3]. Polyamides can be produced via two routes: either by the condensation of dicarboxylic acids with diamines or by the anionic ring-opening polymerization of lactams, which can be formed from ω-amino acids via cyclization [4]. Examples of these main building blocks range from succinate to sebacate for dicarboxylic acids, from putrescine to hexamethylenediamine for diamines, and from γ-aminobutyrate (GABA) to ε-aminocaproate (6ACA) for ω-amino acids.
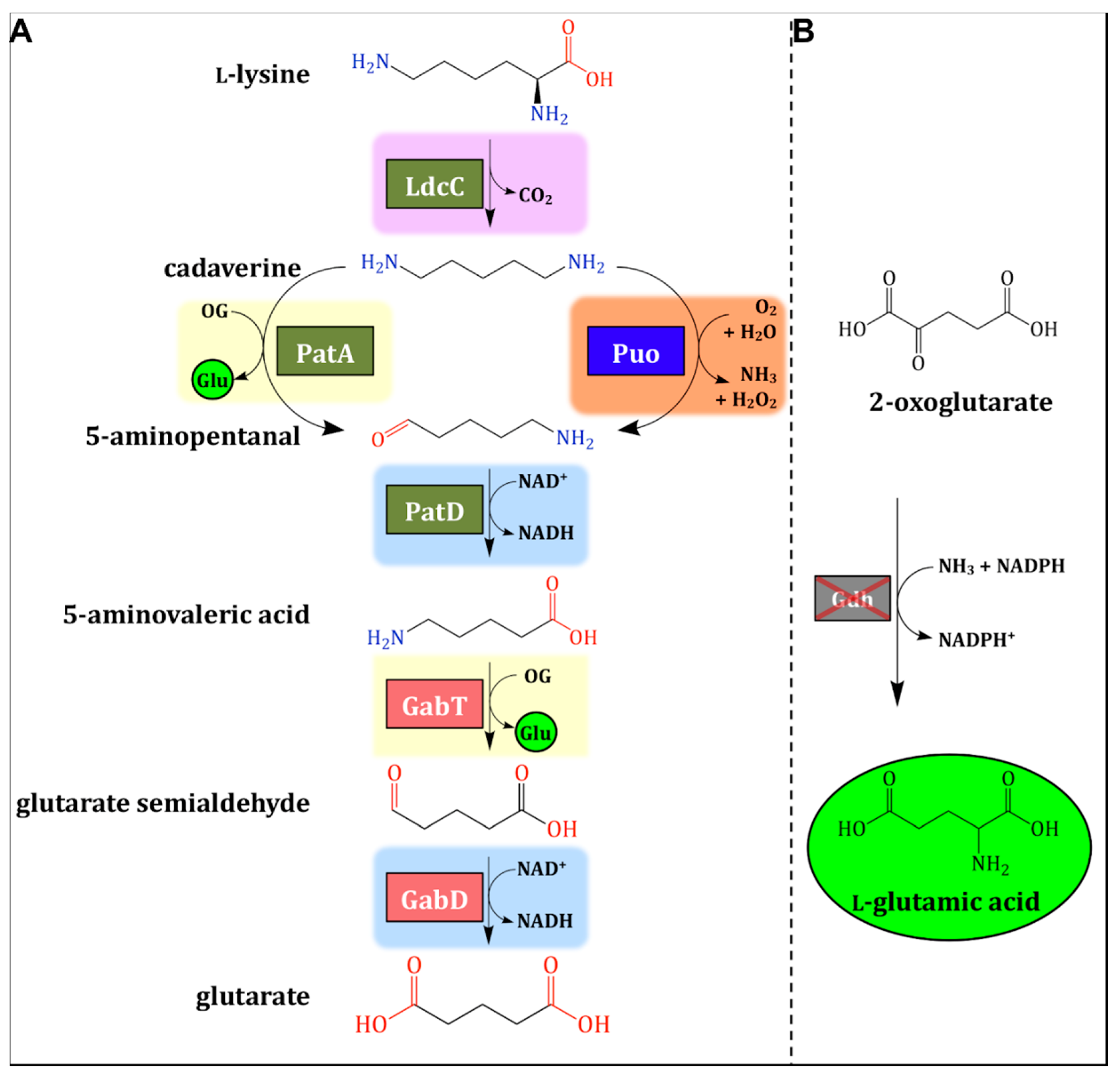
Monomeric polyamide precursors with a carbon chain length of 5 (C5) comprise the ω-amino acid 5-aminovalerate (5AVA) for ring-opening polymerization to polyamide 5 (PA 5), the diamine cadaverine, and the dicarboxylic acid glutarate for the co-polycondensation to PA 5.5 [5]. All three precursors can be synthesized from the amino acid l-lysine by a series of oxidation, decarboxylation, and deamination reactions; thus, fermentative routes for their production have been established. Corynebacterium glutamicum is well-known for the industrial production of l-lysine by fermentation, a process operated at about 2.6 million tons in 2018 [6]. l-lysine-overproducing C. glutamicum strains have been converted to cadaverine producers through the expression of a heterologous l-lysine decarboxylase gene (Figure 1A) [7], and high titers, yields, and productivities have been reached, e.g., using lignocellulose derived wastes [8,9].
For the production of 5AVA from l-lysine, two metabolic pathways have been developed. A three-step route to 5AVA that does not require molecular oxygen (Figure 1A) was established in our group, and it is compatible with aerobic fermentation that is typically limited by low oxygen transfer rates. This three-step LdcC–PatA–PatD route involves l-lysine decarboxylase, putrescine transaminase, and γ-aminobutyraldehyde dehydrogenase (encoded by the genes ldcC, patA, and patD, respectively) [10]. In shake flasks, 5AVA product titers of about 5 g/L, a yield of 0.13 g/g, and a volumetric productivity of 0.12 g/L/h were reached [10]. Independently, a two-step route based on davAB-encoded δ-aminovaleramidase and l-lysine monooxygenase from Pseudomonas putida has been used in Escherichia coli and C. glutamicum with product titers up to about 33 g/L [11,12,13]. The first reaction in the DavB–DavA route requires the presence of molecular oxygen for the oxidative decarboxylation prior to hydrolyzation from l-lysine via 5-aminovaleramide to 5AVA.
Glutarate can be derived from 5AVA by transamination and oxidation (Figure 1A). C. glutamicum possesses the gabTD operon encoding enzymes responsible for the catabolism and import of GABA, an ω-amino acid with a shorter chain length than 5AVA (C4 instead of C5). The gabTD-encoded GABA/5AVA aminotransferase, and succinate/glutarate semialdehyde dehydrogenase accept both C4 and C5 substrates [14]. These genes can be deleted to increase the production of 5AVA by preventing its degradation [10] or overexpressed to improve the conversion of 5AVA to glutarate [15]. Glutarate production via the five-step LdcC–PatA–PatD–GabT–GabD route (Figure 1A) was found to enable the fermentative production of glutarate up to 25 g/L [15]. Glutarate production has also been realized by employing the DavB–DavA–GabT–GabD route, and the produced glutarate was used with diaminohexane to prepare PA 6,5 [12,16].
The metabolic engineering strategy of flux enforcement couples a biosynthetic production pathway to a pathway required for growth [6]. The coupling of the production of 4-hydroxy-l-isoleucine via 2-oxoglutarate-dependent hydroxylase to growth by the deletion of the 2-oxoglutarate dehydrogenase subunit gene sucA was first described for E. coli [17]. In this application and in 4-hydroxy-l-proline production [18], the production pathways become part of an artificial TCA cycle. Flux enforcement by replacing a reaction of the TCA cycle was also used to improve l-lysine production by a succinyl-CoA synthetase-negative (ΔsucCD), l-lysine-producing C glutamicum strain where the succinylase branch of l-lysine production metabolically complemented the disrupted TCA cycle [19].
Flux enforcement has also been applied to improve glutarate production by C. glutamicum [15]. The transamination reactions catalyzed by cadaverine/putrescine transaminase PatA and GABA/5AVA amino transferase GabT during glutarate production were coupled to growth in the absence of the major ammonium assimilating enzyme l-glutamic acid dehydrogenase Gdh [15] (Figure 1B). The resulting strain reached glutarate titers of about 5 g/L in shake flasks and about 25 g/L during fed-batch fermentation [15]. Here, the monooxyogenase enzyme putrescine oxidase encoded by puo from Rhodococcus qingshengii or Paenarthrobacter aurescens was tested for the conversion of cadaverine to 5-aminopentanal in pathways for the production of 5AVA and glutarate (Figure 1A). We analyzed whether flux enforcement by coupling gdh deletion to a single transaminase activity using this novel pathway was superior to its coupling to the two transaminases PatA and GabT for glutarate production.
2. Results
2.1. Design of the Study: Comparing Flux Enforcement with Either a Single or Two Coupling Sites
The five-step LdcC–PatA–PatD–GabT–GabD route from l-lysine to glutarate conserves the energy available to the cell in two ways. First, the oxidoreductases PatD and GabD yield the reduced redox cofactor NADH. Second, during the conversion of cadaverine to 5-aminopentanal and 5AVA to glutarate semialdehyde, the transaminases PatA and GabT simultaneously yield l-glutamic acid from the TCA cycle intermediate 2-oxoglutarate. The operation of the reactions of PatA and/or GabT in the glutarate production pathway obviates the need to synthesize l-glutamic acid from 2-oxoglutarate and ammonium by NADPH-dependent l-glutamic acid dehydrogenase, which is encoded by gdh. Indeed, flux enforcement by the deletion of gdh was found to improve glutarate production via the five-step LdcC–PatA–PatD–GabT–GabD route [15] because the deletion of gdh was compensated for by the activities of the two transaminases PatA and GabT. This prompted us to analyze whether flux enforcement by gdh deletion coupled to a single transaminase reaction in the glutarate pathway was superior to coupling to the two transaminases PatA and GabT.
The fourth reaction in the five-step reaction sequence (transamination by GabT) was kept in order to maintain the “metabolic pull” of flux enforcement by gdh deletion, while the second reaction (transamination by PatA) was chosen to be replaced by an oxidative deamination reaction. Unlike the 2-oxoglutarate-dependent transamination catalyzed by PatA, oxidative deamination by an oxidase does not yield l-glutamic acid; thus, it cannot bypass NADPH-dependent l-glutamic acid dehydrogenase. A search for oxidases that accept substrates similar to cadaverine was conducted. Indeed, putrescine oxidase (EC 1.4.3.10) encoded by puo from Rhodococcus erythropolis accepts cadaverine as a substrate with a catalytic efficiency (kcat/Km) of 220 s−1 mM−1 [20]. Another candidate from Paenarthrobacter aurescens also showed enzyme activity towards cadaverine, albeit with magnitudes lower kcat/Km (4.78 s−1 mM−1) [21]. Both enzymes contain non-covalently bound FAD. The putrescine oxidase gene puoRq from Rhodococcus qingshengii was used due to the 98% identity similarity of its amino acid sequence to the characterized putrescine oxidase proteins from R. erythropolis [20] and the fact that genomic DNA from R. qingshengii was available from the strain collection Deutsche Sammlung von Mikroorganismen und Zellkulturen (DSMZ). The putrescine oxidases from P. aurescens (puoPa) and R. qingshengii (puoRq)were tested in the second position of the five-step pathway for conversion of l-lysine to glutarate in order to compare the new routes (LdcC–PuoRq–PatD–GabT–GabD and LdcC–PuoPa–PatD–GabT–GabD) to the reference route of LdcC–PatA–PatD–GabT–GabD.
2.2. Proof of Principle: Putrescine Oxidases Oxidatively Deaminate Cadaverine in C. glutamicum
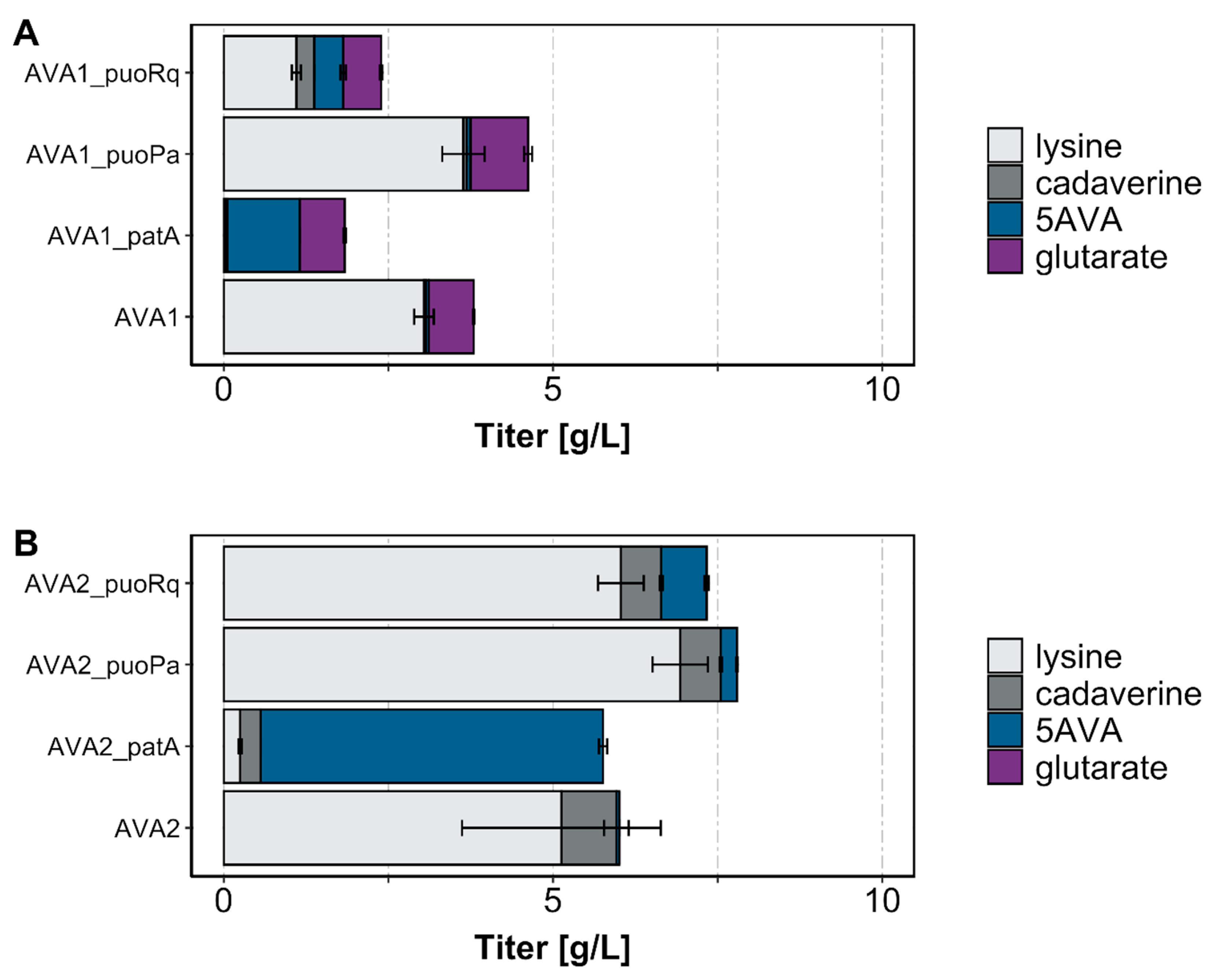
To test whether the putrescine oxidases from P. aurescens and R. qingshengii catalyzed the oxidative deamination of cadaverine to 5AVA in C. glutamicum, 5AVA production via the three-step pathway was analyzed after replacing transamination by PatA with a putrescine oxidase. The putrescine oxidase genes from P. aurescens and R. qingshengii, respectively, were heterologously expressed in the l-lysine-producing C. glutamicum strain GSLA2 to generate the LdcC–PuoRq–PatD and LdcC–PuoPa–PatD variants of the reference three-step LdcC–PatA–PatD route. GSLA2 is based on GRLys1, which was obtained by genome reduction and rational metabolic engineering of l-lysine production [22,23,24], but it also lacks the genes sugR, ldhA, snaA and cgmA to improve glucose consumption and to avoid N-acetylation and the export of cadaverine [8,23,25,26,27]. The AVA1_patA reference strain produced 1.1 ± 0.0 g/L 5AVA in 3 mL microtiter plates (Figure 2A). Notably, the strains with the new LdcC–PuoRq–PatD and LdcC–PuoPa–PatD routes produced 0.4 ± 0.0 g/L 5AVA (AVA1_puoRq) and 0.1 ± 0.0 g/L 5AVA (AVA1_puoPa), respectively. Thus, a proof of concept for the fermentative production of 5AVA with putrescine oxidases was achieved.
The spectrum of by-products formed indicated the incomplete conversion of l-lysine to 5AVA since 1.1 ± 0.1 and 3.6 ± 0.3 g/L of l-lysine and 0.3 ± 0.0 and 0.1 ± 0.0 g/L of cadaverine accumulated in the cultivations with AVA1_puoRq and AVA1_puoPa, respectively (Figure 2A). This was not observed with the AVA1_patA reference strain. All strains produced glutarate (Figure 2A), which may be synthesized from 5AVA by endogenous, chromosomally encoded enzymes, namely GabT and GabD.
2.3. Deletion of Genes for Conversion of 5AVA to Glutarate and Change of the Microcultivation System Improved 5AVA Production
The accumulation of glutarate by the strains with the different three-step routes (LdcC–PatA–PatD, LdcC–PuoRq–PatD and LdcC–PuoPa–PatD) (Figure 2A) prompted us to delete the GABA utilization operon. The deletion of the gabTDP operon has previously been shown to abrogate glutarate formation when using the LdcC–PatA–PatD reference route [10]. The assumption was that this would abolish glutarate formation as a by-product of the new AVA2_puoRq and AVA2_puoPa strains. It was, in fact, shown to be correct because glutarate was not detected after 48 h of the 3 mL microtiter plate cultivation of AVA2_puoRq, AVA2_puoPa, and the reference strain AVA2_patA (Figure 2B). Compared to the strains possessing gabTDP operon (Figure 2A), the production of 5AVA increased two-to-five fold to final titers of 0.7 ± 0.0 g/L for AVA2_puoRq, 0.2 ± 0.0 g/L for AVA2_puoPa, and 5.2 ± 0.1 g/L for the reference strain AVA2_patA (Figure 2B). While the accumulation of cadaverine was comparable for the three strains, the C. glutamicum strains AVA2_puoRq and AVA2_puoPa accumulated l-lysine to high concentrations (6.9 ± 0.4 and 6.0 ± 0.3 g/L, respectively; Figure 2B). This indicated the incomplete conversion of l-lysine to 5AVA by the AVA2_puoRq and AVA2_puoPa strains.
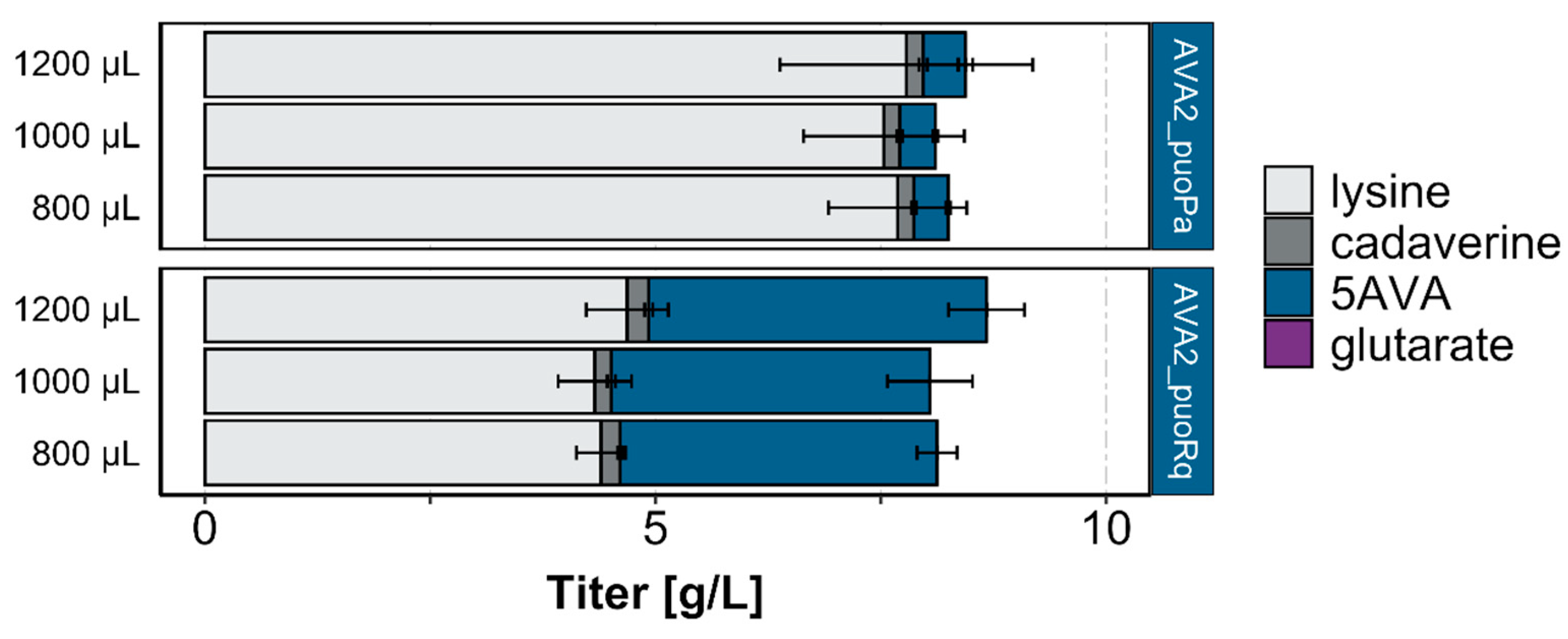
Since the putrescine oxidase reaction requires molecular oxygen, we analyzed 5AVA production in another microcultivation system while allowing for a higher oxygen transfer rate than the current system (i.e., a Duetz system with a cultivation volume of 3 mL at 220 rpm). We used a BioLector with different filling volumes (800, 1000, and 1200 μL) at 1300 rpm, which are known to provide superior oxygen transfer rates [28,29]. Cultivation in the BioLector system lead to increased production of 5AVA by AVA2_puoRq about five-fold (3.5 ± 0.2, 3.5 ± 0.5, and 3.7 ± 0.4 g/L with filling volumes of 800, 1000, and 1200 μL) in comparison to the Duetz system. 5AVA production by AVA2_puoPa was increased by about two-fold (0.4 ± 0.0, 0.4 ± 0.0, and 0.5 ± 0.1 g/L with filling volumes of 800, 1000, and 1200 μL, respectively), as can be seen by comparing Figure 3 with Figure 2B. Notably, significantly less l-lysine was accumulated by the strain overexpressing puoRq than the strain overexpressing puoPa (4.3–4.7 g/L compared to 6.9 ± 0.4 g/L, respectively), as can be seen in Figure 3.
Taken together, cultivation with high oxygen transfer rates in the BioLector system improved 5AVA production by the new three step route and allowed us to identify the better performance of the LdcC–PuoRq–PatD variant than of the LdcC–PuoPa–PatD variant; however, the LdcC–PuoRq–PatD variant was found to be inferior to the LdcC–PatA–PatD route in regards to 5AVA production. Nevertheless, we continued to use the LdcC–PuoRq–PatD variant with the putrescine oxidase from R. qingshengii to test its performance for glutarate production and flux enforcement.
2.4. Flux Enforcement by Deletion of the l-Glutamic Acid Dehydrogenase Gene Improved Glutarate Production
Production can be coupled to growth requirements by flux enforcement. The flux enforcement of glutarate production has been achieved by the deletion of the l-glutamic acid dehydrogenase gene in C. glutamicum [15]. l-glutamic acid dehydrogenase is the major reactant that is necessary for the assimilation of ammonium into l-glutamic acid in C. glutamicum [30]. 2-Oxoglutarate-dependent transaminase reactions in synthetic cascades can substitute for l-glutamic acid dehydrogenase if they operate in the direction of l-glutamic acid formation. In the five-step pathway from l-lysine to glutarate with LdcC, PatA, PatD, GabT, and GabD, l-glutamic acid is synthesized from 2-oxoglutarate by two reactions, namely those catalyzed by PatA and by GabT. Flux enforcement by the deletion of the l-glutamic acid dehydrogenase gene gdh has been shown to improve glutarate production [15].
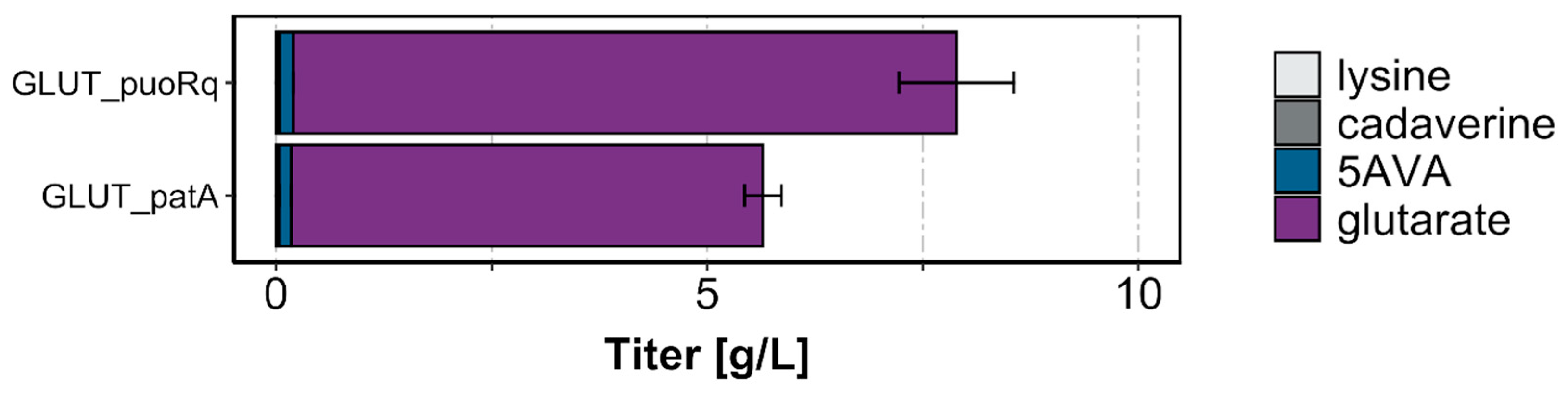
We reasoned that the beneficial effect of gdh deletion for the flux enforcement of glutarate production may be more pronounced in a five-step pathway from l-lysine to glutarate that only involves a single transaminase reaction instead of two. Therefore, we constructed a strain that expressed the oxidase gene puoRq instead of the transaminase gene patA. The GSLA2G strain, which lacked gdh coding for l-glutamic acid dehydrogenase, was transformed with plasmids for the expression of either the LdcC–PuoRq–PatD–GabT–GabD variant with a single transaminase (GLUT_puoRq) or, as a reference, the LdcC–PatA–PatD–GabT–GabD variant with two transaminases (GLUT_patA). The GLUT_puoRq strain grew slower (0.08 ± 0.00 compared to 0.11 ± 0.00 1/h, respectively) and to an about 20% lower maximal biomass concentration (3.6 ± 0.1 compared to 4.5 ± 0.2 g/L cell dry weight, respectively) than the GLUT_patA reference strain. Importantly, while the by-product formation was similar for both strains, GLUT_puoRq produced about 40% more glutarate (7.7 ± 0.7 compared to 5.5 ± 0.2 g/L, respectively; Figure 4) than the reference strain GLUT_patA. This confirmed our hypothesis that flux enforcement by gdh deletion coupled to a production pathway with a single transaminase was superior to a route using two transaminase reactions as metabolic coupling sites.
3. Discussion
This study showed that the oxidative deamination of cadaverine by putrescine oxidase Puo can operate in C. glutamicum to replace cadaverine transaminase PatA in synthetic pathways that convert cadaverine to 5AVA or glutarate. Studying glutarate biosynthesis pathways with either Puo or PatA allowed us to determine the effectiveness of flux enforcement (Figure 4). We showed how l-glutamic acid synthesis (which is required for growth) that is conducted by either one or two transamination reactions in the synthetic glutarate biosynthesis pathways can compensate for the absence of l-glutamic acid dehydrogenase caused by the deletion of its gene gdh. Glutarate production was higher when it was coupled to l-glutamic acid synthesis by one transamination (GabT) compared to a 1:2 stoichiometry, i.e., when l-glutamic acid was formed in reactions catalyzed by two transaminases (PatA and GabT) in the glutarate production pathway. Puo is not more efficient than PatA per se, because 5AVA production via the LdcC–PuoRq–PatD route was found to be lower than production via the LdcC–PatA–PatD route (Figure 2B). Thus, the better glutarate production via the LdcC–PuoRq–PatD–GabT–GabD route compared to the LdcC–PatA–PatD–GabT–GabD route (Figure 4) resulted from the better efficiency of flux enforcement with a single coupling site as compared to two coupling sites (Figure 1).
Putrescine oxidase belongs to the group of diamine oxidases found in bacteria, archaea and eukaryotes that are usually active with short aliphatic diamines in order to utilize them as carbon and/or nitrogen sources [31]. Applications in enzyme catalysis benefit from the fact that these flavoproteins utilize molecular oxygen as an electron acceptor instead of costly redox cofactors such as NAD(P)H [20,21,31,32], e.g., for the enzymatic transformation of cadaverine to 5-aminopentanal by a diamine oxidase [33]. One constraint to their application is the concomitant formation of hydrogen peroxide, an oxidative stressor that causes significant damage to the enzymes used in biocatalysis and to DNA and proteins in vivo [34]. As C. glutamicum, the production host chosen here, shows a high natural resistance to hydrogen peroxide due to its very active catalase (commercialized as CAS Number 9001-05-2 [34]), pathways operating with putrescine oxidase functioned well. However, the decrease in the growth rate of the strains with Puo-supported glutarate production in comparison to strain with PatA-supported glutarate production may be attributed to H2O2 accumulation.
Aerobic fermentation processes are often limited by low oxygen transfer rates, which may be detrimental to growth and production, in particular for biosynthesis routes requiring molecular oxygen. Since putrescine oxidase requires molecular oxygen [31,35], cultivation platforms supporting different oxygen transfer rates (Duetz microtiter plates (MTPs) and BioLector MTPs) were compared [28,29]. Production was higher in the platform with the higher oxygen transfer rates (as can be seen by comparing Figure 2B and Figure 3) in the strain where 5AVA production was based on oxidase activity. The observed benefit may not only have been due to the better provision of molecular oxygen for putrescine oxidase, since even when assessing only the production of precursor l-lysine, C. glutamicum requires a minimum oxygen transfer rate of 14 mmol L−1 h−1 for optimal biomass and l-lysine production, which cannot be reached in Duetz plates [29,36]. This oxygen bottleneck was also observed in closed tubes for an enzyme activity assay that employed the DavA–DavB route from l-lysine via 5-aminovaleramide to 5AVA, where davB encodes a l-lysine monooxygenase [12]. Thus, the better performance of the three-step LdcC–PatA–PatD pathway for the conversion of l-lysine to 5AVA compared to the variants with putrescine oxidase PuoRq or PuoPa instead of transaminase PatA (LdcC–PuoRq–PatD and LdcC–PuoPa–PatD) may also have been due the fact that transaminase PatA does not require molecular oxygen in oxygen-restricted conditions (Duetz MTPs).
The substrate spectrum of putrescine oxidase is relatively broad because, besides the C4 diamine putrescine and the C5 cadaverine, longer diamines (hexamethylenediamine, spermine, spermidine) are also accepted—though the shorter C3 diaminopropane is not [20]. This feature is not limited to the oxidases, as it is also true for the transaminases PatA and GabT (accepting substrates of different chain lengths: diamines such as putrescine and cadaverine and ω-amino acids such as GABA and 5AVA, respectively,). Transaminases, in particular, are known for their promiscuity towards a broad range of substrates [37,38], which proves advantageous in the evolutionary sense but can translate to a drawback for short-term production experiments [39]. Nevertheless, the broad substrate spectra have been employed for the metabolic engineering of diamines, ω-amino acids, and dicarboxylic acids [14], though it has had to be ensured that only a precursor of one chain length was synthesized. This specificity has been achieved by narrow spectrum decarboxylases such as l-ornithine decarboxylase yielding putrescine from l-ornithine for the C4 products or, as also used here, l-lysine decarboxylase for C5 products [10,14,40].
The cascading of enzymes in biocatalysis [41] helps to overcome the need for the purification of intermediates, to regenerate redox cofactors [42], or to displace the reaction equilibrium towards product formation when coproducts tautomerize, dimerize, cyclize, or polymerize, as has been shown for self-sufficient transamination reactions [43]. Cascaded enzymes in metabolic pathways typically allow for coupling to cellular respiration, e.g., for redox cofactor regeneration in whole-cell biotransformations [44]. For fermentation processes, flux enforcement is a powerful tool to link production to an essential metabolic pathway. In C. glutamicum, l-lysine and glutarate production titers have benefitted from this technique [6,15,19]. Similarly, the metabolic cutoffs of genes in the pentose phosphate pathway have been found to enforce the co-utilization of pentoses with methanol by complementation with genes for methanol and formaldehyde assimilation [45,46].
Here, we compared flux enforcements with two or one coupling sites, specifically to see whether the synthesis of l-glutamic acid in the absence of l-glutamic acid dehydrogenase was enabled by GabT alone or by PatA and GabT if glutarate was produced. With one coupling site (GabT alone), the titer of glutarate was increased by 40% compared to two coupling sites (Figure 4). Theoretically, a change from a 2:1 stoichiometry to a 1:1 stoichiometry in flux enforcement should have increased the glutarate titer by 100%. Similar observations were published for the flux enforcement of l-lysine production by the deletion of the genes for the TCA cycle enzyme succinyl-CoA synthetase genes, and it was hypothesized that bypassing reactions such as the glyoxylate shunt compensated, to some extent, for the lack of succinyl-CoA synthetase [19]. Similarly, bypassing reactions compensating for the lack of l-glutamic acid dehydrogenase may have reduced the beneficial effect of flux enforcement. In addition to l-glutamic acid dehydrogenase, C. glutamicum possesses the glutamine synthetase/glutamine-oxoglutarate amidotransferase (GS/GOGAT) system for l-glutamic acid synthesis and ammonium assimilation [47]. The combined reactions of glutamine synthetase and glutamine-oxoglutarate amidotransferase differ from l-glutamic acid dehydrogenase with the requirement for ATP; thus, the GS/GOGAT-encoding genes are only upon induced nitrogen starvation with basal expression at the nitrogen concentrations used here for growth and production [48,49]. Thus, it is conceivable that GS/GOGAT activity reduced the beneficial effect of flux enforcement by the gdh deletion observed here (Figure 4). The additional deletion of the GS/GOGAT genes in the gdh deletion strain is not helpful because l-glutamic acid dehydrogenase and the GS/GOGAT system are the only pathways for net ammonium assimilation, whereas transaminases only transfer ammonium from an existing donor to an oxoacid acceptor.
Glutarate production may be rationally improved through metabolic engineering targeting, e.g., precursor supply, redox cofactor regeneration, or by-product formation [50], as well as by enzyme engineering, e.g., targeting catalytic efficiency or the product inhibition of the synthetic glutarate biosynthesis pathway [51]. Recently, the application of adaptive laboratory evolution (ALE), which has revealed answers to fundamental evolutionary questions in E. coli [52,53], gained attention in strain development for growth-associated fermentative processes [54]. In C. glutamicum, ALE has been exploited for increased methanol-dependent growth and tolerance, followed by whole-genome sequencing and mutational analysis, respectively [45,55,56]. Notably, flux enforcement by gdh deletion in the glutarate-producing strains described here and previously [15] reduced the growth rate. Thus, it may be possible to accelerate glutarate production by this strain through ALE and to rationalize the selected genomic changes by reverse genetics. Importantly, reaching the industrial maturity of glutarate production by recombinant C. glutamicum genetic and metabolic robustness, as well the scaling-up of such production, remains to be achieved.
4. Materials and Methods
4.1. Microorganisms and Cultivation Conditions
The E. coli DH5α strain was used as a cloning host [57], grown in lysogeny broth (LB) at 37 °C, and supplemented with antibiotics (50 μg mL−1 kanamycin, 100 μg mL−1 spectinomycin, and 10 μg mL−1 tetracycline) when appropriate. C. glutamicum ATCC13032-derived strains were cultivated in brain heart infusion with 0.5 M sorbitol (BHIS) or an Eggeling and Bott 2005 CGXII minimal medium supplemented with 25 μg mL−1 kanamycin, 100 μg mL−1 spectinomycin, 5 μg mL−1 tetracycline, and 1 mM IPTG when appropriate. All bacterial strains and plasmids are listed in Table 1 and Table 2. For growth experiments with C. glutamicum, overnight cultures in 50 mL of BHIS were harvested and washed twice in a CGXII medium before inoculation to an OD600 of 1 and supplementation with 4% (w/v) glucose as a sole carbon source. Growth in 10 mL Duetz MTPs with culture volumes of 3 mL at 220 rpm in an Ecotron ET25-TA-RC (Infors HT, Einsbach, Germany) was monitored for the determination of the OD600 with a V-1200 Spectrophotometer (VWR, Radnor, PA, USA). The cultivations in the BioLector micro fermentation system (m2p-labs, Baesweiler, Germany) were performed in 3.2 mL FlowerPlates at 1300 rpm with filling volumes of 800, 1000, and 1200 μL.
4.2. Molecular Biology Methods
The isolation of the genomic DNA of C. glutamicum and classical methods that include the plasmid isolation, molecular cloning, and heat-shock transformation of E. coli and the electroporation of C. glutamicum were performed as described previously [58,59]. ALLin HiFi DNA Polymerase (HighQu, Kraichtal, Germany) was used to amplify DNA sequences, with plasmid or genomic DNA used as the template. The oligonucleotides that were used as primers in this study are listed in Table 3.
The genomic DNA (gDNA) of Rhodococcus qingshengii (DSM 45257) and Paenarthrobacter aurescens (DSM 20116) was obtained from the German Collection of Microorganisms and Cell Cultures (DSMZ), and the gDNA of E. coli MG1655 was isolated in [58]. The pBV2xp plasmid was digested with the restriction enzymes BamHI and SacI (New England Biolabs, Ipswich, MA, USA). The DNA fragments were joined by the means of isothermal DNA assembly [60].
To overexpress puoRq, puoPa, and patD in C. glutamicum, their genes were amplified from pBV2xp-puoRq-patD, pBV2xp-puoPa-patD, and the genomic DNA of C. glutamicum, respectively, and they were assembled into BamHI-linearized pEKEx3 and pEC-XT99A by Gibson Assembly using the respective primers. C. glutamicum was transformed with the constructed plasmids and empty vectors.
4.3. HPLC Analysis
The quantification of amino acids and cadaverine and glutarate in the cultivation medium was performed with a high-pressure liquid chromatography system (1200 series, Agilent Technologies Deutschland GmbH, Böblingen, Germany), as described previously [61]. After the centrifugation of 1 mL cell cultures at 14000 rpm for 10 min, the supernatant was stored at −20°C prior to analysis. The amino acids l-lysine and 5AVA and the diamine cadaverine were detected with a fluorescence detector (FLD G1321A, 1200 series, Agilent Technologies) after the derivatization of the samples with OPA (ortho-phthaldialdehyde). The detection of glutarate was done with a refractive index detector (RID G1362A, 1200 series, Agilent Technologies) and a diode array detector (DAD G1315B, 1200 series, Agilent Technologies).
5. Conclusions
By exchanging a putrescine transaminase with an oxidase in a synthetic glutarate pathway in C. glutamicum, a proof of concept for the production of 5AVA and glutarate was achieved. The heterologous expression of putrescine oxidase from two strains, R. qingshengii and P. aurescens, showed 5AVA production titers of 0.4 ± 0.0 and 0.1 ± 0.0 g/L 5AVA, respectively, and could be improved by the deletion of the glutarate module gabTDP and improved oxygen availability to a maximum of 3.7 ± 0.4 g/L 5AVA with the LdcC–PuoRq–PatA–GabT–GabD route. Upon the disruption of gdh, which encodes the enzyme that is responsible for the major nitrogen assimilation reaction in C. glutamicum, the focus of the flux enforcement towards glutarate production tightened on a single transaminase reaction, as compared to the two transaminases PatA and GabT, resulting in a product titer increase of 40% (7.7 ± 0.7 vs. 5.5 ± 0.2 g/L). These results highlight requirements regarding the modularity and stoichiometry of the pathway-specific flux enforcement for microbial production.
Author Contributions
C.H. and V.F.W. conceptualized this work. B.D. investigated the underlying pathways. C.H. and M.I. constructed strains. C.H. performed the experiments. C.H. and V.F.W. analyzed the data. C.H. drafted, all authors revised, and V.F.W. finalized the manuscript. V.F.W. and S.H. supervised this work. All authors have read and agreed to the final version of the manuscript.
Funding
This work was funded by the ERA CoBioTech project C1Pro (FNR-22023617; ANR-17-COBI-0003-05).
Acknowledgments
The authors thank Carina Prell and Helena Schulz-Mirbach for technical support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bioplastics Market. Available online: https://www.european-bioplastics.org/market/ (accessed on 15 September 2020).
- Ali, M.A.; Kaneko, T. Polyamide Syntheses. In Encyclopedia of Polymeric Nanomaterials; Kobayashi, S., Müllen, K., Eds.; Springer: Berlin/Heidelberg, Germany, 2015; pp. 1750–1762. ISBN 978-3-642-29647-5. [Google Scholar]
- Ligon, S.C.; Liska, R.; Stampfl, J.; Gurr, M.; Mülhaupt, R. Polymers for 3D Printing and Customized Additive Manufacturing. Chem. Rev. 2017, 117, 10212–10290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radzik, P.; Leszczyńska, A.; Pielichowski, K. Modern biopolyamide-based materials: Synthesis and modification. Polym. Bull. 2020, 77, 501–528. [Google Scholar] [CrossRef] [Green Version]
- Adkins, J.; Jordan, J.; Nielsen, D.R. Engineering Escherichia coli for renewable production of the 5-carbon polyamide building-blocks 5-aminovalerate and glutarate. Biotechnol. Bioeng. 2013, 110, 1726–1734. [Google Scholar] [CrossRef] [PubMed]
- Wendisch, V.F. Metabolic engineering advances and prospects for amino acid production. Metab. Eng. 2020, 58, 17–34. [Google Scholar] [CrossRef]
- Mimitsuka, T.; Sawai, H.; Hatsu, M.; Yamada, K. Metabolic Engineering of Corynebacterium glutamicum for Cadaverine Fermentation. Biosci. Biotechnol. Biochem. 2007, 71, 2130–2135. [Google Scholar] [CrossRef] [Green Version]
- Kind, S.; Kreye, S.; Wittmann, C. Metabolic engineering of cellular transport for overproduction of the platform chemical 1,5-diaminopentane in Corynebacterium glutamicum. Metab. Eng. 2011, 13, 617–627. [Google Scholar] [CrossRef]
- Wendisch, V.F.; Mindt, M.; Pérez-García, F. Biotechnological production of mono- and diamines using bacteria: Recent progress, applications, and perspectives. Appl. Microbiol. Biotechnol. 2018, 102, 3583–3594. [Google Scholar] [CrossRef]
- Jorge, J.M.P.; Pérez-García, F.; Wendisch, V.F. A new metabolic route for the fermentative production of 5-aminovalerate from glucose and alternative carbon sources. Bioresour. Technol. 2017. [Google Scholar] [CrossRef]
- Fothergill, J.C.; Guest, J.R. Catabolism of l-Lysine by Pseudomonas aeruginosa. J. Gen. Microbiol. 1977, 99, 139–155. [Google Scholar] [CrossRef] [Green Version]
- Rohles, C.M.; Gießelmann, G.; Kohlstedt, M.; Wittmann, C.; Becker, J. Systems metabolic engineering of Corynebacterium glutamicum for the production of the carbon-5 platform chemicals 5-aminovalerate and glutarate. Microb. Cell Factories 2016, 15. [Google Scholar] [CrossRef] [Green Version]
- Shin, J.H.; Park, S.H.; Oh, Y.H.; Choi, J.W.; Lee, M.H.; Cho, J.S.; Jeong, K.J.; Joo, J.C.; Yu, J.; Park, S.J.; et al. Metabolic engineering of Corynebacterium glutamicum for enhanced production of 5-aminovaleric acid. Microb. Cell Factories 2016, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chae, T.U.; Ahn, J.H.; Ko, Y.-S.; Kim, J.W.; Lee, J.A.; Lee, E.H.; Lee, S.Y. Metabolic engineering for the production of dicarboxylic acids and diamines. Metab. Eng. 2020, 58, 2–16. [Google Scholar] [CrossRef] [PubMed]
- Pérez-García, F.; Jorge, J.M.P.; Dreyszas, A.; Risse, J.M.; Wendisch, V.F. Efficient Production of the Dicarboxylic Acid Glutarate by Corynebacterium glutamicum via a Novel Synthetic Pathway. Front. Microbiol. 2018, 9, 2589. [Google Scholar] [CrossRef] [PubMed]
- Rohles, C.M.; Gläser, L.; Kohlstedt, M.; Gießelmann, G.; Pearson, S.; del Campo, A.; Becker, J.; Wittmann, C. A bio-based route to the carbon-5 chemical glutaric acid and to bionylon-6,5 using metabolically engineered Corynebacterium glutamicum. Green Chem. 2018, 20, 4662–4674. [Google Scholar] [CrossRef] [Green Version]
- Smirnov, S.V.; Kodera, T.; Samsonova, N.N.; Kotlyarova, V.A.; Rushkevich, N.Y.; Kivero, A.D.; Sokolov, P.M.; Hibi, M.; Ogawa, J.; Shimizu, S. Metabolic engineering of Escherichia coli to produce (2S, 3R, 4S)-4-hydroxyisoleucine. Appl. Microbiol. Biotechnol. 2010, 88, 719–726. [Google Scholar] [CrossRef]
- Theodosiou, E.; Breisch, M.; Julsing, M.K.; Falcioni, F.; Bühler, B.; Schmid, A. An artificial TCA cycle selects for efficient α-ketoglutarate dependent hydroxylase catalysis in engineered Escherichia coli: Strain Design for Proline Hydroxylation in Vivo. Biotechnol. Bioeng. 2017, 114, 1511–1520. [Google Scholar] [CrossRef] [Green Version]
- Kind, S.; Becker, J.; Wittmann, C. Increased lysine production by flux coupling of the tricarboxylic acid cycle and the lysine biosynthetic pathway—Metabolic engineering of the availability of succinyl-CoA in Corynebacterium glutamicum. Metab. Eng. 2013, 15, 184–195. [Google Scholar] [CrossRef]
- van Hellemond, E.W.; van Dijk, M.; Heuts, D.P.H.M.; Janssen, D.B.; Fraaije, M.W. Discovery and characterization of a putrescine oxidase from Rhodococcus erythropolis NCIMB 11540. Appl. Microbiol. Biotechnol. 2008, 78, 455–463. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.-I.; Jang, J.-H.; Yu, M.-J.; Kim, Y.-W. Construction of a Bifunctional Enzyme Fusion for the Combined Determination of Biogenic Amines in Foods. J. Agric. Food Chem. 2013, 61, 9118–9124. [Google Scholar] [CrossRef]
- Baumgart, M.; Unthan, S.; Rückert, C.; Sivalingam, J.; Grünberger, A.; Kalinowski, J.; Bott, M.; Noack, S.; Frunzke, J. Construction of a Prophage-Free Variant of Corynebacterium glutamicum ATCC 13032 for Use as a Platform Strain for Basic Research and Industrial Biotechnology. Appl. Environ. Microbiol. 2013, 79, 6006–6015. [Google Scholar] [CrossRef] [Green Version]
- Pérez-García, F.; Peters-Wendisch, P.; Wendisch, V.F. Engineering Corynebacterium glutamicum for fast production of l-lysine and l-pipecolic acid. Appl. Microbiol. Biotechnol. 2016, 100, 8075–8090. [Google Scholar] [CrossRef] [PubMed]
- Unthan, S.; Baumgart, M.; Radek, A.; Herbst, M.; Siebert, D.; Brühl, N.; Bartsch, A.; Bott, M.; Wiechert, W.; Marin, K.; et al. Chassis organism from Corynebacterium glutamicum—A top-down approach to identify and delete irrelevant gene clusters. Biotechnol. J. 2015, 10, 290–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engels, V.; Lindner, S.N.; Wendisch, V.F. The global repressor SugR controls expression of genes of glycolysis and of the l-lactate dehydrogenase LdhA in Corynebacterium glutamicum. J. Bacteriol. 2008, 190, 8033–8044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, A.Q.D.; Schneider, J.; Wendisch, V.F. Elimination of polyamine N-acetylation and regulatory engineering improved putrescine production by Corynebacterium glutamicum. J. Biotechnol. 2015, 201, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Lubitz, D.; Jorge, J.M.P.; Pérez-García, F.; Taniguchi, H.; Wendisch, V.F. Roles of export genes cgmA and lysE for the production of l-arginine and l-citrulline by Corynebacterium glutamicum. Appl. Microbiol. Biotechnol. 2016, 100, 8465–8474. [Google Scholar] [CrossRef]
- Funke, M.; Diederichs, S.; Kensy, F.; Müller, C.; Büchs, J. The Baffled Microtiter Plate: Increased Oxygen Transfer and Improved Online Monitoring in Small Scale Fermentations. Biotechnol. Bioeng. 2009, 103, 1118–1128. [Google Scholar] [CrossRef]
- Duetz, W.A.; Rüedi, L.; Hermann, R.; O’Connor, K.; Büchs, J.; Witholt, B. Methods for Intense Aeration, Growth, Storage, and Replication of Bacterial Strains in Microtiter Plates. Appl. Environ. Microbiol. 2000, 66, 2641–2646. [Google Scholar] [CrossRef] [Green Version]
- Tesch, M.; de Graaf, A.A.; Sahm, H. In Vivo Fluxes in the Ammonium-Assimilatory Pathways in Corynebacterium glutamicum Studied by 15N Nuclear Magnetic Resonance. Appl. Environ. Microbiol. 1999, 65, 1099–1109. [Google Scholar] [CrossRef] [Green Version]
- Floris, G.; Finazzi Agrò, A. Amine Oxidases. In Encyclopedia of Biological Chemistry; Elsevier: Amsterdam, The Netherlands, 2013; pp. 87–90. ISBN 978-0-12-378631-9. [Google Scholar]
- Lee, J.-I.; Kim, Y.-W. Characterization of amine oxidases from Arthrobacter aurescens and application for determination of biogenic amines. World J. Microbiol. Biotechnol. 2013, 29, 673–682. [Google Scholar] [CrossRef]
- Nau, W.M.; Ghale, G.; Hennig, A.; Bakirci, H.; Bailey, D.M. Substrate-Selective Supramolecular Tandem Assays: Monitoring Enzyme Inhibition of Arginase and Diamine Oxidase by Fluorescent Dye Displacement from Calixarene and Cucurbituril Macrocycles. J. Am. Chem. Soc. 2009, 131, 11558–11570. [Google Scholar] [CrossRef]
- Milse, J.; Petri, K.; Rückert, C.; Kalinowski, J. Transcriptional response of Corynebacterium glutamicum ATCC 13032 to hydrogen peroxide stress and characterization of the OxyR regulon. J. Biotechnol. 2014, 190, 40–54. [Google Scholar] [CrossRef]
- Romero, E.; Gómez Castellanos, J.R.; Gadda, G.; Fraaije, M.W.; Mattevi, A. Same Substrate, Many Reactions: Oxygen Activation in Flavoenzymes. Chem. Rev. 2018, 118, 1742–1769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Käß, F.; Prasad, A.; Tillack, J.; Moch, M.; Giese, H.; Büchs, J.; Wiechert, W.; Oldiges, M. Rapid assessment of oxygen transfer impact for Corynebacterium glutamicum. Bioprocess Biosyst. Eng. 2014, 37, 2567–2577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, R.A. Enzyme Recruitment in Evolution of New Function. Annu. Rev. Microbiol. 1976, 30, 409–425. [Google Scholar] [CrossRef] [Green Version]
- Wilding, M.; Peat, T.S.; Kalyaanamoorthy, S.; Newman, J.; Scott, C.; Jermiin, L.S. Reverse engineering: Transaminase biocatalyst development using ancestral sequence reconstruction. Green Chem. 2017, 19, 5375–5380. [Google Scholar] [CrossRef] [Green Version]
- Atkins, W.M. Biological messiness vs. biological genius: Mechanistic aspects and roles of protein promiscuity. J. Steroid Biochem. Mol. Biol. 2015, 151, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Jorge, J.M.P.; Leggewie, C.; Wendisch, V.F. A new metabolic route for the production of gamma-aminobutyric acid by Corynebacterium glutamicum from glucose. Amino Acids 2016, 48, 2519–2531. [Google Scholar] [CrossRef]
- Gröger, H. Biocatalytic concepts for synthesizing amine bulk chemicals: Recent approaches towards linear and cyclic aliphatic primary amines and ω-substituted derivatives thereof. Appl. Microbiol. Biotechnol. 2019, 103, 83–95. [Google Scholar] [CrossRef]
- Breuer, M.; Ditrich, K.; Habicher, T.; Hauer, B.; Keßeler, M.; Stürmer, R.; Zelinski, T. Industrial Methods for the Production of Optically Active Intermediates. Angew. Chem. Int. Ed. 2004, 43, 788–824. [Google Scholar] [CrossRef]
- Grigoriou, S.; Kugler, P.; Kulcinskaja, E.; Walter, F.; King, J.; Hill, P.; Wendisch, V.F.; O’Reilly, E. Development of a Corynebacterium glutamicum bio-factory for self-sufficient transaminase reactions. Green Chem. 2020, 22, 4128–4132. [Google Scholar] [CrossRef]
- Klatte, S.; Wendisch, V.F. Redox self-sufficient whole cell biotransformation for amination of alcohols. Bioorganic Med. Chem. 2014, 22, 5578–5585. [Google Scholar] [CrossRef] [PubMed]
- Hennig, G.; Haupka, C.; Brito, L.F.; Rückert, C.; Cahoreau, E.; Heux, S.; Wendisch, V.F. Methanol-Essential Growth of Corynebacterium glutamicum: Adaptive Laboratory Evolution Overcomes Limitation due to Methanethiol Assimilation Pathway. Int. J. Mol. Sci. 2020, 21, 3617. [Google Scholar] [CrossRef] [PubMed]
- Tuyishime, P.; Wang, Y.; Fan, L.; Zhang, Q.; Li, Q.; Zheng, P.; Sun, J.; Ma, Y. Engineering Corynebacterium glutamicum for methanol-dependent growth and glutamate production. Metab. Eng. 2018, 49, 220–231. [Google Scholar] [CrossRef] [PubMed]
- Burkovski, A. Nitrogen Metabolism and Its Regulation. In Handbook of Corynebacterium Glutamicum; Eggeling, L., Bott, M., Eds.; CRC Press LLC: Boca Raton, FL, USA, 2005; pp. 335–352. ISBN 0-8493-1821-1. [Google Scholar]
- Tesch, M.; Eikmanns, B.J.; de Graaf, A.A.; Sahm, H. Ammonia assimilation in Corynebacterium glutamicum and a glutamate dehydrogenase-deficient mutant. Biotechnol. Lett. 1998, 20, 953–957. [Google Scholar] [CrossRef]
- Nolden, L.; Farwick, M.; Krämer, R.; Burkovski, A. Glutamine synthetases of Corynebacterium glutamicum: Transcriptional control and regulation of activity. FEMS Microbiol. Lett. 2001, 201, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Wendisch, V.F.; Lee, J.-H. Metabolic Engineering in Corynebacterium glutamicum. In Corynebacterium Glutamicum; Inui, M., Toyoda, K., Eds.; Microbiology Monographs; Springer International Publishing: Cham, Switzerland, 2020; Volume 23, pp. 287–322. ISBN 978-3-030-39266-6. [Google Scholar]
- Arnold, F.H. Innovation by Evolution: Bringing New Chemistry to Life (Nobel Lecture). Angew. Chem. Int. Ed. 2019, 58, 14420–14426. [Google Scholar] [CrossRef] [Green Version]
- Tenaillon, O.; Barrick, J.E.; Ribeck, N.; Deatherage, D.E.; Blanchard, J.L.; Dasgupta, A.; Wu, G.C.; Wielgoss, S.; Cruveiller, S.; Médigue, C.; et al. Tempo and mode of genome evolution in a 50,000-generation experiment. Nature 2016, 536, 165–170. [Google Scholar] [CrossRef] [Green Version]
- Barrick, J.E.; Yu, D.S.; Yoon, S.H.; Jeong, H.; Oh, T.K.; Schneider, D.; Lenski, R.E.; Kim, J.F. Genome evolution and adaptation in a long-term experiment with Escherichia coli. Nature 2009, 461, 1243–1247. [Google Scholar] [CrossRef]
- Dragosits, M.; Mattanovich, D. Adaptive laboratory evolution—Principles and applications for biotechnology. Microb. Cell Factories 2013, 12, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Fan, L.; Tuyishime, P.; Liu, J.; Zhang, K.; Gao, N.; Zhang, Z.; Ni, X.; Feng, J.; Yuan, Q.; et al. Adaptive laboratory evolution enhances methanol tolerance and conversion in engineered Corynebacterium glutamicum. Commun. Biol. 2020, 3. [Google Scholar] [CrossRef]
- Leßmeier, L.; Wendisch, V.F. Identification of two mutations increasing the methanol tolerance of Corynebacterium glutamicum. BMC Microbiol. 2015, 15. [Google Scholar] [CrossRef] [Green Version]
- Hanahan, D. Techniques for transformation of E. coli. DNA Cloning Pract. Approach 1985, 1, 109–135. [Google Scholar]
- Eikmanns, B.J.; Thum-Schmitz, N.; Eggeling, L.; Lüdtke, K.U.; Sahm, H. Nucleotide sequence, expression and transcriptional analysis of the Corynebacterium glutamicum gltA gene encoding citrate synthase. Microbiology 1994, 140, 1817–1828. [Google Scholar] [CrossRef] [Green Version]
- Simon, R.; Priefer, U.; Pühler, A. A broad host range mobilization system for in vivo genetic engineering: Transposon mutagenesis in Gram negative bacteria. Bio/Technology 1983, 1, 784–791. [Google Scholar] [CrossRef]
- Gibson, D.G.; Young, L.; Chuang, R.-Y.; Venter, J.C.; Hutchison, C.A.; Smith, H.O. Enzymatic assembly of DNA molecules up to several hundred kilobases. Nat. Methods 2009, 6, 343–345. [Google Scholar] [CrossRef]
- Schneider, J.; Niermann, K.; Wendisch, V.F. Production of the amino acids l-glutamate, l-lysine, l-ornithine and l-arginine from arabinose by recombinant Corynebacterium glutamicum. J. Biotechnol. 2011, 154, 191–198. [Google Scholar] [CrossRef]
Figure 1.
Fermentative production of 5-aminovalerate (5AVA) and glutarate. (A) A novel three-step metabolic pathway for production of 5AVA and glutarate was established by the exchange of a transaminase of an existing three-step pathway (PatA) with an oxidase (Puo). The anticipated shift of net nitrogen assimilation in the form of l-glutamic acid is counteracted by (B) endogenous l-glutamate synthesis by l-glutamic acid dehydrogenase(Gdh), which was deleted to enforce the flux towards glutarate. Pink areas depict decarboxylation reactions. Yellow areas depict transamination steps. Orange areas depict deamination steps. Blue areas depict oxidations. Red-shadowed genes (gabT, γ-aminobutyrate (GABA)/5AVA amino transferase; gabD, and succinate/glutarate-semialdehyde dehydrogenase) are native genes of Corynebacterium glutamicum. Additionally, copies of gabTD from Pseudomonas stutzeri were added by heterologous overexpression. The grey-shadowed gene (gdh, l-glutamic acid dehydrogenase) is native in C. glutamicum. The dark green-shadowed genes (ldcC, l-lysine decarboxylase; patA, putrescine transaminase; and patD, γ-aminobutyraldehyde dehydrogenase) are originally from Escherichia coli and were added by heterologous overexpression. The blue-shadowed gene (puo, putrescine oxidase; EC 1.4.3.10) was originally from Rhodococcus qingshengii and Paenarthrobacter aurescens, and it was heterologously expressed in C. glutamicum. l-glutamic acid is highlighted in bright green. OG: 2-oxoglutarate, Glu: l-glutamatic acid.
Figure 1.
Fermentative production of 5-aminovalerate (5AVA) and glutarate. (A) A novel three-step metabolic pathway for production of 5AVA and glutarate was established by the exchange of a transaminase of an existing three-step pathway (PatA) with an oxidase (Puo). The anticipated shift of net nitrogen assimilation in the form of l-glutamic acid is counteracted by (B) endogenous l-glutamate synthesis by l-glutamic acid dehydrogenase(Gdh), which was deleted to enforce the flux towards glutarate. Pink areas depict decarboxylation reactions. Yellow areas depict transamination steps. Orange areas depict deamination steps. Blue areas depict oxidations. Red-shadowed genes (gabT, γ-aminobutyrate (GABA)/5AVA amino transferase; gabD, and succinate/glutarate-semialdehyde dehydrogenase) are native genes of Corynebacterium glutamicum. Additionally, copies of gabTD from Pseudomonas stutzeri were added by heterologous overexpression. The grey-shadowed gene (gdh, l-glutamic acid dehydrogenase) is native in C. glutamicum. The dark green-shadowed genes (ldcC, l-lysine decarboxylase; patA, putrescine transaminase; and patD, γ-aminobutyraldehyde dehydrogenase) are originally from Escherichia coli and were added by heterologous overexpression. The blue-shadowed gene (puo, putrescine oxidase; EC 1.4.3.10) was originally from Rhodococcus qingshengii and Paenarthrobacter aurescens, and it was heterologously expressed in C. glutamicum. l-glutamic acid is highlighted in bright green. OG: 2-oxoglutarate, Glu: l-glutamatic acid.

Figure 2.
Production titers of 5AVA and by-products by strains derived from (A) C. glutamicum GSLA2(pVWEx1-ldcC) and (B) GSLA2ΔgabTDP(pVWEx1-ldcC). Three milliliter cultures were grown in the Duetz system with a 4% glucose minimal medium supplemented with 1 mM isopropyl β- d-1-thiogalactopyranoside (IPTG) and harvested after 48 h. Values and error bars represent mean and standard deviation values (n = 3 cultivations).
Figure 2.
Production titers of 5AVA and by-products by strains derived from (A) C. glutamicum GSLA2(pVWEx1-ldcC) and (B) GSLA2ΔgabTDP(pVWEx1-ldcC). Three milliliter cultures were grown in the Duetz system with a 4% glucose minimal medium supplemented with 1 mM isopropyl β- d-1-thiogalactopyranoside (IPTG) and harvested after 48 h. Values and error bars represent mean and standard deviation values (n = 3 cultivations).

Figure 3.
Production titers of 5AVA and by-products by strains derived from C. glutamicum GSLA2ΔgabTDP (pVWEx1-ldcC). Cells were grown in a BioLector using different culture volumes (800–1200 μL) of a 4% glucose minimal medium supplemented with 1 mM IPTG and harvested after 48 h. Values and error bars represent mean and standard deviation values (n = 3 cultivations).
Figure 3.
Production titers of 5AVA and by-products by strains derived from C. glutamicum GSLA2ΔgabTDP (pVWEx1-ldcC). Cells were grown in a BioLector using different culture volumes (800–1200 μL) of a 4% glucose minimal medium supplemented with 1 mM IPTG and harvested after 48 h. Values and error bars represent mean and standard deviation values (n = 3 cultivations).

Figure 4.
Production titers of glutarate and precursors by strains GLUT_puoRq and GLUT_patA. Cells were grown in the BioLector with a 4% glucose minimal medium supplemented with 1 mM IPTG and harvested after 72 h. Values and error bars represent mean and standard deviation values (n = 3 cultivations).
Figure 4.
Production titers of glutarate and precursors by strains GLUT_puoRq and GLUT_patA. Cells were grown in the BioLector with a 4% glucose minimal medium supplemented with 1 mM IPTG and harvested after 72 h. Values and error bars represent mean and standard deviation values (n = 3 cultivations).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Bacterial strains used in this study.
| Strain | Relevant Characteristics | Reference |
|---|---|---|
| E. coli DH5α | ∆lacU169 (φ80lacZ ∆M15), supE44, hsdR17, recA1, endA1, gyrA96, thi-1, relA1 | [54] |
| C. glutamicum GRLys1 (DM1933ΔCGP123) | C. glutamicum ATCC13032 with modifications: Δpck, pycP458S, homV59A, 2 copies of lysCT311I, 2 copies of asd, 2 copies of dapA, 2 copies of dapB, 2 copies of ddh, 2 copies of lysA, 2 copies of lysE, in-frame deletion of prophages CGP1 (cg1507-cg1524), CGP2 (cg1746-cg1752) and CGP3 (cg1890-cg2071). | [55] |
| GSLA2 | GRLys1 with in-frame deletions: sugR (cg2115), ldhA (cg3219), snaA (cg1722), cgmA (cg2893) | [15] |
| GSLA2ΔgabTDP | GSLA2 with in-frame deletions: gabT, gabD and gabP (cg0566-cg0568) | [10] |
| GSLA2G | GSLA2 with in-frame deletion: gdh (cg2280) | [15] |
| AVA1 | GSLA2(pVWEx1-ldcC)(pEC-XT99A) | [10] |
| AVA1_patA | GSLA2(pVWEx1-ldcC)(pEKEx3-patDA) | [10] |
| AVA1_puoPa | GSLA2(pVWEx1-ldcC)(pEC-XT99A-puoPa-patD) | This study |
| AVA1_puoRq | GSLA2(pVWEx1-ldcC)(pEC-XT99A-puoRq-patD) | This study |
| AVA2 | GSLA2ΔgabTDP(pVWEx1-ldcC)(pEC-XT99A) | [15] |
| AVA2_patA | GSLA2ΔgabTDP(pVWEx1-ldcC)(pEKEx3-patDA) | [15] |
| AVA2_puoPa | GSLA2ΔgabTDP(pVWEx1-ldcC)(pEC-XT99A-puoPa-patD) | This study |
| AVA2_puoRq | GSLA2ΔgabTDP(pVWEx1-ldcC)(pEC-XT99A-puoRq-patD) | This study |
| GLUT_patA | GSLA2G(pVWEx1-ldcC)(pEKEx3-patDA)(pEC-XT99A-gabTDPstu) | [15] |
| GLUT_puoRq | GSLA2G(pVWEx1-ldcC)(pEKEx3-puoRq-patD)(pEC-XT99A-gabTDPstu) | This study |
Table 2.
Plasmids used in this study.
| Plasmid | Relevant Characteristics | Reference |
|---|---|---|
| pBV2xp | AmpR, KanR, B. methanolicus/E. coli shuttle vector, pHCMC04 derivative, Pxyl from B. megaterium | [56] |
| pBV2xp-puoRq-patD | pBV2xp-expressing puo from R. qingshengii and patD from E. coli MG1655 | This study |
| pBV2xp-puoPa-patD | pBV2xp-expressing puo from P. aurescens and patD from E. coli MG1655 | This study |
| pEC-XT99A | TetR, C. glutamicum/E. coli shuttle vector (Ptrc, lacIq, pGA1 oriVCg) | [57] |
| pEC-XT99A-gabTDPstu | pEC-XT99A-expressing gabT and gabD from Pseudomonas stutzerii ATCC17588 | [15] |
| pEC-XT99A-puoPa-patD | pEC-XT99A-expressing puo from P. aurescens and patD from E. coli MG1655 | This study |
| pEC-XT99A-puoRq-patD | pEC-XT99A-expressing puo from R. qingshengii and patD from E. coli MG1655 | This study |
| pEKEx3 | SpecR, C. glutamicum/E. coli shuttle vector (Ptac lacIq pBL1, oriVEc) | [58] |
| pEKEx3-patDA | pEKEx3-expressing patD and patA from E. coli MG1655 | [15] |
| pEKEx3-puoRq-patD | pEKEx3-expressing puo from R. qingshengii and patD from E. coli MG1655 | This study |
| pVWEx1-ldcC | pVWEx1-expressing ldcC from E. coli MG1655 | [38] |
Table 3.
Oligonucleotides used as primers in this study.
| Primer | Sequence (5′–3′) | Description |
|---|---|---|
| AVA25 | ttcacttaagggggaaatggcaaatgcagaatcttgatcgcgacgttgtgatcgtcgg | Putrescine oxidase gene from P. aurescens 579_fw |
| AVA30 | tcttactacctcctatttatgtaattgtttactcaggcgacaggtacagaagccaacttgtt | Putrescine oxidase gene from P. aurescens 579_rv |
| AVA27 | ttcacttaagggggaaatggcaaatgcctactctccagagagacgttgcaatcgt | Putrescine oxidase gene from R. qingshengii djl-6-2_fw |
| AVA31 | tcttactacctcctatttatgtaattgtttactcaggccttgctgcgagcgatgatgt | Putrescine oxidase gene from R. qingshengii djl-6-2_rv |
| AVA32 | gtaaacaattacataaataggaggtagtaagaatgcaacataagttactgattaacggagaactggttag | γ-aminobutyraldehyde dehydrogenase gene from E. coli MG1655_fw |
| AVA33 | acgacggccagtgaattcgagctttaatgtttaaccatgacgtggcggacga | γ-aminobutyraldehyde dehydrogenase gene from E. coli MG1655_rv |
| AVA43 | agctggacaccatctccttc | Sequencing of pBV2xp-puoPa-patD |
| AVA44 | gcgcttacgcttccagctac | Sequencing of pBV2xp-puoPa-patD |
| AVA45 | ggcaggcgtgattaacatac | Sequencing of pBV2xp-puoPa-patD and pBV2xp-puoRq-patD |
| AVA48 | cgcgatctcgacacagtctc | Sequencing of pBV2xp-puoRq-patD |
| AVA49 | caccgctacggcgcggattc | Sequencing of pBV2xp-puoRq-patD |
| PXPF | tgtttatccaccgaactaag | Colony PCR primer for pBV2xp_fw |
| BVXR | ccgcacagatgcgtaaggag | Colony PCR primer for pBV2xp_rv |
| patD_F | ccctgtcgggaaattagaagaaaaggaggttttttatgcaacataagttactgattaacggagaactgg | Construction of pEC-XT99A/pEKEx3-puoRq-patD_fw |
| patD_R | cctgcaggtcgactctagagttaatgtttaaccatgacgtggcgg | Construction of pEC-XT99A/pEKEx3-puoRq-patD_rv |
| puoRq_F | attcgagctcggtacccgggccatattcacaaccctaattataaaggaggtcttttatgcctactctccagagagacg | Construction of pEC-XT99A-puoRq-patD_fw |
| puoRq_R | cttctaatttcccgacagggtcaggccttgctgcgagc | Construction of pEC-XT99A-puoRq-patD_rv |
| puoPa_F | attcgagctcggtacccgggcggcccggtaacggccaacagtagaaaggaggtattttatgcagaatcttgatcgcgacg | Construction of pEC-XT99A-puoPa-patD_fw |
| puoPa_R | cttctaatttcccgacagggtcaggcgacaggtacagaagcc | Construction of pEC-XT99A-puoPa-patD_rv |
| puoRq_F2 | cctgcaggtcgactctagagccatattcacaaccctaattataaaggaggtcttttatgcctactctccagagagacg | Construction of pEKEx3-puoRq-patD_fw |
| patD_R2 | attcgagctcggtacccgggttaatgtttaaccatgacgtggcgg | Construction of pEKEx3-puoRq-patD_rv |
| pEC_F | gcgccgacatcataacgg | Colony PCR primer for pEC-XT99A_fw |
| pEC_R | ggcgtttcacttctgagttcgg | Colony PCR primer for pEC-XT99A_rv |
| pEK_F | cgcttccactttttcccgcgt | Colony PCR primer for pEKEx3_fw |
| pEK_R | gcatttatcagggttattgtc | Colony PCR primer for pEKEx3_rv |
| seq_patD | atcgcaccgtggaattatccgc | Sequencing of pEC-XT99A/pEKEx3-puoRq-patD |
| seq1_Rq | gcgatctcgacacagtctcc | Sequencing of pEC-XT99A/pEKEx3-puoRq-patD |
| seq2_Rq | caacaccaaccacgaggacg | Sequencing of pEC-XT99A/pEKEx3-puoRq-patD |
| seq1_Pa | tggacaccatctccttccacc | Sequencing of pEC-XT99A-puoPa-patD |
| seq2_Pa | acaccaaccacggagattcc | Sequencing of pEC-XT99A-puoPa-patD |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Haupka, C.; Delépine, B.; Irla, M.; Heux, S.; Wendisch, V.F. Flux Enforcement for Fermentative Production of 5-Aminovalerate and Glutarate by Corynebacterium glutamicum. Catalysts 2020, 10, 1065. https://doi.org/10.3390/catal10091065
AMA Style
Haupka C, Delépine B, Irla M, Heux S, Wendisch VF. Flux Enforcement for Fermentative Production of 5-Aminovalerate and Glutarate by Corynebacterium glutamicum. Catalysts. 2020; 10(9):1065. https://doi.org/10.3390/catal10091065
Chicago/Turabian StyleHaupka, Carsten, Baudoin Delépine, Marta Irla, Stephanie Heux, and Volker F. Wendisch. 2020. "Flux Enforcement for Fermentative Production of 5-Aminovalerate and Glutarate by Corynebacterium glutamicum" Catalysts 10, no. 9: 1065. https://doi.org/10.3390/catal10091065
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.