Comparative Study on Enzyme Immobilization Using Natural Hydrogel Matrices—Experimental Studies Supported by Molecular Models Analysis


Department of Micro, Nano and Bioprocess Engineering, Faculty of Chemistry, Wrocław University of Science and Technology, Norwida 4/6, 50-373 Wrocław, Poland
*
Author to whom correspondence should be addressed.
Catalysts 2020, 10(5), 489; https://doi.org/10.3390/catal10050489
Submission received: 12 March 2020
/
Revised: 10 April 2020
/
Accepted: 25 April 2020
/
Published: 1 May 2020
(This article belongs to the Special Issue Immobilization of Enzymes)
Abstract
:Currently, great attention is focused on conducting manufacture processes using clean and eco-friendly technologies. This research trend also relates to the production of immobilized biocatalysts of industrial importance using matrices and methods that fulfill specified operational and environmental requirements. For that reason, hydrogels of natural origin and the entrapment method become increasingly popular in terms of enzyme immobilization. The presented work is the comparative research on invertase immobilization using two natural hydrogel matrices—alginate and gelatin. During the study, we provided the molecular insight into the structural characteristics of both materials regarding their applicability as effective enzyme carriers. In order to confirm our predictions of using these hydrogels for invertase immobilization, we performed the typical experimental studies. In this case, the appropriate conditions of enzyme entrapment were selected for both types of carrier. Next, the characterization of received invertase preparations was made. As a final experimental result, the gelatin-based hydrogel was selected as an effective carrier for invertase immobilization. Hereby, using mild conditions and a pro-ecological, biodegradable matrix, it was possible to obtain very stable and reactive biocatalyst. The choice of gelatin-immobilized invertase preparation was compatible with our predictions based on the molecular models of hydrogel matrices and enzyme used.

1. Introduction
There is no doubt that the practical application of enzymes has been the significant technological issue subjected to extensive studies for many years. One of the reasons is a constantly growing importance of the industrial potential of enzymes. They are mainly applied as effective catalysts of various biotransformations, resulting in plenty of commercially justified products [1,2,3,4]. Biocatalysis is widely used in many fields, including food processing, manufacture of fine chemicals, medical diagnostics and therapies, and cosmetics and pharmaceutical industries, as well as agriculture and environmental protection. Nevertheless, the areas directly related to human health (i.e., food processing, production of dietary supplements, drugs, and other therapeutic compounds) are carefully monitored in terms of biocatalysts application. In this case, the crucial point is to ensure the required purity of the final products. This is enforced by strictly defined regulations developed by the Food and Drug Administration (FDA) [5].
In regard to typical chemical catalysts, the application of enzymes brings many tangible benefits—both to the efficiency of a given process and to the protection of the natural environment. The most important include the following: high chemo-, stereo-, and regioselectivity; mild operational conditions; and lack of formation of toxic by-products during catalyzed reaction [6,7,8]. However, the application of enzymes in their native form is limited by the purification costs, sensitivity to environmental factors, and relatively low operational stability, followed by its disposability [9]. To overcome these drawbacks, various methods improving the stability of enzymes have been developed [6,10,11,12]. The most commonly used is immobilization, involving binding the catalyst molecules to the surface or trapping them in the volume of the given carrier [6,7,8,9,13,14], as well as the formation of cross-linked aggregates (CLEAs) or crystals (CLECs) without the additional use of an insoluble matrix [15,16,17,18]. Beside increasing the stability of enzymes [3,14], immobilization offers many other tangible benefits, such as enzyme reusability without significant decline in its catalytic activity and less susceptibility to denaturation caused by various environmental factors (i.e., temperature, pH, ionic strength) [7,8,13].
In order to select the suitable carrier and immobilization method, several important parameters of bound enzymes should be monitored. The most significant are catalytic activity and thermal, pH, operational, and storage stability. The next very important factor is also the degree of enzyme release from the carrier, which is an undesirable phenomenon in the industrial applications. Summing up, an ideal immobilized biocatalyst for commercial utilization should be characterized by the following: (i) durable binding to the carrier, (ii) non-toxicity, (iii) sufficient resistance to physico-chemical factors of the microenvironment, (iv) high catalytic activity, and (v) reusability or (vi) possibility to use in a continuous mode [14,19].
In fact, increasingly more attention is being paid to conducting manufacture processes using clean and eco-friendly technologies. This aspect closely relates to the production of immobilized biocatalysts. In this case, special attention is paid primarily to the proper selection of matrices that should fulfill required operational and environmental demands. Referring to this ecological trend, hydrogels of natural origin are becoming increasingly popular in terms of enzyme immobilization [20,21,22]. At present, this group of materials is the subject of intensive research as matrices for the production of biocatalysts for industrial importance—especially in the field of food processing [20,23,24,25,26,27]. The indisputable advantages of using natural hydrogels are as follows: (i) eco-friendly nature, (ii) non-toxicity, (iii) biodegradability, (iv) mild manufacturing conditions, and (v) relatively low production costs [28,29,30]. One of the most commonly used methods of immobilization using these matrices is trapping the enzyme molecules within the gel lattice [8,31]. Extensive application of this procedure results from its relatively simple protocol, rather than low costs and time expenditures. Moreover, using this method, only minor changes in the native structure of the enzyme could appear. That would enable it to retain the full catalytic functionality after immobilization.
Currently, a relatively new trend in research on the immobilization of enzymes using hydrogels is observed. That is, the theoretical analysis of virtually designed structural models of these materials to check their potential suitability for effective retention of biocatalyst molecules [32,33,34,35,36]. This multi-level approach could be a versatile practical tool. First of all, it enables one to determine the key parameters of the hydrogel network of different chemical composition (e.g., equilibrium swelling degree, Young modulus, Flory–Huggins interaction parameter, degree of cross-linking, density, permeability, pore size distribution) that are essential to determine their suitability for use as matrices for immobilization [37,38,39,40,41]. Secondly, it allows one to track the potential mechanism of immobilization of a given enzyme at the molecular level in parallel for a few carriers. Finally, by performing screening tests in virtual space, it allows for a significant reduction in costs and time of preliminary experimental investigations. With this approach, there is no need to perform research using the classical trial and error method, and actual experiments can be limited only to strictly defined narrow conditions selected in computational studies.
The main aim of our study was to analyze molecular models of two different hydrogel materials of natural origin (sodium alginate and gelatin) in terms of their effective use as immobilization carriers for the model enzyme (invertase from Saccharomyces cerevisiae). Then, by tacit analysis of the molecular structure of hydrogel matrices and the enzyme, provide hypothesis about their structural and chemical properties. Thus, optimization of their application could be performed by comparing the outcomes from theoretical considerations with the results obtained in standard experimental studies of enzyme immobilization.
Our goal was to investigate diverse systems. Therefore, the chosen materials completely differ in the cross-linking mechanism. In the case of sodium alginate, the formation of so-called pseudogels is observed (physical hydrogels). The cross-linking mechanism is based on the exchange of monovalent to divalent ion (most frequently from Na+ to Ca2+) [36,42,43,44]. This process results in the creation of a three-dimensional polymeric structure maintained by reversible ionic interactions [44,45,46]. In turn, gelatin-based materials are mostly cross-linked chemically and defined as permanent hydrogels. In this case, stable hydrogels are obtained by the formation of covalent bonds between polymeric gelatin chains in the presence of a specific cross-linking agent (e.g., microbial transglutaminase, EC 2.3.2.13) [47,48,49,50]. Considering the appropriate selection of a model enzyme, the rather high permeability of alginate-based materials was taken into account [51,52]. Therefore, we applied the invertase from Saccharomyces cerevisiae—an enzyme with relatively high molecular weight (MW ≈ 428 kDa [53]).
Our research represents a multi-level verification of the effectivity of the novel methodology for estimating the suitability of hydrogel materials as carriers for the immobilization of biocatalysts. This comparative studies combine the virtual analysis of possible interactions that could be created in the given enzyme–carrier system, with the experimental results obtained after real immobilization of active protein.
2. Results
In the study, the invertase from Saccharomyces cerevisiae (EC 3.2.1.26, soluble type) was subjected to immobilization by entrapment in two different hydrogel matrices of natural origin—alginate and gelatin.
Initially, according to the currently developing trend of material design research, the structural models of the tested hydrogel matrices were taken under consideration. In this case, their predisposition as carriers for the immobilization of enzymes was analyzed. Next, the possibility of creating stable bonds between the model enzyme molecules and polymer chains of the hydrogel carriers was investigated.
Then, in order to select the appropriate conditions of the immobilization process, factors such as concentration of the primary cross-linking agent as well as concentration and time of the additional step of cross-linking with glutaraldehyde were examined. Verification of the suitability of each of the aforementioned factors was determined based on the correlation between received activity of hydrogel-bound invertase, immobilization yield, and the degree of enzyme leakage from the carrier. After initial selection of immobilization conditions, the invertase preparations entrapped in the alginate and gelatin hydrogels were tested in terms of operational stability.
In the following research step, the immobilized biocatalytic preparations were subjected to comparative characteristic studies. The effect of different pH and temperature on the activity and stability of invertase trapped in alginate and gelatin-based hydrogels was determined. Then, the storage stability of these preparations was tested as well. All outcomes obtained for the immobilized invertase were compared to the data obtained for the native form of this enzyme.
2.1. Molecular Insight into Structural Characteristics of Alginate and Gelatin-Based Materials
2.1.1. Analysis of Molecular Models of Alginate and Gelatin-Based Hydrogels
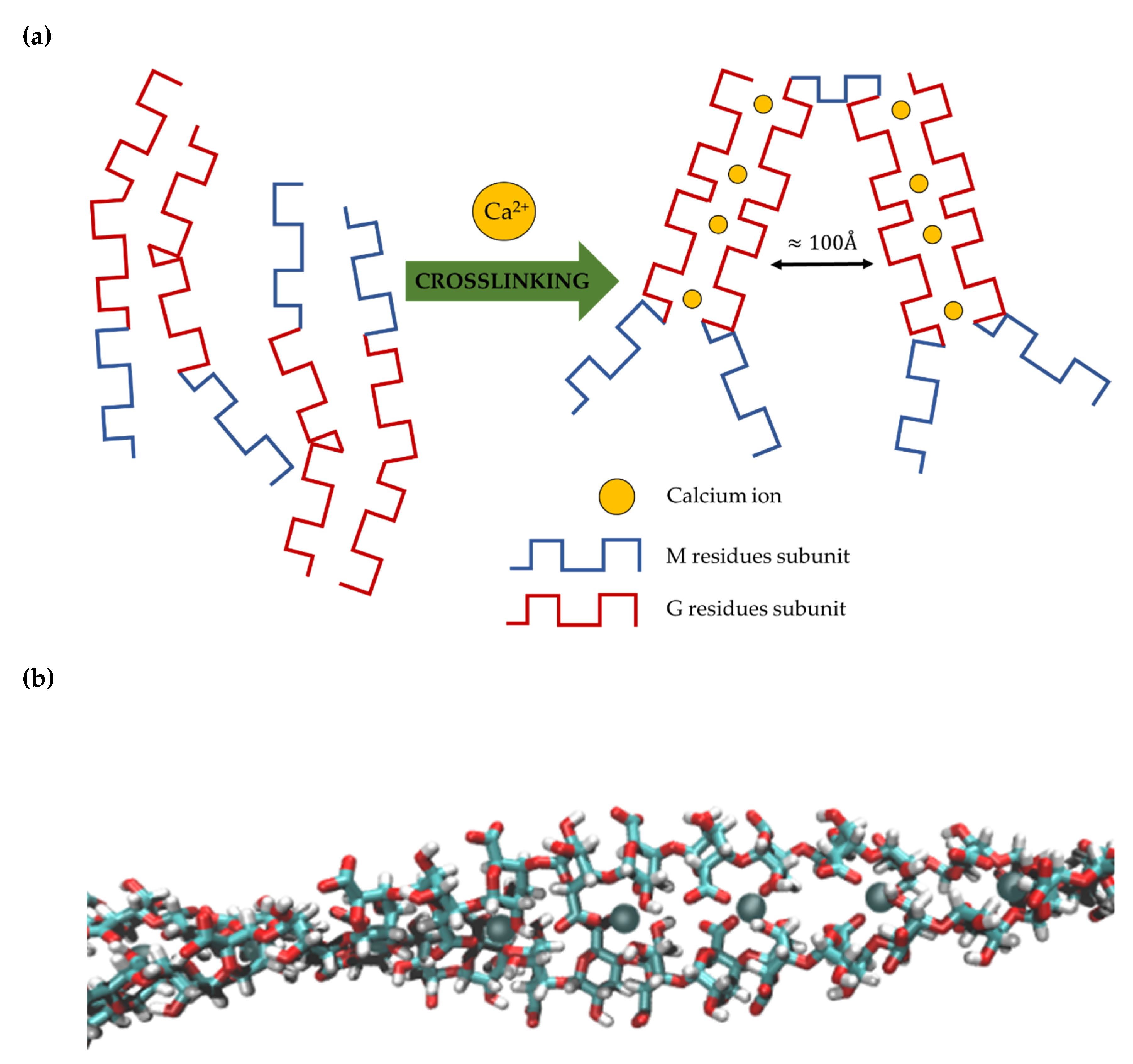
Chemically, alginates are copolymers formed of β-(1→ 4)-linked D-mannuronic acid (M) and α-(1→ 4)-linked L-guluronic acid (G) residues. The M and G units are epimers (D-mannuronic acid residues are enzymatically transformed to L-guluronic after polymerization), differing only at C5 having many different conformations. D-mannuronic acid being 4C1 with di-equatorial links between them and L-guluronic acid being 1C4 with diaxial links between them. Alginates are physically cross-linked by Ca2+ ions, forming coordinated bond around carboxyl groups binding only G residues in an egg-box-like conformation (Figure 1).
The physical cross-linking mediated by Ca2+ ions keeps the adjacent strands together. Relatively weak electrostatic couplings between distant strands provide wide voids that can be penetrated by macromolecules. Thus, alginate gels are widely used as immobilization matrices.
On the other hand, gelatin is a heterogeneous mixture of 19 polypeptides in semi random sequences. Its amino acid composition is very rich in glycine, proline, alanine, and hydroproline (Table 1).
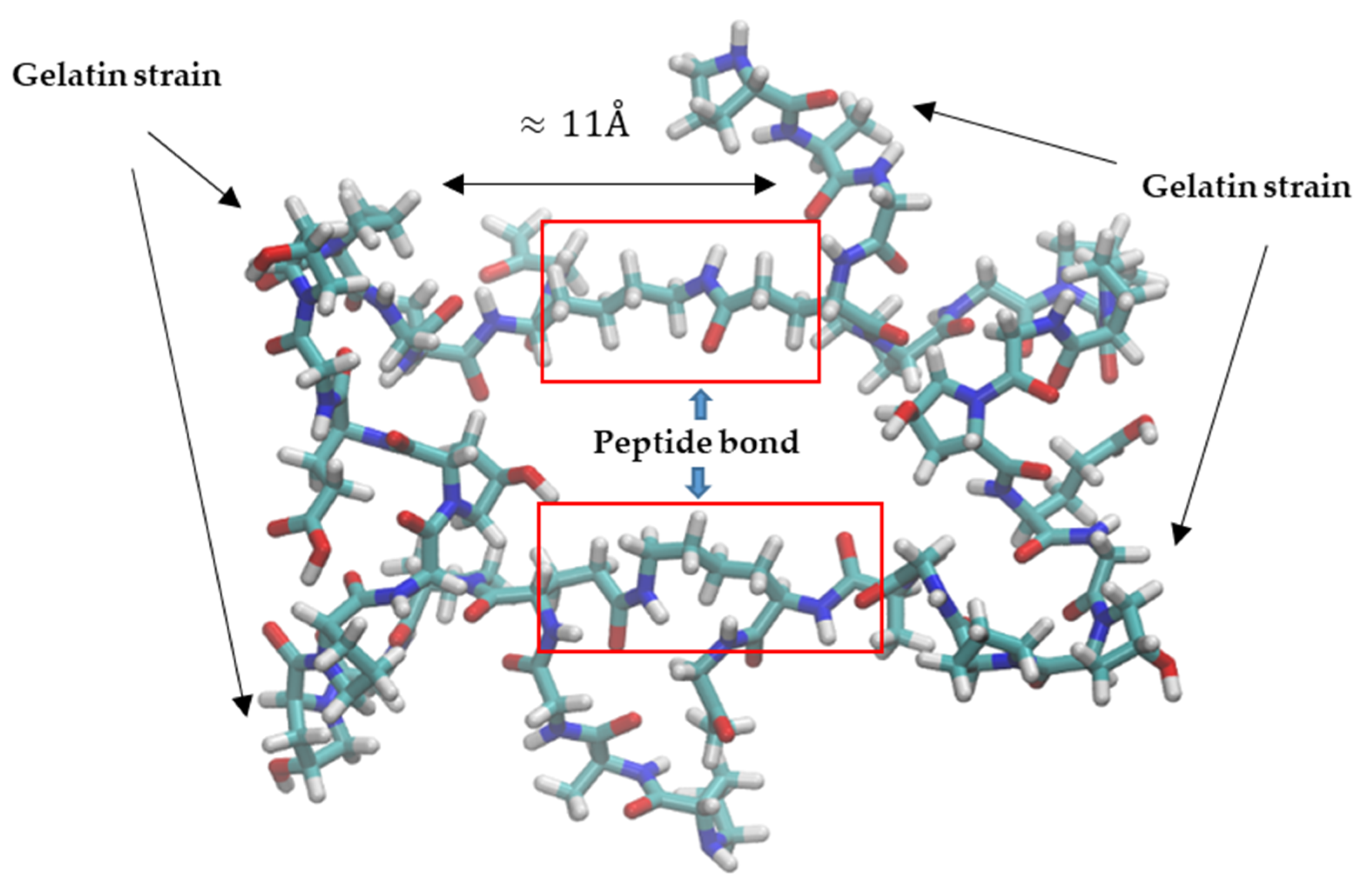
One of the typical sequences is Gly-X-Y, where Gly is glycine and X and Y are any other amino acid (Figure 2). The gelatin strands contain between 50 and 1000 amino acids and form a left-handed helical structure [32].
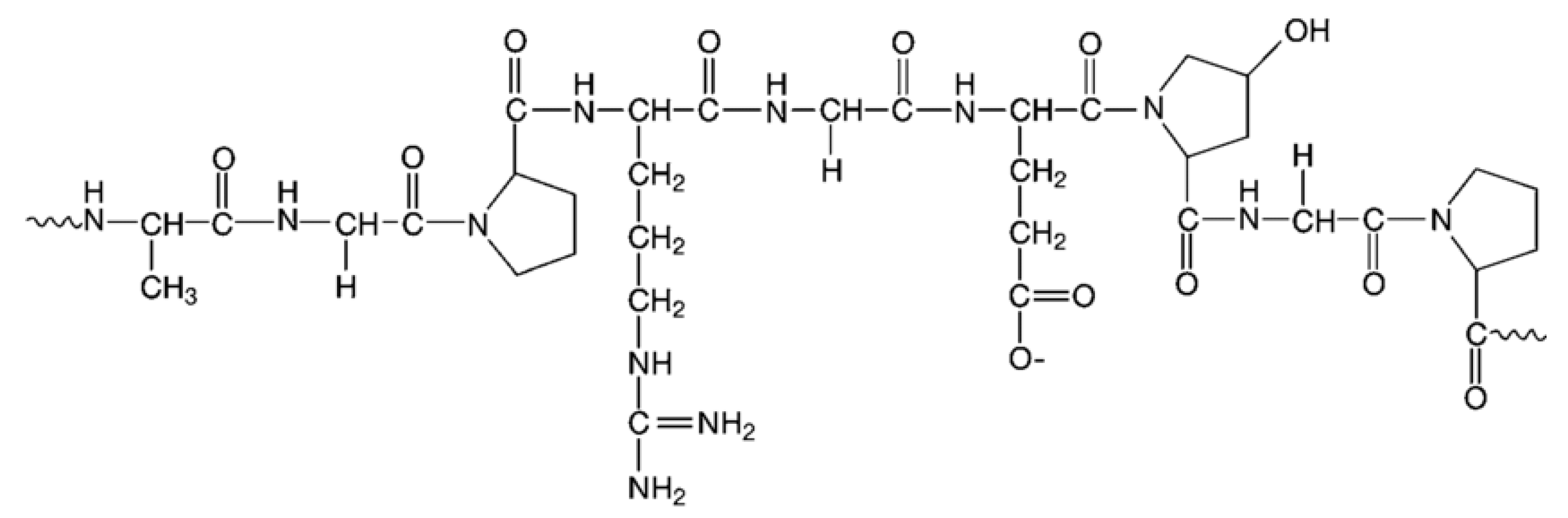
The gelatin polypeptide chains may be chemically cross-linked by microbial transglutaminase (mTGase)—the enzyme that catalyzes formation of the peptide bond between lysine and glutamine residues, as indicated in Figure 3. The pig skin gelatin contains 2.7% of Lys and 2.5% of Gln units (Table 1), which are reactive for mTGase mediated cross-linking.
The peptide bond has a length of approximately 11 A, thus limiting the pore distribution in the gelatin structure compared with that obtained in the case of calcium alginate. Owing to the chemical nature of the interactions between the polymer chains, the strength of peptide bonds is much greater than the coordination linkages created in alginate.
2.1.2. Analysis of Molecular Model of Invertase from Saccharomyces cerevisiae
Yeast invertase (EC 3.2.1.26) is one of the classical model enzymes commonly used in different research fields. Its popularity derived from the widespread utilization of the Saccharomyces strains in early biochemical studies [56]. Owing to the sequence similarity, invertases are classified as belonging to the glycoside hydrolases (family 32, GH32) [57]. The three-dimentional structure of this enzyme is depicted in Figure 4. According to the structural analysis performed by Sainz-Polo et al. [53], invertase from Saccharomyces cerevisiae is a flat square-shaped octamer (130 × 130 × 110 Å). Each polypeptide chain consists of 512 amino acid residues (Table 2).
On the basis of the invertase molecular model (Figure 4), we calculated that, out of 200 Lys and 144 Gln residues, the 148 and 64 are accessible on the surface of the protein molecule (Figure 5).
Using molecular model of the invertase, we estimated that the percentage of accessible Lys and Gln groups is 74% and 44%, respectively. Therefore, there is a high probability that these reactive amino acid moieties exposed in the enzyme molecule could be involved in the formation of additional covalent bonds with gelatin chains. This could lead to obtaining the enzyme immobilized by mixed method of physical entrapment and covalent binding, which will present significantly increased stability.
Summing up, owing to the structural features of model enzyme, both alginate and gelatin hydrogels seem to be suitable matrices for its immobilization. However, on the basis of the performed analysis of molecular predisposition of tested matrices, it could be predict that the retention of invertase molecules should be more efficient using gelatin. Furthermore, the high selectivity of mTGase cross-linking mechanism of connecting Lys and Gln residues with covalent linkages is the main reason for the improved mechanical strength of the received structures, in contrast to rather weak Ca2+–alginate materials. On this basis, we have assumed that gelatin will create a stable scaffold around the invertase, thus providing its stability.
2.2. Experimental Studies on the Process of Invertase Immobilization
The selection of appropriate immobilization conditions is the crucial step in the production of stable enzyme preparations. All experiments were performed for the invertase solution at concentration selected in the preliminary research (55.6 ± 9.93 µg/mL). The catalytic activity of the native enzyme used in the research was 19,550 ± 464 U/mL.
Each step of the selection of immobilization conditions was performed in parallel for both types of hydrogel matrices. Nevertheless, in order to maintain the transparency of the research results, they were discussed separately for each carrier.
2.2.1. Ca2+–Alginate-Based Hydrogel
Initially, the effect of CaCl2 concentration used as a cross-linking agent on the yield of invertase immobilization in alginate carrier was examined. The results of the activity of hydrogel-bound invertase, the immobilization efficiency, and the degree of enzyme leakage from the alginate matrix are summarized in Table 3.
Using data in Table 3, we can conclude that an increase in CaCl2 concentration to 15% w/v was directly proportional to the gradual increase in enzyme activity after trapping, and thereby the yield of immobilization. At the same time, the degree of enzyme leakage was successively reduced from over 20% to 10%. The subsequent increase in CaCl2 concentration did not result in additional reduction of invertase leaking out. Furthermore, the use of a 30% solution of this cross-linking agent caused a significant inactivation of the enzyme revealed in a drastic reduction of catalytic activity (Table 3). Considering these observations, the CaCl2 concentration was set up to 15% w/v and it was used for further research steps.
In the next experiment, the influence of additional cross-linking with glutaraldehyde (GA) on the effectivity of invertase immobilization was examined. Firstly, different concentrations of this agent were taken into consideration under a constant cross-linking time (20 min). Similarly to the primary cross-linking step, enzyme activity after retention in the hydrogel matrix, the immobilization yield, and the degree of leakage were also analyzed in this case (Table 4). It was observed that the use of all glutaraldehyde solutions caused a significant drop in the activity of immobilized invertase in the range from 25% to over 40%. Nevertheless, in this case, it was more important that the use of additional cross-linking with this bifunctional compound results in a considerable reduction of invertase leakage from the alginate network of about 23% to 35%. Following the analysis of the results obtained (Table 4), it was concluded that the use of a 2% v/v concentration of glutaraldehyde was the most preferable.
Afterwards, the optimal cross-linking time was determined using the selected concentration of glutaraldehyde (2% v/v). The results obtained (Table 5) were analyzed based on the correlation between received activity of hydrogel-bound invertase, immobilization yield, and the degree of enzyme leakage from the carrier.
It was observed that the activity of immobilized enzyme decreases with the increase of cross-linking time. The relative activity of the immobilized enzyme for different incubation periods was from 72% to 84.5% of the activity received for the preparation immobilized without cross-linking with glutaraldehyde. However, this treatment lasting for 30 min or longer enabled on the significant reduction of the enzyme leakage from alginate-based carrier.
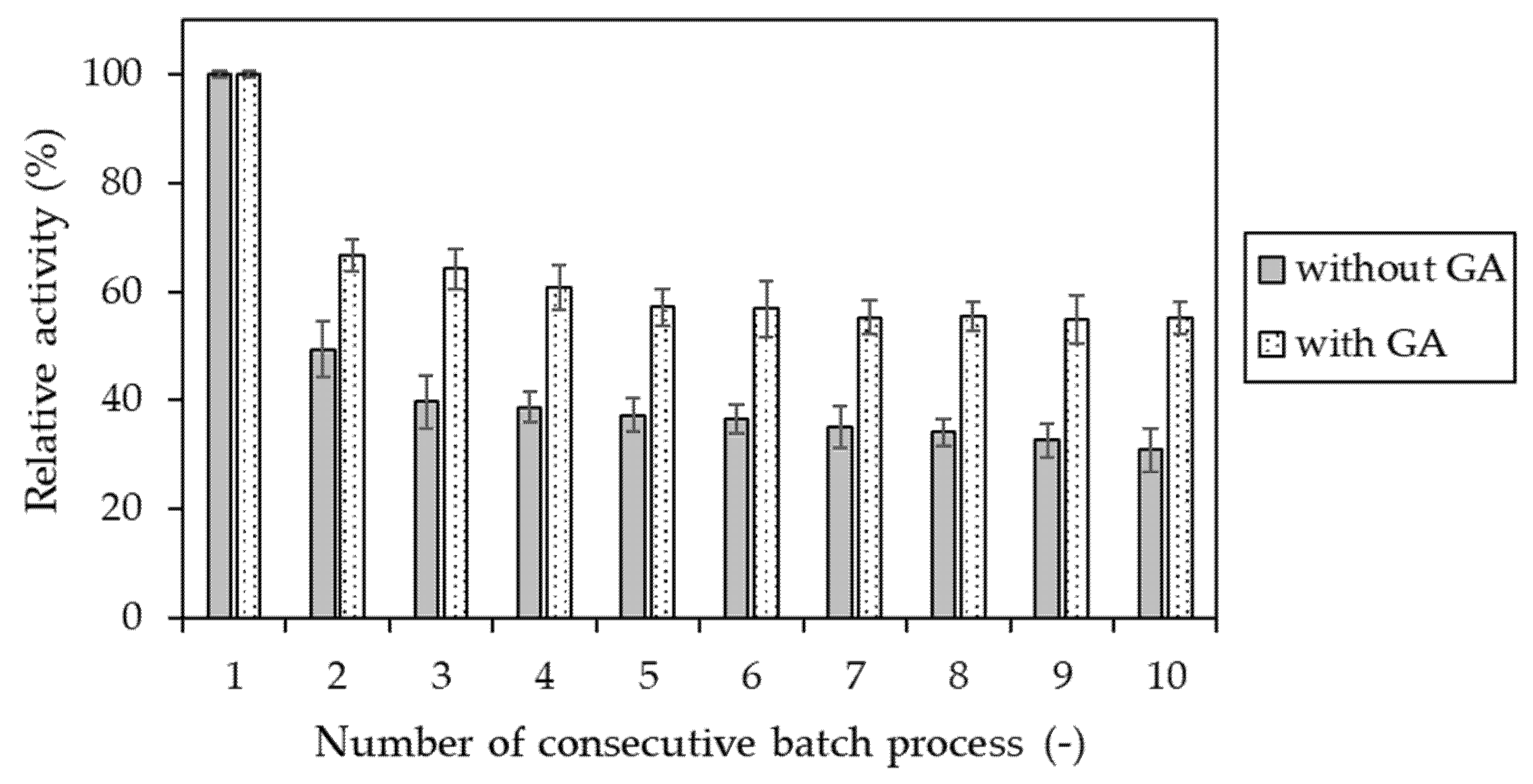
In order to confirm the usefulness of additional cross-linking with glutaraldehyde on the improvement of invertase immobilization effectivity, operational stability studies were performed for two types of alginate-entrapped enzyme—obtained without and with the presence of GA. For this purpose, ten consecutive batch processes were conducted. The obtained experimental results are depicted in Figure 6.
There is no doubt that using an additional step of cross-linking with glutaraldehyde is crucial to obtain the preparation of invertase immobilized in the alginate-based carrier with significantly increased operational stability. It was observed that, after the 10th reaction cycle, the GA cross-linked preparation of enzyme retained about 55% of its initial activity. Meanwhile, the entrapped enzyme formed only 31% without this immobilization step.
2.2.2. Gelatin-Based Hydrogel
The experiments associated with the selection of the immobilization conditions in gelatin-based hydrogel were carried out analogously to those performed for alginate carriers.
Briefly, the impact of the concentration of primary cross-linking agent on the effectivity of invertase immobilization was determined. In this case, different solutions of microbial transglutaminase (mTGase) were taken under consideration. On the basis of the obtained results (Table 6), it could be observed that the enzyme leakage could be completely eliminated by using the mTGase for cross-linking in the concentration of 1.0% w/v or higher. Moreover, the hydrogel-bound invertase obtained using ≥1.0% solution of microbial transglutaminase retained the highest activity after trapping.
Glutaraldehyde is a well-known cross-linking agent that is responsible for stiffening the enzyme structure and increasing its binding to the carrier [58]. A side effect of this can be a remarkable decrease in the catalytic activity of the enzyme after immobilization. Nevertheless, from the practical point of view, it is much more important than biocatalyst preparations additionally cross-linked with glutaraldehyde exhibit significantly increased stability under processing conditions.
In our study, although the selection of the appropriate concentration of the basic cross-linking agent (mTGase) enabled the complete retention of the invertase in the gelatin hydrogel, the effect of different concentrations of glutaraldehyde and different exposure times was examined in order to improve the enzyme stability. After analysis of the obtained results (Table 7), 0.5% v/v glutaraldehyde solution was selected for further studies, for which the appropriate incubation time was selected in the following experiment (Table 8).
Taking into account the yield of immobilization and relative activity of immobilized invertase, the cross-linking time of 10 min was found to be most suitable.
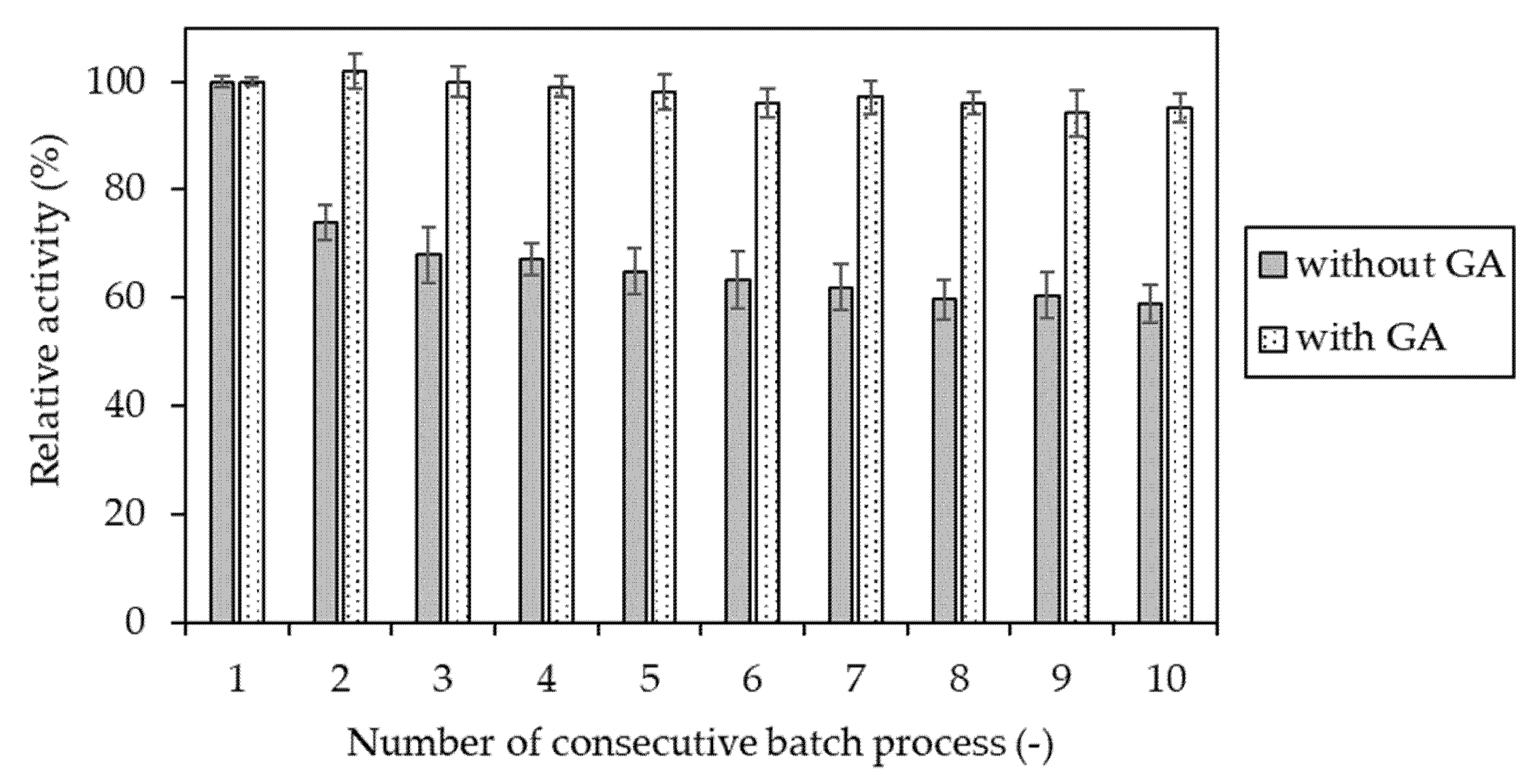
The final verification of the suitability of cross-linking with glutaraldehyde for immobilization of invertase in the gelatin-based hydrogel was checked by determination of the operational stability and comparison with the results obtained for enzyme preparation without cross-linking with GA. In this case, it was observed (Figure 7) that invertase additionaly cross-linked with glutaraldehyde retained almost 100% of the initial activity after ten consecutive reaction cycles. Meanwhile, at the same time, the enzyme preparation without GA treatment retained only about 60%.
Summing up, it was found that the cross-linking using glutaraldehyde was not an obligatory step needed to retain invertase in the gelatin-based carrier. Nevertheless, it was an indispensable procedure for improving the operational stability of the biocatalyst preparation after immobilization.
2.3. Characteristics of Invertase Immobilized in Ca2+–Alginate and Gelatin-Based Hydrogels
Alginate and gelatin-bound invertase preparations, received according to the procedures developed beforehand (Section 2.2), were characterized and compared with the properties of native form of this enzyme. For that purpose, the effect of various pH and temperature values on the activity and stability of the tested invertase forms were determined. Finally, storage stability was examined as well.
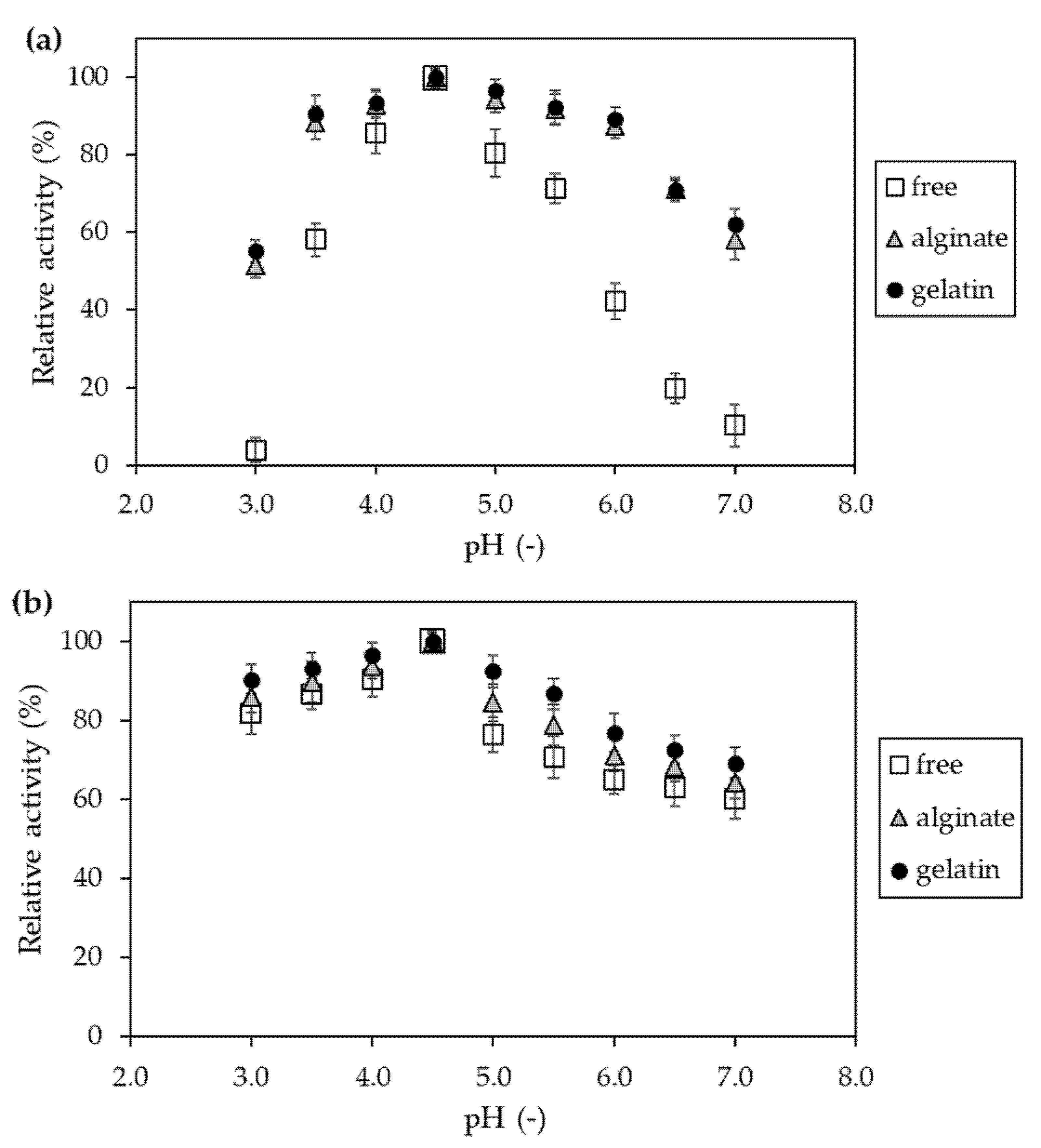
First, the impact of different pH on the retention of the catalytic activity and stability improvement was evaluated (Figure 8). It was observed that, independently of the hydrogel, the invertase after immobilization was characterized by a wider operating range of pH.
This was particularly evident in the case of the effect of pH on activity (Figure 8a), where the values obtained for the immobilized preparations were more than 20% to 50% higher in the range of pH 5.5–7.0 than for the native enzyme. Meanwhile, in the case of pH stability (Figure 8b), it reached over 60% in the whole tested range for both native and hydrogel-bound invertase preparations. However, it should be noted that the highest values were always obtained for the enzyme entrapped in the gelatin carrier.
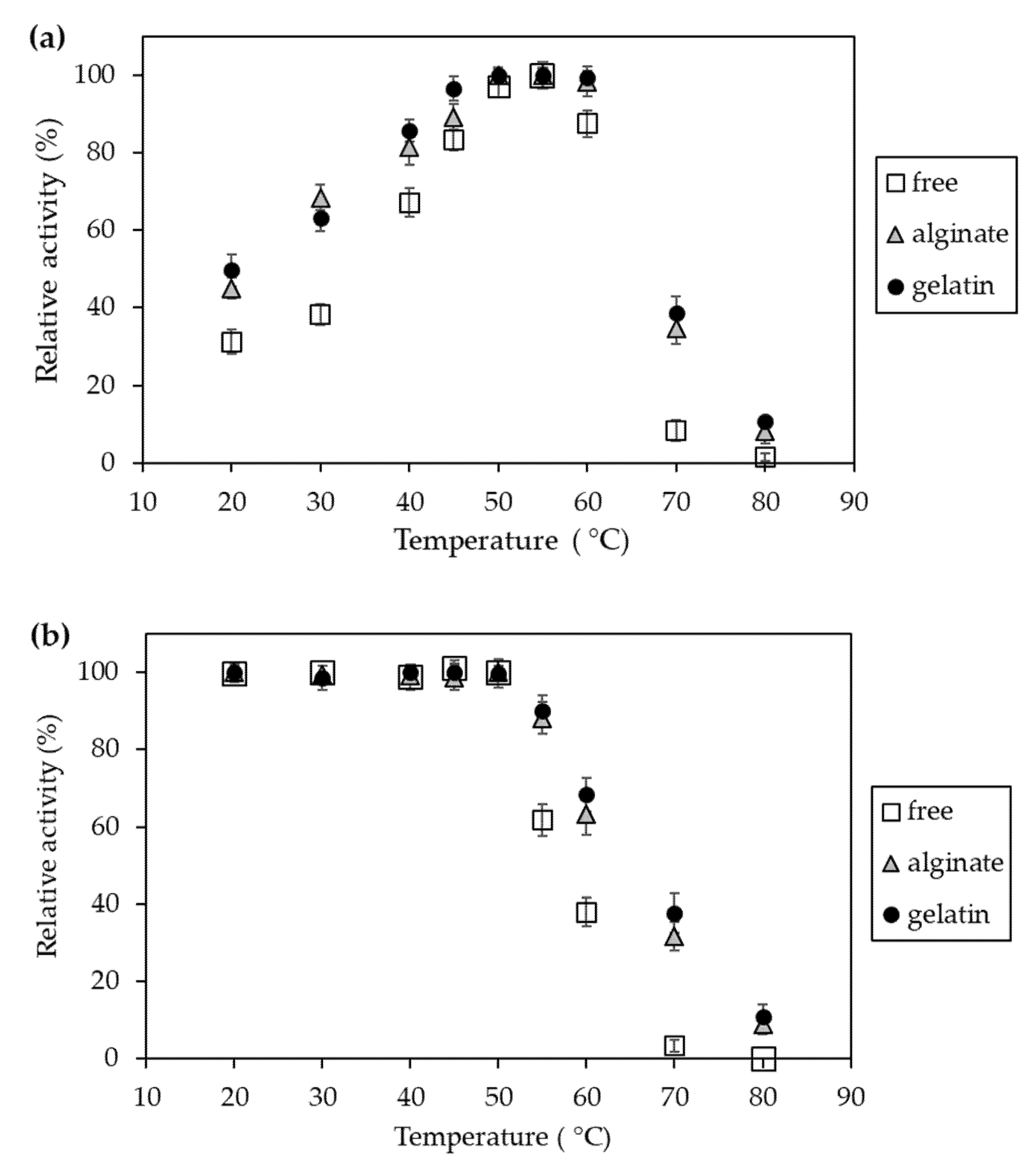
Considering the effect of temperature, in this case, immobilized invertase preparations were also characterized by higher activity (Figure 9a) and stability (Figure 9b), particularly in the increased temperature range of 60–80 °C. Moreover, here, slightly higher values were obtained for gelatin-bound enzymes.
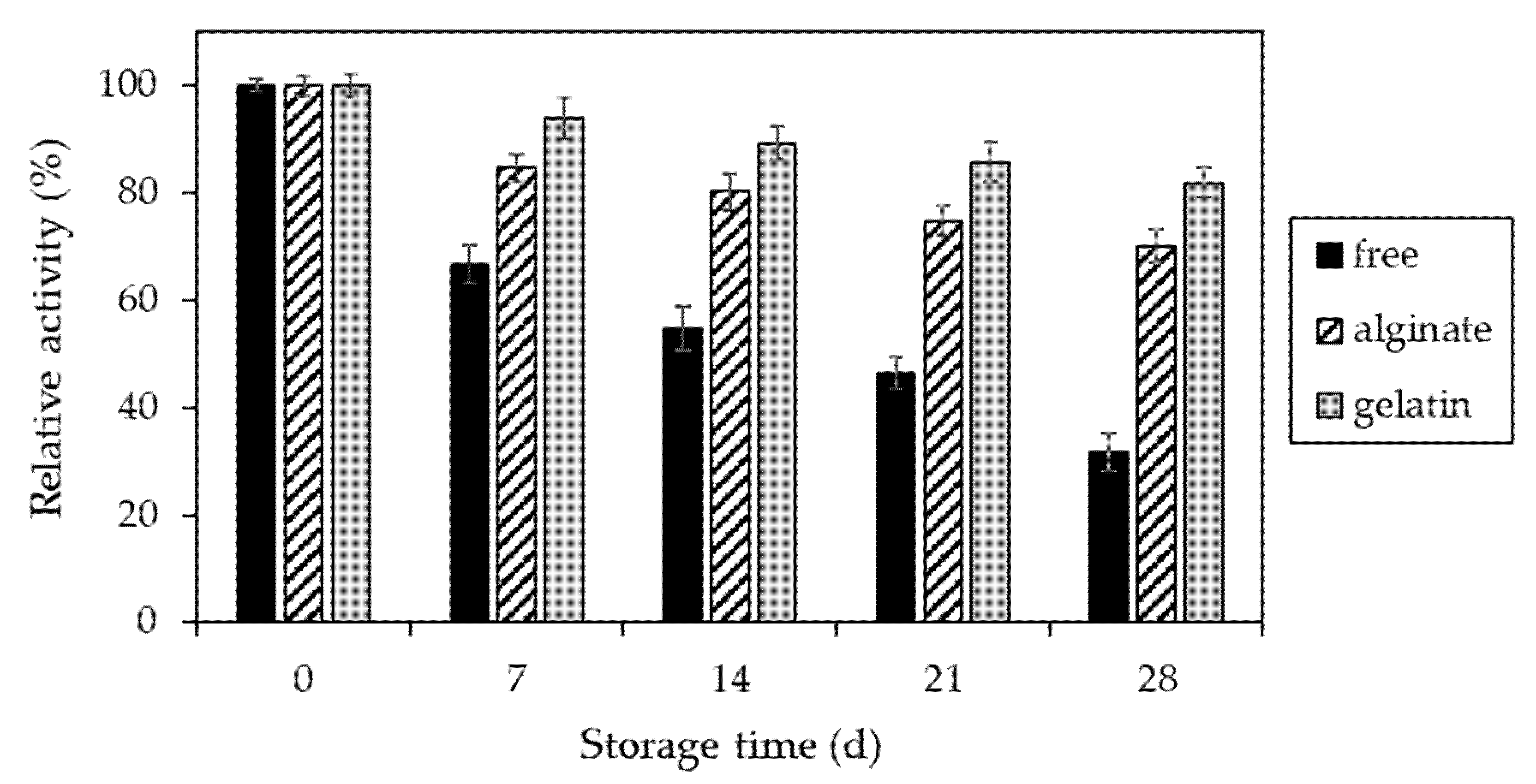
The last parameter studied in our work was the long-term stability of invertase preparations under storage conditions. In this case, the positive effect of immobilization was clearly remarkable and allowed to obtain enzyme preparations with significantly enhanced stability (Figure 10).
After 28 days of storage, the alginate and gelatin-bound invertase retained 70% and 82% of the initial activity, respectively. In contrast, the native enzyme in the same conditions retained only 32%.
3. Discussion
The use of enzymes as biocatalysts for various manufacturing processes of industrial importance is an issue that has been subjected to intensive research and dynamic development for decades. Moreover, at the moment, great importance is attached to sustainable development, the use of environmentally friendly methods, reduction of the waste generation, or their complete elimination from industrial processes. Therefore, increased attention is paid to the use of pro-ecological materials as well as methods in the production of immobilized enzyme preparations with practical application on a commercial scale [59,60]. Referring to this trend, the use of biodegradable carriers of natural origin (such as hydrogels) [20,21,61] in combination with entrapment as the immobilization method [8,31] perfectly fit into the currently considered biotechnological research directions. In this respect, the increasing popularity of using the computational tools for structural studies of various polymeric materials and molecular modeling of possible interactions arising between the reactive groups of carrier and enzyme molecule could be observed.
In our work, two natural hydrogel materials differing in chemical composition and cross-linking methods were used for comparison. For that purpose, the possibility of predicting the behavior of the enzyme (invertase) in a given hydrogel material based on structural models was initially tested. Then, the validity of the assumptions made in these theoretical design studies was tested experimentally by determining the properties of the biocatalyst after its immobilization in the considered carriers.
Molecular model analysis was an important guideline for these investigations. Although there are numerous advanced methods of atomistic calculations like molecular dynamics or Monte Carlo, simple look up of the molecular structures in question gives valuable insight before experimental research. Knowing the mechanism of gelatin chemical cross-linking and the number of Gln and Lys residues on the surface of the model enzyme, it was easy to predict that gelatin will be a much better scaffold for immobilization of that alginate.
There is no doubt that the experimental research presented in our work follows the typical pathway of the procedure used in the selection of an appropriate method for enzyme immobilization. The performed study resulted in the selection of the most favourable conditions for immobilization of soluble type invertase from Saccharomyces cerevisiae using the alginate and gelatin-based hydrogel matrices. Nevertheless, the added value of our research was the possibility of direct comparison of the results obtained in parallel for these two different carriers owing to the use of the same invertase preparation of equal concentration. Because, by using an enzyme of the same origin, purity, and catalytic activity, it is possible to eliminate erroneous interpretation of immobilization effectivity, that could result from differences in biocatalytic preparations used. Furthermore, this enables direct analysis of the enzyme preferences to the carrier or the immobilization method.
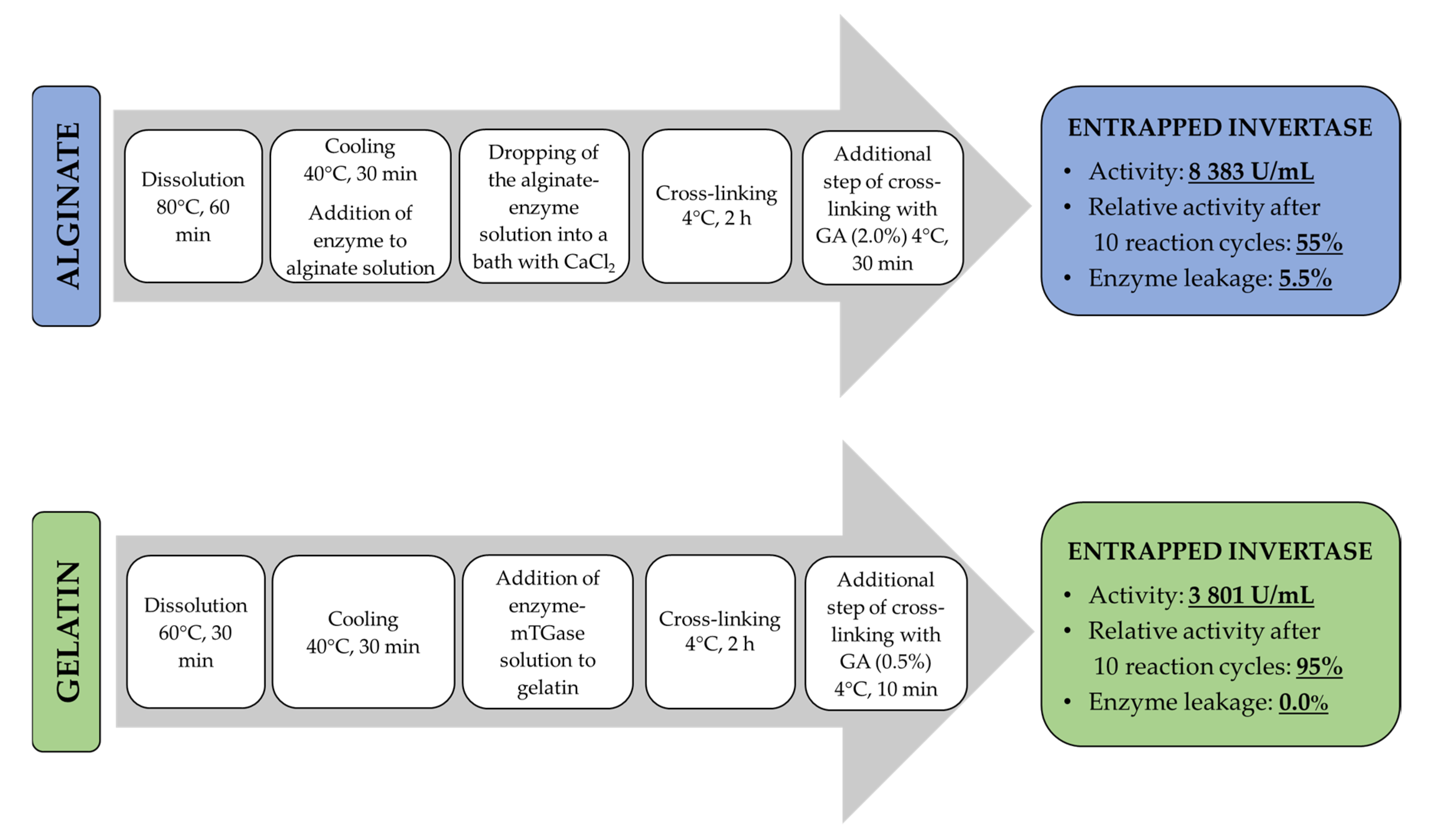
The comparison of both entrapment procedures developed in the current study is depicted in Figure 11.
In the case of alginate-bound invertase, even the introduction of an additional cross-linking step using a fairly high concentration of glutaraldehyde and quite a long exposure time did not result in complete entrapment of the enzyme into the carrier. Moreover, a further increase of GA concentration caused a severe drop in the activity retained by invertase. Therefore, the use of this procedure was not economically justified. In turn, using the gelatin matrix, the complete invertase retention was obtained prior to application of an additional step of cross-linking with glutaraldehyde. Nevertheless, this procedure performed in the presence of a low GA concentration (0.5%) and a short incubation time (10 min) enabled a significant increase in the operational stability of entrapped enzyme preparation (Figure 7). A comparative analysis of the results obtained allowed the unequivocal selection of gelatin-based hydrogel as an effective carrier for immobilization of invertase. This hydrogel matrix enables one to obtain entrapped enzyme preparation characterized by improved operational (Figure 7) and storage (Figure 10) stability, as well as grater resistance in the pH range of 3.0–7.0 (Figure 8) and at elevated temperatures (Figure 9) compared with invertase bound in the alginate beads. Moreover, the use of gelatin as a matrix for immobilization ensures complete retention of invertase in the hydrogel network (Table 7), which is extremely important from a processing side.
The obtained experimental results not only confirm our molecular analysis of the suitability of both materials for the immobilization of invertase, but also fit quite well with the outcomes reported in the literature [62,63,64,65]. Furthermore, the gelatin-entrapped invertase developed in our study successfully competes with similar enzyme preparations received by other research groups (Table 9).
It was observed that the gelatin-bound invertase received in our research was characterized by the highest operational stability and one of the most improved stabilities at the elevated temperatures from all of the preparations taken under consideration.
4. Materials and Methods
4.1. Materials
Porcine skin gelatin, alginic acid sodium salt, invertase from baker’s yeast (S. cerevisiae) (EC 3.2.1.26), glutaraldehyde (GA), calcium chloride, and sucrose were purchased from Sigma-Aldrich (St. Louis, MO, USA). The analytical glucose test was from Biomaxima (Lublin, Poland). Microbial transglutaminase Activa® WM was kindly donated by Ajinomoto (Tokyo, Japan). Other reagents, all of analytical grade, were supplied by Avantor Performance Materials (Gliwice, Poland). For molecular analysis, we used LAMMPS (ver. December 2018, distributed as an open source code under the terms of the GPL by lammps.sandia.gov) molecular package to together with AMBER forcefield and Avogadro 2.0 (ver. December 2018, LAMMPS is distributed as an open source code under the terms of the GPL by lammps.sandia.gov) and VMD 1.9.2 (free license provided by Theoretical and Computational Biophysics Group of University of Illinois https://www.ks.uiuc.edu/) for visualization.
4.2. Methods
4.2.1. Preparation of Molecular Models of Hydrogel Structures
The molecular model of alginate was prepared using polymatic tool run under nanoHub (nonohub.org) and then minimized using LAMMPS software with AMBER forcefield. The methodology of creating initial model of gelatin has been described in [32]. The gelatin model was then reduced and modified by polypeptide bong using Avogadro 2.0 software. All of the structures were visualized using VMD 1.9.2. software.
4.2.2. Preparation of Virtual Model of Invertase
The molecular model of invertase was downloaded from the PDB database (rcsb.org) and then visualized using VMD 1.9.2 software. To present our reasoning, we used the model depicted in the PDB database as 4EQV.
4.2.3. Preparation of Hydrogel Matrices Containing Immobilized Invertase
Immobilization of invertase in alginate-based hydrogel was performed as follows. Firstly, the solution of calcium chloride (cross-linking bath) with an appropriate concentration was prepared in 0.05 M acetate buffer at pH 4.5 and placed at 4 °C for about 2–3 h. Next, a weighed portion of sodium alginate (2.25% w/v) was dissolved in 0.05 M acetate buffer at pH 4.5 in a reactor thermostated at 80 °C for approximately 2 h. After this time, the solution was cooled to 40 °C and incubated at this temperature for a few minutes. In parallel, a buffer solution of invertase with a given concentration was prepared and mixed with sodium alginate at the volume ratio of 1:2. The cross-linking step was initiated by dropping the blend of invertase/sodium alginate to the continuously stirring (250 rpm) solution of chilled calcium chloride. Then, in order to complete the cross-linking process, the moulded Ca2+-alginate particles containing immobilized invertase were stored at 4 °C for 2 h. After this time, the hydrogel beads were drained, washed twice with 0.05 M acetate buffer pH 4.5, and their geometric dimensions were measured (dAVERAGE = 3.5 ± 0.2 mm). In this case, it was also assumed that the total amount of used invertase was immobilized in the alginate-based cross-linked beads.
Immobilization of invertase in gelatin-based hydrogel was performed analogously to the method applied in our previous study for entrapping of another enzyme (β-galactosidase) [24].
However, a few modifications were used to tailor this procedure to a given enzyme. Briefly, a weighed portion of gelatin (15% w/v) was dissolved in 0.05 M acetate buffer at pH 4.5 in a reactor thermostated at 60 °C for approximately 30 min. Next, the solution was cooled to 40 °C and incubated at this temperature for a few minutes. In parallel, a buffer solution of invertase with given concentration was prepared. Then, after total dissolution of the enzyme, a weighted portion of cross-linking agent (microbial transglutaminase, mTGase) was added to obtain its concentration of 3% w/v. The cross-linking step began by mixing the appropriate amount of invertase/mTGase solution with gelatin at the volume ratio of 1:2. Afterwards, the obtained blend was immediately cooled to 4 °C and kept under these conditions for 2 h. After this time, the hydrogel particles (cylinder shape) were drained, washed twice with 0.05 M acetate buffer pH 4.5, and their geometric dimensions were measured (dAVERAGE = 8.0 ± 0.4 mm; hAVERAGE = 10 ± 0.3 mm). It was assumed that the total amount of used invertase was immobilized in the gelatin-based hydrogel lattice.
Immobilization yield was determined as the activity of bound enzyme divided to activity of native form of the enzyme, expressed in (%).
4.2.4. Analytical Test for Determination of Protein Content
The protein concentration was determined using the Lowry’s method [66] with bovine serum albumin as a standard. This method is based on a two-step procedure. In the first step, a biuret reaction occurs—the interaction between peptide bonds and Cu2+ copper ions and the reduction of Cu2+ to Cu+ ions in an alkaline environment. In the second step, the Folina–Ciocalteu reagent (phosphomolybdic acid and phosphotungstic acid) is reduced to the corresponding oxides with Cu+ and tyrosine and tryptophan residues. The intensity of the blue color of the product measured spectrophotometrically at 750 nm is directly proportional to the protein concentration in the analyzed sample.
Then, 0.5 mL of Lowry reagent was added to a 0.5 mL solution of the analyzed sample, mixed, and incubated at room temperature for 20 min. After this time, 0.25 mL of Folina–Ciocalteu reagent was added, immediately mixed, and incubated for another 30 min. Then, the absorbance at 750 nm was measured. Protein concentration was determined using the standard curve (CPROTEIN [µg/mL] = 207.9.A750) obtained for different bovine serum albumin in the range of 10–200 µg/mL.
4.2.5. Analytical Test for Determination of Glucose Concentration
Glucose concentration was determined using the commercially available analytical test (Biomaxima, Poland). This detection method is based on the conversion of glucose to the coloured product, formed as a result of two consecutive enzymatic reactions. Briefly, glucose during the first reaction catalyzed by glucose oxidase is converted to gluconic acid and, simultaneously, one molecule of hydrogen peroxide is generated. Then, in the presence of peroxidase, the hydrogen peroxide reacts with hydroxybenzoic acid (HBA) and 4-aminoantipyrine (AAP) to form a red dye—quinoneimine. The intensity of the colour obtained was measured spectrophotometrically at 500 nm. It is directly proportional to the glucose concentration in the sample.
The test was performed as follows: 10 uL of the sample withdrawn from the reaction mixture was added to 1 mL of analytical reagent and incubated for 5 min at 37 °C. Next, the absorbance at 500 nm was measured. Glucose concentration was determined using the standard curve (CGLUCOSE [µg/mL] = 2703.5·A500) obtained for different glucose concentrations in the range of 50–3000 µg/mL.
4.2.6. Determination of Catalytic Activity of Native and Immobilized Invertase
The catalytic activity of invertase was determined by monitoring the progress of hydrolysis of sucrose to glucose and fructose over time. In all experiments, the sucrose solution at a final concentration of 50 g L−1 prepared in 0.05 M acetate buffer (pH 4.5) was used as a substrate. The activity determination method was based on measuring the concentration of glucose formed as a reaction product. One unit of enzyme activity (1 U) was defined as 1 µg of glucose formed within 1 min. of the reaction of sucrose hydrolysis catalyzed by invertase.
The reaction with the native enzyme was carried out in a sealed test tube. Then, 5 mL of sucrose solution was preincubated for 10 minutes in a thermostatic water bath at 50 °C. After this time, the reaction was started by adding 0.1 mL of invertase to the substrate. Every 0.5 min, the samples were withdrawn from the reaction and the glucose concentration was determined.
The reaction using immobilized invertase was carried out in a thermostated stirred reactor (250 rpm) at 50 °C. The appropriate volume of 50 g/L sucrose solution (25 mL—gelatin hydrogels, 10 mL—alginate hydrogels) was pre-incubated for 10 min, and then the sucrose hydrolysis process was started by adding a proportional amount of the hydrogel containing the immobilized enzyme. During the 10 min of the process, the samples were taken every 1 min for determination of the glucose content.
4.2.7. Determination of Enzyme Leakage from Hydrogel Matrices
The enzyme leakage degree was examined after one day of incubation of hydrogel-bound invertase in the 5 mL of 0.05 M buffer solution (pH 4.5).
Activity of the enzyme released from hydrogel matrices was determined analogously to the procedure used for the native invertase prior to immobilization (Section 4.2.6). The reaction of sucrose hydrolysis was carried out in a sealed tube placed in the water bath thermostated at 50 °C. Briefly, 5 mL of 50 g/L substrate was pre-incubated for 10 min. In order to start the reaction, 0.1 mL of the solution from the buffer suspension of the immobilized enzyme was added to the preheated sucrose solution. The hydrolysis process was carried out for 10 min, taking samples every 2 min.
4.2.8. Influence of the Concentration of the Primary Cross-Linking Agent on the Efficiency of Invertase Immobilization in Hydrogel Matrices
The preparation of durable alginate-based hydrogels required the incubation step in cross-linking bath containing the appropriate concentration of calcium chloride (CaCl2). The following concentrations of CaCl2 were tested: 2%, 4%, 10%, 15%, 20%, and 30%. In turn, for gelatin-based matrices, the addition of a variety of different concentrations (0.25%, 0.5%, 1.0%, 2.0%, 3.0%, and 4.0%) of microbial transglutaminase (mTGase) was examined as valuable cross-linking agent.
In order to verify the effect of these primary cross-linking agents on the efficiency of enzyme immobilization, the invertase activity and its leakage degree from the hydrogel were determined according to the standard procedures described in Section 4.2.6 and Section 4.2.7, respectively.
All results included in the tables are given as average values obtained from measurements performed in triplicate.
4.2.9. Influence of Additional Step of Cross-Linking with Glutaraldehyde on the Efficiency of Invertase Immobilization in Hydrogel Matrices
Different concentrations of glutaraldehyde (0.5%, 1.0%, 2.0%, and 4.0% v/v) were taken under consideration in order to determine the effect of an additional step of cross-linking on the efficiency of invertase immobilization. For that purpose, the preparations of hydrogel-bound enzyme were incubated in glutaraldehyde solutions for 20 minutes. After this time, the hydrogel beads were drained and washed twice with 0.05 M acetate buffer pH 4.5 prior to activity measurements (Section 4.2.6). Moreover, the effectiveness of this procedure was verified by determination of an invertase leakage degree (4.2.7). The concentration of glutaraldehyde, which enabled to receive the immobilized enzyme preparation with most favorable ratio of activity to leakage degree, was chosen for further research.
Subsequently, the influence of cross-linking time with glutaraldehyde on the efficiency of invertase immobilization was examined. In this case, hydrogels with bound enzyme were incubated in the GA solution for different time periods (0, 5, 10, 20, 30, 45, and 60 min). Then, the hydrogel beads were drained and washed twice with 0.05 M acetate buffer pH 4.5 prior to activity (Section 4.2.6) and leakage (Section 4.2.7) measurements.
All results included in tables are given as average values obtained from measurements performed in triplicate.
4.2.10. Determination of Operational Stability of Invertase Immobilized in Hydrogel Matrices
Operational stability of immobilized invertase was determined in consecutive batch processes of sucrose hydrolysis (reaction time 30 min). For that purpose, the enzyme-hydrogel preparations obtained using procedure without and with additional step of cross-linking with glutaraldehyde were examined. Ten subsequent reactions were performed for each sample of entrapped invertase. After the end of one reaction, the hydrogel-bound enzyme was washed twice with the 0.05 M acetated buffer pH 4.5 and the next batch process was started with the same invertase sample. The reaction rate received in the first run was set as 100%.
4.2.11. Determination of the Effect of pH on Activity and Stability of Invertase Immobilized in Hydrogel Matrices
The optimum pH for invertase in native and immobilized form was determined in the pH range of 3.0–7.0 by measuring the enzyme activity in the presence of sucrose solutions adjusted to a certain pH. The highest value of invertase activity obtained during the experiment was assumed as 100%.
The stability of invertase preparations was tested in the range of pH 3.0–7.0. Incubation of the enzyme at a given pH was carried out for 1 h. Then, the solution was adjusted to pH 4.5 and left at room temperature for another hour. After this time, the invertase activity was determined in standard conditions (Section 4.2.6). The highest value of enzyme activity obtained during the study was assumed as 100%.
4.2.12. Determination of the Effect of Temperature on Activity and Stability of Invertase Immobilized in Hydrogel Matrices
The optimum temperature for invertase in native and immobilized form was determined by measuring the enzyme activity in the presence of sucrose solutions brought to the desired temperature from the range of 20–80 °C. The highest value of invertase activity obtained during the experiment was assumed as 100%.
The stability of invertase preparations was tested in the temperature range of 20–80 °C. Incubation of the enzyme at a given temperature was carried out for 1 h. Then, the solution was adjusted to room temperature and left for another hour. After this time, the invertase activity was determined in standard conditions (Section 4.2.6). The highest value of invertase activity obtained during the study was assumed as 100%.
4.2.13. Determination of Storage Stability of Invertase Immobilized in Hydrogel Matrices
Storage stability was tested by keeping preparation in the buffer at 4 °C for 7, 14, 21, and 28 days. The enzyme activity measured before storage (day 0) was set as 100%.
5. Conclusions
In the current work, we analyzed the predisposition of two natural hydrogels (alginate and gelatin) as efficient matrices for immobilization of yeast invertase. During the study, the experimental immobilization procedures were preceded by the analysis of structural models of these materials and the enzyme. The outcomes from practical research confirmed our assumptions made in preliminary theoretical considerations. As the final result, it was found that a gelatin-based hydrogel matrix is the most preferred choice for invertase immobilization.
In the paper, we also indicate the importance of molecular analysis in guiding the researcher towards the right conclusions in the experimental. Although studying in detail the complex systems like enzyme and biopolymer in solution is possible using methods like molecular dynamics, it is still a state-of-the-art calculation requiring large computation power, specialized software, and a relatively long computational time. Such an approach is being presented more and more widely in this field of study. However, the interplay between molecular models and experimental setup may be accessible also in much simpler version, as presented in the paper. Currently, the researcher does not need to be an expert in computations to have access to molecular models of complex molecular systems using widely available databases of molecular systems such as, for example, PDB.org, and draw conclusions upon them. Three-dimensional molecular models provide valuable insight into possible mechanism of the mutual interaction in the enzyme-carrier system, and hence a selection of specific conditions prior to performing only strictly targeted experimental research.
In this work using aforementioned analysis on the molecular level, we pointed out the reason gelatin will be a more suitable choice for invertase immobilization. Namely, we indicated that because there are a number accessible Lys and Gln groups on the surface of the enzyme, there is a large probability of gelatin being chemically cross-linked with the enzyme. The experimental findings also justify our theoretical assumptions.
However, detailed molecular modeling analysis is also possible and needed in the research of enzyme immobilization. In the future perspective, the molecular dynamics and coarse-grained studies can provide additional information on the diffusion rate of product and substrate within the hydrogel matrix, how mechanical properties of hydrogel matrix changes upon presence of the enzyme, the swelling properties of the hydrogel, and the detailed nature of enzyme–hydrogel interactions.
Author Contributions
Conceptualization, K.L. and Ł.R.; methodology, K.L. and Ł.R.; software, Ł.R.; validation, K.L. and Ł.R.; formal analysis, K.L.; investigation, K.L., K.W., and Ł.R.; resources, K.L.; data curation, K.L. and Ł.R.; writing—original draft preparation, K.L.; writing—review and editing, K.L. and Ł.R.; visualization, K.L. and Ł.R.; project administration, K.L and Ł.R.; funding acquisition, K.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by NATIONAL SCIENCE CENTRE POLAND grant number 2015/19/D/ST8/01899.
Acknowledgments
Authors would like to thank Ajinomoto Company (Tokyo, Japan) for the kind donation of transglutaminase preparation (trade name ACTIVA®WM).
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyzes, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Kirk, O.; Borchert, T.V.; Fuglsang, C.C. Industrial enzyme applications. Curr. Opin. Biotechnol. 2002, 13, 345–351. [Google Scholar] [CrossRef]
- Choi, J.M.; Han, S.S.; Kim, H.S. Industrial applications of enzyme biocatalysis: Current status and future aspects. Biotechnol. Adv. 2015, 33, 1443–1454. [Google Scholar] [CrossRef] [PubMed]
- Basso, A.; Serban, S. Industrial applications of immobilized enzymes—A review. Mol. Catal. 2019, 479, 110607. [Google Scholar] [CrossRef]
- Chapman, J.; Ismail, A.; Dinu, C. Industrial Applications of Enzymes: Recent Advances, Techniques, and Outlooks. Catalysts 2018, 8, 238. [Google Scholar] [CrossRef] [Green Version]
- Miller, A.; Nordenberg, T. Food and Drug Administration. In Encyclopedia of Food Sciences and Nutrition; Caballero, B., Toldra, F., Finglas, P., Eds.; Academic Press: Cambridge, MA, USA, 2003; pp. 2593–2599. [Google Scholar]
- Singh, R.K.; Tiwari, M.K.; Singh, R.; Lee, J.K. From protein engineering to immobilization: Promising strategies for the upgrade of industrial enzymes. Int. J. Mol. Sci. 2013, 14, 1232–1277. [Google Scholar] [CrossRef]
- Bilal, M.; Zhao, Y.; Noreen, S.; Shah, S.Z.H.; Bharagava, R.N.; Iqbal, H.M.N. Modifying bio-catalytic properties of enzymes for efficient biocatalysis: A review from immobilization strategies viewpoint. Biocatal. Biotransform. 2019, 37, 159–182. [Google Scholar] [CrossRef]
- Sheldon, R.A. Enzyme immobilization: The quest for optimum performance. Adv. Synth. Catal. 2007, 349, 1289–1307. [Google Scholar] [CrossRef]
- Homaei, A.A.; Sariri, R.; Vianello, F.; Stevanato, R. Enzyme immobilization: An update. J. Chem. Biol. 2013, 6, 185–205. [Google Scholar] [CrossRef] [Green Version]
- Ó’Fágáin, C. Enzyme stabilization—Recent experimental progress. Enzyme Microb. Technol. 2003, 33, 137–149. [Google Scholar] [CrossRef]
- Liu, Q.; Xun, G.; Feng, Y. The state-of-the-art strategies of protein engineering for enzyme stabilization. Biotechnol. Adv. 2019, 37, 530–537. [Google Scholar] [CrossRef]
- Iyer, P.V.; Ananthanarayan, L. Enzyme stability and stabilization—Aqueous and non-aqueous environment. Process Biochem. 2008, 43, 1019–1032. [Google Scholar] [CrossRef]
- Guzik, U.; Hupert-Kocurek, K.; Wojcieszynska, D. Immobilization as a strategy for improving enzyme properties- Application to oxidoreductases. Molecules 2014, 19, 8995–9018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mateo, C.; Palomo, J.M.; Fernandez-Lorente, G.; Guisan, J.M.; Fernandez-Lafuente, R. Improvement of enzyme activity, stability and selectivity via immobilization techniques. Enzyme Microb. Technol. 2007, 40, 1451–1463. [Google Scholar] [CrossRef]
- Noritomi, H.; Sasanuma, A.; Kato, S.; Nagahama, K. Catalytic properties of cross-linked enzyme crystals in organic media. Biochem. Eng. J. 2007, 33, 228–231. [Google Scholar] [CrossRef]
- Voběrková, S.; Solčány, V.; Vršanská, M.; Adam, V. Immobilization of ligninolytic enzymes from white-rot fungi in cross-linked aggregates. Chemosphere 2018, 202, 694–707. [Google Scholar] [CrossRef]
- Wang, S.; Zheng, D.; Yin, L.; Wang, F. Preparation, activity and structure of cross-linked enzyme aggregates (CLEAs) with nanoparticle. Enzyme Microb. Technol. 2017, 107, 22–31. [Google Scholar] [CrossRef]
- Abraham, T.E.; Joseph, J.R.; Bindhu, L.B.V.; Jayakumar, K.K. Crosslinked enzyme crystals of glucoamylase as a potent catalyst for biotransformations. Carbohydr. Res. 2004, 339, 1099–1104. [Google Scholar] [CrossRef]
- Zdarta, J.; Meyer, A.S.; Jesionowski, T.; Pinelo, M. A general overview of support materials for enzyme immobilization: Characteristics, properties, practical utility. Catalysts 2018, 8, 92. [Google Scholar] [CrossRef] [Green Version]
- Bilal, M.; Iqbal, H.M.N. Naturally-derived biopolymers: Potential platforms for enzyme immobilization. Int. J. Biol. Macromol. 2019, 130, 462–482. [Google Scholar] [CrossRef]
- Chou, C.; Syu, S.; Chang, J.H.; Aimar, P.; Chang, Y. Bioinspired Pseudozwitterionic Hydrogels with Bioactive Enzyme Immobilization via pH-Responsive Regulation. Langmuir 2019, 35, 1909–1918. [Google Scholar] [CrossRef] [Green Version]
- Elnashar, M.M.M. The Art of Immobilization Using Biopolymers, Biomaterials and Nanobiotechnology. In Biotechnology of Biopolymers; InTech: London, UK, 2011; ISBN 978-953-307-179-4. [Google Scholar]
- Trusek-Holownia, A.; Noworyta, A. Efficient utilisation of hydrogel preparations with encapsulated enzymes—A case study on catalase and hydrogen peroxide degradation. Biotechnol. Rep. 2015, 6, 13–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labus, K.; Drozd, A.; Trusek-Holownia, A. Preparation and characterisation of gelatine hydrogels predisposed to use as matrices for effective immobilisation of biocatalystst. Chem. Pap. 2016, 70, 523–530. [Google Scholar]
- Czyzewska, K.; Trusek, A. Encapsulated catalase from Serratia genus for H2O2 decomposition in food applications. Polish J. Chem. Technol. 2018, 20, 39–43. [Google Scholar] [CrossRef] [Green Version]
- Batista, R.A.; Espitia, P.J.P.; Quintans, J.S.S.; Freitas, M.M.; Cerqueira, M.Â.; Teixeira, J.A.; Cardoso, J.C. Hydrogel as an alternative structure for food packaging systems. Carbohydr. Polym. 2019, 205, 106–116. [Google Scholar] [CrossRef] [Green Version]
- Joye, I.J. Cereal biopolymers for nano- and microtechnology: A myriad of opportunities for novel (functional) food applications. Trends Food Sci. Technol. 2019, 83, 1–11. [Google Scholar] [CrossRef]
- Kaczmarek, B.; Nadolna, K.; Owczarek, A. The physical and chemical properties of hydrogels based on natural polymers. In Hydrogels Based on Natural Polymers; Chen, Y., Ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2020; pp. 151–172. ISBN 9780128164211. [Google Scholar]
- Bao, Z.; Xian, C.; Yuan, Q.; Liu, G.; Wu, J. Natural Polymer-Based Hydrogels with Enhanced Mechanical Performances: Preparation, Structure, and Property. Adv. Healthc. Mater. 2019, 8, 1900670. [Google Scholar] [CrossRef]
- Ha, T.L.B.; Quan, T.M.; Vu, D.N.; Si, D.M. Chapter 11: Naturally Derived Biomaterials: Preparation and Application. In Regenerative Medicine and Tissue Engineering; InTech: London, UK, 2013; pp. 247–272. [Google Scholar]
- Koch-Schmidt, A. Gel-Entrapment of Enzymes. In Biomedical Applications of Immobilized Enzymes and Proteins; Chang, T.M.S., Ed.; Springer: Boston, MA, USA, 1977; pp. 47–67. [Google Scholar]
- Radosinski, L.; Labus, K.; Zemojtel, P.; Wojciechowski, J.W. Development and validation of a virtual gelatin model using molecular modeling computational tools. Molecules 2019, 24, 3365. [Google Scholar] [CrossRef] [Green Version]
- Košovan, P.; Richter, T.; Holm, C. Molecular Simulations of Hydrogels. In Intelligent Hydrogels; Springer Nature: Basel, Switzerland, 2013; pp. 205–220. ISBN 9783319016832. [Google Scholar]
- Saunders, J.R.; Abu-Salih, S.; Khaleque, T.; Hanula, S.; Moussa, W. Modeling theories of intelligent hydrogel polymers. J. Comput. Theor. Nanosci. 2008, 5, 1942–1960. [Google Scholar] [CrossRef]
- Casalini, T.; Perale, G. From Microscale to Macroscale: Nine Orders of Magnitude for a Comprehensive Modeling of Hydrogels for Controlled Drug Delivery. Gels 2019, 5, 28. [Google Scholar] [CrossRef] [Green Version]
- Braccini, I.; Pe, S. Molecular Basis of Ca2+-Induced Gelation in Alginates and Pectins: The Egg-Box Model Revisited. Biomacromolecules 2001, 2, 1089–1096. [Google Scholar] [CrossRef]
- Li, X.; Jia, J.; Mei, Y.; Latour, R.A. Molecular modeling to predict peptide accessibility for peptide-functionalized hydrogels. Biointerphases 2017, 12, 031008. [Google Scholar] [CrossRef] [PubMed]
- Ou, X.; Han, Q.; Dai, H.H.; Wang, J. Molecular dynamic simulations of the water absorbency of hydrogels. J. Mol. Model. 2015, 21, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Eichenbaum, G.M.; Kiser, P.F.; Shah, D.; Simon, S.A.; Needham, D. Investigation of the swelling response and drug loading of ionic microgels: The dependence on functional group composition. Macromolecules 1999, 32, 8996–9006. [Google Scholar] [CrossRef]
- Kamerlin, N.; Elvingson, C. Tracer diffusion in a polymer gel: Simulations of static and dynamic 3D networks using spherical boundary conditions. J. Phys. Condens. Matter 2016, 28. [Google Scholar] [CrossRef]
- Fujiyabu, T.; Yoshikawa, Y.; Chung, U.I.; Sakai, T. Structure-property relationship of a model network containing solvent. Sci. Technol. Adv. Mater. 2019, 20, 608–621. [Google Scholar] [CrossRef] [Green Version]
- Chan, L.W.; Jin, Y.; Heng, P.W.S. Cross-linking mechanisms of calcium and zinc in production of alginate microspheres. Int. J. Pharm. 2002, 242, 255–258. [Google Scholar] [CrossRef]
- Haug, A.; Smidsrød, O.; Högdahl, B.; Øye, H.A.; Rasmussen, S.E.; Sunde, E.; Sørensen, N.A. Selectivity of Some Anionic Polymers for Divalent Metal Ions. Acta Chem. Scand. 1970, 24, 843–854. [Google Scholar] [CrossRef] [Green Version]
- Russo, R.; Malinconico, M.; Santagata, G. Effect of cross-linking with calcium ions on the physical properties of alginate films. Biomacromolecules 2007, 8, 3193–3197. [Google Scholar] [CrossRef]
- Borgogna, M.; Skjåk-bræk, G.; Paoletti, S.; Donati, I. On the Initial Binding of Alginate by Calcium Ions. The Tilted Egg-Box Hypothesis. J. Phys. Chem. B 2013, 117, 7277–7282. [Google Scholar] [CrossRef]
- Brus, J.; Urbanova, M.; Czernek, J.; Pavelkova, M.; Kubova, K.; Vyslouzil, J.; Abbrent, S.; Konefal, R.; Horsky, J.; Vetchy, D.; et al. Structure and Dynamics of Alginate Gels Cross-Linked by Polyvalent Ions Probed via Solid State NMR Spectroscopy. Biomacromolecules 2017, 18, 2478–2488. [Google Scholar] [CrossRef] [Green Version]
- Orban, J.M.; Wilson, L.B.; Kofroth, J.A.; El-kurdi, M.S.; Maul, T.M.; Vorp, D.A. P14986330.pdf. J. Biomed. Mater. Res. A 2004, 68, 756–762. [Google Scholar] [CrossRef] [PubMed]
- Haiyan, L.; Kunlong, M.; Zhenghua, X.; Xiaomei, R.; Gang, Y. Preparation and characteristics of gelatin sponges crosslinked by microbial transglutaminase. PeerJ 2017, 5, e3665. [Google Scholar]
- Dardelle, G.; Subramaniam, A.; Normand, V. Determination of covalent cross-linker efficacy of gelatin strands using calorimetric analyses of the gel state. Soft Matter 2011, 23, 3315–3322. [Google Scholar] [CrossRef]
- Yung, C.W.; Wu, L.Q.; Ullman, J.A.; Payne, G.F.; Bentley, W.E.; Barbari, T.A. Transglutaminase crosslinked gelatin as a tissue engineering scaffold. J. Biomed. Mater. Res. A 2007, 83, 1039–1046. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, H.; Kurosawa, H.; Kokufuta, E.; Veliky, I.A. Preparation of immobilized glucomylase using ca-alginate gel coated with partially quaterized poly(ethleneimine). Biotechnol. Bioeng. 1984, 26, 1393–1394. [Google Scholar] [CrossRef]
- Ro, H.S.; Kim, H.S. Continuous production of gluconic acid and sorbitol from sucrose using invertase and an oxidoreductase of Zymomonas mobilis. Enzyme Microb. Technol. 1991, 13, 920–924. [Google Scholar] [CrossRef]
- Sainz-Polo, M.A.; Ramírez-Escudero, M.; Lafraya, A.; González, B.; Marín-Navarro, J.; Polaina, J.; Sanz-Aparicio, J. Three-dimensional structure of Saccharomyces invertase: Role of a non-catalytic domain in oligomerization and substrate specificity. J. Biol. Chem. 2013, 288, 9755–9766. [Google Scholar] [CrossRef] [Green Version]
- Stewart, W.W.; Swaisgood, H.E. Characterization of calcium alginate pore diameter by size-exclusion chromatography using protein standards. Enzyme Microb. Technol. 1993, 15, 922–927. [Google Scholar] [CrossRef]
- Schrieber, R.; Gareis, H. Gelatine Handbook: Theory and Industrial Practice; WILEY-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2007; ISBN 9783527315482. [Google Scholar]
- Barnett, J.A. Beginnings of microbiology and biochemistry: The contribution of yeast research. Microbiology 2003, 149, 557–567. [Google Scholar] [CrossRef] [Green Version]
- Cantarel, B.I.; Coutinho, P.M.; Rancurel, C.; Bernard, T.; Lombard, V.; Henrissat, B. The Carbohydrate-Active EnZymes database (CAZy): An expert resource for glycogenomics. Nucleic Acids Res. 2009, 37, 233–238. [Google Scholar] [CrossRef]
- Barbosa, O.; Ortiz, C.; Berenguer-Murcia, Á.; Torres, R.; Rodrigues, R.C.; Fernandez-Lafuente, R. Glutaraldehyde in bio-catalysts design: A useful crosslinker and a versatile tool in enzyme immobilization. RSC Adv. 2014, 4, 1583–1600. [Google Scholar] [CrossRef] [Green Version]
- Bilal, M.; Iqbal, H.M.N. Sustainable bioconversion of food waste into high-value products by immobilized enzymes to meet bio-economy challenges and opportunities—A review. Food Res. Int. 2019, 123, 226–240. [Google Scholar] [CrossRef] [PubMed]
- Alcántara, A.R. Biocatalysis and Pharmaceuticals: A Smart Tool for Sustainable Development. Catalysts 2019, 9, 792. [Google Scholar] [CrossRef] [Green Version]
- Schmieg, B.; Döbber, J.; Kirschhöfer, F.; Pohl, M.; Franzreb, M. Advantages of hydrogel-based 3D-printed enzyme reactors and their limitations for biocatalysis. Front. Bioeng. Biotechnol. 2019, 6, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Bagal-Kestwal, D.; Kestwal, R.M.; Chiang, B.H.; Karve, M.S. Development of dip-strip sucrose sensors: Application of plant invertase immobilized in chitosan-guar gum, gelatin and poly-acrylamide films. Sens. Actuators B Chem. 2011, 160, 1026–1033. [Google Scholar] [CrossRef]
- Emregul, E.; Sungur, S.; Akbulut, U. Polyacrylamide-gelatine carrier system used for invertase immobilization. Food Chem. 2006, 97, 591–597. [Google Scholar] [CrossRef]
- Vujčić, Z.; Miloradović, Z.; Milovanović, A.; Božić, N. Cell wall invertase immobilisation within gelatin gel. Food Chem. 2011, 126, 236–240. [Google Scholar] [CrossRef]
- Kotwal, S.M.; Shankar, V. Immobilized invertase. Biotechnol. Adv. 2009, 27, 311–322. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar]
Figure 1.
(a) Schematic representation of calcium ion mediated cross-linking mechanism of alginate strands. The calcium ions form a coordinative bond between G residues forming large pores (approximately 100 Å according to [54]) between cross-linked strands; (b) molecular model of alginate-Ca2+ hydrogel. Green indicates carbon atoms; red, oxygen; white, hydrogen; grey balls between alginate strands, calcium ions.
Figure 1.
(a) Schematic representation of calcium ion mediated cross-linking mechanism of alginate strands. The calcium ions form a coordinative bond between G residues forming large pores (approximately 100 Å according to [54]) between cross-linked strands; (b) molecular model of alginate-Ca2+ hydrogel. Green indicates carbon atoms; red, oxygen; white, hydrogen; grey balls between alginate strands, calcium ions.

Figure 2.
One of the representative gelatin peptide sequences Ala-Gly-Pro-Arg-Gly-Glu-Hyp-Gly-Pro.

Figure 3.
Molecular model of gelatin-based hydrogel cross-linked with microbial transglutaminase (mTGase). The peptide bond is formed between Gln and Lys residues of two gelatin strands, thus limiting the accessible volume between them and enhancing the mechanical resistivity of the obtained polymeric structure.
Figure 3.
Molecular model of gelatin-based hydrogel cross-linked with microbial transglutaminase (mTGase). The peptide bond is formed between Gln and Lys residues of two gelatin strands, thus limiting the accessible volume between them and enhancing the mechanical resistivity of the obtained polymeric structure.

Figure 4.
Molecular model of invertase from Saccharomyces cerevisiae [https://www.rcsb.org/structure/4EQV].
Figure 4.
Molecular model of invertase from Saccharomyces cerevisiae [https://www.rcsb.org/structure/4EQV].

Figure 5.
Molecular model of invertase from Saccharomyces cerevisiae with underlined Lys (purple) and Gln (green) residues.
Figure 5.
Molecular model of invertase from Saccharomyces cerevisiae with underlined Lys (purple) and Gln (green) residues.

Figure 6.
Relative activity of invertase immobilized in alginate-based hydrogel determined after each of 10 consecutive batch processes. Enzyme entrapped without (gray) and with (dotted) additional step of cross-linking with glutaraldehyde (GA). The value of activity obtained in the first reaction cycle was assumed as 100%.
Figure 6.
Relative activity of invertase immobilized in alginate-based hydrogel determined after each of 10 consecutive batch processes. Enzyme entrapped without (gray) and with (dotted) additional step of cross-linking with glutaraldehyde (GA). The value of activity obtained in the first reaction cycle was assumed as 100%.

Figure 7.
Relative activity of invertase immobilized in gelatin-based hydrogel determined after each of 10 consecutive batch processes. Enzyme entrapped without (gray) and with (dotted) additional step of cross-linking with glutaraldehyde. The value of activity obtained in the first reaction cycle was assumed to be 100%.
Figure 7.
Relative activity of invertase immobilized in gelatin-based hydrogel determined after each of 10 consecutive batch processes. Enzyme entrapped without (gray) and with (dotted) additional step of cross-linking with glutaraldehyde. The value of activity obtained in the first reaction cycle was assumed to be 100%.

Figure 8.
Effect of pH on relative activity (a) and stability (b) of free invertase (white square) and immobilized in alginate (grey triangle) and gelatin-based hydrogels (black circle). The highest activity value obtained during the experiments was assumed to be 100%.
Figure 8.
Effect of pH on relative activity (a) and stability (b) of free invertase (white square) and immobilized in alginate (grey triangle) and gelatin-based hydrogels (black circle). The highest activity value obtained during the experiments was assumed to be 100%.

Figure 9.
Effect of temperature on relative activity (a) and stability (b) of free invertase (white square) and immobilized in alginate (grey triangle) and gelatin-based hydrogels (black circle). The highest activity value obtained during the experiments was assumed to be 100%.
Figure 9.
Effect of temperature on relative activity (a) and stability (b) of free invertase (white square) and immobilized in alginate (grey triangle) and gelatin-based hydrogels (black circle). The highest activity value obtained during the experiments was assumed to be 100%.

Figure 10.
Storage stability of free invertase (black) and immobilized in alginate (striped) and gelatin-based hydrogels (grey) over 28 days of storage at 4 °C. The value of activity obtained on the beginning of storage (day 0) was assumed to be 100%.
Figure 10.
Storage stability of free invertase (black) and immobilized in alginate (striped) and gelatin-based hydrogels (grey) over 28 days of storage at 4 °C. The value of activity obtained on the beginning of storage (day 0) was assumed to be 100%.

Figure 11.
Comparison of the entrapment procedures of invertase in alginate and gelatin-based hydrogels.
Figure 11.
Comparison of the entrapment procedures of invertase in alginate and gelatin-based hydrogels.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Amino acid composition of gelatin strands [55].
Table 1.
Amino acid composition of gelatin strands [55].
| Amino Acid | Percentage (%) | Amino Acid | Percentage (%) |
|---|---|---|---|
| Glycine (Gly) | 32.9 | Threonine (Thr) | 1.80 |
| Proline (Pro) | 13.2 | Phenylalanine (Phe) | 1.4 |
| Alanine (Ala) | 11.2 | Isoleucine (Ile) | 1.0 |
| Hydroxyproline (HPro) | 9.1 | Hydroxylysine (HLys) | 0.6 |
| Glutamic acid (Glu) | 4.8 | Asparagine (Asn) | 2.9 |
| Arginine (Arg) | 4.9 | Histidine (His) | 0.6 |
| Aspartic acid (Asp) | 2.9 | Tyrosine (Tyr) | 0.3 |
| Serine (Ser) | 3.5 | Methionine (Met) | 0.4 |
| Lysine (Lys) | 2.7 | Glutamine (Gln) | 2.5 |
| Leucine (Leu) | 2.3 | Cysteine (Cys) | 0.0 |
Table 2.
Amino acid composition of one polypeptide chain of invertase from Saccharomyces cerevisiae, calculated based on the FASTA code [https://www.uniprot.org/uniprot/P00724.fasta].
Table 2.
Amino acid composition of one polypeptide chain of invertase from Saccharomyces cerevisiae, calculated based on the FASTA code [https://www.uniprot.org/uniprot/P00724.fasta].
| Amino Acid | Percentage (%) | Amino Acid | Percentage (%) |
|---|---|---|---|
| Glycine (Gly) | 5.8 | Threonine (Thr) | 7.6 |
| Proline (Pro) | 5.1 | Phenylalanine (Phe) | 7.0 |
| Alanine (Ala) | 5.7 | Isoleucine (Ile) | 3.5 |
| Valine (Val) | 5.7 | Tryptophan (Trp) | 3.1 |
| Glutamic acid (Glu) | 5.8 | Asparagine (Asn) | 8.6 |
| Arginine (Arg) | 2.5 | Histidine (His) | 0.8 |
| Aspartic acid (Asp) | 6.2 | Tyrosine (Tyr) | 6.0 |
| Serine (Ser) | 9.0 | Methionine (Met) | 2.0 |
| Lysine (Lys) | 4.9 | Glutamine (Gln) | 3.5 |
| Leucine (Leu) | 6.6 | Cysteine (Cys) | 0.4 |
Table 3.
Effect of the concentration of CaCl2 on invertase immobilization in alginate-based hydrogel.
Table 3.
Effect of the concentration of CaCl2 on invertase immobilization in alginate-based hydrogel.
| CaCl2 Concentration (% w/v) | Activity of Bound Enzyme (U mL−1) | Immobilization Yield (%) | Enzyme Leakage (%) |
|---|---|---|---|
| 2 | 7570 | 38.0 | 20.7 |
| 4 | 9210 | 46.2 | 17.9 |
| 10 | 10,300 | 51.7 | 16.2 |
| 15 | 12,202 | 61.2 | 10.4 |
| 20 | 11,513 | 57.7 | 11.0 |
| 30 | 4012 | 20.1 | 10.7 |
Table 4.
Effect of the concentration of glutaraldehyde (GA) on invertase immobilization in alginate-based hydrogel (cross-linking time 10 min).
Table 4.
Effect of the concentration of glutaraldehyde (GA) on invertase immobilization in alginate-based hydrogel (cross-linking time 10 min).
| GA Concentration (% v/v) | Activity of Bound Enzyme (U mL−1) | Immobilization Yield (%) | Relative Activity of Bound Enzyme (%) | Enzyme Leakage (%) |
|---|---|---|---|---|
| 0.0 | 11,975 | 60.8 | 100 | 11.2 |
| 0.5 | 9292 | 47.2 | 77.6 | 8.6 |
| 1.0 | 8924 | 45.3 | 74.5 | 8.5 |
| 2.0 | 8620 | 43.7 | 72.0 | 7.4 |
| 4.0 | 6900 | 35.0 | 57.6 | 7.3 |
Table 5.
Effect of the time of cross-linking with 2.0% glutaraldehyde (GA) on invertase immobilization in alginate-based hydrogel.
Table 5.
Effect of the time of cross-linking with 2.0% glutaraldehyde (GA) on invertase immobilization in alginate-based hydrogel.
| Cross-Linking Time (min) | Activity of Bound Enzyme (U mL−1) | Immobilization Yield (%) | Relative Activity of Bound Enzyme (%) | Enzyme Leakage (%) |
|---|---|---|---|---|
| 0 | 11,471 | 59.1 | 100 | 11.0 |
| 5 | 9693 | 49.9 | 84.5 | 9.2 |
| 10 | 8631 | 44.5 | 75.2 | 7.9 |
| 20 | 8493 | 43.8 | 74.0 | 7.0 |
| 30 | 8383 | 43.2 | 73.1 | 5.5 |
| 45 | 8300 | 42.8 | 72.4 | 5.3 |
| 60 | 8245 | 42.5 | 71.9 | 5.4 |
Table 6.
Effect of the concentration of microbial transglutaminase (mTGase) on invertase immobilization in gelatin-based hydrogel.
Table 6.
Effect of the concentration of microbial transglutaminase (mTGase) on invertase immobilization in gelatin-based hydrogel.
| mTgase Concentration (% w/v) | Activity of Bound Enzyme (U mL−1) | Immobilization Yield (%) | Enzyme Leakage (%) |
|---|---|---|---|
| 0.25 | 3932 | 19.7 | 8.4 |
| 0.50 | 4503 | 22.6 | 3.3 |
| 1.00 | 4884 | 24.5 | 0.0 |
| 2.00 | 4820 | 24.2 | 0.0 |
| 3.00 | 4725 | 23.7 | 0.0 |
| 4.00 | 4852 | 24.3 | 0.0 |
Table 7.
Effect of the concentration of glutaraldehyde (GA) on invertase immobilization in gelatin-based hydrogel (cross-linking time 10 min).
Table 7.
Effect of the concentration of glutaraldehyde (GA) on invertase immobilization in gelatin-based hydrogel (cross-linking time 10 min).
| GA Concentration (% v/v) | Activity of Bound Enzyme (U mL−1) | Immobilization Yield (%) | Relative Activity of Bound Enzyme (%) | Enzyme Leakage (%) |
|---|---|---|---|---|
| 0.0 | 4826 | 24.5 | 100 | 0.0 |
| 0.5 | 4338 | 22.0 | 89.9 | 0.0 |
| 1.0 | 3884 | 19.7 | 80.5 | 0.0 |
| 2.0 | 3091 | 15.7 | 64.0 | 0.0 |
| 4.0 | 2352 | 11.9 | 48.7 | 0.0 |
Table 8.
Effect of the time of cross-linking with 0.5% glutaraldehyde (GA) on invertase immobilization in gelatin-based hydrogel.
Table 8.
Effect of the time of cross-linking with 0.5% glutaraldehyde (GA) on invertase immobilization in gelatin-based hydrogel.
| Cross-Linking Time (min) | Activity of Bound Enzyme (U mL−1) | Immobilization Yield (%) | Relative Activity of Bound Enzyme (%) | Enzyme Leakage (%) |
|---|---|---|---|---|
| 0 | 4249 | 22.4 | 100 | 0.0 |
| 5 | 3832 | 20.2 | 90.2 | 0.0 |
| 10 | 3801 | 20.0 | 89.4 | 0.0 |
| 20 | 3203 | 16.8 | 75.4 | 0.0 |
| 30 | 3076 | 16.2 | 72.4 | 0.0 |
| 45 | 3013 | 15.8 | 70.9 | 0.0 |
| 60 | 3044 | 16.0 | 71.6 | 0.0 |
Table 9.
Comparison of selected features of the invertase immobilized in gelatin-based carrier developed in the current study with other literature reports.
Table 9.
Comparison of selected features of the invertase immobilized in gelatin-based carrier developed in the current study with other literature reports.
| No. | Temperature Optimum [°C] | Thermostability* [%] | Operational Stability* after 10 Cycles [%] | Storage Stability* after 1 month [%] | Ref. [-] |
|---|---|---|---|---|---|
| 1 | 55 | 70 (in 60 °C) 40 (in 70 °C) | 95 | 82 | Current study |
| 2 | 45 | 28 (in 65 °C) | 71 | 93 | [62] |
| 3 | 55 | 30 (in 70 °C) | 30–80 | - | [63] |
| 4 | 60 | 75 (in 60 °C) 10 (in 70 °C) | 90 | - | [64] |
* relative activity in defined conditions expressed in %.
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Labus, K.; Wolanin, K.; Radosiński, Ł. Comparative Study on Enzyme Immobilization Using Natural Hydrogel Matrices—Experimental Studies Supported by Molecular Models Analysis. Catalysts 2020, 10, 489. https://doi.org/10.3390/catal10050489
AMA Style
Labus K, Wolanin K, Radosiński Ł. Comparative Study on Enzyme Immobilization Using Natural Hydrogel Matrices—Experimental Studies Supported by Molecular Models Analysis. Catalysts. 2020; 10(5):489. https://doi.org/10.3390/catal10050489
Chicago/Turabian StyleLabus, Karolina, Kamila Wolanin, and Łukasz Radosiński. 2020. "Comparative Study on Enzyme Immobilization Using Natural Hydrogel Matrices—Experimental Studies Supported by Molecular Models Analysis" Catalysts 10, no. 5: 489. https://doi.org/10.3390/catal10050489
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.