
Plasma Exosome Proteins ILK1 and CD14 Correlated with Organ-Specific Metastasis in Advanced Gastric Cancer Patients

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients and Samples
2.2. Exosome Isolation and Purification
2.3. Nano-HPLC–MS/MS Analysis
2.4. Bioinformation Analysis
2.5. ELISA Analysis
2.6. Cell Lines and Transfection
2.7. In Vitro Experiments and Western Blot
2.8. Single-Cell RNA Sequencing (scRNA-Seq) Analysis
2.9. Statistical Analysis
3. Results
3.1. Clinical Characteristics of 12 AGC Patients with Organ-Specific Metastasis
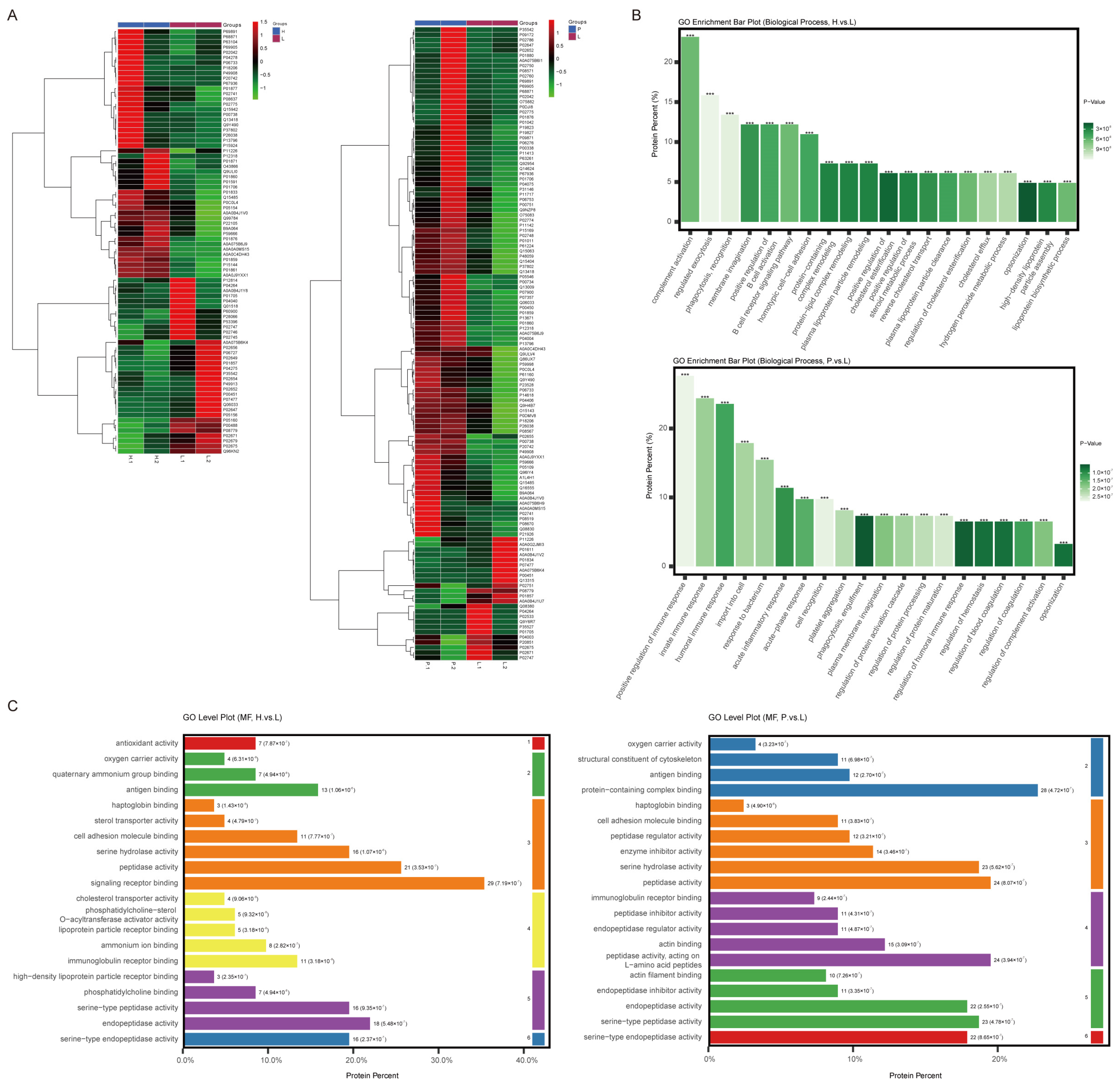
3.2. Distinct Biological Functions of Differential Expression Proteins (DEPs) Detected in Plasma Exosomes from H Group and P Group
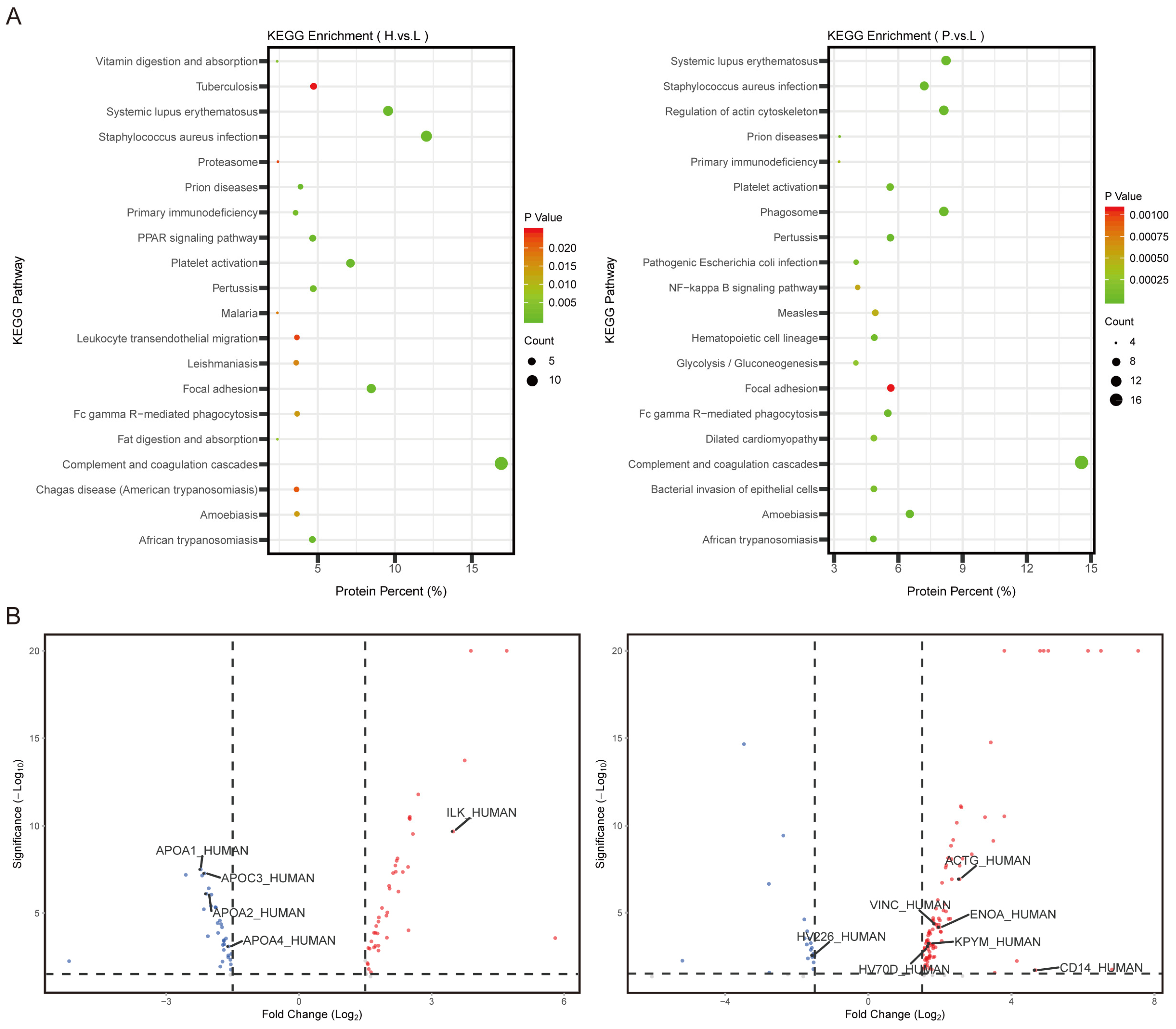
3.3. Signaling Pathways Involved in Lipid Metabolism in the H Group and in Cytoskeleton Regulation in the P Group Were Identified
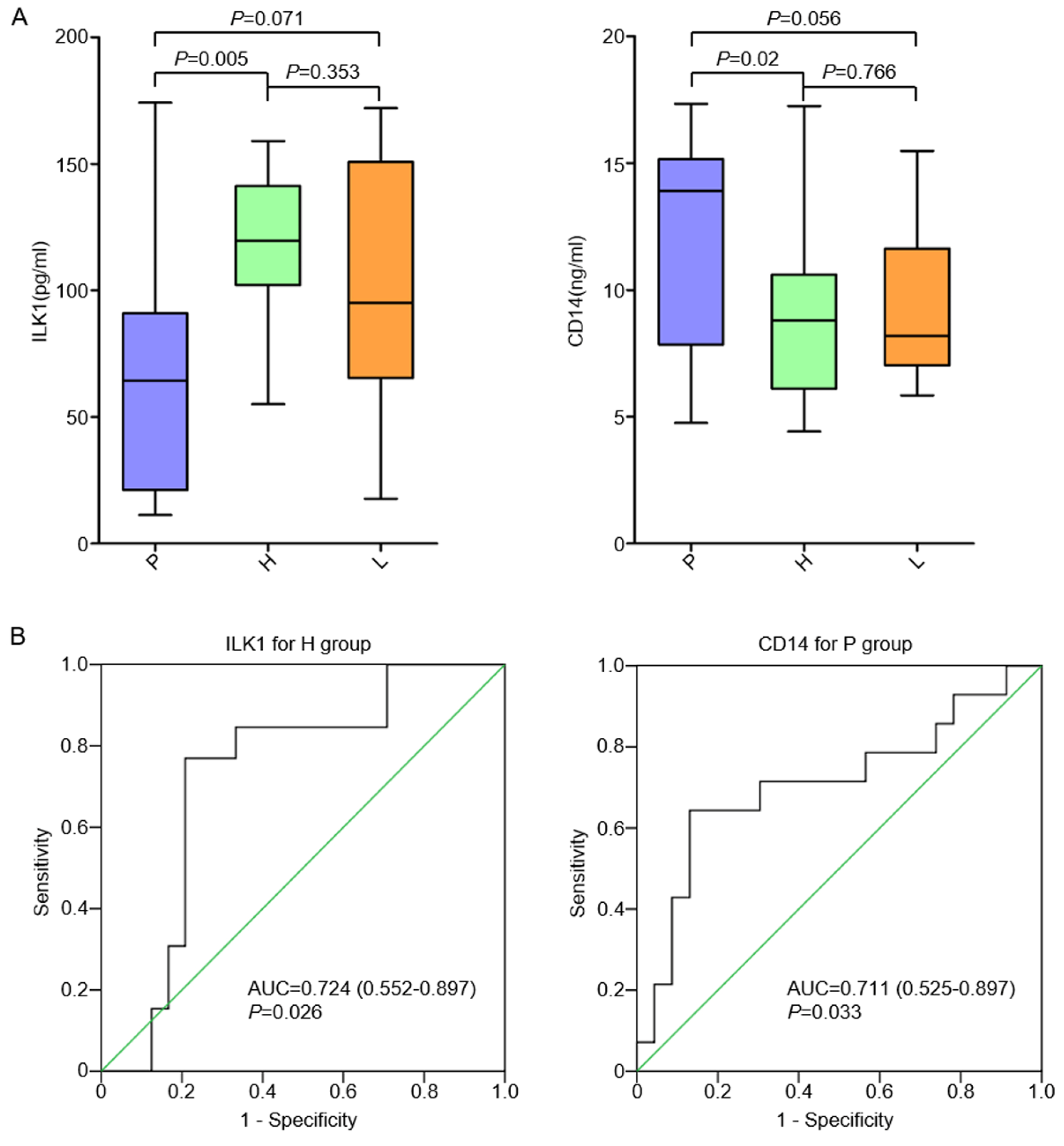
3.4. Plasma ILK1 and CD14 Correlated with Hepatic and Peritoneal Metastasis in AGC, Respectively
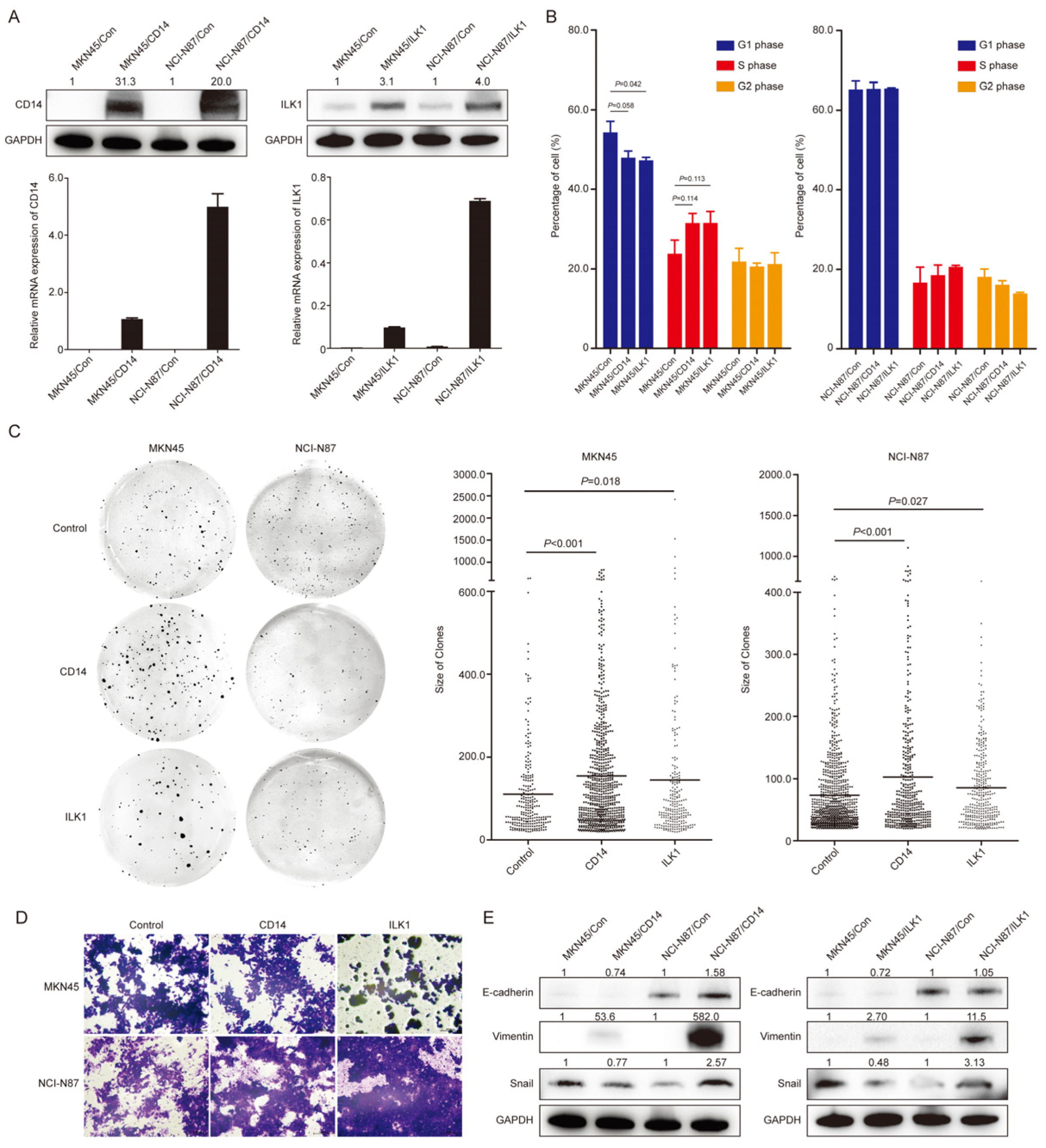
3.5. Overexpression of CD14 and ILK1 Impacted Biological Behaviors in Gastric Cancer Cells
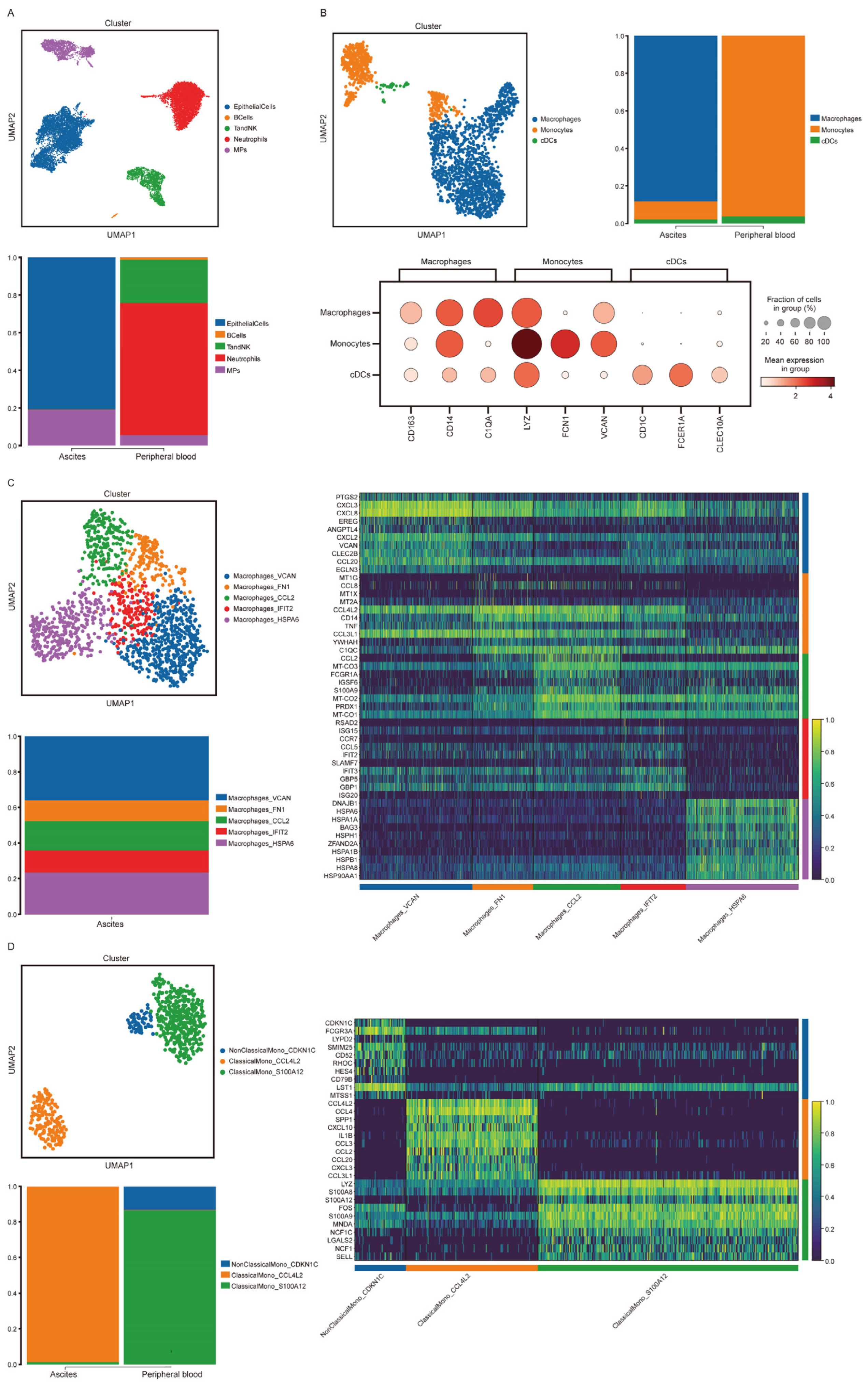
3.6. Immune Cells in Malignant Ascites in a Gastric Cancer Patient Showed High CD14 Expression
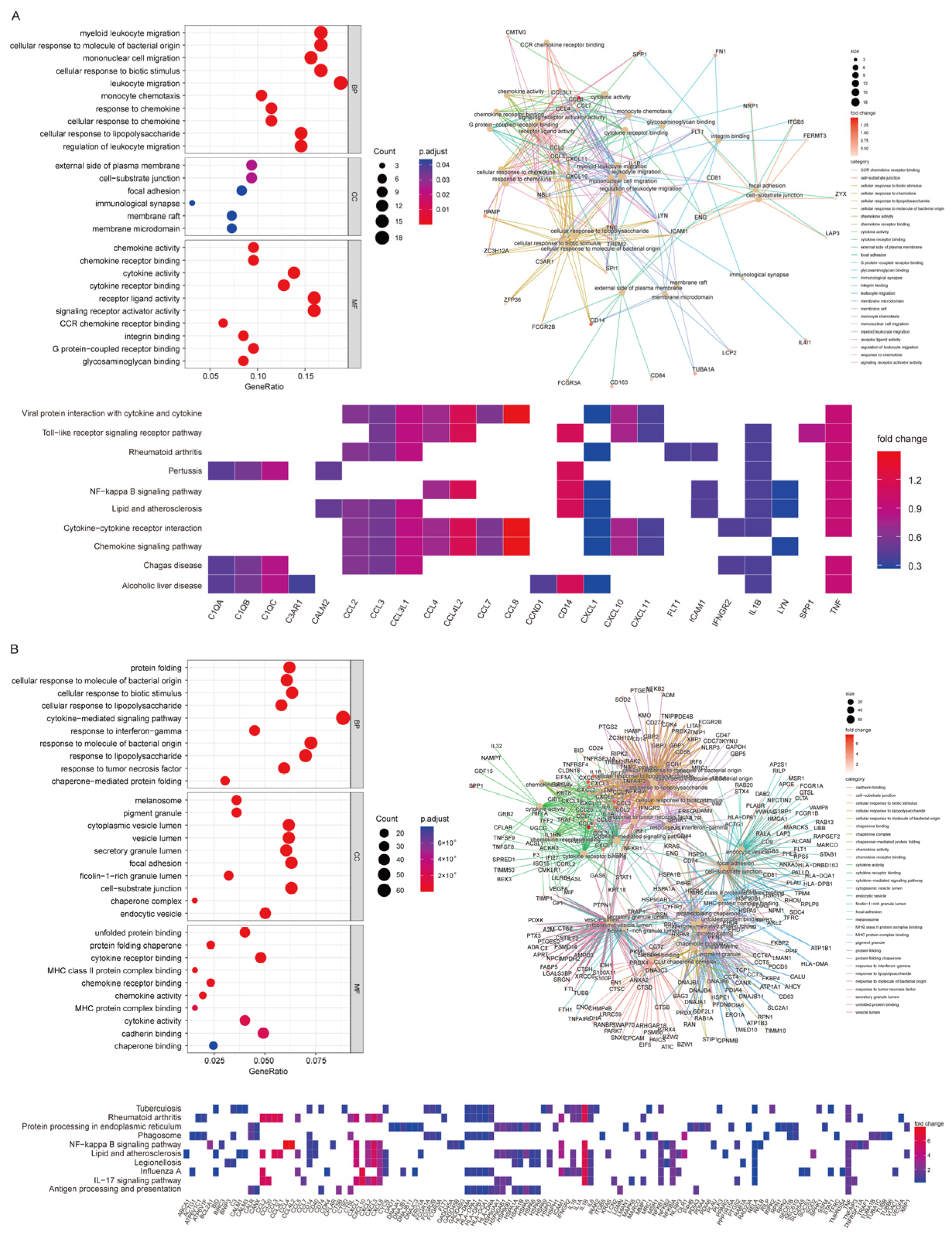
3.7. High CD14 Expression in Macrophages and Monocytes Featured High Activation of Chemokine- and Cytokine-Mediated Signaling Pathways
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shimizu, D.; Kanda, M.; Kodera, Y. Emerging evidence of the molecular landscape specific for hematogenous metastasis from gastric cancer. World J. Gastrointest. Oncol. 2018, 10, 124–136. [Google Scholar] [CrossRef] [PubMed]
- Mikula-Pietrasik, J.; Uruski, P.; Tykarski, A.; Ksiazek, K. The peritoneal “soil” for a cancerous “seed”: A comprehensive review of the pathogenesis of intraperitoneal cancer metastases. Cell. Mol. Life Sci. 2018, 75, 509–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cunningham, D.; Starling, N.; Rao, S.; Iveson, T.; Nicolson, M.; Coxon, F.; Middleton, G.; Daniel, F.; Oates, J.; Norman, A.R.; et al. Capecitabine and oxaliplatin for advanced esophagogastric cancer. N. Engl. J. Med. 2008, 358, 36–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Psaila, B.; Lyden, D. The metastatic niche: Adapting the foreign soil. Nat. Rev. Cancer 2009, 9, 285–293. [Google Scholar] [CrossRef]
- Jiang, H.; Yu, D.; Yang, P.; Guo, R.; Kong, M.; Gao, Y.; Yu, X.; Lu, X.; Fan, X. Revealing the transcriptional heterogeneity of organ-specific metastasis in human gastric cancer using single-cell RNA Sequencing. Clin. Transl. Med. 2022, 12, e730. [Google Scholar] [CrossRef]
- Jin, X.; Demere, Z.; Nair, K.; Ali, A.; Ferraro, G.B.; Natoli, T.; Deik, A.; Petronio, L.; Tang, A.A.; Zhu, C.; et al. A metastasis map of human cancer cell lines. Nature 2020, 588, 331–336. [Google Scholar] [CrossRef]
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef]
- Peinado, H.; Zhang, H.; Matei, I.R.; Costa-Silva, B.; Hoshino, A.; Rodrigues, G.; Psaila, B.; Kaplan, R.N.; Bromberg, J.F.; Kang, Y.; et al. Pre-metastatic niches: Organ-specific homes for metastases. Nat. Rev. Cancer 2017, 17, 302–317. [Google Scholar] [CrossRef]
- Kalluri, R.; LeBleu, V.S. The biology, function, and biomedical applications of exosomes. Science 2020, 367, eaau6977. [Google Scholar] [CrossRef]
- Ma, S.; Zhou, M.; Xu, Y.; Gu, X.; Zou, M.; Abudushalamu, G.; Yao, Y.; Fan, X.; Wu, G. Clinical application and detection techniques of liquid biopsy in gastric cancer. Mol. Cancer 2023, 22, 7. [Google Scholar] [CrossRef]
- Fu, M.; Gu, J.; Jiang, P.; Qian, H.; Xu, W.; Zhang, X. Exosomes in gastric cancer: Roles, mechanisms, and applications. Mol. Cancer 2019, 18, 41. [Google Scholar] [CrossRef] [Green Version]
- Thery, C.; Amigorena, S.; Raposo, G.; Clayton, A. Isolation and characterization of exosomes from cell culture supernatants and biological fluids. Curr. Protoc. Cell Biol. 2006, 30, 3.22.1–3.22.29. [Google Scholar] [CrossRef]
- Zhou, C.; Ji, J.; Cai, Q.; Shi, M.; Chen, X.; Yu, Y.; Liu, B.; Zhu, Z.; Zhang, J. MTA2 promotes gastric cancer cells invasion and is transcriptionally regulated by Sp1. Mol. Cancer 2013, 12, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dura, B.; Choi, J.Y.; Zhang, K.; Damsky, W.; Thakral, D.; Bosenberg, M.; Craft, J.; Fan, R. scFTD-seq: Freeze-thaw lysis based, portable approach toward highly distributed single-cell 3′ mRNA profiling. Nucleic Acids Res. 2019, 47, e16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, G.; Wang, L.G.; Han, Y.; He, Q.Y. clusterProfiler: An R package for comparing biological themes among gene clusters. OMICS 2012, 16, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Al-Sowayan, B.S.; Al-Shareeda, A.T.; Al-Hujaily, E.M. Exosomes, cancer’s little army. Stem Cell Investig. 2019, 6, 9. [Google Scholar] [CrossRef]
- Hoshino, A.; Costa-Silva, B.; Shen, T.L.; Rodrigues, G.; Hashimoto, A.; Tesic Mark, M.; Molina, H.; Kohsaka, S.; Di Giannatale, A.; Ceder, S.; et al. Tumour exosome integrins determine organotropic metastasis. Nature 2015, 527, 329–335. [Google Scholar] [CrossRef] [Green Version]
- Hoshino, A.; Kim, H.S.; Bojmar, L.; Gyan, K.E.; Cioffi, M.; Hernandez, J.; Zambirinis, C.P.; Rodrigues, G.; Molina, H.; Heissel, S.; et al. Extracellular Vesicle and Particle Biomarkers Define Multiple Human Cancers. Cell 2020, 182, 1044–1061.e18. [Google Scholar] [CrossRef]
- Huang, T.; Song, C.; Zheng, L.; Xia, L.; Li, Y.; Zhou, Y. The roles of extracellular vesicles in gastric cancer development, microenvironment, anti-cancer drug resistance, and therapy. Mol. Cancer 2019, 18, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Xian, H.C.; Tang, Y.J.; Liang, X.H.; Tang, Y.L. Fatty acid oxidation: Driver of lymph node metastasis. Cancer Cell Int. 2021, 21, 339. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Yu, W.; Cao, X.; Gu, H.; Huang, J.; Wu, C.; Wang, L.; Sha, X.; Shen, B.; Wang, T.; et al. Exosomal CD44 Transmits Lymph Node Metastatic Capacity Between Gastric Cancer Cells via YAP-CPT1A-Mediated FAO Reprogramming. Front. Oncol. 2022, 12, 860175. [Google Scholar] [CrossRef]
- Li, Y.; Su, X.; Rohatgi, N.; Zhang, Y.; Brestoff, J.R.; Shoghi, K.I.; Xu, Y.; Semenkovich, C.F.; Harris, C.A.; Peterson, L.L.; et al. Hepatic lipids promote liver metastasis. JCI Insight 2020, 5, e136215. [Google Scholar] [CrossRef]
- Yang, Z.; Qin, W.; Chen, Y.; Yuan, B.; Song, X.; Wang, B.; Shen, F.; Fu, J.; Wang, H. Cholesterol inhibits hepatocellular carcinoma invasion and metastasis by promoting CD44 localization in lipid rafts. Cancer Lett. 2018, 429, 66–77. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, Z.; Wang, J.; Chen, C.; Tang, X.; Zhu, J.; Liu, J. Metabolic reprogramming results in abnormal glycolysis in gastric cancer: A review. OncoTargets Ther. 2019, 12, 1195–1204. [Google Scholar] [CrossRef] [Green Version]
- Archid, R.; Solass, W.; Tempfer, C.; Konigsrainer, A.; Adolph, M.; Reymond, M.A.; Wilson, R.B. Cachexia Anorexia Syndrome and Associated Metabolic Dysfunction in Peritoneal Metastasis. Int. J. Mol. Sci. 2019, 20, 5444. [Google Scholar] [CrossRef] [Green Version]
- Jiang, K.; Dong, C.; Yin, Z.; Li, R.; Mao, J.; Wang, C.; Zhang, J.; Gao, Z.; Liang, R.; Wang, Q.; et al. Exosome-derived ENO1 regulates integrin alpha6beta4 expression and promotes hepatocellular carcinoma growth and metastasis. Cell Death Dis. 2020, 11, 972. [Google Scholar] [CrossRef]
- Chen, Y.; Cai, G.; Jiang, J.; He, C.; Chen, Y.; Ding, Y.; Lu, J.; Zhao, W.; Yang, Y.; Zhang, Y.; et al. Proteomic profiling of gastric cancer with peritoneal metastasis identifies a protein signature associated with immune microenvironment and patient outcome. Gastric Cancer 2023, 26, 504–516. [Google Scholar] [CrossRef]
- Almasabi, S.; Ahmed, A.U.; Boyd, R.; Williams, B.R.G. A Potential Role for Integrin-Linked Kinase in Colorectal Cancer Growth and Progression via Regulating Senescence and Immunity. Front. Genet. 2021, 12, 638558. [Google Scholar] [CrossRef]
- Tseng, P.C.; Chen, C.L.; Shan, Y.S.; Chang, W.T.; Liu, H.S.; Hong, T.M.; Hsieh, C.Y.; Lin, S.H.; Lin, C.F. An increase in integrin-linked kinase non-canonically confers NF-kappaB-mediated growth advantages to gastric cancer cells by activating ERK1/2. Cell Commun. Signal. 2014, 12, 69. [Google Scholar] [CrossRef] [Green Version]
- Cortez, V.; Nair, B.C.; Chakravarty, D.; Vadlamudi, R.K. Integrin-linked kinase 1: Role in hormonal cancer progression. Front. Biosci. (Sch. Ed.) 2011, 3, 788–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, D.; Sun, T.; Zhang, X.; Guo, Y.; Yu, D.; Yang, M.; Tan, W.; Wang, G.; Lin, D. Role of CD14 promoter polymorphisms in Helicobacter pylori infection--related gastric carcinoma. Clin. Cancer Res. 2007, 13, 2362–2368. [Google Scholar] [CrossRef] [Green Version]
- Li, K.; Dan, Z.; Hu, X.; Gesang, L.; Ze, Y.; Bianba, Z. CD14 regulates gastric cancer cell epithelialmesenchymal transition and invasion in vitro. Oncol. Rep. 2013, 30, 2725–2732. [Google Scholar] [CrossRef] [Green Version]
- Baumann, C.L.; Aspalter, I.M.; Sharif, O.; Pichlmair, A.; Bluml, S.; Grebien, F.; Bruckner, M.; Pasierbek, P.; Aumayr, K.; Planyavsky, M.; et al. CD14 is a coreceptor of Toll-like receptors 7 and 9. J. Exp. Med. 2010, 207, 2689–2701. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Yang, W.; Sun, A.; Wei, Z.; Lin, Q. The Role of CXC Chemokines in Cancer Progression. Cancers 2022, 15, 167. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Case | Gender | Age | Site | Pathology | Primary Lesion Resection |
|---|---|---|---|---|---|---|
| Lymph node metastasis | L-1 | M | 65 | Corpus | Adenocarcinoma | Yes |
| L-2 | M | 69 | Corpus | Adenocarcinoma | No | |
| L-3 | M | 65 | Corpus | Adenocarcinoma | No | |
| L-4 | F | 69 | Corpus | Adenocarcinoma | No | |
| Hepatic metastasis | H-1 | M | 52 | Cardiac | Adenocarcinoma | No |
| H-2 | M | 66 | Corpus | Poorly differentiated carcinoma | No | |
| H-3 | M | 62 | Antrum | Poorly differentiated carcinoma | No | |
| H-4 | M | 66 | Antrum | Adenocarcinoma | No | |
| Peritoneal metastasis | P-1 | M | 54 | Corpus | Poorly differentiated carcinoma, partial signet-ring cell carcinoma | No |
| P-2 | F | 44 | Cardiac | Poorly differentiated carcinoma, partial signet-ring cell carcinoma | Yes | |
| P-3 | F | 57 | Corpus | Adenocarcinoma, partial signet-ring cell carcinoma | No | |
| P-4 | F | 26 | Corpus | Signet-ring cell carcinoma | No |
| ILK1 n (%) | CD14 n (%) | |||||
|---|---|---|---|---|---|---|
| Group | Low | High | p Value | Low | High | p Value |
| P | 12 (85.7) | 2 (14.3) | 0.003 | 5 (35.7) | 9 (64.3) | 0.004 |
| H | 3 (23.1) | 10 (76.9) | 12 (92.3) | 1 (7.7) | ||
| L | 7 (70.0) | 3 (30.0) | 8 (80.0) | 2 (20.0) | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, C.; Qiao, C.; Ji, J.; Xi, W.; Jiang, J.; Guo, L.; Wu, J.; Qi, F.; Cai, Q.; Damink, S.W.M.O.; et al. Plasma Exosome Proteins ILK1 and CD14 Correlated with Organ-Specific Metastasis in Advanced Gastric Cancer Patients. Cancers 2023, 15, 3986. https://doi.org/10.3390/cancers15153986
Zhou C, Qiao C, Ji J, Xi W, Jiang J, Guo L, Wu J, Qi F, Cai Q, Damink SWMO, et al. Plasma Exosome Proteins ILK1 and CD14 Correlated with Organ-Specific Metastasis in Advanced Gastric Cancer Patients. Cancers. 2023; 15(15):3986. https://doi.org/10.3390/cancers15153986
Chicago/Turabian StyleZhou, Chenfei, Changting Qiao, Jun Ji, Wenqi Xi, Jinling Jiang, Liting Guo, Junwei Wu, Feng Qi, Qu Cai, Steven W. M. Olde Damink, and et al. 2023. "Plasma Exosome Proteins ILK1 and CD14 Correlated with Organ-Specific Metastasis in Advanced Gastric Cancer Patients" Cancers 15, no. 15: 3986. https://doi.org/10.3390/cancers15153986