


Ianus Bifrons: The Two Faces of Metformin
by
 and
and
Umberto Goglia
1,*,
Iderina Hasballa
2,3,
Claudia Teti
4,
Mara Boschetti
2,3,
Diego Ferone
2,3 and
Manuela Albertelli
2,3 1
Endocrinology and Diabetology Unit, Local Health Authority CN1, 12100 Cuneo, Italy
2
Endocrinology Unit, IRCCS Ospedale Policlinico San Martino, 16132 Genova, Italy
3
Endocrinology Unit, Department of Internal Medicine and Medical Specialties (DiMI), University of Genova, 16132 Genoa, Italy
4
Endocrinology and Diabetology Unit, Local Health Autorithy Imperia 1, 18100 Imperia, Italy
*
Author to whom correspondence should be addressed.
Cancers 2024, 16(7), 1287; https://doi.org/10.3390/cancers16071287
Submission received: 18 January 2024
/
Revised: 10 March 2024
/
Accepted: 20 March 2024
/
Published: 26 March 2024
(This article belongs to the Special Issue Cancer and Diabetes: What Connections Lie between Them?)
Abstract
:Simple Summary
Metformin is one of the most widely used drugs in the world, since its first use in the second part of the 1950s in France (Aron, 1957) and in the United Kingdom (Rona, 1958), and is on the World Health Organization’s List of Essential Medicines. Traditionally, metformin is used in diabetes mellitus, mainly in overweight and obese subjects, but also in other conditions of impaired glucose metabolism, such as insulin resistance. In recent years, in vitro and in vivo data on this molecule as an antiproliferative modulator have suggested a series of clinical trials in several tumors that—at the moment—have not confirmed the expected positive results. The aim of this paper is to offer a double vision of this molecule, with a very light side about the metabolic aspects and a darker side concerning its antineoplastic activity, focusing on molecular and subcellular pathways involved in both fields and discussing the contrast between preliminary cellular data and real clinical outcomes in different neoplasms.
Abstract
The ancient Roman god Ianus was a mysterious divinity with two opposite faces, one looking at the past and the other looking to the future. Likewise, metformin is an “old” drug, with one side looking at the metabolic role and the other looking at the anti-proliferative mechanism; therefore, it represents a typical and ideal bridge between diabetes and cancer. Metformin (1,1-dimethylbiguanidine hydrochloride) is a drug that has long been in use for the treatment of type 2 diabetes mellitus, but recently evidence is growing about its potential use in other metabolic conditions and in proliferative-associated diseases. The aim of this paper is to retrace, from a historical perspective, the knowledge of this molecule, shedding light on the subcellular mechanisms of action involved in metabolism as well as cellular and tissue growth. The intra-tumoral pharmacodynamic effects of metformin and its possible role in the management of different neoplasms are evaluated and debated. The etymology of the name Ianus is probably from the Latin term ianua, which means door. How many new doors will this old drug be able to open?
1. Introduction
…cur de caelestibus unus
Sitque quod a tergo sitque quod ante vides.
Tum sacer ancipiti mirandus imagine Ianus
bina repens oculis obtulit ora meis [1].
In ancient Italian and Latin religions, Ianus is considered a singular divinity with mysterious features. In fact, he is the god of material and immaterial primordial times in human history, and his figure is strictly connected to the concept of time.
The picture of Ianus shows a double facial image in opposite vision, one to the past, the other to the future (Figure 1).
In the divinity pantheon, Ianus occupies a very relevant and superior seat; indeed, he is also named Divus Deus (“God of Gods”), Divum Pater (“Father of the Gods”), and Ianus Bifrons (“God with two faces”), and he is also the only idol without his own parents.
Moreover, in a mythological vision of the ancient history of Rome, Ianus was the first king of the land of Latium, founding the ancient town on top of Gianicolo Hill, receiving the feature of the vision of the past and of the future from the god Saturnus.
Just like the Father of Gods, metformin is an old, ambivalent molecule, presenting at least a double pathophysiological connection, looking to metabolic effects and to antiproliferative actions.
1.1. Historic Perspective
In 1772, the famous botanist Sir John Hill described the Galega Officinalis as “Perennial, native of Spain and Italy; of Greece and Africa, a specious plant, of a yard high, that flowers in August. The Stalk is juicy, and green… the Flowers are purples; sometimes white” (Figure 2). This plant—also named goat’s rue, French lilac, Italian fitch, Spanish sainfoin, professor weed, or Herba routae caprariae—was used in medieval Europe as a traditional cure for worms, epilepsy, fever, and pestilence and to treat conditions of thirst and frequent urination [2,3].
In the nineteenth century, the works of Adolf Strecker and Bernhard Rathke, with the preparation of guanidine and the fusion of two guanidine molecules to form biguanides, respectively, were determinants [4,5]. Therefore, the synthesis of metformin (dimethyl-biguanide) was carried out in 1922 by Wermer and Bell, showing lower toxicity than the other mono- and diguanidines [6,7]. Later, an independent research line about antimalaric agents documented that metformin caused a lowering of blood glucose in animal studies [8,9]. The most persevering and passionate scientist who dedicated his own life to the study of metformin was Jean Sterne, a physician at the hospital in Casablanca and later at Aron Laboratories in Suresnes, in the west of Paris, France [10]. Between 1957 and 1958, Stern showed that N,N-dimethylamine guanyl guanidine (metformin) had a blood sugar-lowering effect, replacing the need for insulin in subjects with a relative deficiency of insulin [11,12,13]. To market metformin, Aron adopted the trade name ‘glucophage’ (from the ancient Greek language, meaning glucose eater); later, Jean Stern played a prominent role in further research and physician education to assist the introduction of metformin into clinical practice in Europe [14,15,16,17,18]. Metformin, unlike the other biguanides (mainly phenphormin, removed from the market in the USA in 1978), showed a more favorable safety profile, with distinct and specific differences in pharmacokinetic features [19,20].
1.2. Chemical and Pharmacological Features
The chemical structures of guanine, guanidine, and biguanides are shown in Table 1. Metformin (1,1-dimethylbiguanide hydrochloride) is a relatively planar hydrophilic molecule, monoprotonated at neutral pH with several tautomeric configurations.
Absorption. After the oral intake (approximately 35 mg/kg/day, that is 500–2550 mg/day), metformin is absorbed mainly in the small intestine cells (20% duodenum, 60% jejunum, and ileum, 20% colon). After 1.5 h is the onset of action, about 1.5–4.9 h is the half-life in circulation, and 16–20 h is the duration of action. Metformin hydrophilicity represents a limiting step of oral absorption due to the low intestinal permeability [21,22,23].
Distribution. The distribution of metformin is rapid and without binding to plasma proteins [24,25]. In hepatic tissue, the concentration is three- to five-fold higher than in the portal vein (40 µmol/L), so the hepatocyte is the primary site of drug function [26]. Generally, within 24–48 h, steady-state plasma concentrations are reached (approximately <1 mcg/mL).
The presence and expression of transporters for cationic compounds (OCT), plasma membrane monoamine transporter (PMAT), and multidrug and toxin extrusion proteins (MATE) are critical for the biodistribution and pharmacodynamics of the biguanide [27,28,29,30,31] (Figure 3).
Metabolism and elimination. Metformin, not metabolized, is present unchanged in the urine, with a half-life of about 5 h. In the kidney, active tubular secretion is the main route of removal, with a plasma elimination rate of about 500 mL/min [22,32]. Low molecular weight, absence of plasma protein binding, presence of renal transporters, and low lipid solubility are the main factors inhibiting passive reabsorption. The impairment of renal function reduces the clearance of drugs [33].
Therapeutic range. The therapeutic range of metformin is unclear, but it should not exceed 5 mg/L [22,34]. Both formulations of metformin (immediate and extended release) displayed similar areas under curve (AUC) and equal safety profiles and efficacy [22]. Lactic acidosis is the most dangerous potential risk, so in critical cases and frail subjects, an accurate monitoring should be performed, maintaining a drug concentration below 2.5 mg/L [25].
2. Metformin Target Organs
2.1. Liver
The liver and skeletal muscle have been considered the major target organs of metformin action for many years, while, more recently, other sites of action have been highlighted for an important role beyond simple glycemic control, among these: the gastrointestinal tract, intestinal microbial communities, and tissue-resident immune cells. The pharmacodynamics, even at the molecular level, appear to be influenced by the dose and duration of treatment with metformin [35]. Metformin has been defined as an insulin sensitizer, leading to a reduction in insulin resistance and therefore optimizing cellular glucose uptake, mainly in skeletal muscle, leading to a reduction in plasma glucose and insulin plasma values. This effect of metformin could be attributed to its positive effects on insulin receptor expression and tyrosine kinase activity on the phosphorylation of insulin receptors [36]. Most studies in the literature conclude that the main function through which metformin reduces hepatic glucose production is the inhibition of gluconeogenesis [37,38]. Inhibition of gluconeogenesis has been explained by both demonstrating changes in enzymatic activity such as pyruvate kinase flux and modifications of citric acid cycle activity and also highlighting a reduction in hepatic uptake of gluconeogenic substrates [39,40,41,42].
2.2. Gastrointestinal System
More recently, the gastrointestinal tract has been identified as an additional site of action for metformin; in fact, a high accumulation of metformin in the intestine has been reported, both in humans and in animal models, with concentrations up to a couple of hundred times higher than those of plasma and other tissues, suggesting that the intestine acts as an important reservoir of metformin. Metformin increases glucose absorption in the basolateral intestine and, through gut–liver communication, influences hepatic glucose production [43,44,45,46]. Several findings suggest that the hypoglycemic capacity of metformin depends both on intestinal glucose absorption along the gastrointestinal tract and on bloodstream absorption. After entering enterocytes, anaerobic glucose metabolism causes the accumulation of lactate and acetate in the wall of the small intestine and its release into the circulation, creating an intestinal–liver communication to attenuate gluconeogenesis [44,47,48,49]. Metformin can therefore exert its beneficial metabolic actions through modulation of the incretin axis, enhancing circulating levels of glucagon-like peptide 1 (GLP-1), inducing glucagon-like peptide-1 (GLP-1) receptor gene expression, and reducing plasma dipeptidyl peptidase-4 activity through a mechanism that depends on PPAR (peroxisome proliferator-activated receptor)-α [50,51].
2.3. Brown Adipose Tissue
An increasing number of studies have shown that BAT, in addition to having a role in dissipating energy through heat production, contributes to the regulation of glucose homeostasis [52,53]. The presence of metformin has been demonstrated in the interscapular BAT of mice using 11C-metformin PET imaging, which supports the hypothesis that BAT could be another important target of metformin [54]. At the BAT level, metformin also appears to be involved in lipid metabolism; in fact, by increasing the activity of hormone-sensitive lipase and AMP-activated protein kinase (AMPK), metformin promotes intracellular triglyceride uptake, lipolysis, and subsequent mitochondrial fatty acid oxidation [55,56].
3. Molecular Mechanisms Involved in Metformin’s Action
3.1. Metformin-Induced Reduction in Blood Levels of Glucose (“Metabolic Face”)
The main mechanisms producing the anti-hyperglycemic action of metformin reside in mitochondria and lysosomes, after the interaction of metformin with organic cationic transporter 1 [57,58].
Organic Cation Transporter 1 (OCT1), a member of the family of membrane transporters named Solute Carrier 22 (SLC22), facilitates the movement of endogenous and exogenous compounds across cell membranes and is primarily localized on the basolateral membrane of hepatocytes. Metformin is a well-known substrate of OCT1, and genetic polymorphisms of OCT1 are known to reduce the effectiveness of metformin by lowering absorption and causing gastrointestinal intolerance to metformin [59,60,61]. A recent paper by Zeng et al., using cryo-electron microscopy, showed the interaction between drug and OCT1 in different conditions, substrate-free and substrate-bound, with a resolution of 3.5 Å. Conformational changes from outward-to inward-facing states are demonstrated for the first time in a very suggestive way [58].
3.1.1. Complex I Inhibition-Dependent Mechanism
Mitochondrial Complex-I (NADH: ubiquinone oxidoreductase) is a crucial component for respiration in aerobic organisms, oxidizing NADH from the tricarboxylic acid cycle and β-oxidation. In hepatocytes, metformin induces a reversible inhibition of mitochondrial respiratory chain complex I, as documented by studies combining cryo-electron microscopy and enzyme kinetics [62,63]. After the intracellular uptake of metformin, three main phenomena demonstrated in hepatic cells are a higher NADH/NAD+ ratio, a reduction in ATP concentration, and increased levels of AMP.
3.1.2. Mitochondrial Glycerol-3-Phosphate Dehydrogenase (mGPDH)-Dependent and Complex IV Inhibition-Dependent Mechanism
Complex-IV is a cytochrome c oxidase involved in the final steps of energy conservation. At the hepatic level, metformin acts directly and indirectly via Complex-IV, inhibiting mGPDH, with the following results: a higher NADH/NAD+ ratio, a reduction in gluconeogenesis from lactate, and a reduction in the activity of the glycerol–phosphate shuttle, which transfers NADH from the cytosol to mitochondria [64,65]. Finally, the hepatic redox state is raised through an increase in the glutathione to oxidized glutathione ratio (GSH:GSSG), with inhibitions of genes involved in the process of gluconeogenesis.
3.1.3. AMPK Activation-Dependent Mechanisms in Lysosomes
AMP-activated protein kinase (AMPK) is a master controller of metabolic homeostasis. At low concentrations, metformin binds presenilin enhancer 2 (PEN2), which is recruited to ATPase H+ transporting accessory protein 1 (ATP6AP1) independent of changes in AMP levels, leading to inhibition of v-ATPase and phosphorylation/activation of AMPK through the formation of a supercomplex containing the v-ATPase, AXIN, liver kinase B1 (LKB1), and AMPK [66]. Metformin-activated AMPK from lysosomes reduces lipid accumulation in the liver via acetyl-CoA carboxylase (ACC) inhibition and increases glucagon-like peptide 1 (GLP1) secretion in the gut, inducing reductions in blood levels of glucose.
3.2. Anticancer Molecular Mechanisms of Metformin (“Anti-Proliferative Face”)
Accumulating data from preclinical studies support the anti-neoplastic activity of metformin in several malignancies, thus providing the rationale for further exploration of the biguanide in more than 130 clinical trials [67]. Nevertheless, its underlying molecular mechanisms have not been fully clarified. To date, metformin anti-carcinogenic impact has been generally classified as direct, i.e., glucose- and insulin-independent, or indirect, i.e., glucose- and insulin-dependent, both activities not mutually exclusive [68] (Figure 4).
3.2.1. Direct Anticancer Mechanisms of Metformin
The mTOR Pathway
Cancer cells, in comparison to normal ones, present an aberrant metabolism with higher requests for catabolite uptake and utilization, which are necessary for survival and growth. Reprogramming cellular energy homeostasis, in fact, represents one of the essential mechanisms through which metformin appears to directly attenuate tumorigenesis and progression [68]. A crucial anti-tumor target of the biguanide concerns the mechanistic target of rapamycin complex 1 (mTORC1), which is a major driver of protein biosynthesis, cell growth, and metabolism as a response to different stimuli such as growth factors, nutrient availability, energy, and oxygen intracellular levels [69]. In particular, metformin selectively inhibits mitochondrial respiratory-chain complex 1 (NADH coenzyme Q oxidoreductase), ultimately leading to decreased cell respiration, reduced oxidative phosphorylation (OXPHOS), and ATP depletion. Metformin-induced energy stress triggers the activation of AMP-kinase (AMPK), which plays a key role in modulating critical pathways such as mTORC1 signaling. Specifically, AMPK suppresses mTORC1 activity directly via phosphorylation of S722 and S792 on Raptor, its scaffolding protein, as well as through the activation of the tuberous sclerosis complex (TSC). The inhibitory impact of TSC1 and TSC2 is also exerted on the mTORC1 downstream major effectors such as eukaryotic initiation factor 4E-binding protein 1 (4EBP1) and ribosomal protein kinase S6 (S6K) [68]. Furthermore, metformin may suppress mTORC1 activity in an AMPK-independent way by inhibiting Rag GTPases, essential for the amino acid signaling to mTORC1, as well as by promoting the activation during hypoxic stress of REDD1 (regulated in development and DNA damage responses), a hypoxia-inducible factor-1 target gene involved in cell survival regulation [70,71,72].
The anticancer activities of the biguanide via repression of the mTORC1 pathway, as above-mentioned, were documented in several preclinical studies performed in different malignancies such as lung cancer, pancreatic cancer, prostate cancer, breast cancer, thyroid cancer, meningioma, leukemia, and lymphoma [73,74,75,76,77,78,79,80].
The PI3K/AKT/mTOR Pathway
Another anticancer metformin mechanism of action involves the PI3K/AKT/mTOR (PAM) pathway, which is a major signaling network modulating cell growth, metabolism, proliferation, as well as apoptosis and autophagy [68,81]. The overactivation of the PAM axis represents one of the main drivers of tumor pathogenesis and progression, as well as of anti-tumor therapeutic resistance. Several preclinical studies in different cancer types have suggested that metformin may exhibit its antiproliferative effects through the inhibition of PI3K/AKT/mTOR. In bladder cancer cells, the biguanide in a concentration-dependent manner led to the reduction of PI3K, AKT, and mTOR phosphorylation and was ultimately associated with the suppression of cell proliferation and migration, the activation of the caspase cascade, and the induction of apoptosis [82]. In a study performed by Nozhat et al. in anaplastic thyroid cancer (ATC) cell lines, metformin in a time- and dose-dependent way repressed cell growth, significantly altered ATC cellular morphology, and decreased cell migration, likely by reducing the mRNA expression of PI3K and AKT, with no impact, however, on their phosphorylation status [83]. Tang and colleagues demonstrated the antiproliferative activity of biguanide also in esophageal cancer cells, mediated partly by the suppressed expression of the insulin-like growth factor 1 receptor (IGF-1R) and its downstream targets PI3K, AKT, mTOR, p70S65, and PKM2 [84]. Furthermore, metformin not only inhibited cervical cancer cell survival and proliferation, but also increased NK cell cytotoxicity by modulating through the PI3K/AKT axis the expression of MICA and HSP70 proteins on the cellular surface [85]. In colorectal cancer cells (CRCs), biguanide exhibited its inhibitory effect on CRC growth by down-regulating the PI3K/AKT pathway and repressing the expression of inhibin BetaA (INHBA), an oncogene member of the TGF-beta family [86]. Finally, the antiproliferative activity of metformin mediated by the interference with the PAM axis was also documented in preclinical studies conducted in ovarian, endometrial, and breast cancer, hepatocellular carcinoma, and mouse melanoma B16 cells [87,88,89,90].
The K-Ras Pathway
A novel anti-tumor target of metformin is the K-Ras pathway, which plays a crucial role in cell signal transduction, differentiation, and proliferation. Aberrant K-Ras contributes to oncogenesis and is frequently associated with poor outcomes and resistance to anti-EGFR therapy. Several studies in vitro and in vivo documented the inhibitory effects of the biguanide on the growth of K-Ras-mutated cancer cells. In particular, in endometrial cancer models, metformin inhibited cell proliferation and triggered apoptosis in a concentration-dependent manner by displacing the oncogenic K-Ras from the plasma membrane with the subsequent suppression of its biological activity, as well as by down-regulating downstream MAPK signaling [91]. In K-Ras-mutated CRC, metformin exerted antiproliferative effects by inactivating both the RAS/ERK and AKT/mTOR pathways [92]. Moreover, in K-Ras mutant lung adenocarcinoma and pancreatic cancer cell lines, biguanide, in a dose-dependent manner, induced apoptosis and suppressed cell proliferation by targeting crucial downstream effectors of K-Ras signaling such as MAPK and AKT [93].
The NKL Pathway
The anti-tumor activity of metformin is also exerted by interfering with nemo-like kinase (NLK), a member of the MAPK family, which carries out a relevant role in cell cycle progression and contributes to the oncogenesis of several neoplasms, including colon, prostate, lung, and hepatocellular cancer. Data in vitro and in vivo in non-small-cell lung cancer (NSCLC) cell lines showed that the biguanide—via inhibition of NLK expression—induced cell cycle arrest and significant reduction of the stem cell tumor population [94].
The JNK Pathway
Metformin may exert anti-proliferative activity also by interfering with c-Jun-N-terminal kinase (JNK, also known as stress-activated protein kinase 1—SAPK1) signaling, another MAPK-involved pathway that regulates cell growth, survival, proliferation, and migration [95]. In osteosarcoma cell lines, the biguanide activates the JNK cascade and promotes cell cycle arrest and programmed cell death processes, including apoptosis and autophagy [96]. Similarly, in lung cancer cell lines, metformin led to increased apoptosis and suppression of cell proliferation in a dose- and time-dependent manner, either by activating JNK/p38 MAPK signaling or upregulating the expression of DNA damage inducible gene 153 (GADD153) [97]. In gastric adenocarcinoma cell lines, biguanide was associated with remarkable anti-proliferative activity and apoptosis induction also through the phosphorylation reduction of several MAPKs such as JNK, ERK, and p38, in addition to the activation of AMPK and repression of the AKT/mTOR pathway [98].
Moreover, He et colleagues documented the inhibitory effects of metformin on the viability of thyroid cancer TPC-1 cells via down-regulation of LRP2, a transmembrane receptor mainly involved in lipid metabolism, ultimately leading to the suppression of JNK signaling in a concentration-dependent way [99].
The STAT3 Pathway
Another pathway involved in the anti-neoplastic effects of metformin concerns STAT3 (signal transducer and activator of transcription 3), which is a promising cancer therapeutic target due to its crucial role in cell survival, proliferation, and migration. Preclinical studies suggest that the biguanide activity is mediated by reducing STAT3 phosphorylation and/or suppressing its nuclear translocation, thus also leading to the downregulation of its target genes such as cyclin D1, Bcl-XL, and Bcl2. Cyclin D1 is a vital regulator of cell cycle progression, which promotes cell cycle G1/S phase transition, while Bcl-XL and Bcl2 are pro-survival genes directly transcribed by STAT3 and involved in programmed cell death modulation [100]. In fact, by repressing the STAT3 pathway in in vitro and in vivo models, metformin inhibited cell proliferation and induced apoptosis and autophagy in a dose- and time-dependent manner in several cancer types, including esophageal squamous cell carcinoma, triple-negative breast cancer, endometrial, ovarian, bladder cancer, glioblastoma, chronic neutrophilic leukemia with mutated CSF3R, and cholangiocarcinoma [101,102,103,104,105,106,107]. Furthermore, in bladder cancer models, metformin also suppressed cell migration and invasion, ultimately interfering with the progression of precancerous lesions. The underlying mechanism of action suggested is the biguanide-induced inhibition of STAT3 signaling, which regulates the activity of matrix-metalloproteinases (MMPs), Rho, and Rac proteins that carry out a key role in modulating cellular migration [108]. Additionally, in primary breast cancer cells, high doses of metformin attenuated cancer progression via suppression of the STAT3 and NF-kB pathways. In particular, the biguanide inhibited the IL-6 epithelial–mesenchymal transition (ETM) and decreased the expression of mesenchymal markers, which are crucial for tumor metastasis [109].
The HER2 Pathway
Metformin is also associated with anticancer activity in HER-2 positive cancer via interference with human epidermal growth factor receptor-2 (HER-2) signaling. HER2 is a relevant oncogene able to modulate several key genes such as TP53, CDK12, PI3KCA, and PTEN, which furthermore contribute to tumor aggressiveness and progression [110,111,112]. In particular, in the study performed by Vazquez-Martin on breast carcinoma (BC) cells, biguanide in a dose- and time-dependent manner led to a significant reduction of HER-2 expression, mainly by inhibiting the downstream effector of mTOR p70S6K1, also in an AMPK-independent way [113]. In another preclinical model of HER-2 BC cells, metformin inhibited cell proliferation and induced apoptosis, likely due to the inhibition of heat shock protein 90 (HSP90), which downregulates the AKT and MAPK pathways [114].
The NF-κB Pathway
Other antitumor effects of metformin are mediated by interference with the nuclear factor κB pathway (NF-κB), which is involved in cancer development, invasion, and metastasis via modulation of EMT, as well as in therapeutic resistance [116]. In the study of Li and colleagues, conducted in EGFR-mutant lung cancer models with acquired resistance to EGFR tyrosine kinase inhibitors (TKIs), the biguanide inhibited cell proliferation, induced apoptosis, reversed or delayed the TKIs resistance, and repressed cancer cell stemness by inactivating ERK/NF-κB signaling in an AMPK-dependent way [117]. Metformin is able to trigger caspase3/GSDME-mediated pyroptosis of cancer cells via stimulation of the AMPK/SIRT1/NF-κB pathway and mitochondrial dysregulation [118]. Furthermore, in primary breast cancer cell lines, metformin presents an inhibitory effect on tumor invasion and metastasis due to the dual suppression of NF-kB activity and nuclear translocation mediated by MMP-9 downregulation [119], and in prostate cancer models, metformin appeared to attenuate metastasis by repressing NF-Kb signaling, thus leading to the suppression of tumor necrosis factor-α-induced EMT [120]. In a preclinical study performed on ovarian cancer cells, biguanide was shown to suppress cancer progression and reduce chemoresistance via modulation of NF-κB and IL-6 signaling [121].
3.2.2. Indirect Anticancer Mechanisms of Metformin
The indirect antineoplastic effects of metformin are exerted by modulating blood glucose and insulin concentration. In fact, the biguanide down-regulates insulin and insulin-binding proteins, leading to decreased levels of insulin growth factor-1 [68]. The IGF axis plays a crucial role in cell growth and metabolism. Moreover, IGF-1 and IGF-2, in particular, promote both mitogenic as well as antiapoptotic signaling via activation of other key pathways such as PI3K/AKT/mTOR and RAS/RAF/MAPK [122]. Additionally, metformin-induced AMPK activation reduces insulin receptor substrate-1 (IRS-1) phosphorylation, which interferes with the PI3K/AKT/mTOR axis too [68]. Finally, in endometrial cancer cells, metformin presented antiproliferative effects by reducing IGF-1 secretion and IGF-1R expression, ultimately leading to the inhibition of downstream PI3K/AKT signaling [123], whereas considering breast tumors, in triple-negative cancer cell lines, a synergistic effect of metformin combined with an insulin/IGF-1 inhibitor in suppressing cell growth and proliferation was documented [124,125].
4. Clinical Trials on Metformin Use
4.1. Clinical Studies on Hyperglicemic Conditions (“Metabolic Face”)
4.1.1. Type 2 Diabetes Mellitus
The series of United Kingdom Prospective Diabetes Studies (UKPDS) represent the main source about the normoglycemic role of metformin, involving subjects with type 2 diabetes mellitus in intensive and conventional dietary treatment. In overweight individuals taking metformin, reductions in the risk of myocardial infarction of 39% (p = 0.01) and of death from any cause of 36% (p = 0.01) were observed [126,127,128,129].
Moreover, Holman and colleagues conducted post-trial monitoring of UKPDS, demonstrating that in the overweight group (342 subjects in metformin), compared with subjects in conventional therapy, differences in glycated hemoglobin levels were lost after the first year, but significant risk reductions persisted for several end points. This phenomenon was called the “legacy effect”, the persistence of clinical benefits despite the early loss of within-trial differences in glycometabolic balance [130].
In a systematic review and meta-analysis about diabetes medications as monotherapy or metformin-based combination treatment, Maruthur et al. demonstrated that major hypoglycemic risk and cardiovascular mortality were lower for metformin versus sulfonylureas and that reductions in glycated hemoglobin values were similar across monotherapies and metformin-based combinations, except for DPP-4 inhibitors, which showed a smaller effect. Metformin, DPP-4 inhibitors, and GLP-1 receptor agonists were similar in reducing or maintaining body weight, which increased with sulfonylureas, thiazolididinediones, and insulin (differences up to 5 kg). As expected, genital infections were increased with glifozins, while gastrointestinal adverse events were highest with metformin and GLP-1 receptor agonists [131].
Furthermore, it is also important considering the pleiotropic actions of metformin and the worsening of neuropathic symptoms in subjects with vitamin B12 deficiency due to the chronic assumption of biguanide, so periodic testing of vitamin B12 is recommended [132,133].
Several institutional guidelines do not suggest metformin as a first-line treatment in secondary cardiovascular prevention, lacking protective impact on major adverse cardiovascular events, cardiovascular death, myocardial infarction, heart failure, and stroke, as demonstrated in an umbrella review of a group from Sun Yat-sen University [134].
4.1.2. Pre-Diabetes
The main projects aimed at analyzing subjects at very high risk of developing diabetes were the Diabetes Prevention Program (DPP) at the end of the last century (1996–2001) and later the DPP Outcome Study (DPPOS, 2002–2013). These trials demonstrated that intensive lifestyle treatment and biguanide therapy were favorable vs. placebo in preventing the onset of diabetes (7.0%, 5.7%, and 5.2% per year for placebo, metformin, and lifestyle, respectively) and 27% and 18% lower for lifestyle and metformin vs. placebo, respectively [135].
In a very recent review, Patel and colleagues—in a total sample size of 30,474 subjects—showed the effectiveness of metformin in diabetes prevention in terms of a reduction in the risk of progressing to type 2 diabetes mellitus in prediabetic individuals receiving the drug (pooled RR 0.58, indicating a 42% lower risk [136].
4.1.3. Type 1 Diabetes
In the Standards of Care by the American Diabetes Association (ADA), metformin is also considered in paragraph “Non insulin treatment for type 1 diabetes” [137]. Meng et al. evaluated the effect of metformin in 1183 subjects with type 1 diabetes, demonstrating that metformin was associated with reductions in BMI (−1.14, 95% CI −2.05 to −0.24, p = 0.01), insulin requirements (−0.47, 95% CI −0.70 to −0.23, p = 0.0001), total cholesterol (−0.23, 95% CI −0.34 to −0.12, p < 0.0001), and low-density lipoprotein cholesterol (−0.20, 95% CI −0.29 to −0.11, p < 0.0001) in type 1 diabetic patients. No clear evidence indicated that metformin improved HbA1c, triglyceride, or high-density lipoprotein cholesterol levels [138]. The REMOVAL study focused on cardiovascular and metabolic effects of metformin in adults aged 40 years and older with type 1 diabetes of at least 5 years’ duration, demonstrating that progression of mean carotid intima-media thickness (IMT) was not significantly reduced with metformin, and that glycated hemoglobin was reduced on average over 3 years by metformin, but this was accounted for by a reduction at the 3-month timepoint that was not sustained thereafter. Bodyweight and LDL cholesterol were reduced with metformin over 3 years of treatment, and eGFR was increased. Finally, the insulin requirement was not reduced on average over 3 years [139].
4.2. Clinical Studies on Tumoral Conditions (“Anti-Proliferative Face”)
The promising in vitro and in vivo evidence regarding the antiproliferative activity of metformin has provided the rationale for the conduct of more than 130 clinical studies included up-to-date in the registry of clinicaltrials.gov. The aim of our paper is to focus on phase III clinical trials and major phase II trials when phase III is absent. The trials to evaluate metformin in oncology were prevalently performed in colorectal, breast, endometrial, and prostate cancer. Despite the increasing number of clinical studies, the data available concerning the potential antitumor effect of biguanide remain limited and inconsistent; therefore, more robust and large-scale trials are warranted to further investigate metformin administration in the neoplastic setting (Table 2).
Colorectal Cancer (CRC)
The antiproliferative efficacy of metformin explored in the therapeutic strategy for colorectal cancer (CRC) is still controversial. In a few prospective studies, the potential chemopreventive role of biguanide in CRC carcinogenesis was documented [142]. Hosono et al. evidenced that low-dose metformin treatment (250 mg daily) led to the inhibition of colonic epithelial proliferation and of colorectal aberrant crypt foci (ACF) in non-diabetic patients with ACF [160]. Furthermore in a phase III, multicenter, double-blind, and placebo-controlled RCT, metformin (250 mg daily)—administered for 1 year—was associated with a significantly lower prevalence of metachronous adenomas or polyps in non-diabetic patients at high risk of CRA relapse after polypectomy [161].
A similar chemopreventive effect was seen in a particular setting of patients with an increased risk of colonic polyps: acromegalic patients. In a recent exploratory cross-sectional study [162], we evaluated the prevalence of colonic polyps in acromegalic patients treated or not with metformin and explored its possible protective role against the development of colon polyps. We subdivided our cohort into patients with and without polyps. Only 24% of subjects with polyps were under metformin vs. 57% of patients without polyps, data confirmed by multivariate analysis (OR 0.224, 95% CI 0.065–0.770, p = 0.01). In our study, metformin was used as an antidiabetic treatment, so this finding could suggest that metformin therapy may counterbalance the dual risk factors of diabetes and acromegaly. These data suggest a potential protective role for metformin in a subset of individuals affected by this specific endocrine disease.
On the other hand, as usual in the scientific literature, other experiences yield different results. A phase IIa study involving non-diabetic and obese (BMI ≥ 30 kg/m2) subjects with a recent history of CRA undergoing metformin therapy 1000 mg twice daily for 12 weeks showed no reduction in rectal tissue pS6Ser235/236 or Ki67 immunostaining levels [163]. Similarly, in individuals already affected by CRC, the impact of the biguanide is still unclear and warrants further exploration. In a retrospective study conducted by Han et al. in 232 patients with rectal cancer who underwent curative resection after preoperative chemo-radiotherapy (CCRT), neoadjuvant metformin administration before CCRT resulted as a relevant factor in predicting tumor downstaging and good response rates of tumor regression grade [164]. Moreover, in a phase II trial, Miranda et al. described an overall modest activity of metformin 850 mg twice daily added to 5-fluorouralcil (5-FU) in 50 subjects with refractory CRC. In particular, 11 (22%) patients met the primary endpoint, which was disease control rate at 8 weeks, and among all patients, those with BMI ≥ 30 kg/m2 appeared to benefit more from the combined therapy. The median Progression Free Survival (mPFS) was 2 months and the median Overall Survival (mOS) was 7.9 months [140]. Less promising findings were detected in a phase II study by Akce et al., who documented that in patients with refractory microsatellite stable metastatic CRC undergoing combined treatment of metformin and nivolumab, no objective response was observed, hence the trial did not proceed with further enrollment; mOS and mPFS were respectively 5.1 and 2.3 months [141]. Accordingly, in a sub-study of the TOSCA trial involving individuals with high-risk stage II or stage III colon cancer undergoing 3 months versus 6 months of fluoropyrimidine–oxaliplatin adjuvant therapy, the addition of metformin impacted neither the OS nor the relapse-free survival (RFS), regardless of its dosage [142]. Similarly, no correlation between the biguanide and the survival outcomes, including OS, disease-free survival (DFS), and time to recurrence (TTR), was identified by Singh et al. in the setting of the phase III N0147 study involving 1958 patients with stage III colon cancer (CC) receiving adjuvant chemotherapy [165]. At present, there are two ongoing phase III RCTs enrolling individuals affected by CRC. In particular, Abdelhafeez et al. are investigating in stage IV CC the effect of metformin combined with the standard therapy FOLFOX/XELOX in comparison with their counterparts receiving only FOLFOX/XELOX. The primary outcomes concern the Disease Control Rate (DCR) and PFS [166]. Kim et al. are also conducting an open-label RCT to examine the impact of adjunctive metformin in patients with recurrent stage II high-risk and stage III colorectal cancer who have already undergone surgery and/or neo-adjuvant chemoradiation. The primary endpoint is the comparison of the 3-year DFS between metformin and non-metformin cohorts [167].
5. Breast Cancer (BC)
The potential antineoplastic role of metformin explored in the setting of both neo- and adjuvant therapy in patients affected by breast cancer is still unclear. In the phase III, double-blind and placebo-controlled RCT conducted in 3649 non-diabetic patients with BC, the addition of metformin to standard adjuvant therapy did not lead to a significant improvement in invasive disease-free survival [143]. Similarly, no significant survival benefits were detected in terms of PFS or response rate (RR) related to the combination of biguanide and chemotherapy in a study involving 107 non-diabetic patients with metastatic BC [144]. However, slightly higher OS and PFS, but with no statistical significance, were associated with metformin and adjuvant chemotherapy in the phase II trial of Salah. et al., enrolling 50 individuals with stage IV BC. The radiological response, instead, resulted significantly better in the metformin cohort in comparison to the placebo one [145]. Instead, more encouraging results were observed when metformin was administered in the neoadjuvant setting. In fact, in the METTEN phase II trial involving 79 individuals affected by HER2-positive BC bearing the rs11212617 C allele, metformin combined with neoadjuvant chemotherapy (anthracycline/taxane-based regimens) and ERBB2-targeted therapy (i.e., trastuzumab) was associated with a higher pathological complete response (pCR) compared to the non-metformin counterparts (81.2% vs. 35.3%, respectively) [146]. Moreover, Othman et al., in a phase II/III RCT, placebo-controlled study, involving 140 subjects with invasive non-metastatic BC, demonstrated that positive Her2 or negative estrogen receptor (ER) status seemed to predict pCR in the metformin-treated arm. In particular, ER-negative BC metformin therapy presented a significantly higher pCR compared to the control group (63.2% vs. 22.2%, respectively, p = 0.02). In HER-2-positive BC, a greater pCR was documented in the metformin arm, but no statistical significance was reached [147]. Similarly, Azazy et al. performed a phase II randomized and placebo-controlled trial in 60 non-diabetic patients with stage II–III BC and documented a higher pCR in the group receiving metformin (850 mg b.i.d.) and neoadjuvant chemotherapy without reaching any significance (p = 0.09) [148]. It is also documented that metformin has a positive impact when combined with neoadjuvant chemotherapy on the pCR rate, particularly in individuals with stage II-III triple-positive BC without diabetes and with a BMI ≥ 25 kg/m2 [149]. Discordantly, no improvement of pCR in patients affected by BC and metabolic abnormalities was observed in the phase II NeoMET study in the arm receiving both the biguanide and neoadjuvant chemotherapy (docetaxel, epirubicin, and cyclophosphamide) [150]. Also in the setting of metastatic or locally advanced, unresectable BC metformin treatment in conjunction with chemotherapy or hormone therapy tested in several phase II studies failed to present a relevant impact on the survival outcomes [151,152,153]. In conclusion, the anticancerogenic effect of biguanide in breast cancer remains controversial and warrants further investigation in robust RCTs. An ongoing phase III RCT is currently exploring the eventual role of metformin in preventing BC development in patients affected by atypical hyperplasia or in situ breast cancer. The results are going to be published in the forthcoming years and could also provide information regarding the potential chemopreventive activity of metformin [168].
6. Prostate Cancer
In prostate cancer, evidence shows inconsistent findings regarding the antitumor role of biguanide. In a meta-analysis comprising three phase III, double-blind, and place-controlled RCTs (AFFIRM, PREVAIL, and PROSPER) aimed at studying enzalutamide in castration-resistant prostate cancer (CRPC) patients, metformin treatment failed to present a significant impact on the survival outcomes [169]. Similarly, no benefits in terms of disease control rate (DCR) and OS were observed in the metformin-treated cohort in combination with enzalutamide compared to enzalutamide alone in SAKK 08/14, a phase II multicenter RCT involving 169 subjects with metastatic CRPC [154]. Also in TAXOMET, a phase II multicenter study comparing docetaxel plus metformin versus docetaxel plus placebo in metastatic CRPC, no meaningful benefits in terms of OS, PFS, ORR, and PSA response rate were documented [155]. Discordantly, a significantly higher prostate cancer-free survival (29 months vs. 20 months, p = 0.01), especially in individuals with high-risk localized disease or metastatic low tumor volume disease, was observed in MANSMED, a phase II RCT enrolling 124 patients with CRPC in the metformin plus androgen-deprivation therapy (ADT) arm versus ADT only [156]. Due to the insufficient data available, some randomized trials are currently ongoing. The MAST study, a phase III, double-blind, and placebo-controlled RCT, is evaluating the eventual impact of metformin in reducing disease progression in men affected by low-risk, localized prostate cancer on expectant management [170]. Another active study is STAMPEDE, a multi-arm and multi-stage phase II/III RCT already involving 11992 participants, aimed at exploring several therapeutic strategies, including metformin, in high-risk locally advanced and metastatic hormone-naïve prostate cancer [171]. In the forthcoming future, we are going to publish the findings of METAL, a phase IV placebo-controlled RCT finalized to investigate the role of biguanide in a neoadjuvant setting in early-stage prostate cancer patients [172].
7. Endometrial Cancer (EC)
There is limited data provided by prospective clinical trials regarding the potential antitumor effect of metformin in endometrial cancer. PREMIUM, a multi-center, placebo-controlled phase III RCT, conducted in 88 patients with atypical hyperplasia or endometrioid endometrial cancer, showed that neoadjuvant treatment with metformin (850 mg daily for 3 days and twice daily thereafter) for 1 to 5 weeks did not present a favorable impact on tumor proliferation. No differences were detected regarding the immunohistochemical expression of Ki-67, neither of the markers of PI3K-Akt-mTOR nor the insulin signaling pathway [157]. Conversely, Mitsuhashi et al., in a phase II, single-arm study enrolling 17 women with atypical endometrial hyperplasia (AEH) and 19 with EC, documented that metformin (750–2250 mg/day) was beneficial in preventing disease recurrence after medroxyprogesterone acetate (MPA) administration for fertility-sparing therapy, showing an 89% 3-year relapse-free survival (RFS) rate [158]. Such findings have provided the rationale for the commencement in 2019 of the FELICIA trial, a randomized phase IIb study, finalized particularly to investigate the adequate dose of metformin in addition to MPA in AEH and EC patients and identify the 3-year RFS rate. The trial results are still to be published [173]. In another ongoing phase III, randomized and placebo-controlled study, the role of biguanide is also being explored in the chemoprevention setting in obese (BMI ≥ 30 kg/m2) and hyperinsulinemic women affected by EC [174]. Other currently active trials are evaluating, in advanced or recurrent EC, the impact of metformin combined with everolimus and letrozole in a single-arm study [175], as well as metformin combined with paclitaxel plus carboplatin in a phase II/III placebo-controlled RCT [176].
8. Other Malignancies
Some interesting data on the antineoplastic efficacy of metformin derive from an Italian multicenter retrospective study on neuroendocrine tumors [177]. In this trial (PRIME-NET Study), the authors demonstrated a significantly longer PFS in diabetic patients with pancreatic neuroendocrine tumors (panNET) treated with metformin, everolimus, and/or somatostatin analogs, compared to diabetic patients with panNET treated with the same oncological therapy but with another antidiabetic agent instead of metformin. The PFS in the first group of diabetic patients with panNET was significantly longer, even compared to those of non-diabetic patients with panNET treated in the same manner, but without metformin. Clinical prospective data about the antineoplastic effect of metformin on other malignancies is still scarce and inconclusive. Regarding neuroendocrine neoplasms (NENs), METNET, a phase II single-arm trial, showed a modest antineoplastic activity of the biguanide (850 mg twice daily) in progressive metastatic well-differentiated NENs of gastroenteropancreatic (GEP) or pulmonary origin. In this study, 46% of the patients presented with DCR at 6 months and a mPFS of 6.3 months; however, this study enrolled only 28 subjects [159]. In consideration of the beneficial activity of metformin when added to everolimus and/or somatostatin analogs in NENs patients with diabetes, as evidenced by the preliminary findings of the PRIME-NET study [177], two phase II studies have been initiated. The MetNET-1 and MetNET-2 trials aim at evaluating the antiproliferative potential of the biguanide combined, respectively, with everolimus plus octreotide LAR in advanced well-differentiated pancreatic NENs or combined with lanreotide in well-differentiated GEP and lung NENs [178,179]. The results are still to be published and could provide the rationale for further exploration of metformin combined with target therapies in this cohort of patients. Preclinical evidence of the antiproliferative activity of metformin in other malignancies has also led to the design of some phase II or III RCTs, aimed at investigating the efficacy of the biguanide in lung cancer when added to tyrosine kinase inhibitors [180], in melanoma when combined with dacarbazine or with pembrolizumab [181,182], in hepatocellular carcinoma when administered with sorafenib [183], as well as in pancreatic cancer when combined with chemotherapeutic agents [184,185].
9. Conclusions and Future Research Directions
Metformin is an old drug with multiple target organs. Over the years, different mechanisms of action, such as the subcellular mechanisms involved in metabolism in cellular and tissue growth, not previously known, have been highlighted, arousing interest about the possibility of using this molecule in fields other than the one for which it is commonly used (T2DM). In addition to confirming the safety and effectiveness of metformin in reducing hba1c in the treatment of T2DM, this review confirms its role in situations where prediabetes are able to prevent the evolution of T2DM. In this last context, in our opinion, it should also be considered what it means to mask an evolution to T2DM and to establish the timing of complications checks and targets of other metabolic or clinical parameters (for example, blood pressure), which would otherwise be defined in the case of an evolution to T2DM. Data are not enough to allow important conclusions about the use of metformin in T1DM, where most likely the patient’s constitutional habit, beyond insulin deficiency, makes the difference in the sensitivity of organs to the response to metformin.
As regards the “antiproliferative face” of metformin, scientific knowledge to date suffers from a contrast between the encouraging in vitro results and the still inconclusive in vivo findings of some trials.
The most promising areas appear to be colon and breast cancers, although some very encouraging data derive from neuroendocrine neoplasms, though from retrospective data, on a large series of cases.
Therefore, further, more rigorous observational studies, or RCTs, should be designed properly, aimed at exploring the potential benefits of the biguanide in specific population subgroups accurately selected according to glycemic and metabolic status, cancer histological features, molecular profiling, and staging.
Considering the current data globally, it seems that the use of metformin in RCTs for a short period has not led to clinical benefits, while in some more prolonged experiences or in retrospective studies, in which patients had been taking the drug for other reasons for several years, the data are more suggestive of a certain benefit.
However, we have probably also considered this aspect, the duration of the patient’s exposure to the drug metformin, in order to evaluate the potential antiproliferative effect and its effectiveness.
Hence, proper data analysis methods should be implemented to avoid time-related biases that could have erroneously led to beneficial effects of metformin in cancer prevention and treatment in the non-randomized studies conducted since 2005, as highlighted by Hoi Yun Yu et al. [186]. In particular, several observational studies have been affected by the immortal time bias, which is defined as the time between cohort entry and metformin initiation, which could amplify the potential benefits of metformin by extending the exposure time to the biguanide and the survival time of the patient [186]. Therefore, a careful definition of drug exposure is another fundamental criterion for providing robust and accurate evidence.
The most interesting role of metformin could be its preventive impact against the development of neoplasia in populations at increased risk, rather than its implementation as an antiproliferative therapy in already diagnosed neoplasia.
As reported by Lord and Harris in a recent perspective article, metformin and its efficacy in cancer prevention have been underexplored in prospective studies [187].
Some data in the literature [161,162] provide a strong rationale for testing this drug in selected groups of patients, for example, acromegalic, obese patients, or insulin-resistant subjects [187].
Given the difficulty in obtaining results in the cancer prevention aspect, because the studies would require very large populations and very long follow-up, Lord and Harris emphasize that an attempt should instead be made to study the preventive power of metformin in those populations who have a higher risk of cancer, such as in some genetic syndromes or metabolic conditions [187].
An interesting translational study published a few years ago in the Journal of Clinical Investigation demonstrated an advantage in terms of increased cancer-free survival in a mouse model of Li–Fraumeni syndrome treated with metformin [188].
This effect was determined by the inhibition of mitochondrial respiration exerted by metformin and occurred in the same way both in the murine model and in vivo in patients affected by this syndrome. The authors therefore concluded that, having demonstrated that metformin was able to inhibit mitochondrial respiration in humans as well, it could be used to prevent the onset of cancer in this particular population of patients with Li–Fraumeni syndrome [188].
These data are also suggested by the literature available so far and will certainly be clarified in the future when the results of several currently ongoing studies are published [166,167,168,172,178,179].
For the future, instead, we should expect to find metformin increasingly under the magnifying glass of many authors as a possible association with immunotherapies. In fact, despite its high efficacy in many cancers, immunotherapy could be limited by the modulation of the tumor immune microenvironment.
A very recent review shows how metformin can be an important “booster” for immunotherapy, according to some preclinical studies (in vitro and animal models) and a few phase II studies on human tumor specimens derived from patients pretreated or not with metformin [189,190,191,192].
Some evidence from these studies shows how metformin, through mechanisms not yet fully elucidated, can modulate the tumor immune microenvironment, thus enhancing the response to immunotherapy.
In summary, we do not yet have a conclusive vision of the potential applications of metformin in the oncology field. At this moment, the evidence about its effectiveness in the treatment of cancers is disappointing; however, the data regarding its role in preventing cancer are intriguing and worthy of further exploration with ongoing trials and others that may clarify the “anti-neoplastic face” of metformin in the future.
Author Contributions
Conceptualization: U.G. and M.A.; Methodology: U.G. and M.B.; Figures and Tables: U.G., M.B. and I.H.; Writing—Original Draft Preparation: U.G., M.A., M.B., C.T. and I.H.; Writing—Review and Editing: U.G. and M.A.; Supervision: D.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in this article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Naso, P.O. Fasti I. In Ovidio—Opere; UTET: Torino, Italy, 1999; Volume IV, p. 104. [Google Scholar]
- Hill, J. The Vegetable System. Or the Internal Structure and the Life of Plants; Their Parts, and Nourishment, Explained; Their Classes, Orders, Genera, and Species, Ascertained, and Described; in a Methods Altogether New: Comprehending an Artificial Index, and a Natural System. With Figures of All the Plants; Designed and Engraved by the Author. The Whole from Nature Only; Containing Plants and Four-Petal’d Irregular Flowers; Pater-Noster-Row: London, UK, 1772; Volume 21, p. 54. [Google Scholar]
- Hadden, D.R. Goat’s rue—French lilac—Italian fitch—Spanish sainfoin: Gallega officinalis and metformin: The Edinburgh connection. J. R. Coll. Physicians Edinb. 2005, 35, 258–260. [Google Scholar]
- Pasik, C. Diabetes and the biguanides: The mystery of each. In Glucophage: Serving Diabetology for 40 Years; Pasik, C., Ed.; Groupe Lipha: Lyon, France, 1997; p. 79. [Google Scholar]
- Rathke, B. Uber Biguanid. Ber. Dtsch. Chem. Ges. 1879, 12, 776–784. (In German) [Google Scholar] [CrossRef]
- Werner, E.A.; Bell, J. The preparation of methylguanidine, and of ββ-dimethylguanidine by the interaction of dicyandiamide, and methylammonium and dimethylammonium chlorides respectively. J. Chem. Soc. Trans. 1922, 121, 1790–1794. [Google Scholar] [CrossRef]
- Slotta, K.H.; Tschesche, R. Uber Biguanide, II.: Die blutzuckersenkende Wirkung der Biguanides. Ber. Dtsch. Chem. Ges. 1929, 62, 1398–1405. (In German) [Google Scholar] [CrossRef]
- Curd, F.H.S.; Davey, D.G.; Rose, F.L. Studies on synthetic antimalarial drugs. Some biguanide derivatives as new types of antimalarial substances with both therapeutic and causal prophylactic activity. Ann. Trop. Med. Parasitol. 1945, 39, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.K.; Anderson, R.C. The toxicity and general pharmacology of N1-p-chlorophenyl-N5-isopropyl biguanide. J. Pharmacol. Exp. Ther. 1947, 91, 157–160. [Google Scholar]
- Pasik, C. Jean Sterne: A passion for research. In Glucophage: Serving Diabetology for 40 Years; Pasik, C., Ed.; Groupe Lipha: Lyon, France, 1997; pp. 29–31. [Google Scholar]
- Sterne, J. Du nouveau dans les antidiabétiques. La NN dimethylamine guanyl guanidine (N.N.D.G.). Maroc. Med. 1957, 36, 1295–1296. (In French) [Google Scholar]
- Sterne, J. Blood sugar-lowering effect of 1,1-dimethylbiguanide. Therapie 1958, 13, 650–659. (In French) [Google Scholar] [PubMed]
- Sterne, J. Treatment of diabetes mellitus with N,N-dimethylguanylguanidine (LA. 6023, glucophage). Therapie 1959, 14, 625–630. (In French) [Google Scholar]
- Sterne, J. Report on 5-years’ experience with dimethylbiguanide (metformin, glucophage) in diabetic therapy. Wien. Med. Wochenschr. 1963, 113, 599–602. (In German) [Google Scholar]
- Sterne, J.; Hirsch, C. Experimental basis for combined treatment of diabetes with the biguanide-sulfonamide association. Diabete 1964, 12, 171–175. (In French) [Google Scholar] [PubMed]
- Sterne, J. Mechanism of action of antidiabetic biguanides. Presse Med. 1964, 72, 17–19. (In French) [Google Scholar]
- Sterne, J. Pharmacology and mode of action of the hypoglycemic guanidine derivatives. In Oral Hypoglycemic Agents; Campbell, G.D., Ed.; Academic Press: London, UK, 1969; pp. 193–245. [Google Scholar]
- Sterne, J.; Duval, D.; Junien, J.L. Aspects of pharmacology and mechanisms of action. In Metformin: Current Aspects and Future Developments; Cudwoth, A.G., Ed.; Research and Clinical Forums: Tunbridge Wells, UK, 1979; Volume 1, pp. 13–20. [Google Scholar]
- Schäfer, G. Biguanides: A review of history, pharmacodynamics and therapy. Diabete Metab. 1983, 9, 148–163. [Google Scholar] [PubMed]
- Hermann, L.S. Metformin: A review of its pharmacological properties and therapeutic use. Diabete Metab. 1979, 5, 233–245. [Google Scholar] [PubMed]
- Apampa, B. Pharmacology and safe prescribing of metformin. Nurse Prescr. 2012, 10, 597–602. [Google Scholar] [CrossRef]
- Glossmann, H.H.; Lutz, O.M. Pharmacology of metformin—An update. Eur. J. Pharmacol. 2019, 865, 172782. [Google Scholar] [CrossRef] [PubMed]
- Jeong, Y.-S.; Jusko, W.J. Meta-assessment of metformin absorption and disposition pharmacokinetics in nine species. Pharmaceuticals 2021, 14, 545. [Google Scholar] [CrossRef]
- Scheen, A.J. Clinical pharmacokinetics of metformin. Clin. Pharmacokinet. 1996, 30, 359–371. [Google Scholar] [CrossRef] [PubMed]
- Graham, G.G.; Punt, J.; Arora, M.; Day, R.O.; Doogue, M.P.; Duong, J.K.; Furlong, T.J.; Greenfield, J.R.; Greenup, L.C.; Kirkpatrick, C.M.; et al. Clinical pharmacokinetics of metformin. Clin. Pharmacokinet. 2011, 50, 81–98. [Google Scholar] [CrossRef]
- Song, R. Mechanism of metformin: A tale of two sites. Diabetes Care 2016, 39, 187–189. [Google Scholar] [CrossRef]
- Chan, P.; Shao, L.; Tomlinson, B.; Zhang, Y.; Liu, Z.M. Metformin transporter pharmacogenomics: Insights into drug disposition—Where are we now? Expert. Opin. Drug Metab. Toxicol. 2018, 14, 1149–1159. [Google Scholar] [CrossRef] [PubMed]
- Sundelin, E.; Jensen, J.B.; Jakobsen, S.; Gormsen, L.C.; Jessen, N. Metformin biodistribution: A key to mechanisms of action? J. Clin. Endocrinol. Metab. 2020, 105, dgaa332. [Google Scholar] [CrossRef]
- Owen, M.R.; Doran, E.; Halestrap, A.P. Evidence that metformin exerts its anti-diabetic effects through inhibition of complex 1 of the mitochondrial respiratory chain. Biochem. J. 2000, 348, 607–614. [Google Scholar] [CrossRef]
- El-Mir, M.Y.; Nogueira, V.; Fontaine, E.; Avéret, N.; Rigoulet, M.; Leverve, X. Dimethylbiguanide inhibits cell respiration via an indirect effect targeted on the respiratory chain complex I. J. Biol. Chem. 2000, 275, 223–228. [Google Scholar] [CrossRef]
- He, L.; Wondisford, F.E. Metformin action: Concentrations matter. Cell Metab. 2015, 21, 159–162. [Google Scholar] [CrossRef]
- Gong, L.; Goswami, S.; Giacomini, K.M.; Altman, R.B.; Klein, T.E. Metformin pathways: Pharmacokinetics and pharmacodynamics. Pharmacogenet. Genom. 2012, 22, 820. [Google Scholar] [CrossRef] [PubMed]
- Vecchio, S.; Giampreti, A.; Petrolini, V.; Lonati, D.; Protti, A.; Papa, P. Metformin accumulation: Lactic acidosis and high plasmatic metformin levels in a retrospective case series of 66 patients on chronic therapy. Clin. Toxicol. 2014, 52, 129–135. [Google Scholar] [CrossRef]
- Duong, J.K.; Kumar, S.S.; Kirkpatrick, C.M.; Greenup, L.C.; Arora, M.; Lee, T.C. Population pharmacokinetics of metformin in healthy subjects and patients with type 2 diabetes mellitus: Simulation of doses according to renal function. Clin. Pharmacokinet. 2013, 52, 373–384. [Google Scholar] [CrossRef]
- Foretz, M.; Guigas, B.; Viollet, B. Metformin: Update on mechanisms of action and repurposing potential. Nat. Rev. Endocrinol. 2023, 19, 460–476. [Google Scholar] [CrossRef] [PubMed]
- Gunton, J.E.; Delhanty, P.J.; Takahashi, S.; Baxter, R.C. Metformin rapidly increases insulin receptor activation in human liver and signals preferentially through insulin-receptor substrate-2. J. Clin. Endocrinol. Metab. 2003, 88, 1323–1332. [Google Scholar] [CrossRef]
- Cusi, K.; Consoli, A.; DeFronzo, R.A. Metabolic effects of metformin on glucose and lactate metabolism in noninsulin-dependent diabetes mellitus. J. Clin. Endocrinol. Metab. 1996, 81, 4059–4067. [Google Scholar]
- Hundal, R.S.; Krssak, M.; Dufour, S.; Laurent, D.; Lebon, V.; Chandramouli, V.; Inzucchi, S.E.; Schumann, W.C.; Petersen, K.F.; Landau, B.R.; et al. Mechanism by which metformin reduces glucose production in type 2 diabetes. Diabetes 2000, 49, 2063–2069. [Google Scholar] [CrossRef]
- Argaud, D.; Roth, H.; Wiernsperger, N.; Leverve, X.M. Metformin decreases gluconeogenesis by enhancing the pyruvate kinase flux in isolated rat hepatocytes. Eur. J. Biochem. 1993, 213, 1341–1348. [Google Scholar] [CrossRef] [PubMed]
- Large, V.; Beylot, M. Modifications of citric acid cycle activity and gluconeogenesis in streptozotocin-induced diabetes and effects of metformin. Diabetes 1999, 48, 1251–1257. [Google Scholar] [CrossRef]
- Mithieux, G.; Guignot, L.; Bordet, J.C.; Wiernsperger, N. Intrahepatic mechanisms underlying the effect of metformin in decreasing basal glucose production in rats fed a high-fat diet. Diabetes 2002, 51, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Radziuk, J.; Zhang, Z.; Wiernsperger, N.; Pye, S. Effects of metformin on lactate uptake and gluconeogenesis in the perfused rat liver. Diabetes 1997, 46, 1406–1413. [Google Scholar] [CrossRef]
- Wilcock, C.; Bailey, C.J. Accumulation of metformin by tissues of the normal and diabetic mouse. Xenobiotica 1994, 24, 49–57. [Google Scholar] [CrossRef]
- Tobar, N.; Rocha, G.Z.; Santos, A.; Guadagnini, D.; Assalin, H.B.; Camargo, J.A.; Goncalves, A.E.S.S.; Pallis, F.R.; Oliveira, A.G.; Rocco, S.A.; et al. Metformin acts in the gut and induces gut–liver crosstalk. Proc. Natl. Acad. Sci. USA 2023, 120, e2211933120. [Google Scholar] [CrossRef] [PubMed]
- Bailey, C.J.; Wilcock, C.; Scarpello, J.H. Metformin and the intestine. Diabetologia 2008, 51, 1552–1553. [Google Scholar] [CrossRef]
- Proctor, W.R.; Ming, X.; Bourdet, D.; Han, T.K.; Everett, R.S.; Thakker, D.R. Why does the intestine lack basolateral eflux transporters for cationic compounds? A provocative hypothesis. J. Pharm. Sci. 2016, 105, 484–496. [Google Scholar] [CrossRef]
- Wu, T.; Xie, C.; Wu, H.; Jones, K.L.; Horowitz, M.; Rayner, C.K. Metformin reduces the rate of small intestinal glucose absorption in type 2 diabetes. Diabetes Obes. Metab. 2017, 19, 290–293. [Google Scholar] [CrossRef] [PubMed]
- Bailey, C.J. Metformin and intestinal glucose handling. Diabetes Metab. Rev. 1995, 11, S23–S32. [Google Scholar] [CrossRef] [PubMed]
- Zubiaga, L.; Brinad, O.; Auger, F.; Touche, V.; Hubert, T.; Thvenet, J.; Marciniak, C.; Quenon, Q.; Bonner, C.; Peschard, S.; et al. Oral metformin transiently lowers post-prandial glucose response by reducing the apical expression of sodium-glucose co-transporter 1 in enterocytes. iScience 2023, 26, 106057. [Google Scholar] [CrossRef] [PubMed]
- Maida, A.; Lamont, B.J.; Cao, X.; Drucker, D.J. Metformin regulates the incretin receptor axis via a pathway dependent on peroxisome proliferator-activated receptor-α in mice. Diabetologia 2011, 54, 339–349. [Google Scholar] [CrossRef]
- Borg, M.J.; Boung, M.; Grivell, J.; Sun, Z.; Jones, K.L.; Horowitz, M.; Rayner, C.K.; Wu, T. Comparative effects of proximal and distal small intestinal administration of metformin on plasma glucose and glucagon-like peptide-1, and gastric emptying after oral glucose, in type 2 diabetes. Diabetes Obes. Metab. 2019, 21, 640–647. [Google Scholar] [CrossRef]
- Chondronikola, M.; Volpi, E.; Børsheim, E.; Porter, C.; Annamalai, P.; Enerbäck, S.; Lidell, M.E.; Saraf, M.K.; Labbe, S.M.; Hurren, N.M.; et al. Brown adipose tissue improves whole-body glucose homeostasis and insulin sensitivity in humans. Diabetes 2014, 63, 4089–4099. [Google Scholar] [CrossRef]
- Sponton, C.H.; Hosono, T.; Taura, J.; Jedrychowski, M.P.; Yoneshiro, T.; Wang, Q.; Takahashi, M.; Matsui, Y.; Ikeda, K.; Oguri, Y.; et al. The regulation of glucose and lipid homeostasis via PLTP as a mediator of BAT–liver communication. EMBO Rep. 2020, 21, e49828. [Google Scholar] [CrossRef] [PubMed]
- Breining, P.; Jensen, J.B.; Sundelin, E.I.; Gormsen, L.C.; Jakobsen, S.; Busk, M.; Rolighed, L.; Bross, P.; Fernandez-Guerra, P.; Markussen, L.K.; et al. Metformin targets brown adipose tissue in vivo and reduces oxygen consumption in vitro. Diabetes Obes. Metab. 2018, 20, 2264–2273. [Google Scholar] [CrossRef] [PubMed]
- Geerling, J.J.; Boon, M.R.; Van der Zon, G.C.; Van den Berg, S.A.; Van den Hoek, A.M.; Lombès, M.; Princen, H.M.G.; Havekes, L.M.; Rensen, P.C.N.; Guigas, B. Metformin lowers plasma triglycerides by promoting VLDLtriglyceride clearance by brown adipose tissue in mice. Diabetes 2014, 63, 880–891. [Google Scholar] [CrossRef]
- Tokubuchi, I.; Tajiri, Y.; Iwata, S.; Hara, K.; Wada, N.; Hashinaga, T.; Nakayama, H.; Mifune, H.; Yamada, K. Beneficial effects of metformin on energy metabolism and visceral fat volume through a possible mechanism of fatty acid oxidation in human subjects and rats. PLoS ONE 2017, 12, e0171293. [Google Scholar] [CrossRef]
- Riedmaier, A.E.; Nies, A.T.; Schaeffeler, E.; Schwab, M. Organic anion transporters and their implications in pharmacotherapy. Pharmacol. Rev. 2012, 64, 421–449. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.C.; Sobti, M.; Quinn, A.; Smith, N.J.; Brown Simon, H.J.; Vanderberg, J.I.; Ryan, R.M.; O’Mara, M.L.; Stewart, A.G. Structural basis of promiscuous substrate transport by Organic Cation Transporter 1. Nat. Commun. 2023, 14, 6374. [Google Scholar] [CrossRef] [PubMed]
- Dujic, T.; Zhou, K.; Donnelly, L.A.; Tavendale, R.; Palmer, C.N.A.; Pearson, E.R. Association of organic cation transporter 1 with intolerance to metformin in type 2 diabetes: A GoDARTS study. Diabetes 2015, 64, 1786–1793. [Google Scholar] [CrossRef] [PubMed]
- Shu, Y.; Sheardown, S.A.; Brown, C.; Owen, R.P.; Zhang, S.; Castro, R.A.; Ianculescu, A.G.; Yue, L.; Lo, J.C.; Burchard, E.G.; et al. Effect of genetic variation in the organic cation transporter 1 (OCT1) on metformin action. J. Clin. Investig. 2007, 117, 1422–1431. [Google Scholar] [CrossRef]
- Kawoosa, F.; Zafar, A.S.; Masoodi, S.R.; Amin, A.; Rasool, R.; Fazili, K.M.; Dar, A.H.; Lone, A.; Bashir, S.U.I. Role of human organic cation transporter-1 (OCT1/SLC22A1) in modulating the response to metformin in patients with type 2 diabetes. BMC Endocr. Disord. 2022, 22, 140. [Google Scholar] [CrossRef]
- Bridges, H.R.; Blaza, J.N.; Yin, Z.; Chung, I.; Pollak, M.N.; Hirst, J. Structural basis of mammalian respiratory complex I inhibition by medicinal biguanides. Science 2023, 379, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Bridges, H.R.; Jones, A.J.; Pollak, M.N.; Hirst, J. Effects of metformin and other biguanides on oxidative phosphorylation in mitochondria. Biochem. J. 2014, 462, 475–487. [Google Scholar] [CrossRef]
- LaMoia, T.E.; Butrico, G.M.; Kalpage, H.A.; Goedeke, L.; Hubbard, B.T.H.; Vatner, D.F.; Gaspar, R.C.; Zhang, X.M.; Cline, G.W.; Nakahara, K.; et al. Metformin, phenformin, and galegine inhibit complex IV activity and reduce glycerol-derived gluconeogenesis. Proc. Natl. Acad. Sci. USA 2022, 119, e2122287119. [Google Scholar] [CrossRef] [PubMed]
- Madiraju, A.K.; Erion, D.M.; Rahimi, Y.; Zhang, X.M.; Braddock, D.T.; Albright, R.A.; Prigaro, B.J.; Wood, J.L.; Bhanot, S.; MacDonald, M.J.; et al. Metformin suppresses gluconeogenesis by inhibiting mitochondrial glycerophosphate dehydrogenase. Nature 2014, 510, 542–546. [Google Scholar] [CrossRef]
- Ma, T.; Tian, X.; Zhang, B.; Li, M.; Wang, Y.; Yang, C.; Wu, J.; Wei, X.; Qu, Q.; Yu, Y.; et al. Low-dose metformin targets the lysosomal AMPK pathway through PEN2. Nature 2022, 603, 159–165. [Google Scholar] [CrossRef]
- Aljofan, M.; Riethmacher, D. Anticancer activity of metformin: A systematic review of the literature. Future Sci. OA 2019, 5, FSO410. [Google Scholar] [CrossRef]
- Hua, Y.; Zheng, Y.; Yao, Y.; Jia, R.; Ge, S.; Zhuang, A. Metformin and cancer hallmarks: Shedding new lights on therapeutic repurposing. J. Transl. Med. 2023, 21, 403. [Google Scholar] [CrossRef] [PubMed]
- Howell, J.J.; Hellberg, K.; Turner, M.; Talbott, G.; Kolar, M.J.; Ross, D.S.; Hoxhaj, G.; Saghatelian, A.; Shaw, R.J.; Manning, B.D. Metformin Inhibits Hepatic mTORC1 Signaling via Dose-Dependent Mechanisms Involving AMPK and the TSC Complex. Cell Metab. 2017, 25, 463–471. [Google Scholar] [CrossRef]
- Lama-Sherpa, T.D.; Jeong, M.H.; Jewell, J.L. Regulation of mTORC1 by the Rag GTPases. Biochem. Soc. Trans. 2023, 51, 655–664. [Google Scholar] [CrossRef] [PubMed]
- Katiyar, S.; Liu, E.; Knutzen, C.A.; Lang, E.S.; Lombardo, C.R.; Sankar, S.; Toth, J.I.; Petroski, M.D.; Ronai, Z.; Chiang, G.G. REDD1, an inhibitor of mTOR signalling, is regulated by the CUL4A-DDB1 ubiquitin ligase. EMBO Rep. 2009, 10, 866–872. [Google Scholar] [CrossRef] [PubMed]
- Sahra, I.; Regazzetti, C.; Robert, G.; Laurent, K.; Le Marchand-Brustel, Y.; Auberger, P.; Tanti, J.F.; Giorgetti-Peraldi, S.; Bost, F. Metformin, Independent of AMPK, Induces mTOR Inhibition and Cell-Cycle Arrest through REDD1. Cancer Res. 2011, 71, 4366–4372. [Google Scholar] [CrossRef]
- Han, P.; Zhou, J.; Xiang, J.; Liu, Q.; Sun, K. Research progress on the therapeutic effect and mechanism of metformin for lung cancer (Review). Oncol. Rep. 2023, 49, 3. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.-W.; Zhou, N.; Jin, F.; Wang, R.; Zhao, J.-Q. Metformin reduces pancreatic cancer cell proliferation and increases apoptosis through MTOR signaling pathway and its dose-effect relationship. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 5336–5344. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Cui, J.; Wang, H.; Medina, R.; Zhang, S.; Zhang, X.; Zhuang, Z.; Lin, Y. Metformin enhances anti-cancer effects of cisplatin in meningioma through AMPK-mTOR signaling pathways. Mol. Ther.-Oncolytics 2021, 20, 119–131. [Google Scholar] [CrossRef]
- Grimaldi, C.; Chiarini, F.; Tabellini, G. Amp-dependent kinase/mammalian target of rapamycin complex 1 signaling in t-cell acute lymphoblastic leukemia: Therapeutic implications. Leukemia 2012, 26, 91–100. [Google Scholar] [CrossRef]
- Green, A.S.; Chapuis, N.; Maciel, T.T. The lkb1/ampk signaling pathway has tumor suppressor activity in acute myeloid leukemia through the repression of mtor-dependent oncogenic mrna translation. Blood 2010, 116, 4262–4273. [Google Scholar] [CrossRef]
- Rocha, G.Z.; Dias, M.M.; Ropelle, E.R. Metformin amplifies chemotherapy-induced ampk activation and antitumoral growth. Clin. Cancer Res. 2011, 17, 3993–4005. [Google Scholar] [CrossRef]
- Shi, W.Y.; Xiao, D.; Wang, L. Therapeutic metformin/ampk activation blocked lymphoma cell growth via inhibition of mTOR pathway and induction of autophagy. Cell Death Dis. 2012, 3, e275. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.W.; Yi, K.H.; Han, S.K. Therapeutic potential of metformin in papillary thyroid cancer in vitro and in vivo. Mol. Cell. Endocrinol. 2014, 393, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Glaviano, A.; Foo, A.S.C.; Lam, H.Y. PI3K/AKT/mTOR signaling transduction pathway and targeted therapies in cancer. Mol. Cancer 2023, 22, 138. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Xue, D.; Wang, K.; Zhang, F.; Shi, J.; Jia, B.; Yang, D.; Zhang, Q.; Zhang, S.; Jiang, H.; et al. Metformin exerts an antitumor effect by inhibiting bladder cancer cell migration and growth, and promoting apoptosis through the PI3K/AKT/mTOR pathway. BMC Urol. 2022, 22, 79. [Google Scholar] [CrossRef] [PubMed]
- Nozhat, Z.; Mohammadi-Yeganeh, S.; Azizi, F.; Zarkesh, M.; Hedayati, M. Effects of metformin on the PI3K/AKT/FOXO1 pathway in anaplastic thyroid cancer cell lines. DARU 2018, 26, 93–103. [Google Scholar] [CrossRef]
- Tang, J.C.; An, R.; Jiang, Y.Q.; Yang, J. Effects and Mechanisms of Metformin on the Proliferation of Esophageal Cancer Cells In Vitro and In Vivo. Cancer Res. Treat. 2017, 49, 778–789. [Google Scholar] [CrossRef] [PubMed]
- Xia, C.; Liu, C.; He, Z.; Cai, Y.; Chen, J. Metformin inhibits cervical cancer cell proliferation by modulating PI3K/Akt-induced major histocompatibility complex class I-related chain A gene expression. J. Exp. Clin. Cancer Res. 2020, 39, 127. [Google Scholar] [CrossRef]
- Xiao, Q.; Xiao, J.; Liu, J. Metformin suppresses the growth of colorectal cancer by targeting INHBA to inhibit TGF-β/PI3K/AKT signaling transduction. Cell Death Dis. 2022, 13, 202. [Google Scholar] [CrossRef]
- Cui, Y.; Zhou, J.; Rong, F. Combination of metformin and RG7388 enhances inhibition of growth and induction of apoptosis of ovarian cancer cells through the PI3K/AKT/mTOR pathway. Biochem. Biophys. Res. Commun. 2020, 533, 665–671. [Google Scholar] [CrossRef]
- Qiang, P.; Shao, Y.; Sun, Y.-P.; Zhang, J.; Chen, L.-J. Metformin inhibits proliferation and migration of endometrial cancer cells through regulating PI3K/AKT/MDM2 pathway. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 1778–1785. [Google Scholar] [CrossRef]
- Zheng, Z.; Zhu, W.; Yang, B.; Chai, R.; Liu, T.; Li, F.; Ren, G.; Ji, S.; Liu, S.; Li, G. The cotreatment of metformin with flavone synergistically induces apoptosis through inhibition of PI3K/AKT pathway in breast cancer cells. Oncol. Lett. 2018, 15, 5952–5958. [Google Scholar] [CrossRef] [PubMed]
- Sun, R.; Zhai, R.; Ma, C.; Miao, W. Combination of aloin and metformin enhances the antitumor effect by inhibiting the growth and invasion and inducing apoptosis and autophagy in hepatocellular carcinoma through PI3K/AKT/mTOR pathway. Cancer Med. 2020, 9, 1141–1151. [Google Scholar] [CrossRef]
- Iglesias, D.A.; Yates, M.S.; van der Hoeven, D.; Rodkey, T.L.; Zhang, Q.; Co, N.N.; Burzawa, J.; Chigurupati, S.; Celestino, J.; Bowser, J.; et al. Another surprise from Metformin: Novel mechanism of action via K-Ras influences endometrial cancer response to therapy. Mol. Cancer Ther. 2013, 12, 2847–2856. [Google Scholar] [CrossRef]
- Xie, J.; Xia, L.; Xiang, W.; He, W.; Yin, H.; Wang, F.; Gao, T.; Qi, W.; Yang, Z.; Yang, X.; et al. Metformin selectively inhibits metastatic colorectal cancer with the KRAS mutation by intracellular accumulation through silencing MATE1. Proc. Natl. Acad. Sci. USA 2020, 117, 13012–13022. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Guo, F.; Wang, W.; Shi, H.; Li, D.; Wang, Y. K-ras gene mutation as a predictor of cancer cell responsiveness to metformin. Mol. Med. Rep. 2013, 8, 763–768. [Google Scholar] [CrossRef]
- Dong, S.; Zeng, L.; Liu, Z.; Li, R.; Zou, Y.; Li, Z.; Ge, C.; Lai, Z.; Xue, Y.; Yang, J.; et al. NLK functions to maintain proliferation and stemness of NSCLC and is a target of metformin. J. Hematol. Oncol. 2015, 8, 120. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Singh, U.K.; Kini, S.G.; Garg, V.; Agrawal, S.; Tomar, P.K.; Pathak, P.; Chaudhary, A.; Gupta, P.; Malik, A. JNK pathway signaling: A novel and smarter therapeutic targets for various biological diseases. Future Med. Chem. 2015, 7, 2065–2086. [Google Scholar] [CrossRef]
- Li, B.; Zhou, P.; Xu, K.; Chen, T.; Jiao, J.; Wei, H.; Yang, X.; Xu, W.; Wan, W.; Xiao, J. Metformin induces cell cycle arrest, apoptosis and autophagy through ROS/JNK signaling pathway in human osteosarcoma. Int. J. Biol. Sci. 2020, 16, 74–84. [Google Scholar] [CrossRef]
- Wu, N.; Gu, C.; Gu, H.; Hu, H.; Han, Y.; Li, Q. Metformin induces apoptosis of lung cancer cells through activating JNK/p38 MAPK pathway and GADD153. Neoplasma 2011, 58, 482–490. [Google Scholar] [CrossRef]
- Lu, C.; Chiang, J.; Tsai, F.; Hsu, Y.; Juan, Y.; Yang, J.; Chiu, H. Metformin triggers the intrinsic apoptotic response in human AGS gastric adenocarcinoma cells by activating AMPK and suppressing mTOR/AKT signaling. Int. J. Oncol. 2019, 54, 1271–1281. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Cao, L.; Wang, L.; Liu, L.; Huang, Y.; Gong, X. Metformin Inhibits Proliferation of Human Thyroid Cancer TPC-1 Cells by Decreasing LRP2 to Suppress the JNK Pathway. OncoTargets Ther. 2020, 13, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Montalto, F.I.; De Amicis, F. Cyclin D1 in Cancer: A Molecular Connection for Cell Cycle Control, Adhesion and Invasion in Tumor and Stroma. Cells 2020, 9, 2648. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Ke, C.; Tang, Q. Metformin promotes autophagy and apoptosis in esophageal squamous cell carcinoma by downregulating Stat3 signaling. Cell Death Dis. 2014, 5, e1088. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.-S.; Wang, S.; Deng, A.; Liu, B.; Edgerton, S.; Lind, S.; Wahdan-Alaswad, R.; Thor, A. Metformin targets Stat3 to inhibit cell growth and induce apoptosis in triple-negative breast cancers. Cell Cycle 2012, 11, 367–376. [Google Scholar] [CrossRef]
- Wallbillich, J.J.; Josyula, S.; Saini, U.; Zingarelli, R.A.; Dorayappan, K.D.P. High Glucose-Mediated STAT3 Activation in Endometrial Cancer Is Inhibited by Metformin: Therapeutic Implications for Endometrial Cancer. PLoS ONE 2017, 12, e0170318. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Huang, M.; Zhang, Q. Metformin Antagonizes Ovarian Cancer Cells Malignancy Through MSLN Mediated IL-6/STAT3 Signaling. Cell Transplant. 2021, 30, 9636897211027819. [Google Scholar] [CrossRef]
- Saengboonmee, C.; Seubwai, W.; Cha’on, U.; Sawanyawisuth, K.; Wongkham, S.; Wongkham, C. Metformin Exerts Antiproliferative and Anti-metastatic Effects Against Cholangiocarcinoma Cells by Targeting STAT3 and NF-ĸB. Anticancer Res. 2017, 37, 115–123. [Google Scholar] [CrossRef]
- Fonseca, N.P.; Fenerich, B.A.; Pereira-Martins, D.A.; Garibaldi, P.M.M.; Coelho-Silva, J.L.; Alves Silva, A.B.; Fernandes, J.C.; Silva, C.L.; Rego, M.E.; Machado-Neto, J.A.; et al. Metformin Inhibits STAT3 and MAPK Signaling through AMPK Activation in CSF3RT618I Cells. Blood 2022, 140 (Suppl. 1), 12153–12154. [Google Scholar] [CrossRef]
- Leidgens, V.; Proske, J.; Rauer, L.; Moeckel, S.; Renner, K.; Bogdahn, U.; Riemenschneider, M.J.; Proescholdt, M.; Vollmann-Zwerenz, A.; Hau, P.; et al. Stattic and metformin inhibit brain tumor initiating cells by reducing STAT3-phosphorylation. Oncotarget 2017, 8, 8250–8263. [Google Scholar] [CrossRef]
- Pan, Q.; Yang, G.-L.; Yang, J.-H.; Lin, S.-L.; Liu, N.; Liu, S.-S.; Liu, M.-Y.; Zhang, L.-H.; Huang, Y.-R.; Shen, R.L.; et al. Metformin can block precancerous progression to invasive tumors of bladder through inhibiting STAT3-mediated signaling pathways. J. Exp. Clin. Cancer Res. 2015, 34, 77. [Google Scholar] [CrossRef] [PubMed]
- Esparza-López, J.; Alvarado-Muñoz, J.F.; Escobar-Arriaga, E. Metformin reverses mesenchymal phenotype of primary breast cancer cells through STAT3/NF-κB pathways. BMC Cancer 2019, 19, 728. [Google Scholar] [CrossRef]
- Bashraheel, S.S.; Kheraldine, H.; Khalaf, S.; Al Moustafa, A.-E. Metformin and HER2-positive breast cancer: Mechanisms and therapeutic implications. Biomed. Pharmacother. 2023, 162, 114676. [Google Scholar] [CrossRef]
- Kim, J.-S.; Kim, M.Y.; Hong, S. Synergistic Effects of Metformin and Trastuzumab on HER2 Positive Gastroesophageal Adenocarcinoma Cells In Vitro and In Vivo. Cancers 2023, 15, 4768. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Kwon, H.; Lee, J.W. Metformin increases survival in hormone receptor-positive, HER2-positive breast cancer patients with diabetes. Breast Cancer Res. 2015, 17, 64. [Google Scholar] [CrossRef] [PubMed]
- Vazquez-Martin, A.; Oliveras-Ferraros, C.; Menendez, J.A. The antidiabetic drug metformin suppresses HER2 (erbB-2) oncoprotein overexpression via inhibition of the mTOR effector p70S6K1 in human breast carcinoma cells. Cell Cycle 2009, 8, 88–96. [Google Scholar] [CrossRef]
- Chen, T.W.; Liang, Y.N.; Feng, D.; Tao, L.Y.; Qi, K.; Zhang, H.Y.; Wang, H.X.; Lin, Q.S.; Kong, H. Metformin inhibits proliferation and promotes apoptosis of HER2 positive breast cancer cells by downregulating HSP90. J. Balk. Union Oncol. 2013, 18, 51–56. [Google Scholar]
- Wang, J.; Li, G.; Wang, Y.; Tang, S.; Sun, X.; Feng, X.; Li, Y.; Bao, G.; Li, P.; Mao, X.; et al. Suppression of tumor angiogenesis by metformin treatment via a mechanism linked to targeting of HER2/HIF-1α/VEGF secretion axis. Oncotarget 2015, 6, 44579–44592. [Google Scholar] [CrossRef]
- Tanaka, K.; Babic, I.; Nathanson, D.; Akhavan, D.; Guo, D.; Gini, B. Oncogenic EGFR signaling activates an mTORC2-NF-kappaB pathway that promotes chemotherapy resistance. Cancer Discov. 2011, 1, 524–538. [Google Scholar] [CrossRef]
- Li, L.; Wang, T.; Hu, M.; Zhang, Y.; Chen, H.; Xu, L. Metformin Overcomes Acquired Resistance to EGFR TKIs in EGFR-Mutant Lung Cancer via AMPK/ERK/NF-κB Signaling Pathway. Front. Oncol. 2020, 10, 1605. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Bian, Y.; Zhang, Y.; Ren, G.; Li, G. Metformin activates AMPK/SIRT1/NF-κB pathway and induces mitochondrial dysfunction to drive caspase3/GSDME-mediated cancer cell pyroptosis. Cell Cycle 2020, 19, 1089–1104. [Google Scholar] [CrossRef] [PubMed]
- Yenmis, G.; Yaprak Sarac, E.; Besli, N.; Soydas, T.; Tastan, C.; Dilek Kancagi, D.; Yilanci, M.; Senol, K.; Karagulle, O.O.; Ekmekci, C.G.; et al. Anti-cancer effect of metformin on the metastasis and invasion of primary breast cancer cells through mediating NF-kB activity. Acta Histochem. 2021, 123, 151709. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Liu, X.; Chen, Z.; Chen, H.; Tan, Y. Metformin Suppressed Tumor Necrosis Factor-α-Induced Epithelial- Mesenchymal Transition in Prostate Cancer by Inactivating the NF-κB Signaling Pathway. Transl. Cancer Res. 2020, 9, 6086–6095. Available online: https://tcr.amegroups.org/article/view/44883 (accessed on 17 October 2023). [CrossRef] [PubMed]
- Xu, S.; Yang, Z.; Jin, P.; Yang, X.; Li, X.; Wei, X.; Wang, Y.; Long, S.; Zhang, T.; Chen, G.; et al. Metformin Suppresses Tumor Progression by Inactivating Stromal Fibroblasts in Ovarian Cancer. Mol. Cancer Ther. 2018, 17, 1291–1302. [Google Scholar] [CrossRef] [PubMed]
- Lei, Y.; Yi, Y.; Liu, Y.; Liu, X.; Keller, E.T.; Qian, C.N.; Zhang, J.; Lu, Y. Metformin targets multiple signaling pathways in cancer. Chin. J. Cancer 2017, 36, 17. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, M.-X.; Wang, H.; Zeng, Z.; Li, X.-M. Metformin Down-regulates Endometrial Carcinoma Cell Secretion of IGF-1 and Expression of IGF-1R. Asian Pac. J. Cancer Prev. 2015, 16, 221–225. [Google Scholar] [CrossRef] [PubMed]
- Xue, L.; Chen, F.; Yue, F.; Camacho, L.; Kothapalli, S.; Wei, G.; Huang, S.; Mo, Q.; Ma, F.; Li, Y.; et al. Metformin and an insulin/IGF-1 receptor inhibitor are synergistic in blocking growth of triple-negative breast cancer. Breast Cancer Res. Treat. 2021, 185, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Esposito, A.I.; Amaro, A.; Angelini, G.; Emionite, L.; Gennari, A.; Indraccolo, S.; Maggi, D.; Marini, C.; Salani, B.; Sambuceti, G.; et al. Abstract 1182: Metformin affects breast cancer cell growth and disturbs an IGF1/insulin related gene network that correlates with breast cancer progression. Cancer Res. 2015, 75 (Suppl. 15), 1182. [Google Scholar] [CrossRef]
- UK Prospective Diabetes Study (UKPDS) Group. Intensive blood-glucose control with sulphonylureas or insulin compared with conventional treatment and risk of complications in patients with type 2 diabetes (UKPDS 33). Lancet 1998, 352, 837–853. [Google Scholar] [CrossRef]
- Home, P.D. Impact of the UKPDS—An overview. Diabet. Med. 2008, 25 (Suppl. 2), 2–8. [Google Scholar] [CrossRef]
- UK Prospective Diabetes Study (UKPDS). VIII. Study design, progress and performance. Diabetologia 1991, 34, 877–890.
- Metropolitan Life Insurance Company. New weight standards for men and women. Stat. Bull. Metrop. Insur. Co. 1959, 40, 1–4. [Google Scholar]
- Holman, R.R.; Paul, S.K.; Bethel, M.A.; Matthews, D.R.; Neil, H.A.W. 10-year follow-up of intensive glucose control in type 2 diabetes. N. Engl. J. Med. 2008, 359, 1577–1589. [Google Scholar] [CrossRef] [PubMed]
- Maruthur, N.M.; Tseng, E.; Hutfless, S.; Wilson, L.M.; Suarez-Cuervo, C.; Berger, Z.; Chu, Y.; Iyoha, E.; Segal, J.B.; Bolen, S. Diabetes Medications as Monotherapy or Metformin-Based Combination Therapy for Type 2 Diabetes: A Systematic Review and Meta-analysis. Ann. Intern. Med. 2016, 164, 740–751. [Google Scholar] [CrossRef] [PubMed]
- Out, M.; Kooy, A.; Lehert, P.; Schalkwijk, C.A.; Stehouwer, C.D.A. Long-term treatment with metformin in type 2 diabetes and methylmalonic acid: Post hoc analysis of a randomized controlled 4.3-year trial. J. Diabetes Complicat. 2018, 32, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Aroda, V.R.; Edelstein, S.L.; Goldberg, R.B.; Knowler, W.C.; Marcovina, S.M.; Orchard, T.J.; Bray, G.A.; Schade, D.S.; Temprosa, M.G.; White, N.H.; et al. Diabetes Prevention Program Research Group. Long-term metformin use and vitamin B12 deficiency in the Diabetes Prevention Program Outcomes Study. J. Clin. Endocrinol. Metab. 2016, 101, 1754–1761. [Google Scholar] [CrossRef]
- Zhu, J.; Yu, X.; Zheng, Y.; Li, J.; Wang, Y.; Lin, Y.; He, Z.; Zhao, W.; Chen, C.; Qiu, K.; et al. Association of glucose-lowering medications with cardiovascular outcomes: An umbrella review and evidence map. Lancet Diabetes Endocrinol. 2020, 8, 192–205. [Google Scholar] [CrossRef]
- Diabetes Prevention Program Research Group. Long-term effects of lifestyle intervention or metformin on diabetes development and microvascular complications over 15-year follow-up: The Diabetes Prevention Program Outcomes Study. Lancet Diabetes Endocrinol. 2015, 3, 866–875. [Google Scholar] [CrossRef]
- Patel, D.; Ayesha, I.E.; Monson, N.R.; Klair, N.; Patel, U.; Saxena, A.; Hamid, P. The Effectiveness of Metformin in Diabetes Prevention: A Systematic Review and Meta-Analysis. Cureus 2023, 15, e46108. [Google Scholar] [CrossRef]
- American Diabetes Association. Pharmacologic Approaches to Glycemic Treatment: Standards of Care in Diabetes—2023. Diabetes Care 2023, 46 (Suppl. 1), S140–S157. [Google Scholar] [CrossRef]
- Meng, H.; Zhang, A.; Liang, Y.; Hao, J.; Zhang, X.; Lu, J. Effect of metformin on glycaemic control in patients with type 1 diabetes: A meta-analysis of randomized controlled trials. Diabetes Metab. Res. Rev. 2018, 34, e2983. [Google Scholar] [CrossRef] [PubMed]
- Petrie, J.R.; Chaturvedi, N.; Ford, I.; Brouwers, M.C.G.J.; Greenlaw, N.; Tillin, T.; Hramiak, I.; Hughes, A.D.; Jenkins, A.J.; Klein, B.E.K.; et al. Cardiovascular and metabolic effects of metformin in patients with type 1 diabetes (REMOVAL): A double-blind, randomised, placebo controlled trial. Lancet Diabetes Endocrinol. 2017, 5, 597–609. [Google Scholar] [CrossRef] [PubMed]
- Miranda, V.C.; Braghiroli, M.I.; Faria, L.D.; Bariani, G.; Alex, A.; Bezerra Neto, J.E.; Capareli, F.C.; Sabbaga, J.; Lobo Dos Santos, J.F.; Hoff, P.M.; et al. Phase 2 Trial of Metformin Combined with 5-Fluorouracil in Patients with Refractory Metastatic Colorectal Cancer. Clin. Color. Cancer 2016, 15, 321–328.e1. [Google Scholar] [CrossRef]
- Akce, M.; Farran, B.; Switchenko, J.M.; Rupji, M.; Kang, S.; Khalil, L.; Ruggieri-Joyce, A.; Olson, B.; Shaib, W.L.; Wu, C.; et al. Phase II trial of nivolumab and metformin in patients with treatment-refractory microsatellite stable metastatic colorectal cancer. J. Immunother. Cancer 2023, 11, e007235. [Google Scholar] [CrossRef] [PubMed]
- Vernieri, C.; Galli, F.; Ferrari, L.; Marchetti, P.; Lonardi, S.; Maiello, E.; Iaffaioli, R.V.; Zampino, M.G.; Zaniboni, A.; De Placido, S.; et al. Impact of Metformin Use and Diabetic Status During Adjuvant Fluoropyrimidine-Oxaliplatin Chemotherapy on the Outcome of Patients with Resected Colon Cancer: A TOSCA Study Subanalysis. Oncologist 2019, 24, 385–393. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, P.J.; Chen, B.E.; Gelmon, K.A.; Whelan, T.J.; Ennis, M.; Lemieux, J.; Ligibel, J.A.; Hershman, D.L.; Mayer, I.A.; Hobday, T.J.; et al. Effect of Metformin vs Placebo on Invasive Disease-Free Survival in Patients with Breast Cancer: The MA.32 Randomized Clinical Trial. JAMA 2022, 327, 1963–1973. [Google Scholar] [CrossRef] [PubMed]
- Essa, N.M.; Salem, H.F.; Elgendy, M.O.; Gabr, A.; Omran, M.M.; Hassan, N.A.; Tashkandi, H.M.; Harakeh, S.; Boshra, M.S. Efficacy of Metformin as Adjuvant Therapy in Metastatic Breast Cancer Treatment. J. Clin. Med. 2022, 11, 5505. [Google Scholar] [CrossRef] [PubMed]
- Salah, H.; Rabea, H.; Hassan, A.; Elberry, A.A. Metformin as an Adjuvant Treatment in Non-Diabetic Metastatic Breast Cancer. Bahrain Med. Bull. 2021, 43, 2050312119865114. [Google Scholar]
- Cuyàs, E.; Buxó, M.; Ferri Iglesias, M.J.; Verdura, S.; Pernas, S.; Dorca, J.; Álvarez, I.; Martínez, S.; Pérez-Garcia, J.M.; Batista-López, N.; et al. The C Allele of ATM rs11212617 Associates with Higher Pathological Complete Remission Rate in Breast Cancer Patients Treated with Neoadjuvant Metformin. Front. Oncol. 2019, 28, 193. [Google Scholar] [CrossRef]
- Othman, A.S.; Fatehy, H.; Khadiga, M.A.; Nasef, K.; Hussein, O.; Elkodary, T.R. Neoadjuvant chemotherapy with or without metformin in invasive nonmetastatic breast cancer. Randomized controlled trial. Eur. Chem. Bull. 2023, 12, 7543–7550. [Google Scholar] [CrossRef]
- Azazy, H.A.; Gado, N.M.; Salem, D.A.; El-Ghamry, W.R. Metformin with neoadjuvant chemotherapy in stage II-III breast cancer: A phase II clinical trial. Ann. Oncol. 2020, 31 (Suppl. 4), S323–S324. [Google Scholar] [CrossRef]
- El-Khayat, S.M.; Abouegylah, M.; Abdallah, D.; Geweil, A.G.; Elenbaby, A.M.; Zahra, O.S. The effect of metformin when combined with neoadjuvant chemotherapy in breast cancer patients. Med. Oncol. 2021, 39, 1. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Tong, Y.; Hong, J.; Huang, O.; Wu, J.; He, J.; Chen, W.; Li, Y.; Chen, X.; Shen, K. Neoadjuvant docetaxel, epirubicin, and cyclophosphamide with or without metformin in breast cancer patients with metabolic abnormality: Results from the randomized Phase II NeoMET trial. Breast Cancer Res. Treat. 2023, 197, 525–533. [Google Scholar] [CrossRef] [PubMed]
- Pimentel, I.; Lohmann, A.E.; Ennis, M.; Dowling, R.J.O.; Cescon, D.; Elser, C.; Potvin, K.R.; Haq, R.; Hamm, C.; Chang, M.C.; et al. A phase II randomized clinical trial of the effect of metformin versus placebo on progression-free survival in women with metastatic breast cancer receiving standard chemotherapy. Breast 2019, 48, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Nanni, O.; Amadori, D.; De Censi, A.; Rocca, A.; Freschi, A.; Bologna, A.; Gianni, L.; Rosetti, F.; Amaducci, L.; Cavanna, L.; et al. Metformin plus chemotherapy versus chemotherapy alone in the first-line treatment of HER2-negative metastatic breast cancer. The MYME randomized, phase 2 clinical trial. Breast Cancer Res. Treat. 2019, 174, 433–442. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Gong, C.; Wang, Z.; Zhang, J.; Wang, L.; Zhang, S.; Cao, J.; Tao, Z.; Li, T.; Wang, B.; et al. A randomized phase II study of aromatase inhibitors plus metformin in pre-treated postmenopausal patients with hormone receptor positive metastatic breast cancer. Oncotarget 2017, 8, 84224–84236. [Google Scholar] [CrossRef] [PubMed]
- Rothermundt, C.A.; Cathomas, R.; Gysel, K.; Fischer, N.; Pereira Mestre, N.; Hermanns, T.; Rothschild, S.I.; Mach, N.; Mingrone, W.; Ciriolo, M.; et al. SAKK 08/14—IMPROVE Investigation of metformin in patients with metastatic castration-resistant prostate cancer (mCRPC) in combination with enzalutamide vs. enzalutamide alone. A randomized, open label, phase II trial. Ann. Oncol. 2022, 33 (Suppl. 7), S808–S869. [Google Scholar] [CrossRef]
- Martin, P.J.; Borchiellini, D.; Guillot, B.T.A.; Paoli, J.-B.; Besson, D.; Hilgers, W.; Priou, F.; El Kouri, C.; Hoch, B.; Deville, J.L.; et al. TAXOMET: A French prospective multicentric randomized controlled phase II study comparing docetaxel plus metformin versus docetaxel plus placebo in mCRPC. Clin. Genitourin. Cancer 2021, 19, 501–509. [Google Scholar] [CrossRef] [PubMed]
- Alghandour, R.; Ebrahim, M.; Elshal, A.; Ghobrial, F.; Elzaafarany, M.; ELbaiomy, A.M. Repurposing metformin as anticancer drug: Randomized controlled trial in advanced prostate cancer (MANSMED). Urol. Oncol. Semin. Orig. Investig. 2021, 39, 831.e1–831.e10. [Google Scholar] [CrossRef]
- Kitson, S.J.; Maskell, Z.; Sivalingam, V.N.; Allen, J.L.; Ali, S.; Burns, S.; Gilmour, K.; Latheef, R.; Slade, R.J.; Pemberton, P.W.; et al. PRE-surgical Metformin in Uterine Malignancy (PREMIUM): A Multi-Center, Randomized Double-Blind, Placebo-Controlled Phase III Trial. Clin. Cancer Res. 2019, 25, 2424–2432. [Google Scholar] [CrossRef]
- Mitsuhashi, A.; Sato, Y.; Kiyokawa, T.; Koshizaka, M.; Hanaoka, H.; Shozu, M. Phase II study of medroxyprogesterone acetate plus metformin as a fertility-sparing treatment for atypical endometrial hyperplasia and endometrial cancer. Ann. Oncol. 2016, 27, 262–266. [Google Scholar] [CrossRef] [PubMed]
- Glasberg, J.; Talans, A.; Giollo, T.R.; Recchimuzzi, D.Z.; Neto, J.E.B.; Lopez, R.V.M.; Hoff, P.M.G.; Riechelmann, R.P. METNET: A phase II trial of metformin in patients with well-differentiated neuroendocrine tumours. Ecancermedicalscience 2022, 16, 1369. [Google Scholar] [CrossRef] [PubMed]
- Hosono, K.; Endo, H.; Takahashi, H.; Sugiyama, M.; Sakai, E.; Uchiyama, T.; Suzuki, K.; Iida, H.; Sakamoto, Y.; Yoneda, K.; et al. Metformin suppresses colorectal aberrant crypt foci in a short-term clinical trial. Cancer Prev. Res. 2010, 3, 1077–1083. [Google Scholar] [CrossRef] [PubMed]
- Higurashi, T.; Hosono, K.; Takahashi, H.; Komiya, Y.; Umezawa, S.; Sakai, E.; Uchiyama, T.; Taniguchi, L.; Hata, Y.; Uchiyama, S.; et al. Metformin for chemoprevention of metachronous colorectal adenoma or polyps in post-polypectomy patients without diabetes: A multicentre double-blind, placebo-controlled, randomised phase 3 trial. Lancet Oncol. 2016, 17, 475–483. [Google Scholar] [CrossRef]
- Albertelli, M.; Nazzari, E.; Dotto, A.; Grasso, L.F.; Sciallero, S.; Pirchio, R.; Rebora, A.; Boschetti, M.; Pivonello, R.; Ricci Bitti, S.; et al. Possible protective role of metformin therapy on colonic polyps in acromegaly: An exploratory cross-sectional study. Eur. J. Endocrinol. 2021, 184, 419–425. [Google Scholar] [CrossRef] [PubMed]
- Zell, J.A.; McLaren, C.E.; Morgan, T.R.; Lawson, M.J.; Rezk, S.; Albers, C.G.; Chen, W.-P.; Carmichael, J.C.; Chung, J.; Richmond, E.; et al. A Phase IIa Trial of Metformin for Colorectal Cancer Risk Reduction among Individuals with History of Colorectal Adenomas and Elevated Body Mass Index. Cancer Prev. Res. 2020, 13, 203–212. [Google Scholar] [CrossRef]
- Han, J.; Kim, J.H.; Lee, J.W.; Han, S.H.; Kim, H. Neoadjuvant therapy of metformin is associated with good tumor response after preoperative concurrent chemoradiotherapy for rectal cancer. Sci. Rep. 2022, 12, 3716. [Google Scholar] [CrossRef]
- Singh, P.P.; Shi, Q.; Foster, N.R.; Grothey, A.; Nair, S.G.; Chan, E.; Shields, A.F.; Goldberg, R.M.; Gill, S.; Kahlenberg, M.S.; et al. Relationship Between Metformin Use and Recurrence and Survival in Patients with Resected Stage III Colon Cancer Receiving Adjuvant Chemotherapy: Results from North Central Cancer Treatment Group N0147 (Alliance). Oncologist 2016, 21, 1509–1521. [Google Scholar] [CrossRef]
- Available online: https://www.clinicaltrials.gov/study/NCT05921942 (accessed on 18 October 2023).
- Available online: https://clinicaltrials.gov/study/NCT02614339 (accessed on 18 October 2023).
- Available online: https://clinicaltrials.gov/study/NCT01905046 (accessed on 18 October 2023).
- Joshua, A.M.; Armstrong, A.; Crumbaker, M.; Scher, H.I.; de Bono, J.; Tombal, B.; Hussain, M.; Sternberg, C.N.; Gillessen, S.; Carles, J.; et al. Statin and metformin use and outcomes in patients with castration-resistant prostate cancer treated with enzalutamide: A meta-analysis of AFFIRM, PREVAIL and PROSPER. Eur. J. Cancer 2022, 170, 285–295. [Google Scholar] [CrossRef]
- Available online: https://clinicaltrials.gov/study/NCT01864096 (accessed on 18 October 2023).
- Available online: https://clinicaltrials.gov/study/NCT00268476 (accessed on 18 October 2023).
- Available online: https://clinicaltrials.gov/study/NCT02511665 (accessed on 18 October 2023).
- Mitsuhashi, A.; Kawasaki, Y.; Hori, M.; Fujiwara, T.; Hanaoka, H.; Shozu, M. Medroxyprogesterone acetate plus metformin for fertility-sparing treatment of atypical endometrial hyperplasia and endometrial carcinoma: Trial protocol for a prospective, randomised, open, blinded-endpoint design, dose-response trial (FELICIA trial). BMJ Open 2020, 10, e035416. [Google Scholar] [CrossRef]
- Available online: https://clinicaltrials.gov/study/NCT01697566 (accessed on 18 October 2023).
- Available online: https://www.clinicaltrials.gov/study/NCT01797523 (accessed on 18 October 2023).
- Available online: https://clinicaltrials.gov/study/NCT02065687 (accessed on 18 October 2023).
- Pusceddu, S.; Vernieri, C.; Di Maio, M.; Marconcini, R.; Spada, F.; Massironi, S.; Ibrahim, T.; Brizzi, M.P.; Campana, D.; Faggiano, A.; et al. Metformin use is associated with longer progression-free survival of patients with diabetes and pancreatic neuroendocrine tumors receiving everolimus and/or somatostatin analogues. Gastroenterology 2018, 155, 479–489.e7. [Google Scholar] [CrossRef]
- Available online: https://www.clinicaltrials.gov/study/NCT02294006 (accessed on 27 February 2024).
- Available online: https://www.clinicaltrials.gov/study/NCT02823691 (accessed on 27 February 2024).
- Available online: https://clinicaltrials.gov/study/NCT05445791 (accessed on 27 February 2024).
- Available online: https://clinicaltrials.gov/study/NCT02190838 (accessed on 27 February 2024).
- Available online: https://clinicaltrials.gov/study/NCT03311308 (accessed on 27 February 2024).
- Available online: https://www.clinicaltrials.gov/study/NCT02672488 (accessed on 27 February 2024).
- Available online: https://clinicaltrials.gov/study/NCT02005419 (accessed on 27 February 2024).
- Available online: https://clinicaltrials.gov/study/NCT01666730 (accessed on 27 February 2024).
- Yu, O.H.Y.; Suissa, S. Metformin and Cancer: Solutions to a Real-World Evidence Failure. Diabetes Care 2023, 46, 904–912. [Google Scholar] [CrossRef]
- Lord, S.R.; Harris, A.L. Is it still worth pursuing the repurposing of metformin as a cancer therapeutic? Br. J. Cancer 2023, 128, 958–966. [Google Scholar] [CrossRef]
- Wang, P.Y.; Li, J.; Walcott, F.L.; Kang, J.G.; Starost, M.F.; Talagala, S.L.; Zhuang, J.; Park, J.H.; Huffstutler, R.D.; Bryla, C.M.; et al. Inhibiting mitochondrial respiration prevents cancer in a mouse model of Li-Fraumeni syndrome. J. Clin. Investig. 2017, 127, 132–136. [Google Scholar] [CrossRef]
- Panaampon, J.; Zhou, Y.; Saengboonmee, C. Metformin as a booster of cancer immunotherapy. Int. Immunopharmacol. 2023, 121, 110528. [Google Scholar] [CrossRef]
- Amin, D.; Richa, T.; Mollaee, M.; Zhan, T.; Tassone, P.; Johnson, J.; Luginbuhl, A.; Cognetti, D.; Martinez-Outschoorn, U.; Stapp, R.; et al. Metformin Effects on FOXP3+ and CD8+ T Cell Infiltrates of Head and Neck Squamous Cell Carcinoma. Laryngoscope 2020, 130, E490–E498. [Google Scholar] [CrossRef]
- Curry, J.M.; Johnson, J.; Mollaee, M.; Tassone, P.; Amin, D.; Knops, A.; Whitaker-Menezes, D.; Mahoney, M.G.; South, A.; Rodeck, U.; et al. Metformin clinical trial in HPV+ and HPV–head and neck squamous cell carcinoma: Impact on cancer cell apoptosis and immune infiltrate. Front. Oncol. 2018, 8, 436. [Google Scholar] [CrossRef]
- Wang, S.; Lin, Y.; Xiong, X.; Wang, L.; Guo, Y.; Chen, Y.; Chen, S.; Wang, G.; Lin, P.; Chen, H.; et al. Low-Dose Metformin Reprograms the Tumor Immune Microenvironment in Human Esophageal Cancer: Results of a Phase II Clinical Trial. Clin. Cancer Res. 2020, 26, 4921–4932. [Google Scholar] [CrossRef]
Figure 1.
Ianus Bifrons sculpture (https://iStock.com/ accessed on 22 May 2023).
Figure 1.
Ianus Bifrons sculpture (https://iStock.com/ accessed on 22 May 2023).

Figure 2.
(A,B) Frontspiece of Volume XXI of John Hill’s huge work and graphic reproduction of Galena officinalis, p54, 1772 [2]. (C) Photo of Galena officinalis by Peter Smith, Aylestone Meadows, 10 July 2013, www.natarespot.org.uk (accessed on 22 May 2023).
Figure 2.
(A,B) Frontspiece of Volume XXI of John Hill’s huge work and graphic reproduction of Galena officinalis, p54, 1772 [2]. (C) Photo of Galena officinalis by Peter Smith, Aylestone Meadows, 10 July 2013, www.natarespot.org.uk (accessed on 22 May 2023).

Figure 3.
Absorption, uptake, and elimination of metformin. After oral intake, absorption in the gastrointestinal tract is mediated by specific molecular transporters that allow a drug concentration in the portal system of 40–70 µmol/L, higher than that in systemic circulation (10–40 µmol/L). Metformin is excreted unchanged in the urine, and active tubular secretion in the kidney is the main route of drug elimination. PMAT: plasma membrane monoamine transporter (expressed within the apical membranes of enterocytes in the small intestine, and variants are associated with poor tolerance in subjects affected by diabetes mellitus). OCT1/2/3: organic cation transporter. MATE1/2: multidrug and toxin extrusion proteins.
Figure 3.
Absorption, uptake, and elimination of metformin. After oral intake, absorption in the gastrointestinal tract is mediated by specific molecular transporters that allow a drug concentration in the portal system of 40–70 µmol/L, higher than that in systemic circulation (10–40 µmol/L). Metformin is excreted unchanged in the urine, and active tubular secretion in the kidney is the main route of drug elimination. PMAT: plasma membrane monoamine transporter (expressed within the apical membranes of enterocytes in the small intestine, and variants are associated with poor tolerance in subjects affected by diabetes mellitus). OCT1/2/3: organic cation transporter. MATE1/2: multidrug and toxin extrusion proteins.

Figure 4.
Direct (left) and indirect (right) pathways involved in the molecular mechanisms of the antineoplastic role of metformin. For specific actions of subcellular signals, see the text.
Figure 4.
Direct (left) and indirect (right) pathways involved in the molecular mechanisms of the antineoplastic role of metformin. For specific actions of subcellular signals, see the text.


{kind=link}
{kind=link}
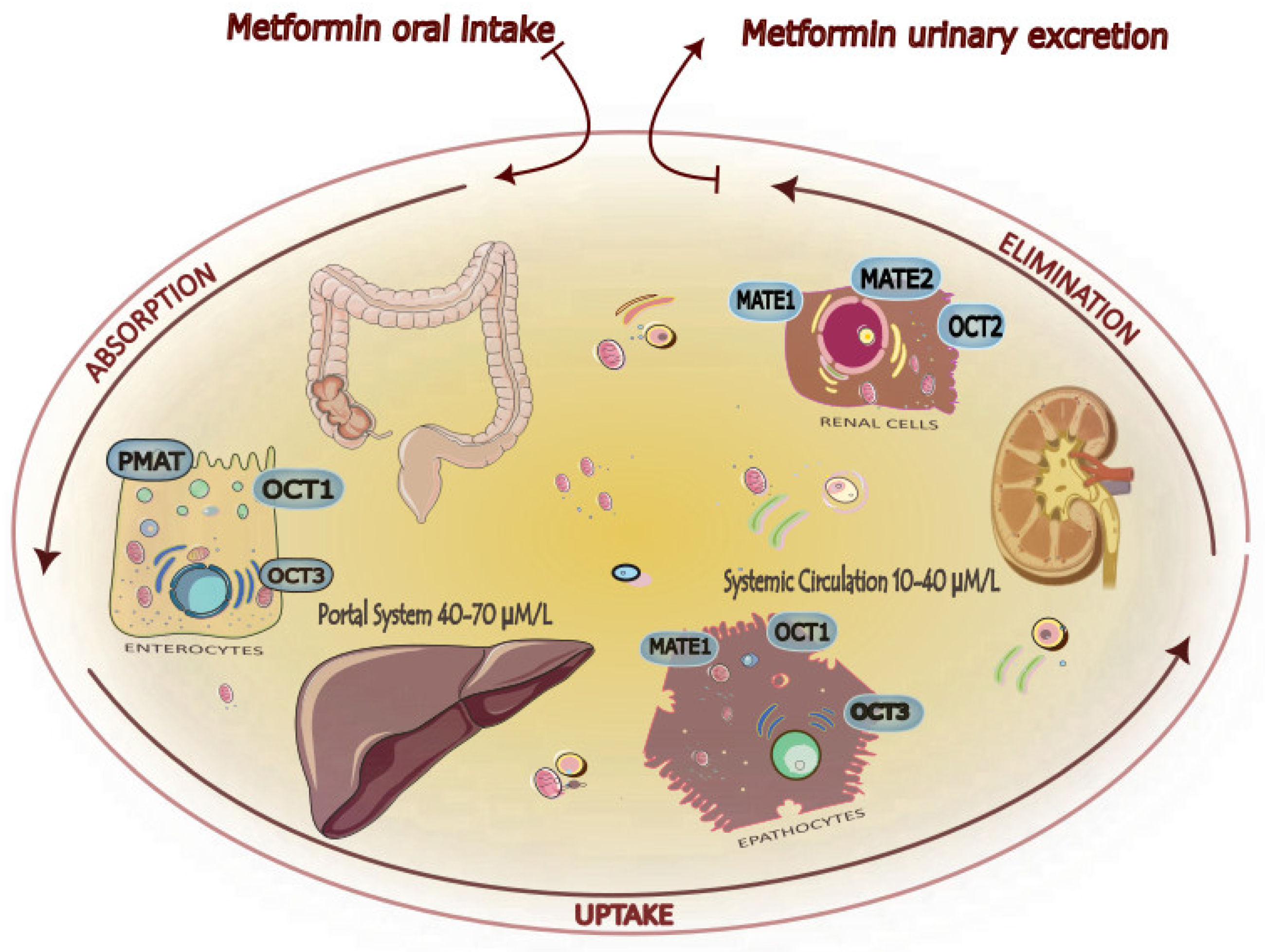{kind=link}
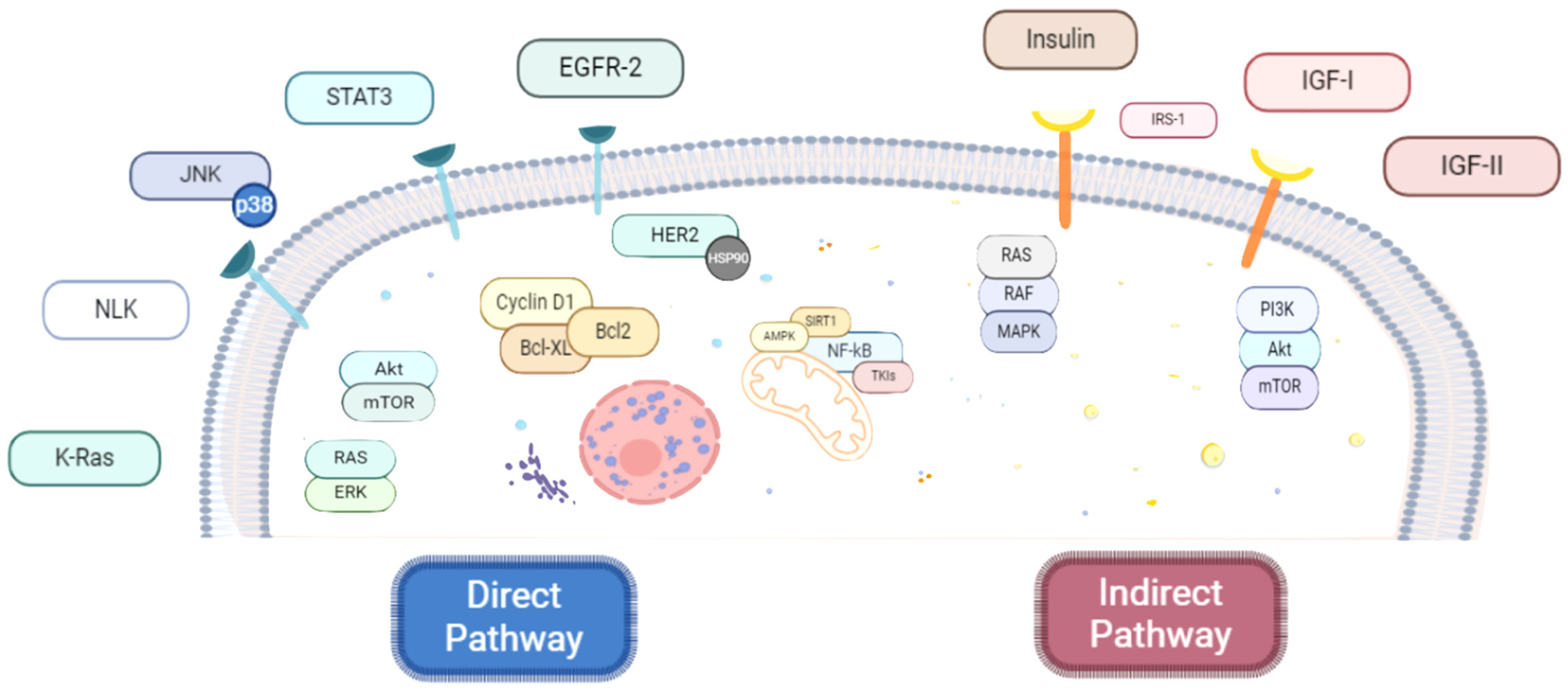{kind=link}
Table 1.
Comparisons between guanine, guanidine, and biguanides (metformin, phenformin, and buformin). LogP: octanol-water partition coefficient (Log P is positive for lipophilic and negative for hydrophilic substances or species).
Table 1.
Comparisons between guanine, guanidine, and biguanides (metformin, phenformin, and buformin). LogP: octanol-water partition coefficient (Log P is positive for lipophilic and negative for hydrophilic substances or species).
| Name | Chemical Formula | Chemical Structure | Features and Origin | Solubility LogP | Binding to Mithocondrial Membranes | Tissue of Anaerobic Glycolysis | Metabolism | Risk of Lactic Acidosis (Events for 1000 Subjects/Year) |
|---|---|---|---|---|---|---|---|---|
| GUANINE | C5H5N5O |  | In 1844, the German chemist Julius Bodo Unger obtained it as a mineral formed from the excreta of sea birds (guano). | Insoluble in water | ||||
| GUANIDINE | HNC(NH2)2 |  | It is a strong base, obtained from natural source, via the oxidative degradation of guanine | Soluble in water and ethanol −1.7 | ||||
| METFORMIN | C4H11N5 |  | N,N-dimethylamine guanylguanidine (chemical syntesis) | More hydrophilic −1.43 | Weaker | Mostly intestinal tissue exposed to high drug concentration | Not metabolized, eliminated unchanged | 0.03–0.09 |
| PHENFORMIN | C10H15N5 |  | It was developed in 1957 by Ungar, Freedman, and Shapiro | More lipophilic −0.83 | Stronger | More generalized, including muscle | Not metabolized, eliminated unchanged | 0.40–0.90 |
| BUFORMIN | C6H15N5 |  | It is a strong base, and is freely soluble in water, methanol, and ethanol | Intermediate −1.20 | Stronger | More generalised, including muscle | Not metabolized, eliminated unchanged | >0.1 |
Table 2.
Type of study, intervention/treatment, patients, and outcomes in main clinical trials in different neoplasms (colorectal cancer, breast cancer, prostate cancer, and GEP-NET).
Table 2.
Type of study, intervention/treatment, patients, and outcomes in main clinical trials in different neoplasms (colorectal cancer, breast cancer, prostate cancer, and GEP-NET).
| Trial Title | Tumor | Type of Study | Intervention/Treatment | Patients, n | Primary Outcome (s) | Results | References |
|---|---|---|---|---|---|---|---|
| Phase 2 Trial of Metformin Combined With 5-Fluorouracil in Patients With Refractory Metastatic Colorectal Cancer | Colorectal cancer | Single-arm, phase II study | Metformin 850 mg bid plus 5-FU 425 mg/m2 and leucovorin 50 mg i.v. weekly | 50 | DCR at 8 weeks | Modest activity of metformin plus 5-FU with major benefits observed in patients with BMI ≥ 30 kg/m2 11 (22%) patients presented DCR at 8 weeks with mPFS = 5.6 months and mOS = 16.2 months | Miranda et al., 2016 [140] |
| Phase II trial of nivolumab and metformin in patients with treatment-refractory microsatellite stable metastatic colorectal cancer. | Colorectal cancer | Single-arm, phase II study | Metformin 1000 mg bid plus Nivolumab 480 mg i.v. every 4 weeks | 24 | ORR | No ORR was observed; the study did not proceed with further enrollment | Akce et al., 2023 [141] |
| Impact of Metformin Use and Diabetic Status During Adjuvant Fluoropyrimidine-Oxaliplatin Chemotherapy on the Outcome of Patients with Resected Colon Cancer: A TOSCA Study Subanalysis | Colon cancer | Substudy of phase III TOSCA trial | Metformin plus adjuvant fluoropyrimidine-oxaliplatin | 3759 | OS and RFS | No impact on OS or RFS | Vernieri et al., 2019 [142] |
| Effect of Metformin vs. Placebo on Invasive Disease-Free Survival in Patients With Breast Cancer: The MA.32 Randomized Clinical Trial | Breast cancer | Phase III, placebo-controlled, double-blind RCT | Drug: metformin 850 mg/day for 4 weeks, then 850 mg bid for 5 years plus adjuvant standard breast cancer treatment Control arm: placebo plus adjuvant standard breast cancer treatment | 3649 | Invasive disease-free survival | No significant improvement of the invasive disease-free survival | Goodwin et al., 2022 [143] |
| Efficacy of Metformin as Adjuvant Therapy in Metastatic Breast Cancer Treatment | Breast cancer | Prospective, placebo-controlled RCT | Drug: metformin 500 mg bid plus adjuvant CT Control arm: adjuvant CT | 107 | PFS and RR | No significant benefits on PFS and RR | Essa et al., 2022 [144] |
| Metformin as an Adjuvant Treatment in Non-Diabetic Metastatic Breast Cancer | Breast cancer | Phase II RCT | Drug: metformin 1000 mg bid plus adjuvant CT Control arm: CT | 50 | OS and PFS | Metformin group vs. control group presented no significant improvement of OS and PFS, but higher radiological response (p = 0.002) | Salah et al., 2021 [145] |
| The C Allele of ATM rs11212617 Associates With Higher Pathological Complete Remission Rate in Breast Cancer Patients Treated With Neoadjuvant Metformin (METTEN) | Breast cancer | Phase II, open-label, multicenter RCT | Drug: metformin 850 mg bid for 24 weeks plus anthracycline/taxane-based CT and trastuzumab Control arm: neoadjuvant anthracycline/taxane-based CT and trastuzumab | 79 | PCR | Metformin group was associated with a higher PCR than control group | Cuyàs et al., 2019 [146] |
| Neoadjuvant chemotherapy with or without metformin in invasive nonmetastatic breast cancer. Randomized controlled trial | Breast cancer | Phase II/III RCT | Drug: metformin 850 mg/day, then 850 mg bid plus neoadjuvant CT Control arm: neoadjuvant CT | 140 | Tumor RR | Metformin plus neoadjuvant CT was associated with higher PCR, especially in ER-negative BC (63.2% vs. 22.2%, p = 0.02) and ER-negative BC (50% vs. 34.6%, p = 0.3) | Othman et al., 2023 [147] |
| Metformin with neoadjuvant chemotherapy in stage II-III breast cancer: A phase II clinical trial. | Breast cancer | Phase II RCT | Drug: metformin 850 mg bid plus neoadjuvant CT Control arm: neoadjuvant CT | 60 | OPR | A higher pCR was observed in the metformin group, but without statistical significance (p = 0.09) | Azazy et al., 2020 [148] |
| The effect of metformin when combined with neoadjuvant chemotherapy in breast cancer patients. | Breast cancer | Prospective study | Drug: metformin 850 mg bid plus neoadjuvant CT Control arm: neoadjuvant CT | 59 | PCR | The addition of metformin may improve PCR particularly in individuals with triple-positive BC and BMI ≥ 25 kg/m2 | El-Khayat et al., 2021 [149] |
| Neoadjuvant docetaxel, epirubicin, and cyclophosphamide with or without metformin in breast cancer patients with metabolic abnormality: results from the randomized Phase II NeoMET trial | Breast cancer | Phase II RCT | Drug: metformin 850 mg/day for the first cycle, then 850 mg bid plus neoadjuvant CT (TEC) Control arm: neoadjuvant CT (TEC) | 92 | PCR | No improvement of PCR | Huang et al., 2023 [150] |
| A phase II randomized clinical trial of the effect of metformin versus placebo on progression-free survival in women with metastatic breast cancer receiving standard chemotherapy | Breast cancer | Phase II, double-blind, RCT | Drug: metformin 850 mg bid plus CT Control arm: placebo plus CT | 40 | PFS | No significant impact on PFS | Pimentel et al., 2019 [151] |
| Metformin plus chemotherapy versus chemotherapy alone in the first-line treatment of HER2-negative metastatic breast cancer. The MYME randomized, phase 2 clinical trial. | Breast cancer | Phase II RCT | Drug: metformin 2000 mg/day plus CT Control arm: CT | 122 | PFS | No significant impact on PFS | Nanni et al., 2019 [152] |
| A randomized phase II study of aromatase inhibitors plus metformin in pre-treated postmenopausal patients with hormone receptor positive metastatic breast cancer. | Breast cancer | Phase II RCT | Drug: metformin 500 mg bid plus aromatase inhibitor (exemestane 25 mg/d or letrozole 2.5 mg/d) Control arm: aromatase inhibitor (exemestane 25 mg/d or letrozole 2.5 mg/d) | 60 | PFS | No significant impact on PFS | Zhao et al., 2017 [153] |
| SAKK 08/14—IMPROVE Investigation of metformin in patients with metastatic castration-resistant prostate cancer (mCRPC) in combination with enzalutamide vs. enzalutamide alone. A randomized, open label, phase II trial. | Prostate cancer | Phase II, open-label, multicenter RCT | Drug: metformin 850 mg bid plus enzalutamide 160 mg Control arm: enzalutamide 160 mg | 169 | DCR at 15 months | No benefits on DCR | Rothermundt et al., 2022 [154] |
| TAXOMET: A French prospective multicentric randomized controlled phase II study comparing docetaxel plus metformin versus docetaxel plus placebo in mCRPC. | Prostate cancer | Phase II, placebo-controlled, multicenter RCT | Drug: metformin 850 mg bid plus docetaxel 75 mg/m2 every 21 days plus prednisone 5 mg bid Control arm: placebo plus docetaxel 75 mg/m2 every 21 days plus prednisone 5 mg bid | 99 | PSA response ≥ 50% from baseline | No significant benefits on PSA response, PFS, OS or ORR | Martin et al., 2021 [155] |
| Repurposing metformin as anticancer drug: Randomized controlled trial in advanced prostate cancer (MANSMED) | Prostate cancer | Phase II RCT | Drug: metformin plus standard of care Control arm: standard of care | 124 | CRPC-FS | A significantly higher CRPC-FS (29 months vs. 20 months, p = 0.01) was observed in the metformin group, especially in individuals with high-risk localized disease or metastatic low tumor volume disease | Alghandour et al., 2021 [156] |
| PRE-surgical Metformin In Uterine Malignancy (PREMIUM): a Multi-Center, Randomized Double-Blind, Placebo-Controlled Phase III Trial | Endometrial cancer or AEH | Phase III, multicenter, double-blind RCT | Drug: neoadjuvant metformin 850 mg/d for 3 days, then 850 mg bid for 1 to 5 weeks Control arm: placebo for 1 to 5 weeks until surgery | 88 | Post-treatment IHC expression of Ki-67 | No differences in Ki67 expression were detected | Kitson et al., 2019 [157] |
| Phase II study of medroxyprogesterone acetate plus metformin as a fertility-sparing treatment for atypical endometrial hyperplasia and endometrial cancer. | Endometrial cancer or AEH | Single-arm, phase II trial | Drug: metformin (750–2250 mg/day) plus medroxyprogesterone acetate 400 mg/day | 17 with AEH and 19 with EC | RFS after remission | Beneficial effect of metformin in inhibiting disease relapse (3-year RFS rate = 89%) | Mitsuhashi et al., 2016 [158] |
| METNET: a phase II trial of metformin in patients with well-differentiated neuroendocrine tumours. | GEP-NET or pulmonary NET | Single-arm, phase II trial | Drug: metformin 850 mg bid | 28 | DCR at 6 months | Modest antineoplastic activity of metformin in well-differentiated GEP or lung NET 26 patients had progression, 13 (46%) of whom presented DCR at 6 months and mPFS 6.3 months | Glasberg et al., 2022 [159] |
Abbreviations: RCT: randomized clinical trial, bid: twice a day, 5-FU: 5-flurouracil, i.v.: intravenous, DCR: disease control rate, BMI: body mass index, PFS: progression-free survival, mPFS: median PRS, OS: overall survival, mOS: median OS, ORR: overall response rate, RFS: relapse-free survival, CT: chemotherapy, RR: response rate, PCR: pathological complete response, OPR: overall pathological response, BC: breast cancer, TEC: TEC docetaxel 75 mg/m2, epirubicin 75 mg/m2, and cyclophosphamide 500 mg/m2, d1, q3w; PSA: prostate-specific antigen, CRPC-FS: castration-resistant prostate cancer-free survival, AEH: atypical endometrial hyperplasia, IHC: immunohistochemical, GEP: gastreoenteropancreatic, and NET: neuroendocrine tumor.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Goglia, U.; Hasballa, I.; Teti, C.; Boschetti, M.; Ferone, D.; Albertelli, M. Ianus Bifrons: The Two Faces of Metformin. Cancers 2024, 16, 1287. https://doi.org/10.3390/cancers16071287
AMA Style
Goglia U, Hasballa I, Teti C, Boschetti M, Ferone D, Albertelli M. Ianus Bifrons: The Two Faces of Metformin. Cancers. 2024; 16(7):1287. https://doi.org/10.3390/cancers16071287
Chicago/Turabian StyleGoglia, Umberto, Iderina Hasballa, Claudia Teti, Mara Boschetti, Diego Ferone, and Manuela Albertelli. 2024. "Ianus Bifrons: The Two Faces of Metformin" Cancers 16, no. 7: 1287. https://doi.org/10.3390/cancers16071287
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.