
Inflammatory Networks in Renal Cell Carcinoma
by
 , ,
, ,
Linus Kruk
1,2,
Medina Mamtimin
1,2,
Attila Braun
1,
Hans-Joachim Anders
2,
Joachim Andrassy
3,
Thomas Gudermann
1,4 and
Elmina Mammadova-Bach
1,2,* 1
Walther-Straub-Institute for Pharmacology and Toxicology, Ludwig-Maximilian-University, 80336 Munich, Germany
2
Division of Nephrology, Department of Medicine IV, Hospital of the Ludwig-Maximilian-University, 80336 Munich, Germany
3
Division of General, Visceral, Vascular and Transplant Surgery, Hospital of LMU, 81377 Munich, Germany
4
German Center for Lung Research (DZL), 80336 Munich, Germany
*
Author to whom correspondence should be addressed.
Cancers 2023, 15(8), 2212; https://doi.org/10.3390/cancers15082212
Submission received: 5 February 2023
/
Revised: 23 March 2023
/
Accepted: 4 April 2023
/
Published: 9 April 2023
{kind=link}
{kind=link}
Abstract
:Simple Summary
Inflammation is an important hallmark of advanced renal cell carcinoma and plays a pivotal role in tumor progression, metastasis, and anti-cancer resistance. Inflammation regulates the crosstalk between tumor cells and the surrounding tumor microenvironment, thereby promoting immune cell infiltration and release of inflammatory mediators and inducing immunosuppressive functions. Although inflammatory pathways sustain the function of renal cancer cells, the activatory effects are highly dependent on the immune cell landscape and the stage of RCC tumors. The aim of this review is to summarize the recent works describing the role of inflammation in the growth and metastasis of RCC tumors. This review also critically discusses the clinically relevant inflammatory pathways as potential targets for disease prevention and treatment.
Abstract
Cancer-associated inflammation has been established as a hallmark feature of almost all solid cancers. Tumor-extrinsic and intrinsic signaling pathways regulate the process of cancer-associated inflammation. Tumor-extrinsic inflammation is triggered by many factors, including infection, obesity, autoimmune disorders, and exposure to toxic and radioactive substances. Intrinsic inflammation can be induced by genomic mutation, genome instability and epigenetic remodeling in cancer cells that promote immunosuppressive traits, inducing the recruitment and activation of inflammatory immune cells. In RCC, many cancer cell-intrinsic alterations are assembled, upregulating inflammatory pathways, which enhance chemokine release and neoantigen expression. Furthermore, immune cells activate the endothelium and induce metabolic shifts, thereby amplifying both the paracrine and autocrine inflammatory loops to promote RCC tumor growth and progression. Together with tumor-extrinsic inflammatory factors, tumor-intrinsic signaling pathways trigger a Janus-faced tumor microenvironment, thereby simultaneously promoting or inhibiting tumor growth. For therapeutic success, it is important to understand the pathomechanisms of cancer-associated inflammation, which promote cancer progression. In this review, we describe the molecular mechanisms of cancer-associated inflammation that influence cancer and immune cell functions, thereby increasing tumor malignancy and anti-cancer resistance. We also discuss the potential of anti-inflammatory treatments, which may provide clinical benefits in RCCs and possible avenues for therapy and future research.
1. Introduction
Renal cell carcinoma (RCC) accounts for 95% of kidney-related malignancies and is considered a heterogeneous group of diseases that are classified according to their histological characteristics [1,2]. The clear-cell RCC (ccRCC) is the most predominant form, followed by non-clear-cell RCCs such as papillary (pRCC) and chromophobe (chRCC), clear-cell papillary (ccpRCC), collecting duct (cdRCC) and medullary (mRCC) and unclassified (uRCC) RCC types [2]. Although the ontogenesis of RCC is not fully elucidated, RCC can be either sporadic (non-inherited) or familial (inherited) and associated with distinct genetic mutations. The most frequently implicated genes in the pathogenesis of ccRCC are von Hippel–Lindau (VHL), PBRM1, SETD2, BAP1, KDM5C and MTOR. Non-clear-cell RCCs present similar genetic alterations with additional genetic mutations such as CDKN2A, NRF2, PTEN, TP53, TFEB, TFE3 and SMARCB1 and with numerous mitochondrial gene dysfunctions [1]. RCC possesses the highest number of indel mutations on a pan-cancer basis. Sporadic RCC can arise as a consequence of kidney injury where injury-related deoxyribonucleic acid (DNA) damage introduces somatic mutations, and subsequent proliferative healing responses drive the monoclonal expansion of mutated epithelial cell clones, e.g., in pRCC. These genomic changes of RCC induce the release of high numbers of tumor-specific antigens (neo-antigens), thereby eliciting the adaptive immune response [3,4,5]. RCC is a highly vascularized tumor, susceptible to the recruitment of immune and stromal cells, forming an inflammatory microenvironment [6,7]. The early stage of RCC is treated by curative surgical resection. The treatment of advanced and metastatic RCC is difficult, based on the combination of several agents, targeting multiple receptor tyrosine kinases, including growth-factor receptors such as vascular endothelial growth factor receptor (VEGFR), or directly targeting VEGF, often in combination with immune checkpoint inhibitors (ICIs) [8,9,10,11,12,13]. The heterogeneity of the genetic background and tumor microenvironment (TME) varies the treatment response among patients [14]. Cancer-associated inflammation is the most recurrent hallmark of many solid cancer types. Recent studies have linked inflammation to metastatic progression in patients with RCCs [15,16,17,18,19,20]. Cancer cell-extrinsic pathological conditions, such as systemic and local infections and inflammatory diseases, can activate oncogenic pathways leading to the development of solid tumors.
On the other hand, cancer cell-intrinsic oncogenic or epigenetic changes lead to pro-inflammatory reactions, including the release of cytokines and growth factors into the TME, thereby inducing cancer progression [21]. Accordingly, high levels of pro-inflammatory cytokines and consequent enhanced systemic inflammatory responses are observed. Elevated C-reactive protein concentration and hypo-albuminuria, elevated neutrophil and platelet counts, and a high neutrophil-lymphocyte ratio have been shown to correlate with poor RCC survival [15,16,17,18,19,20,22,23,24]. In TME, intricate crosstalk between cancer and host cells (i.e., stroma, endothelial, immune cells) induces an immune response, thereby modulating tumor growth [25]. ccRCC tumors are characterized by rich leukocyte infiltrates and an abundance of intratumoral CD8+ and CD4+ T cells, which are associated with high tumor grade and shorter patient survival [26]. RCC tumors express human leukocyte antigen G (HLA-G), and typically, the loss of major histocompatibility complex I (MHCI), which correlates with inhibition of CD8+ T cells, induction of regulatory T cells (Treg) and inhibition of natural killer cells (NKs) and dendritic cells (DCs) [27,28,29]. Studies analyzing the TME-associated gene expression signature could divide RCC into non-inflamed and inflamed subtypes. Based on the histological analysis of RCC, the non-inflamed subtype is characterized by increased angiogenesis and plasmacytoid DCs and mast cell infiltrates, while the inflamed subtype showed high levels of complement system proteins and infiltrates of different immune cell types, such as Treg, NKs, Th, CD8+, B and activated DCs, as well as neutrophils, macrophages and eosinophils [30,31,32]. The inflamed subtype is frequently associated with the clinical manifestation of thrombocytosis or anemia, which account for worse patient survival, highlighting the impact of renal and systemic inflammation on the progression of RCCs and patient outcome [30,31,32]. In this review, we describe the molecular mechanisms of inflammation that influence tumor cell function and immune cell activity, thereby increasing tumor malignancy and anti-cancer resistance. We also discuss potential anti-inflammatory treatments of RCCs, which may provide clinical benefits in patients with advanced tumor and metastases.
2. Inflammatory Signaling Pathways in RCC
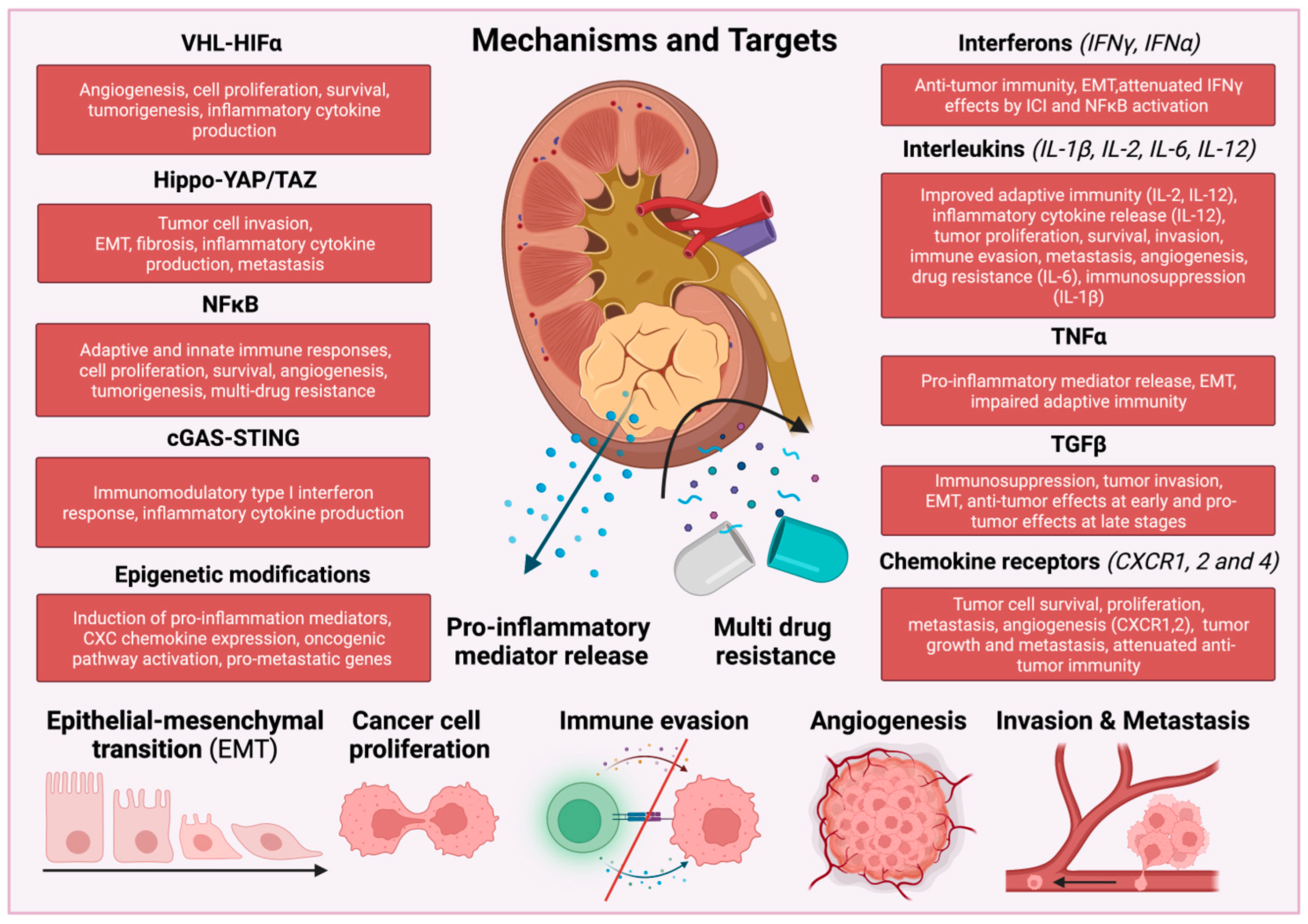
RCCs produce and release several pro-inflammatory cytokines that activate different subtypes of immune cells, thereby promoting tumor progression and metastasis. In RCC tumors, activation of transcription factors, such as nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB), signal transducer and activator of transcription 3 (STAT3) and hypoxia-inducible factor 1α (HIF1α) increase the production of angiogenic factors and inflammatory mediators, such as interleukins IL-1β, IL-6, IL-23 and tumor necrosis factor α (TNFα) and chemokines, creating a cancer-associated inflammatory TME [33,34,35] (Figure 1).
Tumor cell-extrinsic inflammation can be triggered by many factors, including infection, obesity, autoimmune disorders, and exposure to tobacco and toxic substances [36]. Inflammation can also be induced by epigenetic remodeling of paracrine or autocrine signaling, genomic mutation, genome instability of cancer cells, called tumor-intrinsic inflammation, that fuel immunosuppressive or tumor-promoting traits, leading to the recruitment and activation of inflammatory cells [36]. Although due to the complexity of the molecular mechanisms, the distinction between systemic response and tumor-intrinsic activity is challenging.
3. vHL-HIFα
The majority of sporadic ccRCCs represent mutations in the von Hippel–Lindau tumor-suppressor gene (VHL). Therefore, VHL was considered a main target in conventional therapy of ccRCC [37]. The protein VHL is responsible for targeting the alpha (α) subunits of hypoxia-inducible transcription factors (HIF1α and HIF2α) for degradation through a process that depends on oxygen and ubiquitin [38,39]. VHL protein can destabilize hypoxia-induced genes, thereby influencing cellular and tissue response to hypoxia, such as VEGF-mediated angiogenesis, energy metabolism through glucose transporter 1 (GLUT1), or production of platelet-derived growth factor subunit B (PDGFB) [40]. Additionally, VHL protein activates signaling pathways that control the production of several inflammatory cytokines, including TNFα, chemokine receptor CXCR4, and NF-κB [41,42,43].
When VHL is genetically inactivated, HIF1α and HIF2α become stabilized, which leads to changes in cellular metabolism, angiogenesis, epithelial-mesenchymal transition (EMT), invasion, and tumor metastasis that contribute to the progression of ccRCC [44]. HIF2α is believed to have a pro-tumorigenic role in ccRCC, while HIF1α inhibits aggressive tumor behavior, thereby acting as a tumor suppressor. The loss of the region of chromosome 14q that harbors the HIF1A gene is associated with poor survival and is often found in ccRCC metastases [45]. ccRCC tumors expressing only HIF2α have higher proliferation index than those expressing both HIF1α and HIF2α [46]. ccRCC tumor cell lines frequently exhibit intragenic deletions of HIF1A but express wild-type HIF2α [47]. HIF2α is necessary for the growth of ccRCC xenografts, whereas knockdown of HIF1α enhances xenograft tumor formation in cell lines expressing both HIF1α and HIF2α [39,47,48]. Therefore, various therapeutic approaches have been elaborated to target HIF2α signaling routes in ccRCC [49,50]. A recent study based on the introduction of floxed alleles of HIF1α and HIF2α into an autochthonous mouse model of ccRCC challenged this concept [51]. It was shown that the presence of HIF1α is essential for the formation of tumors, whereas the absence of HIF2α has a limited effect on tumor initiation and growth [51]. Although HIF1α plays a crucial role in glycolysis, HIF2α regulates the expression of genes that are involved in various cellular processes, such as lipoprotein metabolism, ribosome biogenesis, and transcriptional activities of E2F and MYC [51]. Interestingly, in this study, HIF2α-deficient tumors were also characterized by increased antigen presentation, interferon (IFN) signaling, and infiltration and activation of CD8+ T cells [51].
Abolished VHL gene expression in mice induces hyperplastic clear cell lesions with severe inflammation and fibrosis [52]. This inflammatory response was associated with increased inositol-requiring transmembrane kinase/endoribonuclease 1α (IRE1α) kinase activity required for the polarization and recruitment of inflammatory macrophages [53]. IRE1α signaling pathway in macrophages drives obesity and metabolic syndrome through impairing brown adipose tissue activity and white adipose tissue browning [54]. VHL deficiency induces the infiltration of macrophages and their polarization toward M2 pro-inflammatory phenotype. Consequently, these macrophages enhance tumor progression and metastasis by secreting high amounts of chemokine (C-C motif) ligand 18 (CCL18) and transforming growth factor beta 1 (TGFβ1) [55].
AXL is a receptor tyrosine kinase expressed in different cancer cell types, and the gene expression is regulated by HIF1α and HIF2α in hypoxic cancer cells lacking VHL protein [56]. Therefore, invasion and metastasis of ccRCC are tightly regulated by AXL signaling route. Downregulation of AXL gene expression or competitive inhibition of AXL function with a soluble variant of AXL (sAXL) has an inhibitory effect on the metastasis of ccRCC cells [57]. Several tyrosine-kinase inhibitors (TKIs) or monoclonal antibodies (bevacizumab, sunitinib and sorafenib) that block VEGFA mediated tyrosine-kinase pathways have been used to treat advanced RCC [10,58,59]. However, only a few patients positively responded to this treatment with long-lasting tumor suppression because the majority of RCC tumors use an alternative “angiogenic escape” pathway, thereby sustaining tumor revascularization [60]. AXL and fibroblast growth factor receptor (FGFR) are involved in this alternative pathway since increased levels of FGFR and AXL are directly associated with cancer cell resistance to the TKI (sunitinib) [61]. Therefore, the other TKI, lenvatinib, was developed, which effectively inhibited different isoforms of FGFR and vascular endothelial growth factor receptor (VEGFR) [61,62]. The combination of lenvantinib with pembrolizumab or everolimus have been successfully used in the treatment of advanced RCC [63,64]. Another therapeutic strategy was developed to target HIF pathways [65]. Belzutifan is a small-molecule inhibitor of HIF2α, which disrupts the heterodimerization of HIF2α with HIF1β, required to form an active transcription factor [66]. Several clinical studies evaluated the effects of belzutifan in RCCs [66], showing improved bioavailability and efficacy compared to other HIF inhibitors [67]. Recently, belzutifan was approved by the Food and Drug Administration (FDA) for adult patients with VHL-associated RCC, central nervous system hemangioblastomas, or pancreatic neuroendocrine tumors without the need for immediate surgery [68].
Interestingly, HIF2α is recruited by the PAX-8-bound transcriptional enhancers such as cyclin D1 (CCND1) and MYC [69,70]. In ccRCC, rs7948643 is a common genetic variant linked to the significant renal cancer risk locus rs7105934 on chromosome 11q13.3, which coincidently falls within PAX8-binding site E11:69419. Both PAX8 and HIF2α play an important role in E11:69419 transcriptional activity, and genetic inactivation of PAX8 is tolerated in the mouse kidney leading to the assumption that PAX8 synergistically to HIF2α could be a therapeutic target in ccRCC [69]. The synthetic lethality of VHL combined with many additional genetic alterations, offers another therapeutic avenue [71]. For instance, disruption of VHL and EZH1, SLC2A1, ROCK1, SKP2, AURKA, or TBK1 genes can each inhibit cancer progression by inducing the death of tumor cells [72,73,74,75,76,77,78]. A recent study discovered that HIF2α accumulation could enhance the gene expression of proliferative CDK4/6 partner cyclin D1 while the synthetic lethality of VHL-/- and cyclin-dependent kinases 4 and 6 (CDK4/6) is independent of HIF2α accumulation [77]. This may therapeutically represent benefits since HIF2α-inhibitors are only effective in a small group of ccRCC patients, and a large cohort of ccRCC patients have pre-existing or acquired resistance to HIF2α-inhibitors [49,77]. Therefore, future studies are needed to determine whether inhibition of CDK4/6 is a suitable therapeutic approach for ccRCC, alone or in combination with HIF2α inhibitors [77].
4. Hippo-YAP/TAZ
Several studies have linked RCC and inflammation with the Hippo pathway. The core components of the Hippo pathway consist of the MST1/2 and LATS1/2 kinases [79]. Downstream from the Hippo pathway are the YAP/TAZ transcription coactivators, which are suppressed through phosphorylation by MST1/2 and LATS1/2 kinases [79]. The Hippo pathway is regulated by the merlin/neurofibromatosis type 2 (NF2), a well-studied tumor suppressor gene. NF2 enhances the Hippo signaling pathway by inhibiting YAP/TAZ nuclear translocation through phosphorylation, isolation, and degradation [79].
Changes in the biomechanical properties of cells and tissues can activate the YAP/TAZ sensor, leading to the release of pro-inflammatory and pro-fibrotic signals. YAP plays a critical role in fibrosis following ischemic acute kidney injury (AKI) [80] and numerous studies have linked aberrant YAP activation to kidney fibrosis [81,82,83,84]. Wnt5a has been shown to stimulate renal fibrosis by inducing M2 macrophage polarization through YAP/TAZ activation [85]. The polyploidization of kidney tubular cells is considered a rapid mechanism to increase the residual function of differentiated tubular cells during the early phase of injury, thereby avoiding AKI fatality. However, during the late phase of AKI, the cycling polyploid kidney tubular cells were shown to become senescent, developing a pro-fibrotic phenotype that promotes chronic kidney disease (CKD) [86]. Interestingly, the blockade of YAP1 and TEAD coactivator transcriptional activity using CA3 (CIL56) during the recovery phase of AKI was proposed as a promising therapeutic approach to prevent CKD [86].
Abnormal Hippo pathway signaling and YAP activation are observed in NF2-deficient tumors, including uRCC [87,88]. Silencing YAP/TAZ in NF2-deficient tumors induces tumor regression by increasing mitochondrial respiration, reducing glycolic-dependent growth, and causing the accumulation of reactive oxygen species (ROS) and consequent oxidative stress-induced cell death [89]. LATS1 kinase has been proposed as a tumor suppressor and is reduced in many types of cancer, including RCC [90,91]. Consistently, high expression of LATS1 or LATS2 is associated with longer disease-free survival (DFS) and overall survival (OS) in RCC patients [92]. Patients with ccRCC also exhibit a significant increase in TAZ expression, which is associated with poor prognosis [93]. It was also shown that TAZ deletion can counteract ferroptosis, which regulates cell death [94]. TAZ regulates the expression of epithelial membrane protein 1 (EMP1), which induces the expression of nicotinamide adenine dinucleotide phosphate (NADPH) oxidase 4 (Nox4)—an enzyme essential for ROS-mediated ferroptosis in the kidneys [94]. Hypoxia-induced Nox4 and NADPH were shown to increase IL-6 and IL-8 levels in RCC, thereby increasing the invasion of cancer cells, [23] indicating a positive feedback loop between IL-6/8 and YAP/TAZ during the progression of RCC.
The Hippo pathway can also interact with the epidermal growth factor receptor (EGFR) pathway, thereby regulating cancer cell invasion [95]. EGFR was shown to downregulate claudin-2, a type of tight junction protein. Its downregulation results in the disruption of claudin-2-YAP interaction, thereby leading to the activation of YAP and oncogenic proteins that induce mesenchymal plasticity of cancer cells, thereby promoting tumor metastasis [95]. In sporadic ccRCC, the expression of tight junction proteins occludin, claudin 1 and E-cadherin is downregulated upon the loss of VHL [96]. Although the overexpression of occludin and claudin 1 could restore the assembly of tight junctions, the re-expression of E-cadherin was insufficient to restore this process [96]. Altogether, these studies indicated the role of the activated YAP pathway in the disruption of epithelial polarity and consequent metastasis.
High-grade ccRCC is associated with the downregulation of salvador homolog 1 (SAV1), which is a tumor suppressor and a component of the Hippo signaling pathway [97,98]. In contrast, the overexpression of YAP1 was associated with the pathogenesis of mucinous tubular spindle cell carcinoma (MTSCC), a rare subtype of renal cell carcinoma [99]. The use of YAP inhibitors, such as verteporfin (VP), was proposed as a treatment for patients with rare MTSCC with sarcomatoid differentiation or metastatic disease [88,99].
The leukemia inhibitory factor receptor (LIFR) is a potential downstream target of the Hippo pathway. Knockdown of the LIFR gene was shown to increase YAP expression and inhibit Hippo signaling pathway kinase activity, thereby promoting tumor metastasis [100]. SH3 domain-binding glutamate rich protein-like 2 (SH3BGRL2) is involved in the suppression of EMT and metastasis in ccRCC tumors by inhibiting the nuclear translocation of YAP and its interaction with the TEAD coactivator in the nuclei [101]. Conversely, the knockdown of SH3BGRL2 activates the Hippo pathway by binding TEAD, leading to the downregulation of Twist1 and the shifting EMT, ultimately inhibiting the growth and metastasis of RCC cells [101]. Recently, CA3 (CIL56), YAP1/TEAD inhibition was shown to strongly inhibit esophageal adenocarcinoma cell growth and induce strong anti-tumor effects by suppressing tumor cell proliferation, and cancer stem cell phenotype in a xenograft model with no apparent toxicity [102]. However, CA3 (CIL56) has not yet been tested in experimental models of RCC. A Phase 1 study has been planned to study the effect of IK-930, an oral TEAD inhibitor targeting the Hippo pathway in patients with advanced tumors, including RCC (NCT05228015).
5. NF-κB Pathway
The nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) is a transcription factor ubiquitously expressed in almost all mammalian cells, forming heterodimers [103]. The classic “canonical” NF-κB pathway forms a complex with p50 and RelA, while the non-canonical NF-κB pathway is predominantly associated with p52/RelB [103]. NF-κB pathways are involved in response to inflammatory stress, which are activated by cytokines, bacterial toxins, viral products and cell death stimuli. Although NF-κB pathways regulate the innate immune system in healthy conditions, abnormal NF-κB activation can trigger abnormal inflammatory responses during tumor development [104]. NF-κB pathways enhance cancer cell migration and invasion by upregulating multi-drug resistance genes, pro-angiogenic factors and pro-inflammatory cytokines such as VEGFA, EGF, IL-6 and IL-8 [105]. Interestingly, the NF-κB subunit p50 regulates macrophage function in cancer, and its inhibition converts the pro-tumor phenotype to M1-like cytotoxic phenotype and consequently attenuates tumor growth [106]. VHL loss could activate NF-κB pathways [107], thereby inducing gene expression of anti-apoptotic Bcl-xL and Bcl-2, and inhibiting tumor suppressor p53, highlighting the potential role in RCC development [108]. Studies showed that elevated NF-κB expression correlated with a low rate of patient survival with RCC [109]. Activation of NF-κB and STAT3 pathways enhances tumor infiltration of M1/M2-like macrophages and Treg cells, thereby increasing tumor growth and metastasis in RCC [110].
N6-adenosine-methyltransferase (METTL3) is the catalytic subunit of the m6A methyltransferase complex, which is involved in the post-transcriptional methylation of adenosine residues of mRNAs. In tumor-infiltrating myeloid cells, METTL3 regulates the induction of NF-κB through the ERK pathway and response to programmed cell death protein 1 (PD-1) checkpoint blockade [111]. Therefore, it was proposed that a blockade of NF-κB activation in myeloid-derived cells and Treg cells may offer a therapeutic avenue by inducing an immunosuppressive response. Contrary, a recent study showed that adoptive cell transfer of T cells with constitutively active upstream NF-κB activator IκB kinase β (IKKβ) increased the number of functional CD8(+) T cells that are selective for tumors and enhanced cytotoxic activity [112]. This demonstrated the bivalent role of NF-κB targeting when applied to specific immune cell subsets.
Bortezomib is a proteasome inhibitor with anti-tumor activity blocking NF-kB function. Bortezomib increases apoptosis, decreases angiogenic cytokine production, and inhibits cancer cell adhesion to stroma [113]. Although bortezomib was used in the treatment of patients with advanced RCCs [114,115], similar to other NF-κB inhibitors, it also has off-target effects. [113,116,117]. Bortezomib enhances caspase-8 activity, sensing RCC to TNF–related apoptosis-inducing ligand (TRAIL). Without priming caspase-8 activation, bortezomib has limited efficiency to induce TRAIL-mediated cancer cell apoptosis [115].
6. cGAS-STING and BAF Complex
The cGAS–STING signalosome is composed of cyclic GMP-AMP synthase (cGAS) and the stimulator of IFN genes (STING), which detect cytoplasmic DNA in virus-infected cells and triggers IFN regulatory transcription factor 3 (IRF3)-dependent inflammatory gene expressions, thereby accelerating the innate immune response [118]. Activated STING is translocated to the Golgi and activates TANK-binding kinase 1 (TBK1), which further phosphorylates and activates IRF3. STING can also activate MAPK and NF-κB pathways, thereby increasing the production of inflammatory cytokines and chemokines [118]. Although cGAS–STING signalosome is important in the DNA detection of pathogens, it also binds endogenous DNA, which is released from the nucleus of cancer or apoptotic cells. Mitochondria dysfunction and mtDNA release also activate the cGAS-STING pathway. Therefore, the cGAS-STING signalosome was proposed to be contributed to anti-tumor immunity by inducing an immune suppressive TME [119]. Sensing of cytosolic DNA in DCs can sustain anti-tumor activity by inducing IFN production, priming cytotoxic CD8+ T cells and triggering cancer cell senescence. Interestingly, this senescence-associated secretory phenotype eliciting the release of pro-inflammatory cytokines, chemokines, proteases, and growth factors, is shown to restrict tumorigenesis [120]. ENPP1, a transmembrane phosphodiesterase in purinergic signaling, can downregulate cGAS-STING signaling by hydrolyzing cyclic guanosine monophosphate–adenosine monophosphate (cGAMP) [121]. Interestingly, brain metastatic cancer cells can activate cGAS-STING signalosome, transferring cGAMP to neighboring astrocytes, thereby increasing the production of inflammatory cytokines such as IFN and TNF and consequently promoting metastasis [120].
Recently, cGAS-STING was also described as a risk factor in different types of RCCs. The score of differentially expressed genes associated with cGAS-STING was shown to be correlated to poor OS and was significantly higher compared to other cancer types [122]. This result suggests a unique TME of RCCs, which is differently regulated by cGAS-STING signalosome, thus promoting tumor formation and metastasis. The effect of cGAS-STING activation appears to be highly dependent on the disease context. In AKI and CKD, STING induction in mice leads to albuminuria and podocyte loss [123]. Supporting this ambivalent role of cGAS-STING, the novel STING antagonist, H151, significantly ameliorates renal function, kidney morphology, and renal inflammation in cisplatin-induced AKI [124].
Currently, several clinical trials are undergoing to explore the efficacy of STING agonists and DNA damage repair (DDR) targeting drugs in patients with different kidney cancer, including mRCC and metastatic ccRCC (NCT03274258; NCT03587662; NCT03010176). The impact of cGAS-STING targeting drugs in the cancer therapy of RCC remains to be addressed by future studies. Especially, contributing pathways with over-activation, and off-target effects of agents on immune cells have not yet been fully understood [125].
The histone-packed nucleosomes need to be opened to expose chromatin-free gene regulatory elements for transcription factors to maintain gene expression [126]. Nucleosome repositioning is regulated by the chromatin-remodeling complex BAF [126]. Interestingly, many cancer cells carry BAF mutations indicating an important tumor-suppressive role in humans [126]. The SWI/SNF related, matrix-associated, actin-dependent regulator of chromatin, subfamily B, member 1 (SMARCB1) protein is a subunit of the BAF complex. SMARCB1 deficiency resulted in the miss-localization of the BAF complex, which leads to the inhibition of gene expression [127]. However, residual BAF complex can still maintain abnormal gene expression required for cancer progression. SMARCB1 deficiency has been detected in mRCC [128]. SMARCB1 deficiency leads to c-MYC pathway activation and increased DNA replication stress. Consequently, a highly inflammatory phenotype of mRCC with elevated replication stress was associated with the activation of the cGAS-STING innate immune pathway [128]. The potential therapeutic value of targeting the DDR enzyme, poly(ADP-ribose) polymerase (PARP) pathway, alone or in combination with platinum chemotherapy, has been suggested in mRCC [128].
The SWI/SNF protein complex mobilizes nucleosomes that are required for kinetochore localization and also modifies chromatin structure through the hydrolysis of ATP molecules [126]. PBRM1 gene encodes a bromodomain-containing protein (BAF180), which is a member of the DNA-targeting subunit of the SWI/SNF complex, recognizing acetylated histone residues, thereby localizing the complex in a specific chromatin region [129]. In addition to VHL, PBRM1 is the most common mutated gene in ccRCC. PBRM1 deficiency results in increased proliferation, migration, and colony formation of ccRCC [130]. Although the exact pathophysiological role of PBRM1 in ccRCC has not been established, PBRM1 deficiency disturbs p53-dependent chromatin regulation; therefore, ccRCC tumors are able to escape from p53-mediated surveillance [131]. Inhibition of PARP or ATR improved synthetically lethal in PRBM1-deficient cells and induced, rather than attenuate the cGAS-STING pathway to elicit immunomodulatory type I IFN response [132].
Adaptive immunotherapy using chimeric antigen receptor T-cells (CAR-T), genetically engineered T cells expressing antigen-specific non-major histocompatibility complex-restricted receptors is a popular approach in immunotherapy of cancers, including RCCs [133]. CD70 is a member of a type II transmembrane cell-surface protein of the TNF receptor family, which is highly expressed in RCCs and metastatic tissues [134]. Many patients with ccRCC showed abnormally high CD70 expression [135]. Therefore, targeting of CD70 function using CAR-T therapy thus far has only modest benefits, and positively evaluated [136,137].
7. Epigenetic Mechanisms in RCC
Epigenetic mechanisms strongly influence gene expression in mammalians. Several histone-modifying enzymes such as histone acetyltransferases (HATs) and histone deacetylases (HDACs) can regulate promoter activity and chromatin condensation processes by the addition or removal of methyl- and acetyl groups [138]. Gene expression profiles of cancer cells can be drastically modified by epigenetic mechanisms, which propagate tumor progression [139]. Mediators of systemic inflammation, such as nitric oxide, prostaglandins, or cytokines (IL-1β and IL-6), also modify gene expression profile through the activation of DNA methyltransferases (DNMTs) and HDACs, thereby influencing the behavior of cancer cells [140]. The inflamed TME can further change the epigenome in immune and epithelial cells, thereby influencing innate and adaptive immune responses [141].
Epigenetic mechanisms regulate different steps of RCC development since more than 200 genes have been identified with abnormal gene methylation [142]. RCC-related genes such as PBRM1, BAP1 and SETD2 are tightly regulated by histone modification and nucleosome and chromatin remodeling [143]. Recently, a predictive classifier was presented, accurately stratifying RCC patients’ risk based on methylation of five CpG-regions independent of standard clinical prognostic factors [144]. These studies revealed the activation of NF-κB and CCAAT/enhancer-binding protein (C/EBP) pathways, inducing the demethylation of inflammation-related genes in advanced ccRCC [145,146]. HIF, NF-κB and C/EBP pathways can strongly upregulate gene expression of CXC chemokines during cancer progression, thereby resulting in cancer-cell-intrinsic inflammation [147,148]. This inflammatory signal induces epigenetic changes in ccRCC cells, resulting in a “super-enhancer” formation in the genome at chromosome 4, forming a growth-regulated oncogene region [147]. HIF2α and NF-κB pathways also play an important role in RCC metastasis, due to the activation of metastasis-associated enhancer (MAE), which amplifies the expression of chemokine receptor CXCR4, thereby inducing pro-metastatic activity [148].
Lysine-specific histone demethylase 1 (LSD1) is a flavin-dependent monoamine oxidase, which targets mono- and di-methylated lysines on histone 3 (H3K4 and H3K9). LSD1 was detected in half of the patients with ccRCCs. Histone methylation status changed in LSD1-knockdown kidney cancer cells, suggesting an epigenetic control, probably through androgen receptor transcription factors [149]. High LSD1 expression suppresses H3K4 methylation in RCCs, which dysregulates the anti-proliferative check-point regulator p21 [150]. Consequently, inhibition of LSD1 function could restore p21 levels, thereby inducing G1/S cell-cycle arrest in RCCs [150]. Therefore, LSD1 inhibitors were developed such as raloxifene [151] or tranylcypromine derivates [152] and other preclinical tested compounds (ORY-1001 [153], SP2095 [150]). Although no clinical trials with LSD1 inhibitors have been published in patients with RCCs, some compounds are clinically validated in other solid cancer types (NCT02959437, NCT05268666, NCT02712905).
VHL deficiency can stabilize HIF2α function at tumor-specific gain enhancers thereby upregulating the expression of ccRCC-related oncogenes, such as ZNF395 [154]. Class II HDAC inhibitors were proposed to induce anti-tumor effects through the degradation of HIF isoforms [155]. Therefore, early clinical trials used panobinostat as a non-selective pan-HDAC inhibitor, a drug that suppresses the proliferation of immature epithelial progenitor cells in the adult kidney and enforces their differentiation. Later, monotherapy with HDAC inhibitor vorinostat, or combined therapy with mTOR inhibitor were more effective in patients with RCCs than panobinostat alone (NCT00278395, NCT00324740, NCT00324870). Hence, the numbers of clinical studies are limited, future investigations are needed to determine the effectiveness of epigenetic modulators in RCCs, which may improve patient survival.
8. Inflammatory Immune Cell Landscape of RCC
The immune cell landscape is regulated by tumor cell-presenting antigens, immune cell infiltration and immunomodulatory molecules in the TME, which are all engaged in a complex interplay between cancer cell internal and external factors, that influence tumor progression [156], (Figure 2). The immune cell landscape is an important factor contributing to the efficacy of cancer treatments, such as TKIs or ICIs, that target the TME of RCCs [25,157]. Numerous studies described ccRCC as an invasive cancer type with highly enriched T cell infiltrates. An immune profiling study of human ccRCC samples, identified T cells as the main immune cell population, followed by myeloid cells, NKs and B cells. However, in contrast to many other solid tumor types, infiltration of CD8+ and CD4+ T cells in ccRCC tumors is often linked to a poor prognosis and advanced tumor grade [25,158,159,160,161,162,163,164,165,166,167], highlighting T cell dysfunction during the progression of RCC. Interestingly, the presence of lymphocytes positive for PD-1 and tumor cells positive for programmed cell death-ligand 1 (PD-L1) and/or PD-L2 in the invasive margin of ccRCC was correlated with the shortest DFS, OS and higher risk of post-surgical cancer progression [168].
Dysfunctional maturation of DCs is characterized by the expression of lysosomal-associated membrane protein LAMP (DCLamp+), CD83 (DCCD83-) and MHC class II protein (DCMHCII,Low) Such DCs are located outside of tumor-associated tertiary lymphoid structures (NTLS-DC), particularly in the invasive margins of primary ccRCC tumors [168]. Increased density of NTLS-DC at the invasive margins was associated with higher levels of PD-L1+ and/or PD-L2+ and a bad prognosis, whereas high levels of CD8+ cells and low levels of DCs in tertiary lymphoid structures (TLS-DC) served as the strongest independent variable for a poor outcome. A higher density of TLS-DC in CD8+high groups was associated with a good prognosis [168]. Of note, patients’ outcomes were not significantly affected by the immunological profile of DCs in the tumor center. Tumor cells instigate a pro-inflammatory microenvironment, which leads to the recruitment and polarization of T cells towards an exhausted phenotype, potentially due to the presence of dysfunctional DCs in the TME outside of their canonical priming sites [168]. Shorter OS correlated to high densities of CD8+ T cells and DCLamp+ cells at metastatic sites, while higher densities of NKs were linked to improved survival [162]. Additionally, intratumoral PD-1 expression was correlated with tumor size, nuclear grade, TNM stage, coagulative tumor necrosis, a sarcomatoid phenotype and a higher risk of cancer-related death [169].
Several transcriptomic studies have been conducted to classify metastatic ccRCCs, and results identified four subtypes (ccrcc1 to 4). Crcc2 and 3 groups responded better to sunitinib therapy in comparison to ccrcc1 and 4 groups. The latter two included less differentiated cancers with upregulated MYC targets and displayed a hyper-methylated gene profile associated with polycomb stem-cell-like phenotype. The ccrcc4 group exhibited a sarcomatoid, pro-inflammatory, Th1-oriented phenotype with elevated B, T, and cytotoxic-cell-specific transcripts. The ccrcc4 group also displayed an immune suppressive TME with high expression of IL-10, lymphocyte activation gene 3 (Lag-3), PD-1 and PD-1 ligands [170]. Patients with ccRCC were also classified by immune- and inflammation-related gene signature using TCGA cohorts and determined three groups: (1) T cell-enriched (2) heterogeneously infiltrated and (3) non-infiltrated groups. The group with enriched T cells exhibited increased expression levels of granzyme B and INF-γ, PD-1 and cytotoxic T-lymphocyte-associated protein 4 (CTLA-4). This cluster corresponded to the worst survival. The T cell enriched group was further divided into two subgroups, called TCa and TCb. Gene expression of TCa was linked to metabolic and mitochondrial processes and better survival, while TCb was associated with cell cycle, extracellular matrix, cellular proliferation, and macrophage infiltration via stromal cells. Moreover, the infiltration of specific T cell subsets had prognostic significance, as Th17 cells and positive CD8+ T/Treg ratio compared to Th2 cells and Tregs corresponded to an improved prognosis [166,167]. Based on the immune cell profile of TME, RCCs were classified into three groups: (1) immune-silent, (2) immune-regulated and (3) immune-activated RCCs. The immune-silent group of RCCs displayed T cells with low activation markers while the immune-activated group showed oligoclonal cytotoxic T cells (CD8+/PD-1+), and immunoglobulin and mucin-domain containing-3 protein positive T cells (Tim-3+). The hallmarks of the immune-regulated group included an inflammatory microenvironment with the infiltration of polyclonal exhausted CD8+/PD-1+/Tim-3+/Lag-3+ tumor-infiltrating leukocytes (TIL) lacking in cytotoxicity, linked to infiltration of dysfunctional DCs. In addition, this group was enriched in CD4+/ICOS+/Glucocorticoid-induced tumor necrosis factor receptor-related protein (GITR)+/Helios+ T cells displaying a Treg-cell-like phenotype expressing CD25+/CD127-/Foxp3+ [168]. Ultimately, immune-regulated tumors exhibited the highest risk of post-surgical progression, however, also present obvious targets for ICIs [171]. PD-1 expression was widespread among several T cell subsets, as well as CD38, a cyclic ADP-ribosyl hydrolase 1 associated with inflammation, autoimmunity and hematologic malignancies [172], proposed as a new marker of T cell exhaustion [163]. The expression of further co-stimulatory receptors or markers of T cell exhaustion, however, varied among the subsets, such as Tim-3, tumor necrosis factor receptor superfamily 9 (4-1BB), CTLA-4 and tumor necrosis factor receptor superfamily member 4 (OX-40).
Tumor-associated macrophages (TAMs) are heterogeneous immune cell population in the TME. Clustering analysis of TAMs based on surface proteins showed different groups aligning to three branches. Apart from clusters corresponding to transitionary and circulating CD14+ and inflammatory CD16+ monocytes, these studies revealed a branch of mature TAMs only detectable in tumors of grade II or higher. The macrophages in these clusters exhibited diverse expression patterns, such as inflammatory marker CD38, as well as ones corresponding to a pro-tumor (CD163, CD204, CD206) and anti-tumor (CD169) phenotype. Furthermore, a CD68+/CD38+ macrophage population colocalized with exhausted CD8+/CD38+ T cells with elevated levels of PD-1 [163]. As the macrophage population also express PD-1 ligands such as programmed cell death 1 ligand 2 (PDCD1LG2) and PD-L1, it was proposed that these cells play a role in the establishment of an immunosuppressive TME in ccRCC [163]. Studies analyzing single-cell protein activity found that RCC-resident macrophages expressed a unique protein signature correlating with a higher risk of post-nephrectomy regression [173]. Profiling of TME genes in patients with advanced localized and metastatic RCC responding to ICI revealed a polarization towards an M1-like macrophage. Although M1 macrophages are responsible for the elimination of pathogens or tumor cells, M2 macrophages exhibit immune suppressive properties and a tendency towards tissue repair and tumor progression [174,175]. Remarkably, upregulation of IFN signaling and CD8+ T cell immune checkpoint genes, was paralleled by anti-inflammatory genes including V-set immunoregulatory receptor (VSIR), V-set and immunoglobulin domain containing 4 (VSIG4), PD-L2 and sialic acid-binding Ig-like lectin 10 (SIGLEC10). This result suggests a dual pro- and anti-inflammatory role of TAMs in RCC after ICI treatment, which probably leads to either immune suppression or resistance to ICIs [176]. Studies based on single-cell RNA sequencing of immune cell and T cell receptors along a pseudotime trajectory, respective of the tumor stage, showed early pro-inflammatory macrophages, while later in pseudotime or at advanced tumor stages, macrophages displayed an anti-inflammatory (M2-like) and CD8+ T cells an exhausted signature. A composite expression signature between terminally exhausted T cells and tumor macrophages was associated with worse OS across all tumor stages. Interestingly, the response to ICIs did not correlate to the composite signature [177]. Therefore, these studies propose an interplay between T cells and TAMs resulting in inflammation and defective immune response in RCC, leading to a reduced outcome.
In summary, several studies of the TME in RCC have revealed a crucial role in the interaction between immune cells, vasculature, and tumor cells in the development of tumors as well as in their responsiveness to and resistance to targeted therapy. Understanding the connections between key regulators of TME and interactions between various intratumoral cell types may increase therapeutic efficacy, particularly in RCC with more advanced stages.
9. Cytokine and Chemokine-Mediated Inflammatory Responses in RCC
9.1. IFNγ
Interferon-gamma (IFNγ) is an important cytokine that activates both the innate and adaptive immune systems. During the early phases of cancer, activated immune cells (NKs, CD8+, CD4+ T cells) secrete IFNγ, thereby modulating the activity of macrophages, DCs, Th1 CD4+helper T cells, and B cells to eliminate cancer cells. Therefore, IFNγ has anti-tumorigenic effects, which significantly suppress tumor growth. At the later step of tumor progression, IFNγ impairs secondary anti-tumor immune responses. However, long-term exposure to IFNγ can exert pro-tumorigenic effects inducing immunosuppression, angiogenesis and tumor cell proliferation [178]. IFNγ can upregulate the expression of programmed-death ligand 1 (PD-L1) on the surface of cancer cells, which binds its ligand (PD-1) on the activated CD8+ T cells. This cellular interaction leads to apoptosis of CD8+ T cells, thereby reducing anti-tumor CD8+T-cell cytotoxicity and creating an immunosuppressive TME. In addition, IFNγ also increases the expression of checkpoint inhibitors, such as indoleamine-2,3-dioxygenase 1 (IDO1) and CTLA-4, leading to enhanced tumor growth [179]. However, this was dependent on the RCC subtype, as PD-L1 in ccRCC showed a favorable effect on OS and DFS [180].
Other IFNγ-regulated gene is RBCK1, which showed markedly differential expression levels in cancer, and significantly correlated with tumor-infiltrating immune cells, tumor purity, and immune checkpoint molecules, such as PD-L1, CTLA-4, Lag-3, and T-cell immunoglobulin and ITIM domain (TIGIT) in pan-cancer samples [181]. High expression levels of RBCK1 displayed an immunosuppressive phenotype with decreased numbers of infiltrating CD4+ T cells, CD4+FOXP3+ Treg cells, M1 macrophages, and CD56bight/dim NK cells, and was associated with a worse prognosis in RCC patients. Furthermore, RBCK1 promotes p53 degradation through ubiquitination, which further enhances RCC progression.
Interestingly, tumor-associated activation of NF-κB represents a protective mechanism against IFNγ-mediated pathways. In the absence of NF-κB, IFNγ can trigger receptor-interacting protein kinase 1 (RIPK1)-dependent programmed necrosis (also called necroptosis) in resistant RCC cells [182]. Interestingly, treatment of cells with IFNγ/CD70 conjugated antibody could induce RIPK1-dependent necroptosis in RCC cells in the presence of NF-κB inhibitor bortezomib [183]. Although this result opens a gate for a new therapeutic approach, future studies are needed to determine the preclinical potential of this treatment.
FGF and FGFR1 are highly expressed in primary RCCs, and could enhance tumor cell proliferation, survival, and angiogenesis [184,185]. Activation of the FGFR signaling inhibits IFNγ-induced STAT1 phosphorylation and the expression of PD-L1, thus protecting cancer cells from the cytotoxic effects of CD8+ T cells [186], thereby supporting resistance to anti-cancer therapies [185]. In a mouse model of RCC, lenvatinib treatment inhibits both EGFR and FGFR signaling, and could restore the IFNγ-STAT1 signaling pathway and consequently prove the anti-cancer effect of the inhibitor [186,187].
In mammalian tissues, the essential amino acid tryptophan is metabolized by the kynurenine pathway. Kynurenine metabolites inhibit T cell proliferation and NK and Treg cell activities [188]. IDO1 is involved in this metabolic pathway, converting tryptophan to N-formyl-kynurenine. Interestingly, IDO1 is highly expressed in different cancer cell types, therefore IDO1 inhibitors were proposed as a potential therapeutic approach for the treatment of cancer [189]. IFNα therapy in RCC could further induce IDO1 activity, showing elevated tryptophan metabolic products in the urine of RCC patients [190], which indicates impaired immune responses. The combined therapy of IFNα and IDO1 inhibitors resulted in a strong beneficial effect on RCCs, because IFNα could properly stimulate the immune system, while the immunosuppressive activities mediated by IFNα/IDO1 signaling were blocked [189].
IFN effectors such as IFNGR1/2 and STAT1/2 are associated with metastatic RCCs and predict poor OS [191,192]. IFNγ downstream effector IFIT5-XRN1 is found to degrade tumor suppressor miRNA (miR-363 and miR-128) resulting in a pro-tumorigenic axis, which upregulates EMT drivers, such as Slug and zinc finger E-box binding homeobox 1 (ZEB1) [191,193]. Altogether, these results suggest that IFN-regulated genes can potentially be used as biomarkers to predict the efficacy of ICIs.
9.2. IL-2
Interleukin 2 (IL-2) stimulates the proliferation and cytolytic potential of NK and CD8+ T cells and promotes antibody release by B cells [194]. IL-2 binds and activates different receptor complexes, containing alpha (IL-2Rα, CD25), beta (IL-2Rβ, CD122), or common gamma chain receptors (γc, CD132). The CD8+ T cell expansion is mediated through the heterodimeric IL-2Rβγ complex while the expansion of immunosuppressive CD4+ CD25+ Treg cells is regulated by the IL-2Rαβγ complex. PEGylation has been used to conjugate polyethylene glycol (PEG) to IL-2. The PEGylated form of IL-2 can bind these receptor complexes and improves CD8+ T cell induction with minimal toxicity [194]. Although PEGylated IL-2 has been tested in clinical trials and the therapeutic effects were compared to nivolumab to cabozantinib or sunitinib, results are still pending (NCT03729245).
GI101 is a molecular agent comprising the extracellular domain of CD80 together with a long-acting IL-2 variant that preferentially binds to the IL-2Rβ complex [195]. In a preclinical setting, GI101 strongly stimulates CD8+ T and NK cell proliferation without affecting Treg cell population. GI101 elicits improved restoration of immune functions compared to a CD80-Fc without IL-2-associated toxicity [196]. A phase 1/2, open-label, dose-escalation and expansion study has been designed to evaluate GI101 as a single agent or in combination with pembrolizumab, lenvatinib or local radiotherapy in advanced and metastatic solid tumors (NCT04977453).
9.3. IL-12
Antigen-presenting cells produce interleukin 12 (IL-12) in response to infections, which promotes CD4+ T cell differentiation into Th1 cells. IL-12 has a positive synergistic proliferative effect on pre-activated NK and T cells and enhances independently and/or synergistically the cytolytic activity of both immune cell types by upregulating genes encoding cytotoxic cell granule-associated proteins [197,198]. Although preclinical studies proposed IL-12 as a potent immunotherapeutic agent in cancer therapy, disappointing clinical study was reported about the human recombinant IL-12 (rIL-12) treatment, and the negative result was explained by the inability of rIL-12 to reach the TME with effective doses [199]. The rIL-12 treatment also resulted in high levels of pro-inflammatory cytokine production, thereby inducing adverse side effects [200]. Therefore, novel approaches were developed to obtain biologically relevant rIL-12 treatment. Immunocytokine fusion protein of NHS-IL12 was designed, which consists of the tumor necrosis-targeting NHS76 antibody and fused to the human IL-12 heterodimer. This strategy was successful to deliver IL-12 to the necrotic region of the tumor with limited systemic exposure and associated toxicity [201]. Although NHS-IL12 was initially well tolerated by patients, a later study in combination with the anti-PD-L1 antibody avelumab was terminated (NCT02994953), due to the lack of observable efficacy. IL-12 fusion protein with humanized antibody BC1 was developed to target the ED-B splice variant of fibronectin, which is only expressed in the tumor-associated vasculature. Although the drug was well tolerated in the phase I evaluation of RCC, the cohort size was low (n=2) [202]; therefore, follow-up clinical trials are necessary for the final conclusion.
Tumor-targeted oncolytic viruses represent a long-promised revolution in cancer immunotherapy, because viruses can selectively target and eliminate tumor cells. The virus-infected cancer cells (or cancer lysates) release tumor antigens and intracellular proteins, which induce anti-tumor immune responses through IL-12-mediated pathways [203]. Genetically modified adenovirus with triple gene deletions is a safety tumor-sensitive virus vector (Ad-TD-LUC) for cancer cell targeting, and it was used to deliver a modified IL-12 (nsIL-12) to pancreatic cancer [203]. nsIL-12 lacks the N-terminal signal peptide, thereby preventing IL-12 secretion from the intact cell, but it can be released by necrotic cancer cells. Adenovirus-nsIL-12 treatment significantly improved outcomes, compared to mice treated with adenovirus-IL-12, which succumbed to IL-12-induced toxicity [203]. Further studies are necessary to evaluate this treatment in RCCs.
9.4. IL-6/JAK/STAT3
Interleukin-6 (IL-6) is required for inflammatory reactions induced by TNF, IL-1, INF, and bacterial or viral infections [204]. IL-6 is produced by tumor cells and cells in the TME, such as fibroblasts, T and B cells, as well as myeloid, endothelial, and mesangial cells [205,206,207]. IL-6 binds IL-6 receptor (IL-6R), forming IL-6R/gp130 receptor complex which activates Janus-kinase (JAK). JAK subsequently phosphorylates tyrosine residues of gp130, thereby resulting in the activation of the gp130 Tyr759-derived Scr homology 2 domain-containing protein tyrosine phosphatase-2 (Scr-2)/extracellular-signal-regulated kinase (ERK)/mitogen-activated protein kinase (MAPK) pathway and the gp130 YXXQ-mediated JAK/ STAT pathway [205].
During cancer progression, JAK/STAT3 signaling enhances tumor proliferation, survival, invasion, immune evasion, and metastasis [208]. STAT3 was identified as the major mediator of IL-6-dependent cancer cell proliferation in RCCs [209]. In genomic and transcriptional profiling studies of RCCs, JAK/STAT3 gene signature was associated with a poor outcome and was correlated with an increased immunosuppressive Treg cell score and served as an independent prognostic factor in RCCs [210].
In the kidney, different cell types can synthesize IL-6, which confers a pro-inflammatory phenotype to kidney diseases [211,212,213,214,215,216,217,218,219,220,221]. Serum levels of IL-6 correlated with the tumor stage of the RCC and was therefore considered a prediction factor of disease recurrence, progression, and OS and disease-specific (DS) survival [222,223,224,225,226,227,228]. High expression levels of IL-6 were detected in endothelial and vascular smooth muscle cells, indicating the vascular origin of IL-6 in patients with RCC. Furthermore, tumor-associated CD4+ T cells can also release IL-6 in ccRCC [226,229].
Hypoxia increases IL-6 production through the NADPH oxidase 4 (Nox4) signaling pathway in RCCs [23]. The HIF1-mediated VEGF expression is modulated by STAT3 signaling in RCCs [35]. High levels of IL-6 promote tumor invasiveness, and it is strongly associated with reduced response to sunitinib and pazopanib in patients with metastatic RCC [230]. This tumor resistance was explained by the ability of IL-6 to induce the phosphorylation of tumor AKT, mTOR and NF-κB signaling [231]. Activation of p21-activated kinase 1 (PAK1) influences IL-6/NF-κB activation, leading to a stem-like phenotype and consequent sunitinib resistance in RCCs [232]. An alternative mechanism also promotes the proliferation of RCCs through the downregulation of hepatocyte cell adhesion molecule (hepaCAM). Activation of IL-6/STAT3 signaling results in epigenetic changes (hypermethylation) in hepaCAM promoter and consequent inhibition of gene expression [233].
In VHL-deficient RCCs, IL-6 production is increased, which propagates macrophage migration to the tumor site and polarization toward an M2-like phenotype [55]. IL-6-activated macrophages release TGFβ; therefore, renal tubular epithelium develops an adenoma-like phenotype. IL-6 also activates human CD14+ peripheral blood nuclear cells through the STAT3 pathway producing CCL18 and TGFβ, which leads to the induction of the EMT with tumor-promoting effects. Therefore, tumorigenic EMT seems to be promoted by a reciprocal immunosuppressive interplay between tumor cells and macrophages, activated by IL-6/JAK/STAT3 signaling during the early phases of tumor progression [55]. In ccRCC, it has been proposed that cytotoxic T cell activation and consequent response to immunotherapy are probably impaired by enhanced IL-6/JAK/STAT3, IFNα, IFNγ signaling routes and RIPK2-mediated signaling [234,235]. Altogether, these results suggest that direct blockade of IL-6 or its downstream effectors could be a potential strategy to cure RCCs.
Siltuximab is a chimeric mouse–human monoclonal antibody that binds and blocks IL-6 functions. Siltuximab has been tested in a clinical trial (NCT00265135) with beneficial effects, stabilizing progressive metastasis in half of the patients with RCCs [236]. IL-6R and JAK inhibitors have been used in the treatment of inflammatory diseases, including myeloproliferative neoplasms and the management of adverse effects of CAR-T cells [207]. Tocilizumab is a recombinant humanized monoclonal antibody targeting IL-6R, approved for the treatment of arthritis [237,238,239,240] and giant cell arteritis [241], CAR-T cell-induced cytokine-release syndrome [242], systemic sclerosis-associated interstitial lung disease [218,243,244], and severe acute respiratory syndrome coronavirus 2 (SARS-Cov-2 infection) [245,246]. Tocilizumab has been proposed to support the anti-tumor effect of IFNα treatment in RCCs by inhibiting the IL-6-mediated expression of the suppressor of cytokine signaling 3 (SOCS3) [247,248]. In a xenograft mouse model, tocilizumab therapy was combined with TKI and beneficial inhibitory effects were observed in tumor growth and vascularization, compared to TKI monotherapy, implicating a positive clinical evaluation [231].
9.5. IL-1β
Interleukin 1 beta (IL-1β) is an inflammatory agonist that has many immunomodulatory effects in the kidney [249]. Both tumor and stromal cells can release IL-1β, generating an immunosuppressive TME enriched in TAMs, myeloid-derived suppressor cells (MDSCs), and Treg cells [250,251,252]. IL-1β in the TME reduced tumor growth, invasiveness and angiogenesis, and facilitated checkpoint inhibition [250,251,252,253]. IL-1β-mediated expansion of MDSCs in ccRCC tumor is also associated with elevated intratumoral levels of angiogenic IL-8, CXCL5 and chemotactic C-C motif chemokine 3 (also known as MIP-1α) [254]. A recent study showed increased efficacy of TKI, cabozantinib, or anti-PD-1 checkpoint-inhibition, when combined with an anti-IL-1β mAb in a heterotopic mouse model of RCC [255]. The number of intratumoral immunosuppressive MDSCs was strongly TAMs differentiated toward an anti-cancer (M1-like) phenotype [255].
Additionally, pathogen- and damage- associated molecular patterns (PAMPs, DAMPs), and NLR family pyrin domain containing 3 (NRLP3)-inflammasome activation in TME may cause the production of IL-1β. Although IL-1β levels in RCC tumors are frequently increased, it is still an open question whether this inflammasome activation may affect the growth and metastasis of RCC [249]. Drugs used in RCC patients may further drive inflammasome activation [249].
9.6. TNFα
Tumor necrosis factor α (TNFα) is a key mediator of cancer-related inflammation in RCCs, promoting a pro-tumoral milieu by inducing the production of reactive oxygen intermediates, cyclooxygenases, matrix metalloproteinases (MMPs), and cytokines [256,257]. Serum and intratumoral levels of TNFα are strongly elevated in RCCs, compared to other cancer types, and correlated with tumor size [258,259]. Gene expressions of chemokine receptors (CXCR2 or CXCR3) are induced by TNFα and associated with a worse outcome in a high-risk cohort of patients with ccRCC. TNFα also enhances PI3K/AKT signaling and GSK-3β signaling-dependent tumor necrosis factor α-induced protein 8 (TNFAIP8) and MMP-9 expression. The elevated mRNA expression of the EMT-associated transcriptional factors Slug and ZEB1 and increased expression of vimentin, a mesenchymal marker over epithelial marker E-cadherin, resulting in enhanced migratory and invasive potential [260,261,262,263,264,265]. The co-expression of TNFα with MMP-9 or cancer stem cell marker CD44 in metastatic RCC tumors increases the resistance to sunitinib treatment [259,266]. TNFα also has an indirect effect on adaptive immune response by inhibiting T cell activity in the TME. Tumor-derived gangliosides inhibit anti-tumor efficacy via altering TNF-induced NF-κB activation in T cells and inducing T cell apoptosis [267]. TNFα has anti-proliferative effects in RCC when combined with Th2 cytokines IL-4 or IFNγ, highlighting its complex role in the regulation of immune response [268,269,270]. In the early 2000s, a series of phase II clinical studies investigated infliximab, a chimeric anti-TNFα monoclonal antibody. The trials first demonstrated promising results in low and high-dose treatment protocols of chemokine-resistant patients but eventually failed in combination therapy with first-line TKI treatment, producing side effects without observable improvement [271,272]. Studies on TNFα in RCC therapy have stepped back in favor of TKIs, mTOR inhibitors, and other approaches.
9.7. TGFβ
Transforming growth factor beta (TGFβ) signaling is involved in a variety of physiological and pathophysiological processes, including proliferation, migration, differentiation, apoptosis, embryonic development, tissue repair, angiogenesis and inflammation [273]. In the canonical Smad-dependent pathway, TGFβ ligands bind to serine/threonine kinase receptor TGFβ receptor type II (TβRII), leading to the recruitment and phosphorylation of TGFβ receptor type I (TβRI, ALK5, ALK5-Full Length/ALK5-FL). Downstream effectors, Smad-family member 2 and 3 (Smad2/3) are phosphorylated, resulting in a complex formation including Smad4, and activation of target genes [274,275,276]. The non-canonical (pathway involves translocation of the TβRI-intracellular domain (TβRI-ICD, ALK5-ICD) into the nucleus, independent of Smad [277]. TGFβ is considered as an anti-inflammatory cytokine in the context of renal inflammation [278]. However, TGFβ regulates renal fibrotic diseases as well, inducing the production and abnormal accumulation of extracellular matrix components [279,280,281,282,283,284,285,286]. TGFβ signaling is quite complex in kidney diseases since TβRII deficiency in kidney fibroblasts and tubular epithelial cells impairs TGFβ/Smad3-mediated kidney fibrosis, but simultaneously enhances NF-κB-mediated renal inflammation via upregulation of IL-1β and TNFα-mediated pathways [287].
Elevated serum levels of TGFβ are associated with metastatic disease [288]. Tissue expression of TGFβ correlates to tumor stage, histological type and nuclear grade [289]. Increased activity of TGFβ-mediated pathways, as a prognostic signature, was detected in patients with ccRCC [275]. TGFβ/Smad2/3 signaling leads to improved invasiveness of RCCs [290]. TβRII and TβRIII deficiency have been associated with primary tumor formation and metastases of RCCs, respectively [291,292]. A ubiquitin-dependent degradation of TβRII was observed in advanced RCCs which is regulated by the E3 ligase Smad-ubiquitination regulatory factor 2 (Smurf2), thereby dysregulating TGFβ signaling pathways [293]. Cytoplasmic TβRIII domain-induced apoptosis functions through a TβR/Smad-independent manner involving the p38-MAPK pathway [294]. Treatment of RCC cell lines with recombinant TGFβ (rTGFβ) leads to the downregulation of cadherin 1 and claudin 1 (epithelial markers), and the upregulation of vimentin, ZEB1 and zinc-finger transcription factor Snai1 (mesenchymal markers) in a TGFβ/Smad-dependent manner, highlighting the contribution to EMT formation. rTGFβ treatment could downregulate TβRII and upregulate TβRI expressions in ccRCC, which is probably consistent with the link between enhanced tumor growth and TβRII deficiency [295].
Analyzing ccRCC cohorts, an increased tumor immune infiltration was observed and correlated with activation of TGFβ pathway [296]. Suppression of TGFβ-induced paired box gene 2 (PAX2) or SMAD proteins binding to the PAX2 promoter has been suggested to be involved in EMT of RCCs [297]. Integrin signaling potentiated TGFβ1-induced E-cadherin depletion in vitro, mediated by the EMT-associated transcription factor Snai1 [298]. Forkhead activin signal transducer 1 (FAST1, FOXH1) is upregulated in RCC and mediates activation of the TGFβ/Smad signaling pathway, leading to increased cell viability, proliferation, migration, invasion, EMT, and reduced apoptosis as well as increased tumor growth [299]. Neutralizing TGFβ in a heterotopic cancer model decreased the size of tumors and inhibited vascularization [300]. TGFβ-activated kinase 1 (TAK1)-inhibitors could also inhibit cancer cell survival by impairing NF-κB activation and inducing apoptosis [301]. High mRNA levels of IL-10 and TGFβ were detected in the peripheral blood of metastatic RCC patients compared to the healthy control group. Although various T cell subsets could release TGFβ, mainly monocytes were responsible for TGFβ production in the peripheral blood, and this was correlated with favorable prognosis for patients with metastatic RCCs [302]. Exosomes released from primary tumor cells also contain TGFβ, thereby attracting tumor-infiltrating NKs by regulating the TGFβ/Smad pathway [303]. In a preclinical study, tumor-reactive CD8+ T cells were isolated from RCC patients, activated with autologous immature DCs and tumor lysate, and infected with a TβRIIDN retrovirus to inhibit TGFβ signaling. RCC patient-derived peripheral blood mononuclear cells were injected into immunodeficient RCC-xenografted mice, followed by the transfer of autologous patient-derived TGFβ-resistant CD8+ T cells. The autologous TGFβ-insensitive CD8+ T cells displayed cytotoxic activity and decreased tumor burden and lung metastasis, and increased survival [304].
Targeting of TGFβ was extensively investigated in various kidney diseases, such as focal segmental glomerulosclerosis and diabetic nephropathy [285,305,306]. Several studies suggested an immune suppressive role of TGFβ; therefore, many agents blocking downstream effectors of TGFβ have been investigated in both preclinical and clinical studies. Low-dose chemotherapeutic drugs initially inhibited TβRI expression on tumor cells but promoted IFNγ secretion and T-cell infiltration into tumors. At later stages, this effect was reversed, as the TβRI and Smad expression were increased in cancer and T cells [307]. Salinomycin is a monocarboxylic polyether antibiotic against Gram-positive bacteria. Moreover, salinomycin also kills cancer stem cells and potentially inhibits cancer growth and metastasis through DNA damage and inhibition of TGFβ-mediated EMT [308]. Salinomycin also inhibits the proliferation and viability of RCC cell lines. TβRI inhibitor reduces tumor cell invasion by attenuating the TGFβ/Smad3 pathway [309].
Two types of estrogen receptors (ERα, ERβ) regulate cancer development and interestingly, RCC cell lines only expressed the ERβ isoform [310]. Increased ERβ expression was detected during RCC development, and this was associated with worse survival for RCC patients [310]. In a mouse model of RCC, ERβ supported cancer cell proliferation, migration and invasion, and could affect the expression of TGFβ/Smad3 signaling to control cancer cell invasion [311]. In line with these results, anti-estrogen (ICI 182,780) and selective ERβ antagonist (PHTPP) could effectively inhibit tumor growth and invasion in an orthotopic mouse model of RCC [310].
Pirfenidone is a pyridone compound with anti-fibrotic effects due to the downregulation of TGFβ, PDGF and type I collagen synthesis [312,313,314]. Therefore, pirfenidone has been proposed to treat patients with kidney and pulmonary fibrosis. Pirfenidone inhibits TGFβ-mediated EMT, migration, proliferation, and invasion of RCC cells through decreasing Smad2/3 phosphorylation. In a subcutaneous RCC model, pirfenidone could inhibit tumor growth and recruitment of MDSCs [315].
Fresolimumab is a pan-TGF-targeting neutralizing monoclonal antibody, which was subjected to clinical trials, including advanced RCCs and malignant melanoma (NCT00923169) [316]. Fresolimumab displayed an acceptable safety profile and anti-tumor activity [317]. However, many patients did not exhibit increased OS, due to the reduced expression of activating surface proteins in NK cells [318]. Another anti-TGFβ antibody NIS739 was also studied in a phase I/Ib trial as a single agent or in combination with spartalizumab, an anti-PD-1 antibody (PDR001) in patients with RCC and advanced malignancies. Results showed synergic effects in attenuating immunosuppressive responses to α-PD-1 (NCT02947165) [318,319]. Altogether, these results suggest that a TGFβ signalosome could be an attractive therapeutic target in the advanced stages of RCC.
9.8. Chemokine Receptors
Activation of chemokine receptors (CXCR1 and CXCR2) has pro-angiogenic and pro-inflammatory effects, inducing several signaling pathways, such as the protein kinase C, phospholipase C, PI3K/AKT/mTOR, RAS/RAF/MEK/ERK and NF-kB pathways, promoting tumor cell survival, proliferation, and dissemination [320]. The growth-related oncogene (GRO) subgroup CXCL1/GRO-a, CXCL2/GRO-b, CXCL3/ GRO-c, preferentially bind to CXCR2, while CXCL8 binds CXCR1. These interactions increase neutrophil recruitment to the tumor site and consequent VEGFA production and angiogenesis [321]. Blockade of CXCR1/2-ELR+CXCL interactions in malignancies appears to be an attractive therapeutic approach. Reparixin is a CXCR1/2 inhibitor and showed promising results in pilot studies of early and metastatic breast cancer and was evaluated in combination with paclitaxel [322] (NCT02370238).
Analysis of pan-cancer databanks showed that RCCs can expose the highest amounts of C-X-C motif chemokine receptors CXCR1 and CXCR2 [323]. High levels of chemokine expression correlated to the significantly worse OS and progression-free survival (PFS) in RCC patients [323]. Chemokine receptor expression in RCCs has a significant impact on the formation of a pro-inflammatory landscape in the TME. Dufies et al. investigated the effects of various CXCR1/2 blocking agents in ectopic models of RCC and identified compound C29 to inhibit primary tumor cell growth and ELR+CXCL-mediated proliferation and migration of endothelial cells [323]. The impact of C29 was equal in RCC cells that had not been exposed to TKIs and those that had, indicating a common method to prevent angiogenic escape. Further characterization of C29 effects in a pre-clinical study will unveil its potential in the therapy of RCC [323].
CXCR4 binds macrophage migration inhibitory factor (MIF), a factor implicated in the control of innate immunity, as well as CXCL12, a molecule well-known for its function in immunological surveillance, inflammatory response, tissue homeostasis, and tumor development and metastasis [324,325]. CXCR4 expression levels especially increased during the invasive phases of cancer involving EMT formation. In patients with RCC, high CXCR4 expression correlates with worse OS and PFS [326]. High-grade RCC samples showed a signature of CXCR4, CXCR7, VIM, CDH2 and ZEB1, while metastatic samples had even higher expression levels for VIM, CXCR4 and FN1 [327]. VHL deficiency or aberrant β-Catenin (CTNNB1) expression induced CXCR4 expression in RCCs. CTNNB1 is a component of the Wnt signaling pathway, which regulates tumor growth and metastasis through the expression of ICAM-1, VCAM-1, CXCR4, and CCL18 [328].
In mouse models, interfering with the CXCR4–CXCL12 axis promoted CD8+ T cell infiltration and reduced CXCL12-mediated migration of lymphocytes and MDSCs to the TME, as well as inhibited tumor revascularization following anti-angiogenic therapy [320]. Multiple studies currently evaluate X4P-001 (mavorixafor), a CXCR4 inhibitor, in combination with standard therapy options. The combined therapy of X4P-001 with VEGF inhibitor axitinib is well tolerated in patients with advanced RCC and demonstrates encouraging median PFS [329] (NCT02667886). A phase 1b trial of mavorixafor and ICI nivolumab in advanced RCC patients with no prior response to nivolumab monotherapy showed promising tolerability and anti-tumor activity [330].
10. Conclusions
RCC can develop from different genetic insults, including as a consequence of kidney injury. Many hallmarks of RCC are mediated by inflammation, including tumor cell proliferation, migration, angiogenesis, and tumor spread. Inflammation can be also due to extrinsic factors, such as obesity, infection, and exposure to pollutants, bacteria, viruses, and tobacco, which can increase the susceptibility to developing RCC. Although eradication of inflammation-causing factors as earlier as possible is an optimal approach, this requires robust diagnostic markers. Inflammation is significantly reflected by quantitative and qualitative alterations in RCC’s immune cell landscape, such as immune cell infiltration and immunosuppression. Testing systemic inflammation markers in blood and urine samples of RCC patients, including circulating leukocytes, immune cells and inflammatory cytokines, provide important prognostic data. However, predictive accuracy depends on the patient profile, which has to be taken into the account, i.e., patients who already received immunotherapy. In this case, pre- and post-therapeutic assessment of inflammation-related markers can better predict response and survival in patients undergoing treatment. Single-cell RNA-sequencing approaches in clinics offer many benefits; it can determine the source, origin and type of inflammation-inducing cells and identify interconnected resistance pathways, thereby proposing powerful risk-stratification models. Due to the differences between mouse and human RCC oncogenesis, many stochastic mouse RCC models do not mimic all the stages of tumor development and fail to form metastases. Although patient-derived xenograft models retain many characteristics of human patients and serve as valuable tools to dissect specific mechanisms, they lack an immune system. Therefore, additional experimental studies in the field are required to translate the findings from animal models to human RCC. Understanding the intricate inflammatory pathways controlling tumor cell or TME plasticity may help to identify new therapeutic targets in RCC in order to better design personalized medicine approaches.
Author Contributions
L.K., M.M. and E.M.-B. wrote the manuscript. A.B., J.A., H.-J.A. and T.G. contributed to the writing of manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Bayerisches Landesamt für Gesundheit und Lebensmittelsicherheit (BLGL), (project number 15-25) to E.M-B. and A.B., Deutsche Forschungsgemeinschaft; CRC TRR152 to T.G.; L.K. and M.M. were recipients of a fellowship from the Förderprogramm für Forschung und Lehre (FöFoLe) of the LMU, Munich, respectively. H-J.A. received support from the Deutsche Forschungsgemeinschaft (AN372/30-1 and 32-1). J.A. received support from the Deutsche Forschungsgemeinschaft (AN 391/4-1).
Acknowledgments
Figures were created with www.biorender.com.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hsieh, J.J.; Purdue, M.P.; Signoretti, S.; Swanton, C.; Albiges, L.; Schmidinger, M.; Heng, D.Y.; Larkin, J.; Ficarra, V. Renal cell carcinoma. Nat. Rev. Dis. Primers 2017, 3, 17009. [Google Scholar] [CrossRef] [PubMed]
- Muglia, V.F.; Prando, A. Renal cell carcinoma: Histological classification and correlation with imaging findings. Radiol. Bras. 2015, 48, 166–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, U.K.; Ramskov, S.; Bjerregaard, A.M.; Borch, A.; Andersen, R.; Draghi, A.; Donia, M.; Bentzen, A.K.; Marquard, A.M.; Szallasi, Z.; et al. Tumor-Infiltrating T Cells from Clear Cell Renal Cell Carcinoma Patients Recognize Neoepitopes Derived from Point and Frameshift Mutations. Front. Immunol. 2020, 11, 373. [Google Scholar] [CrossRef] [PubMed]
- Peired, A.J.; Lazzeri, E.; Guzzi, F.; Anders, H.J.; Romagnani, P. From kidney injury to kidney cancer. Kidney Int. 2021, 100, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Peired, A.J.; Antonelli, G.; Angelotti, M.L.; Allinovi, M.; Guzzi, F.; Sisti, A.; Semeraro, R.; Conte, C.; Mazzinghi, B.; Nardi, S.; et al. Acute kidney injury promotes development of papillary renal cell adenoma and carcinoma from renal progenitor cells. Sci. Transl. Med. 2020, 12, eaaw6003. [Google Scholar] [CrossRef]
- Qian, C.N.; Huang, D.; Wondergem, B.; Teh, B.T. Complexity of tumor vasculature in clear cell renal cell carcinoma. Cancer 2009, 115, 2282–2289. [Google Scholar] [CrossRef]
- Mertz, K.D.; Demichelis, F.; Kim, R.; Schraml, P.; Storz, M.; Diener, P.A.; Moch, H.; Rubin, M.A. Automated immunofluorescence analysis defines microvessel area as a prognostic parameter in clear cell renal cell cancer. Hum. Pathol. 2007, 38, 1454–1462. [Google Scholar] [CrossRef]
- Choueiri, T.K.; Motzer, R.J. Systemic Therapy for Metastatic Renal-Cell Carcinoma. N. Engl. J. Med. 2017, 376, 354–366. [Google Scholar] [CrossRef]
- Escudier, B.; Eisen, T.; Stadler, W.M.; Szczylik, C.; Oudard, S.; Siebels, M.; Negrier, S.; Chevreau, C.; Solska, E.; Desai, A.A.; et al. Sorafenib in advanced clear-cell renal-cell carcinoma. N. Engl. J. Med. 2007, 356, 125–134. [Google Scholar] [CrossRef]
- Motzer, R.J.; Hutson, T.E.; Tomczak, P.; Michaelson, M.D.; Bukowski, R.M.; Rixe, O.; Oudard, S.; Negrier, S.; Szczylik, C.; Kim, S.T.; et al. Sunitinib versus interferon alfa in metastatic renal-cell carcinoma. N. Engl. J. Med. 2007, 356, 115–124. [Google Scholar] [CrossRef]
- Escudier, B.; Pluzanska, A.; Koralewski, P.; Ravaud, A.; Bracarda, S.; Szczylik, C.; Chevreau, C.; Filipek, M.; Melichar, B.; Bajetta, E.; et al. Bevacizumab plus interferon alfa-2a for treatment of metastatic renal cell carcinoma: A randomised, double-blind phase III trial. Lancet 2007, 370, 2103–2111. [Google Scholar] [CrossRef]
- Sternberg, C.N.; Davis, I.D.; Mardiak, J.; Szczylik, C.; Lee, E.; Wagstaff, J.; Barrios, C.H.; Salman, P.; Gladkov, O.A.; Kavina, A.; et al. Pazopanib in locally advanced or metastatic renal cell carcinoma: Results of a randomized phase III trial. J. Clin. Oncol. 2010, 28, 1061–1068. [Google Scholar] [CrossRef]
- Rini, B.I.; Escudier, B.; Tomczak, P.; Kaprin, A.; Szczylik, C.; Hutson, T.E.; Michaelson, M.D.; Gorbunova, V.A.; Gore, M.E.; Rusakov, I.G.; et al. Comparative effectiveness of axitinib versus sorafenib in advanced renal cell carcinoma (AXIS): A randomised phase 3 trial. Lancet 2011, 378, 1931–1939. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Kadife, E.; Myers, M.; Kannourakis, G.; Prithviraj, P.; Ahmed, N. Determinants of resistance to VEGF-TKI and immune checkpoint inhibitors in metastatic renal cell carcinoma. J. Exp. Clin. Cancer Res. 2021, 40, 186. [Google Scholar] [CrossRef]
- Wu, Y.; Fu, X.; Zhu, X.; He, X.; Zou, C.; Han, Y.; Xu, M.; Huang, C.; Lu, X.; Zhao, Y. Prognostic role of systemic inflammatory response in renal cell carcinoma: A systematic review and meta-analysis. J. Cancer Res. Clin. Oncol. 2011, 137, 887–896. [Google Scholar] [CrossRef] [PubMed]
- Miyata, Y.; Koga, S.; Nishikido, M.; Noguchi, M.; Kanda, S.; Hayashi, T.; Saito, Y.; Kanetake, H. Predictive values of acute phase reactants, basic fetoprotein, and immunosuppressive acidic protein for staging and survival in renal cell carcinoma. Urology 2001, 58, 161–164. [Google Scholar] [CrossRef] [PubMed]
- Roxburgh, C.S.; McMillan, D.C. Role of systemic inflammatory response in predicting survival in patients with primary operable cancer. Future Oncol. 2010, 6, 149–163. [Google Scholar] [CrossRef] [PubMed]
- Ramsey, S.; Lamb, G.W.; Aitchison, M.; McMillan, D.C. Prospective study of the relationship between the systemic inflammatory response, prognostic scoring systems and relapse-free and cancer-specific survival in patients undergoing potentially curative resection for renal cancer. BJU Int. 2008, 101, 959–963. [Google Scholar] [CrossRef]
- Gu, L.; Li, H.; Gao, Y.; Ma, X.; Chen, L.; Li, X.; Zhang, Y.; Fan, Y.; Zhang, X. The association of platelet count with clinicopathological significance and prognosis in renal cell carcinoma: A systematic review and meta-analysis. PLoS ONE 2015, 10, e0125538. [Google Scholar] [CrossRef]
- Motzer, R.J.; Mazumdar, M.; Bacik, J.; Berg, W.; Amsterdam, A.; Ferrara, J. Survival and Prognostic Stratification of 670 Patients with Advanced Renal Cell Carcinoma. J. Clin. Oncol. 1999, 17, 2530. [Google Scholar] [CrossRef] [Green Version]
- Greten, F.R.; Grivennikov, S.I. Inflammation and Cancer: Triggers, Mechanisms, and Consequences. Immunity 2019, 51, 27–41. [Google Scholar] [CrossRef] [PubMed]
- Fox, P.; Hudson, M.; Brown, C.; Lord, S.; Gebski, V.; De Souza, P.; Lee, C.K. Markers of systemic inflammation predict survival in patients with advanced renal cell cancer. Br. J. Cancer 2013, 109, 147–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fitzgerald, J.P.; Nayak, B.; Shanmugasundaram, K.; Friedrichs, W.; Sudarshan, S.; Eid, A.A.; DeNapoli, T.; Parekh, D.J.; Gorin, Y.; Block, K. Nox4 mediates renal cell carcinoma cell invasion through hypoxia-induced interleukin 6- and 8-production. PLoS ONE 2012, 7, e30712. [Google Scholar] [CrossRef] [PubMed]
- Reuter, S.; Gupta, S.C.; Chaturvedi, M.M.; Aggarwal, B.B. Oxidative stress, inflammation, and cancer: How are they linked? Free Radic. Biol. Med. 2010, 49, 1603–1616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Díaz-Montero, C.M.; Rini, B.I.; Finke, J.H. The immunology of renal cell carcinoma. Nat. Rev. Nephrol. 2020, 16, 721–735. [Google Scholar] [CrossRef] [PubMed]
- Godwin, J.L.; Zibelman, M.; Plimack, E.R.; Geynisman, D.M. Immune checkpoint blockade as a novel immunotherapeutic strategy for renal cell carcinoma: A review of clinical trials. Discov Med 2014, 18, 341–350. [Google Scholar] [PubMed]
- Atkins, D.; Ferrone, S.; Schmahl, G.E.; Störkel, S.; Seliger, B. Down-regulation of HLA class I antigen processing molecules: An immune escape mechanism of renal cell carcinoma? J. Urol. 2004, 171, 885–889. [Google Scholar] [CrossRef]
- Dunker, K.; Schlaf, G.; Bukur, J.; Altermann, W.W.; Handke, D.; Seliger, B. Expression and regulation of non-classical HLA-G in renal cell carcinoma. Tissue Antigens 2008, 72, 137–148. [Google Scholar] [CrossRef]
- Seliger, B.; Atkins, D.; Bock, M.; Ritz, U.; Ferrone, S.; Huber, C.; Störkel, S. Characterization of human lymphocyte antigen class I antigen-processing machinery defects in renal cell carcinoma lesions with special emphasis on transporter-associated with antigen-processing down-regulation. Clin. Cancer Res. 2003, 9, 1721–1727. [Google Scholar]
- Wang, T.; Lu, R.; Kapur, P.; Jaiswal, B.S.; Hannan, R.; Zhang, Z.; Pedrosa, I.; Luke, J.J.; Zhang, H.; Goldstein, L.D.; et al. An Empirical Approach Leveraging Tumorgrafts to Dissect the Tumor Microenvironment in Renal Cell Carcinoma Identifies Missing Link to Prognostic Inflammatory Factors. Cancer Discov. 2018, 8, 1142–1155. [Google Scholar] [CrossRef] [Green Version]
- Donskov, F.; Maase, H.v.d. Impact of Immune Parameters on Long-Term Survival in Metastatic Renal Cell Carcinoma. J. Clin. Oncol. 2006, 24, 1997–2005. [Google Scholar] [CrossRef] [PubMed]
- Atkins, M.B.; Tannir, N.M. Current and emerging therapies for first-line treatment of metastatic clear cell renal cell carcinoma. Cancer Treat. Rev. 2018, 70, 127–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altmayr, F.; Jusek, G.; Holzmann, B. The neuropeptide calcitonin gene-related peptide causes repression of tumor necrosis factor-alpha transcription and suppression of ATF-2 promoter recruitment in Toll-like receptor-stimulated dendritic cells. J. Biol. Chem. 2010, 285, 3525–3531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Q.; Xu, L.; Wang, Y.; Jiang, Q.; Liu, Z.; Zhang, J.; Zhou, Q.; Zeng, H.; Tong, S.; Wang, T.; et al. Tumor-associated Macrophage-derived Interleukin-23 Interlinks Kidney Cancer Glutamine Addiction with Immune Evasion. Eur. Urol. 2019, 75, 752–763. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.E.; Lee, H.G.; Cho, I.H.; Chung, D.H.; Yoon, S.H.; Yang, Y.M.; Lee, J.W.; Choi, S.; Park, J.W.; Ye, S.K.; et al. STAT3 is a potential modulator of HIF-1-mediated VEGF expression in human renal carcinoma cells. FASEB J. 2005, 19, 1296–1298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, N.; Baby, D.; Rajguru, J.P.; Patil, P.B.; Thakkannavar, S.S.; Pujari, V.B. Inflammation and cancer. Ann. Afr. Med. 2019, 18, 121–126. [Google Scholar] [CrossRef]
- Cancer Genome Atlas Research Network. Comprehensive molecular characterization of clear cell renal cell carcinoma. Nature 2013, 499, 43–49. [Google Scholar] [CrossRef] [Green Version]
- Maxwell, P.H.; Wiesener, M.S.; Chang, G.W.; Clifford, S.C.; Vaux, E.C.; Cockman, M.E.; Wykoff, C.C.; Pugh, C.W.; Maher, E.R.; Ratcliffe, P.J. The tumour suppressor protein VHL targets hypoxia-inducible factors for oxygen-dependent proteolysis. Nature 1999, 399, 271–275. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.; Kaelin, W.G., Jr. The VHL/HIF axis in clear cell renal carcinoma. Semin. Cancer Biol. 2013, 23, 18–25. [Google Scholar] [CrossRef] [Green Version]
- Iliopoulos, O.; Levy, A.P.; Jiang, C.; Kaelin, W.G., Jr.; Goldberg, M.A. Negative regulation of hypoxia-inducible genes by the von Hippel-Lindau protein. Proc. Natl. Acad. Sci. USA 1996, 93, 10595–10599. [Google Scholar] [CrossRef] [Green Version]
- Labrousse-Arias, D.; Martinez-Alonso, E.; Corral-Escariz, M.; Bienes-Martinez, R.; Berridy, J.; Serrano-Oviedo, L.; Conde, E.; Garcia-Bermejo, M.L.; Gimenez-Bachs, J.M.; Salinas-Sanchez, A.S.; et al. VHL promotes immune response against renal cell carcinoma via NF-kappaB-dependent regulation of VCAM-1. J. Cell Biol. 2017, 216, 835–847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staller, P.; Sulitkova, J.; Lisztwan, J.; Moch, H.; Oakeley, E.J.; Krek, W. Chemokine receptor CXCR4 downregulated by von Hippel-Lindau tumour suppressor pVHL. Nature 2003, 425, 307–311. [Google Scholar] [CrossRef] [PubMed]
- Galban, S.; Fan, J.; Martindale, J.L.; Cheadle, C.; Hoffman, B.; Woods, M.P.; Temeles, G.; Brieger, J.; Decker, J.; Gorospe, M. von Hippel-Lindau protein-mediated repression of tumor necrosis factor alpha translation revealed through use of cDNA arrays. Mol. Cell. Biol. 2003, 23, 2316–2328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazumder, S.; Higgins, P.J.; Samarakoon, R. Downstream Targets of VHL/HIF-alpha Signaling in Renal Clear Cell Carcinoma Progression: Mechanisms and Therapeutic Relevance. Cancers 2023, 15, 1316. [Google Scholar] [CrossRef]
- Monzon, F.A.; Alvarez, K.; Peterson, L.; Truong, L.; Amato, R.J.; Hernandez-McClain, J.; Tannir, N.; Parwani, A.V.; Jonasch, E. Chromosome 14q loss defines a molecular subtype of clear-cell renal cell carcinoma associated with poor prognosis. Mod. Pathol. 2011, 24, 1470–1479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordan, J.D.; Lal, P.; Dondeti, V.R.; Letrero, R.; Parekh, K.N.; Oquendo, C.E.; Greenberg, R.A.; Flaherty, K.T.; Rathmell, W.K.; Keith, B.; et al. HIF-alpha effects on c-Myc distinguish two subtypes of sporadic VHL-deficient clear cell renal carcinoma. Cancer Cell 2008, 14, 435–446. [Google Scholar] [CrossRef] [Green Version]
- Shen, C.; Beroukhim, R.; Schumacher, S.E.; Zhou, J.; Chang, M.; Signoretti, S.; Kaelin, W.G., Jr. Genetic and functional studies implicate HIF1alpha as a 14q kidney cancer suppressor gene. Cancer Discov. 2011, 1, 222–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kondo, K.; Kim, W.Y.; Lechpammer, M.; Kaelin, W.G., Jr. Inhibition of HIF2alpha is sufficient to suppress pVHL-defective tumor growth. PLoS Biol. 2003, 1, e83. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Hill, H.; Christie, A.; Kim, M.S.; Holloman, E.; Pavia-Jimenez, A.; Homayoun, F.; Ma, Y.; Patel, N.; Yell, P.; et al. Targeting renal cell carcinoma with a HIF-2 antagonist. Nature 2016, 539, 112–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuvillier, O. The therapeutic potential of HIF-2 antagonism in renal cell carcinoma. Transl. Androl. Urol. 2017, 6, 131–133. [Google Scholar] [CrossRef] [Green Version]
- Hoefflin, R.; Harlander, S.; Schafer, S.; Metzger, P.; Kuo, F.; Schonenberger, D.; Adlesic, M.; Peighambari, A.; Seidel, P.; Chen, C.Y.; et al. HIF-1alpha and HIF-2alpha differently regulate tumour development and inflammation of clear cell renal cell carcinoma in mice. Nat. Commun. 2020, 11, 4111. [Google Scholar] [CrossRef] [PubMed]
- Pritchett, T.L.; Bader, H.L.; Henderson, J.; Hsu, T. Conditional inactivation of the mouse von Hippel-Lindau tumor suppressor gene results in wide-spread hyperplastic, inflammatory and fibrotic lesions in the kidney. Oncogene 2015, 34, 2631–2639. [Google Scholar] [CrossRef] [PubMed]
- Kuo, C.Y.; Lin, C.H.; Hsu, T. VHL Inactivation in Precancerous Kidney Cells Induces an Inflammatory Response via ER Stress-Activated IRE1α Signaling. Cancer Res. 2017, 77, 3406–3416. [Google Scholar] [CrossRef] [Green Version]
- Shan, B.; Wang, X.; Wu, Y.; Xu, C.; Xia, Z.; Dai, J.; Shao, M.; Zhao, F.; He, S.; Yang, L.; et al. The metabolic ER stress sensor IRE1α suppresses alternative activation of macrophages and impairs energy expenditure in obesity. Nat. Immunol. 2017, 18, 519–529. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.N.; Nguyen-Tran, H.H.; Chen, C.Y.; Hsu, T. IL6 and CCL18 Mediate Cross-talk between VHL-Deficient Kidney Cells and Macrophages during Development of Renal Cell Carcinoma. Cancer Res. 2022, 82, 2716–2733. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Siemann, D.W. Gas6/Axl Signaling Pathway in the Tumor Immune Microenvironment. Cancers (Basel) 2020, 12, 1850. [Google Scholar] [CrossRef]
- Xiao, Y.; Zhao, H.; Tian, L.; Nolley, R.; Diep, A.N.; Ernst, A.; Fuh, K.C.; Miao, Y.R.; von Eyben, R.; Leppert, J.T.; et al. S100A10 Is a Critical Mediator of GAS6/AXL-Induced Angiogenesis in Renal Cell Carcinoma. Cancer Res. 2019, 79, 5758–5768. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.C.; Haworth, L.; Sherry, R.M.; Hwu, P.; Schwartzentruber, D.J.; Topalian, S.L.; Steinberg, S.M.; Chen, H.X.; Rosenberg, S.A. A randomized trial of bevacizumab, an anti-vascular endothelial growth factor antibody, for metastatic renal cancer. N. Engl. J. Med. 2003, 349, 427–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisen, T.; Oudard, S.; Szczylik, C.; Gravis, G.; Heinzer, H.; Middleton, R.; Cihon, F.; Anderson, S.; Shah, S.; Bukowski, R.; et al. Sorafenib for older patients with renal cell carcinoma: Subset analysis from a randomized trial. J Natl Cancer Inst 2008, 100, 1454–1463. [Google Scholar] [CrossRef]
- Makhov, P.; Joshi, S.S.; Ghatalia, P.; Kutikov, A.; Uzzo, R.G.; Kolenko, V. Resistance to Systemic Therapies in Clear Cell Renal Cell Carcinoma: Mechanisms and Management Strategies. Mol. Cancer Ther. 2018, 17, 1355–1364. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Liu, X.D.; Sun, M.; Zhang, X.; German, P.; Bai, S.; Ding, Z.; Tannir, N.; Wood, C.G.; Matin, S.F.; et al. Targeting MET and AXL overcomes resistance to sunitinib therapy in renal cell carcinoma. Oncogene 2016, 35, 2687–2697. [Google Scholar] [CrossRef] [PubMed]
- Yakes, F.M.; Chen, J.; Tan, J.; Yamaguchi, K.; Shi, Y.; Yu, P.; Qian, F.; Chu, F.; Bentzien, F.; Cancilla, B.; et al. Cabozantinib (XL184), a novel MET and VEGFR2 inhibitor, simultaneously suppresses metastasis, angiogenesis, and tumor growth. Mol. Cancer Ther. 2011, 10, 2298–2308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Motzer, R.; Alekseev, B.; Rha, S.-Y.; Porta, C.; Eto, M.; Powles, T.; Grünwald, V.; Hutson, T.E.; Kopyltsov, E.; Méndez-Vidal, M.J.; et al. Lenvatinib plus Pembrolizumab or Everolimus for Advanced Renal Cell Carcinoma. N. Engl. J. Med. 2021, 384, 1289–1300. [Google Scholar] [CrossRef]
- Al-Salama, Z.T.; Syed, Y.Y.; Scott, L.J. Lenvatinib: A Review in Hepatocellular Carcinoma. Drugs 2019, 79, 665–674. [Google Scholar] [CrossRef] [PubMed]
- Iacovelli, R.; Arduini, D.; Ciccarese, C.; Pierconti, F.; Strusi, A.; Piro, G.; Carbone, C.; Foschi, N.; Daniele, G.; Tortora, G. Targeting hypoxia-inducible factor pathways in sporadic and Von Hippel-Lindau syndrome-related kidney cancers. Crit. Rev. Oncol. Hematol. 2022, 176, 103750. [Google Scholar] [CrossRef] [PubMed]
- Deeks, E.D. Belzutifan: First Approval. Drugs 2021, 81, 1921–1927. [Google Scholar] [CrossRef]
- Jonasch, E.; Donskov, F.; Iliopoulos, O.; Rathmell, W.K.; Narayan, V.K.; Maughan, B.L.; Oudard, S.; Else, T.; Maranchie, J.K.; Welsh, S.J.; et al. Belzutifan for Renal Cell Carcinoma in von Hippel-Lindau Disease. N. Engl. J. Med. 2021, 385, 2036–2046. [Google Scholar] [CrossRef] [PubMed]
- Fallah, J.; Brave, M.H.; Weinstock, C.; Mehta, G.U.; Bradford, D.; Gittleman, H.; Bloomquist, E.W.; Charlab, R.; Hamed, S.S.; Miller, C.P.; et al. FDA Approval Summary: Belzutifan for von Hippel-Lindau Disease-Associated Tumors. Clin. Cancer Res. 2022, 28, 4843–4848. [Google Scholar] [CrossRef]
- Patel, S.A.; Hirosue, S.; Rodrigues, P.; Vojtasova, E.; Richardson, E.K.; Ge, J.; Syafruddin, S.E.; Speed, A.; Papachristou, E.K.; Baker, D.; et al. The renal lineage factor PAX8 controls oncogenic signalling in kidney cancer. Nature 2022, 606, 999–1006. [Google Scholar] [CrossRef]
- Bleu, M.; Gaulis, S.; Lopes, R.; Sprouffske, K.; Apfel, V.; Holwerda, S.; Pregnolato, M.; Yildiz, U.; Cordoʹ, V.; Dost, A.F.M.; et al. PAX8 activates metabolic genes via enhancer elements in Renal Cell Carcinoma. Nat. Commun. 2019, 10, 3739. [Google Scholar] [CrossRef] [Green Version]
- Sun, N.; Petiwala, S.; Lu, C.; Hutti, J.E.; Hu, M.; Hu, M.; Domanus, M.H.; Mitra, D.; Addo, S.N.; Miller, C.P.; et al. VHL Synthetic Lethality Signatures Uncovered by Genotype-Specific CRISPR-Cas9 Screens. CRISPR J. 2019, 2, 230–245. [Google Scholar] [CrossRef] [PubMed]
- Bakouny, Z.; Barbie, D.A. TBK1 Activation by VHL Loss in Renal Cell Carcinoma: A Novel HIF-Independent Vulnerability. Cancer Discov. 2020, 10, 348–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, L.; Xie, H.; Liu, X.; Potjewyd, F.; James, L.I.; Wilkerson, E.M.; Herring, L.E.; Xie, L.; Chen, X.; Cabrera, J.C.; et al. TBK1 Is a Synthetic Lethal Target in Cancer with VHL Loss. Cancer Discov. 2020, 10, 460–475. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.M.; Nguyen, Q.H.; Singh, M.; Pavesic, M.W.; Nesterenko, I.; Nelson, L.J.; Liao, A.C.; Razorenova, O.V. Rho-associated kinase 1 inhibition is synthetically lethal with von Hippel-Lindau deficiency in clear cell renal cell carcinoma. Oncogene 2017, 36, 1080–1089. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, A.A.; Nakamura, E.; Qi, J.; Creech, A.; Jaffe, J.D.; Paulk, J.; Novak, J.S.; Nagulapalli, K.; McBrayer, S.K.; Cowley, G.S.; et al. HIF activation causes synthetic lethality between the VHL tumor suppressor and the EZH1 histone methyltransferase. Sci. Transl. Med. 2017, 9, eaal5272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, D.A.; Sutphin, P.D.; Nguyen, P.; Turcotte, S.; Lai, E.W.; Banh, A.; Reynolds, G.E.; Chi, J.T.; Wu, J.; Solow-Cordero, D.E.; et al. Targeting GLUT1 and the Warburg effect in renal cell carcinoma by chemical synthetic lethality. Sci. Transl. Med. 2011, 3, 94ra70. [Google Scholar] [CrossRef] [Green Version]
- Nicholson, H.E.; Tariq, Z.; Housden, B.E.; Jennings, R.B.; Stransky, L.A.; Perrimon, N.; Signoretti, S.; Kaelin, W.G., Jr. HIF-independent synthetic lethality between CDK4/6 inhibition and VHL loss across species. Sci. Signal. 2019, 12, eaay0482. [Google Scholar] [CrossRef]
- Li, P.; Chen, T.; Kuang, P.; Liu, F.; Li, Z.; Liu, F.; Wang, Y.; Zhang, W.; Cai, X. Aurora-A/FOXO3A/SKP2 axis promotes tumor progression in clear cell renal cell carcinoma and dual-targeting Aurora-A/SKP2 shows synthetic lethality. Cell Death Dis. 2022, 13, 606. [Google Scholar] [CrossRef]
- Fu, M.; Hu, Y.; Lan, T.; Guan, K.L.; Luo, T.; Luo, M. The Hippo signalling pathway and its implications in human health and diseases. Signal Transduct. Target. Ther. 2022, 7, 376. [Google Scholar] [CrossRef]
- Xu, J.; Li, P.X.; Wu, J.; Gao, Y.J.; Yin, M.X.; Lin, Y.; Yang, M.; Chen, D.P.; Sun, H.P.; Liu, Z.B.; et al. Involvement of the Hippo pathway in regeneration and fibrogenesis after ischaemic acute kidney injury: YAP is the key effector. Clin. Sci. 2016, 130, 349–363. [Google Scholar] [CrossRef] [Green Version]
- Szeto, S.G.; Narimatsu, M.; Lu, M.; He, X.; Sidiqi, A.M.; Tolosa, M.F.; Chan, L.; De Freitas, K.; Bialik, J.F.; Majumder, S.; et al. YAP/TAZ Are Mechanoregulators of TGF-beta-Smad Signaling and Renal Fibrogenesis. J. Am. Soc. Nephrol. 2016, 27, 3117–3128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNeill, H.; Reginensi, A. Lats1/2 Regulate Yap/Taz to Control Nephron Progenitor Epithelialization and Inhibit Myofibroblast Formation. J. Am. Soc. Nephrol. 2017, 28, 852–861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.H.; Choi, H.I.; Park, J.S.; Kim, C.S.; Bae, E.H.; Ma, S.K.; Kim, S.W. Src-mediated crosstalk between FXR and YAP protects against renal fibrosis. FASEB J. 2019, 33, 11109–11122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Jin, D.; Zhang, Z.; Jin, D.; Xue, J.; Duan, L.; Zhang, Y.; Kang, X.; Lian, F. The critical role of the Hippo signaling pathway in kidney diseases. Front. Pharmacol. 2022, 13, 988175. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Liang, Y.; Zhu, X.; Wang, M.; Gui, Y.; Lu, Q.; Gu, M.; Xue, X.; Sun, X.; He, W.; et al. The signaling protein Wnt5a promotes TGFbeta1-mediated macrophage polarization and kidney fibrosis by inducing the transcriptional regulators Yap/Taz. J. Biol. Chem. 2018, 293, 19290–19302. [Google Scholar] [CrossRef] [Green Version]
- De Chiara, L.; Conte, C.; Semeraro, R.; Diaz-Bulnes, P.; Angelotti, M.L.; Mazzinghi, B.; Molli, A.; Antonelli, G.; Landini, S.; Melica, M.E.; et al. Tubular cell polyploidy protects from lethal acute kidney injury but promotes consequent chronic kidney disease. Nat. Commun. 2022, 13, 5805. [Google Scholar] [CrossRef]
- Chen, Y.B.; Xu, J.; Skanderup, A.J.; Dong, Y.; Brannon, A.R.; Wang, L.; Won, H.H.; Wang, P.I.; Nanjangud, G.J.; Jungbluth, A.A.; et al. Molecular analysis of aggressive renal cell carcinoma with unclassified histology reveals distinct subsets. Nat. Commun. 2016, 7, 13131. [Google Scholar] [CrossRef] [Green Version]
- Yu, F.X.; Zhao, B.; Guan, K.L. Hippo Pathway in Organ Size Control, Tissue Homeostasis, and Cancer. Cell 2015, 163, 811–828. [Google Scholar] [CrossRef] [Green Version]
- White, S.M.; Avantaggiati, M.L.; Nemazanyy, I.; Di Poto, C.; Yang, Y.; Pende, M.; Gibney, G.T.; Ressom, H.W.; Field, J.; Atkins, M.B.; et al. YAP/TAZ Inhibition Induces Metabolic and Signaling Rewiring Resulting in Targetable Vulnerabilities in NF2-Deficient Tumor Cells. Dev. Cell 2019, 49, 425–443 e429. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.H.; He, J.; Wang, D.L.; Cao, J.J.; Li, M.C.; Zhao, X.M.; Sheng, X.; Li, W.B.; Liu, W.J. Methylation-associated inactivation of LATS1 and its effect on demethylation or overexpression on YAP and cell biological function in human renal cell carcinoma. Int. J. Oncol. 2014, 45, 2511–2521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furth, N.; Aylon, Y. The LATS1 and LATS2 tumor suppressors: Beyond the Hippo pathway. Cell Death Differ. 2017, 24, 1488–1501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, L.; Li, W.; Wang, G.; Shi, H.; Wang, K.; Yang, H.; Peng, B. NR1B2 suppress kidney renal clear cell carcinoma (KIRC) progression by regulation of LATS 1/2-YAP signaling. J. Exp. Clin. Cancer Res. 2019, 38, 343. [Google Scholar] [CrossRef] [Green Version]
- Ruan, H.; Bao, L.; Song, Z.; Wang, K.; Cao, Q.; Tong, J.; Cheng, G.; Xu, T.; Chen, X.; Liu, D.; et al. High expression of TAZ serves as a novel prognostic biomarker and drives cancer progression in renal cancer. Exp. Cell Res. 2019, 376, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Heng, C.; Zhou, Y.; Hu, Y.; Chen, S.; Wang, H.; Yang, H.; Jiang, Z.; Qian, S.; Wang, Y.; et al. Targeting mammalian serine/threonine-protein kinase 4 through Yes-associated protein/TEA domain transcription factor-mediated epithelial-mesenchymal transition ameliorates diabetic nephropathy orchestrated renal fibrosis. Metabolism 2020, 108, 154258. [Google Scholar] [CrossRef] [PubMed]
- Kumar, B.; Ahmad, R.; Giannico, G.A.; Zent, R.; Talmon, G.A.; Harris, R.C.; Clark, P.E.; Lokeshwar, V.; Dhawan, P.; Singh, A.B. Claudin-2 inhibits renal clear cell carcinoma progression by inhibiting YAP-activation. J. Exp. Clin. Cancer Res. 2021, 40, 77. [Google Scholar] [CrossRef] [PubMed]
- Harten, S.K.; Shukla, D.; Barod, R.; Hergovich, A.; Balda, M.S.; Matter, K.; Esteban, M.A.; Maxwell, P.H. Regulation of renal epithelial tight junctions by the von Hippel-Lindau tumor suppressor gene involves occludin and claudin 1 and is independent of E-cadherin. Mol. Biol. Cell 2009, 20, 1089–1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tapon, N.; Harvey, K.F.; Bell, D.W.; Wahrer, D.C.; Schiripo, T.A.; Haber, D.; Hariharan, I.K. salvador Promotes both cell cycle exit and apoptosis in Drosophila and is mutated in human cancer cell lines. Cell 2002, 110, 467–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuura, K.; Nakada, C.; Mashio, M.; Narimatsu, T.; Yoshimoto, T.; Tanigawa, M.; Tsukamoto, Y.; Hijiya, N.; Takeuchi, I.; Nomura, T.; et al. Downregulation of SAV1 plays a role in pathogenesis of high-grade clear cell renal cell carcinoma. BMC Cancer 2011, 11, 523. [Google Scholar] [CrossRef]
- Mehra, R.; Vats, P.; Cieslik, M.; Cao, X.; Su, F.; Shukla, S.; Udager, A.M.; Wang, R.; Pan, J.; Kasaian, K.; et al. Biallelic Alteration and Dysregulation of the Hippo Pathway in Mucinous Tubular and Spindle Cell Carcinoma of the Kidney. Cancer Discov. 2016, 6, 1258–1266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, C.; Lv, S.; Wang, H.; Liu, C.; Zhai, Q.; Wang, S.; Cai, G.; Lu, D.; Sun, Z.; Wei, Q. Leukemia Inhibitory Factor Receptor Suppresses the Metastasis of Clear Cell Renal Cell Carcinoma Through Negative Regulation of the Yes-Associated Protein. DNA Cell Biol. 2018, 37, 659–669. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Li, W.; Xu, A.; Shi, H.; Wang, K.; Yang, H.; Wang, R.; Peng, B. SH3BGRL2 inhibits growth and metastasis in clear cell renal cell carcinoma via activating hippo/TEAD1-Twist1 pathway. EBioMedicine 2020, 51, 102596. [Google Scholar] [CrossRef] [Green Version]
- Song, S.; Xie, M.; Scott, A.W.; Jin, J.; Ma, L.; Dong, X.; Skinner, H.D.; Johnson, R.L.; Ding, S.; Ajani, J.A. A Novel YAP1 Inhibitor Targets CSC-Enriched Radiation-Resistant Cells and Exerts Strong Antitumor Activity in Esophageal Adenocarcinoma. Mol. Cancer Ther. 2018, 17, 443–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taniguchi, K.; Karin, M. NF-κB, inflammation, immunity and cancer: Coming of age. Nat. Rev. Immunol. 2018, 18, 309–324. [Google Scholar] [CrossRef]
- Karin, M. NF-kappaB as a critical link between inflammation and cancer. Cold Spring Harb. Perspect. Biol. 2009, 1, a000141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.; Bai, L.; Chen, W.; Xu, S. The NF-kappaB activation pathways, emerging molecular targets for cancer prevention and therapy. Expert. Opin. Ther. Targets 2010, 14, 45–55. [Google Scholar] [CrossRef] [Green Version]
- Saccani, A.; Schioppa, T.; Porta, C.; Biswas, S.K.; Nebuloni, M.; Vago, L.; Bottazzi, B.; Colombo, M.P.; Mantovani, A.; Sica, A. p50 nuclear factor-kappaB overexpression in tumor-associated macrophages inhibits M1 inflammatory responses and antitumor resistance. Cancer Res. 2006, 66, 11432–11440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, H.; Ohh, M. The von Hippel-Lindau tumor suppressor protein sensitizes renal cell carcinoma cells to tumor necrosis factor-induced cytotoxicity by suppressing the nuclear factor-kappaB-dependent antiapoptotic pathway. Cancer Res. 2003, 63, 7076–7080. [Google Scholar] [PubMed]
- Cory, S.; Adams, J.M. The Bcl2 family: Regulators of the cellular life-or-death switch. Nat. Rev. Cancer 2002, 2, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Ng, K.L.; Yap, N.Y.; Rajandram, R.; Small, D.; Pailoor, J.; Ong, T.A.; Razack, A.H.; Wood, S.T.; Morais, C.; Gobe, G.C. Nuclear factor-kappa B subunits and their prognostic cancer-specific survival value in renal cell carcinoma patients. Pathology 2018, 50, 511–518. [Google Scholar] [CrossRef]
- Rebe, C.; Ghiringhelli, F. STAT3, a Master Regulator of Anti-Tumor Immune Response. Cancers 2019, 11, 1280. [Google Scholar] [CrossRef] [Green Version]
- Yin, H.; Zhang, X.; Yang, P.; Zhang, X.; Peng, Y.; Li, D.; Yu, Y.; Wu, Y.; Wang, Y.; Zhang, J.; et al. RNA m6A methylation orchestrates cancer growth and metastasis via macrophage reprogramming. Nat. Commun. 2021, 12, 1394. [Google Scholar] [CrossRef] [PubMed]
- Evaristo, C.; Spranger, S.; Barnes, S.E.; Miller, M.L.; Molinero, L.L.; Locke, F.L.; Gajewski, T.F.; Alegre, M.-L. Cutting Edge: Engineering Active IKKβ in T Cells Drives Tumor Rejection. J. Immunol. 2016, 196, 2933–2938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Nag, S.A.; Zhang, R. Targeting the NFκB signaling pathways for breast cancer prevention and therapy. Curr. Med. Chem. 2015, 22, 264–289. [Google Scholar] [CrossRef] [PubMed]
- Kondagunta, G.V.; Drucker, B.; Schwartz, L.; Bacik, J.; Marion, S.; Russo, P.; Mazumdar, M.; Motzer, R.J. Phase II trial of bortezomib for patients with advanced renal cell carcinoma. J. Clin. Oncol. 2004, 22, 3720–3725. [Google Scholar] [CrossRef] [PubMed]
- Brooks, A.D.; Jacobsen, K.M.; Li, W.; Shanker, A.; Sayers, T.J. Bortezomib Sensitizes Human Renal Cell Carcinomas to TRAIL Apoptosis through Increased Activation of Caspase-8 in the Death-Inducing Signaling Complex. Mol. Cancer Res. 2010, 8, 729–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erstad, D.J.; Cusack, J.C., Jr. Targeting the NF-κB pathway in cancer therapy. Surg. Oncol. Clin. N. Am. 2013, 22, 705–746. [Google Scholar] [CrossRef] [PubMed]
- Gilmore, T.D.; Garbati, M.R. Inhibition of NF-κB signaling as a strategy in disease therapy. Curr. Top. Microbiol. Immunol. 2011, 349, 245–263. [Google Scholar] [CrossRef]
- Hopfner, K.-P.; Hornung, V. Molecular mechanisms and cellular functions of cGAS–STING signalling. Nat. Rev. Mol. Cell Biol. 2020, 21, 501–521. [Google Scholar] [CrossRef]
- Kwon, J.; Bakhoum, S.F. The Cytosolic DNA-Sensing cGAS-STING Pathway in Cancer. Cancer Discov. 2020, 10, 26–39. [Google Scholar] [CrossRef]
- Khoo, L.T.; Chen, L.Y. Role of the cGAS-STING pathway in cancer development and oncotherapeutic approaches. EMBO Rep. 2018, 19, e46935. [Google Scholar] [CrossRef]
- Amouzegar, A.; Chelvanambi, M.; Filderman, J.N.; Storkus, W.J.; Luke, J.J. STING Agonists as Cancer Therapeutics. Cancers 2021, 13, 2695. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Lin, Y.; Liu, L.-M.; Hou, Y.-L.; Qin, W.-T.; Zhang, L.; Jiang, S.-H.; Yang, Q.; Bai, Y.-R. Identification of Cytosolic DNA Sensor cGAS-STING as Immune-Related Risk Factor in Renal Carcinoma following Pan-Cancer Analysis. J. Immunol. Res. 2022, 2022, 7978042. [Google Scholar] [CrossRef] [PubMed]
- Mitrofanova, A.; Fontanella, A.; Tolerico, M.; Mallela, S.; Molina David, J.; Zuo, Y.; Boulina, M.; Kim, J.-J.; Santos, J.; Ge, M.; et al. Activation of Stimulator of IFN Genes (STING) Causes Proteinuria and Contributes to Glomerular Diseases. J. Am. Soc. Nephrol. 2022, 33, 2153–2173. [Google Scholar] [CrossRef] [PubMed]
- Gong, W.; Lu, L.; Zhou, Y.; Liu, J.; Ma, H.; Fu, L.; Huang, S.; Zhang, Y.; Zhang, A.; Jia, Z. The novel STING antagonist H151 ameliorates cisplatin-induced acute kidney injury and mitochondrial dysfunction. Am. J. Physiol. -Ren. Physiol. 2021, 320, F608–F616. [Google Scholar] [CrossRef]
- Larkin, B.; Ilyukha, V.; Sorokin, M.; Buzdin, A.; Vannier, E.; Poltorak, A. Cutting edge: Activation of STING in T cells induces type I IFN responses and cell death. J. Immunol. 2017, 199, 397–402. [Google Scholar] [CrossRef] [Green Version]
- Alfert, A.; Moreno, N.; Kerl, K. The BAF complex in development and disease. Epigenetics Chromatin 2019, 12, 19. [Google Scholar] [CrossRef] [Green Version]
- Cooper, G.W.; Hong, A.L. SMARCB1-Deficient Cancers: Novel Molecular Insights and Therapeutic Vulnerabilities. Cancers 2022, 14, 3645. [Google Scholar] [CrossRef]
- Msaouel, P.; Malouf, G.G.; Su, X.; Yao, H.; Tripathi, D.N.; Soeung, M.; Gao, J.; Rao, P.; Coarfa, C.; Creighton, C.J.; et al. Comprehensive Molecular Characterization Identifies Distinct Genomic and Immune Hallmarks of Renal Medullary Carcinoma. Cancer Cell 2020, 37, 720–734.e713. [Google Scholar] [CrossRef]
- Gao, W.; Li, W.; Xiao, T.; Liu, X.S.; Kaelin, W.G. Inactivation of the PBRM1 tumor suppressor gene amplifies the HIF-response in VHL−/− clear cell renal carcinoma. Proc. Natl. Acad. Sci. USA 2017, 114, 1027–1032. [Google Scholar] [CrossRef] [Green Version]
- Varela, I.; Tarpey, P.; Raine, K.; Huang, D.; Ong, C.K.; Stephens, P.; Davies, H.; Jones, D.; Lin, M.L.; Teague, J.; et al. Exome sequencing identifies frequent mutation of the SWI/SNF complex gene PBRM1 in renal carcinoma. Nature 2011, 469, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Burrows, A.E.; Smogorzewska, A.; Elledge, S.J. Polybromo-associated BRG1-associated factor components BRD7 and BAF180 are critical regulators of p53 required for induction of replicative senescence. Proc. Natl. Acad. Sci. USA 2010, 107, 14280–14285. [Google Scholar] [CrossRef] [Green Version]
- Chabanon, R.M.; Morel, D.; Eychenne, T.; Colmet-Daage, L.; Bajrami, I.; Dorvault, N.; Garrido, M.; Meisenberg, C.; Lamb, A.; Ngo, C.; et al. PBRM1 Deficiency Confers Synthetic Lethality to DNA Repair Inhibitors in Cancer. Cancer Res. 2021, 81, 2888–2902. [Google Scholar] [CrossRef] [PubMed]
- Schepisi, G.; Conteduca, V.; Casadei, C.; Gurioli, G.; Rossi, L.; Galla, V.; Cursano, M.C.; Brighi, N.; Lolli, C.; Menna, C.; et al. Potential Application of Chimeric Antigen Receptor (CAR)-T Cell Therapy in Renal Cell Tumors. Front. Oncol. 2020, 10, 565857. [Google Scholar] [CrossRef]
- Adam, P.J.; Terrett, J.A.; Steers, G.; Stockwin, L.; Loader, J.A.; Fletcher, G.C.; Lu, L.S.; Leach, B.I.; Mason, S.; Stamps, A.C.; et al. CD70 (TNFSF7) is expressed at high prevalence in renal cell carcinomas and is rapidly internalised on antibody binding. Br. J. Cancer 2006, 95, 298–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jilaveanu, L.B.; Sznol, J.; Aziz, S.A.; Duchen, D.; Kluger, H.M.; Camp, R.L. CD70 expression patterns in renal cell carcinoma. Hum. Pathol. 2012, 43, 1394–1399. [Google Scholar] [CrossRef] [Green Version]
- Pal, S.K.; Forero-Torres, A.; Thompson, J.A.; Morris, J.C.; Chhabra, S.; Hoimes, C.J.; Vogelzang, N.J.; Boyd, T.; Bergerot, P.G.; Adashek, J.J.; et al. A phase 1 trial of SGN-CD70A in patients with CD70-positive, metastatic renal cell carcinoma. Cancer 2019, 125, 1124–1132. [Google Scholar] [CrossRef] [PubMed]
- Panowski, S.H.; Srinivasan, S.; Tan, N.; Tacheva-Grigorova, S.K.; Smith, B.; Mak, Y.S.L.; Ning, H.; Villanueva, J.; Wijewarnasuriya, D.; Lang, S.; et al. Preclinical Development and Evaluation of Allogeneic CAR T Cells Targeting CD70 for the Treatment of Renal Cell Carcinoma. Cancer Res. 2022, 82, 2610–2624. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Kelly, T.K.; Jones, P.A. Epigenetics in cancer. Carcinogenesis 2009, 31, 27–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joosten, S.C.; Smits, K.M.; Aarts, M.J.; Melotte, V.; Koch, A.; Tjan-Heijnen, V.C.; van Engeland, M. Epigenetics in renal cell cancer: Mechanisms and clinical applications. Nat Rev Urol 2018, 15, 430–451. [Google Scholar] [CrossRef] [PubMed]
- Maiuri, A.R.; O’Hagan, H.M. Interplay Between Inflammation and Epigenetic Changes in Cancer. Prog. Mol. Biol. Transl. Sci. 2016, 144, 69–117. [Google Scholar] [CrossRef]
- Karin, M.; Shalapour, S. Regulation of antitumor immunity by inflammation-induced epigenetic alterations. Cell. Mol. Immunol. 2022, 19, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Joosten, S.C.; Deckers, I.A.; Aarts, M.J.; Hoeben, A.; van Roermund, J.G.; Smits, K.M.; Melotte, V.; van Engeland, M.; Tjan-Heijnen, V.C. Prognostic DNA methylation markers for renal cell carcinoma: A systematic review. Epigenomics 2017, 9, 1243–1257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, M.R.; Latif, F. The epigenetic landscape of renal cancer. Nat. Rev. Nephrol. 2017, 13, 47–60. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.-H.; Haddad, A.; Wu, K.-J.; Zhao, H.-W.; Kapur, P.; Zhang, Z.-L.; Zhao, L.-Y.; Chen, Z.-H.; Zhou, Y.-Y.; Zhou, J.-C.; et al. A CpG-methylation-based assay to predict survival in clear cell renal cell carcinoma. Nat. Commun. 2015, 6, 8699. [Google Scholar] [CrossRef] [Green Version]
- Ramankulov, A.; Lein, M.; Johannsen, M.; Schrader, M.; Miller, K.; Loening, S.A.; Jung, K. Serum amyloid A as indicator of distant metastases but not as early tumor marker in patients with renal cell carcinoma. Cancer Lett. 2008, 269, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M.; Tomita, Y.; Imai, T.; Saito, T.; Katagiri, A.; Ohara-Mikami, Y.; Matsudo, T.; Takahashi, K. Significance of serum amyloid A on the prognosis in patients with renal cell carcinoma. Cancer 2001, 92, 2072–2075. [Google Scholar] [CrossRef] [PubMed]
- Nishida, J.; Momoi, Y.; Miyakuni, K.; Tamura, Y.; Takahashi, K.; Koinuma, D.; Miyazono, K.; Ehata, S. Epigenetic remodelling shapes inflammatory renal cancer and neutrophil-dependent metastasis. Nat. Cell Biol. 2020, 22, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, P.; Patel, S.A.; Harewood, L.; Olan, I.; Vojtasova, E.; Syafruddin, S.E.; Zaini, M.N.; Richardson, E.K.; Burge, J.; Warren, A.Y.; et al. NF-κB–Dependent Lymphoid Enhancer Co-option Promotes Renal Carcinoma Metastasis. Cancer Discov. 2018, 8, 850–865. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.H.; Kim, B.C.; Jeong, S.H.; Jeong, C.W.; Ku, J.H.; Kwak, C.; Kim, H.H. Histone Demethylase LSD1 Regulates Kidney Cancer Progression by Modulating Androgen Receptor Activity. Int. J. Mol. Sci. 2020, 21, 6089. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Wang, J.; Kong, W.; Huang, J.; Dong, B.; Huang, Y.; Xue, W.; Zhang, J. LSD1 inhibition suppresses the growth of clear cell renal cell carcinoma via upregulating P21 signaling. Acta Pharm. Sin. B 2019, 9, 324–334. [Google Scholar] [CrossRef]
- Ma, Y.; Zheng, Y.; Ji, Y.; Wang, X.; Ye, B. Raloxifene, identified as a novel LSD1 inhibitor, suppresses the migration of renal cell carcinoma. Future Med. Chem. 2021, 13, 533–542. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.C.; Ma, Y.C.; Qin, W.P.; Ding, L.N.; Zheng, Y.C.; Zhu, Y.L.; Zhai, X.Y.; Yang, J.; Ma, C.Y.; Guan, Y.Y. Design and synthesis of tranylcypromine derivatives as novel LSD1/HDACs dual inhibitors for cancer treatment. Eur. J. Med. Chem. 2017, 140, 392–402. [Google Scholar] [CrossRef] [PubMed]
- Maes, T.; Mascaró, C.; Tirapu, I.; Estiarte, A.; Ciceri, F.; Lunardi, S.; Guibourt, N.; Perdones, A.; Lufino, M.M.P.; Somervaille, T.C.P.; et al. ORY-1001, a Potent and Selective Covalent KDM1A Inhibitor, for the Treatment of Acute Leukemia. Cancer Cell 2018, 33, 495–511.e412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, X.; Tan, J.; Lim, K.J.; Koh, J.; Ooi, W.F.; Li, Z.; Huang, D.; Xing, M.; Chan, Y.S.; Qu, J.Z.; et al. VHL Deficiency Drives Enhancer Activation of Oncogenes in Clear Cell Renal Cell Carcinoma. Cancer Discov. 2017, 7, 1284–1305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hutt, D.M.; Roth, D.M.; Vignaud, H.; Cullin, C.; Bouchecareilh, M. The Histone Deacetylase Inhibitor, Vorinostat, Represses Hypoxia Inducible Factor 1 Alpha Expression through Translational Inhibition. PLoS ONE 2014, 9, e106224. [Google Scholar] [CrossRef] [Green Version]
- Blankenstein, T.; Coulie, P.G.; Gilboa, E.; Jaffee, E.M. The determinants of tumour immunogenicity. Nat. Rev. Cancer 2012, 12, 307–313. [Google Scholar] [CrossRef]
- Amin, A.; Hammers, H. The Evolving Landscape of Immunotherapy-Based Combinations for Frontline Treatment of Advanced Renal Cell Carcinoma. Front. Immunol. 2018, 9, 3120. [Google Scholar] [CrossRef] [PubMed]
- Raghubar, A.M.; Roberts, M.J.; Wood, S.; Healy, H.G.; Kassianos, A.J.; Mallett, A.J. Cellular milieu in clear cell renal cell carcinoma. Front. Oncol. 2022, 12, 943583. [Google Scholar] [CrossRef]
- Nakano, O.; Sato, M.; Naito, Y.; Suzuki, K.; Orikasa, S.; Aizawa, M.; Suzuki, Y.; Shintaku, I.; Nagura, H.; Ohtani, H. Proliferative activity of intratumoral CD8(+) T-lymphocytes as a prognostic factor in human renal cell carcinoma: Clinicopathologic demonstration of antitumor immunity. Cancer Res. 2001, 61, 5132–5136. [Google Scholar]
- Zhang, S.; Zhang, E.; Long, J.; Hu, Z.; Peng, J.; Liu, L.; Tang, F.; Li, L.; Ouyang, Y.; Zeng, Z. Immune infiltration in renal cell carcinoma. Cancer Sci. 2019, 110, 1564–1572. [Google Scholar] [CrossRef] [Green Version]
- Yao, J.; Xi, W.; Zhu, Y.; Wang, H.; Hu, X.; Guo, J. Checkpoint molecule PD-1-assisted CD8(+) T lymphocyte count in tumor microenvironment predicts overall survival of patients with metastatic renal cell carcinoma treated with tyrosine kinase inhibitors. Cancer Manag. Res. 2018, 10, 3419–3431. [Google Scholar] [CrossRef] [Green Version]
- Remark, R.; Alifano, M.; Cremer, I.; Lupo, A.; Dieu-Nosjean, M.C.; Riquet, M.; Crozet, L.; Ouakrim, H.; Goc, J.; Cazes, A.; et al. Characteristics and clinical impacts of the immune environments in colorectal and renal cell carcinoma lung metastases: Influence of tumor origin. Clin. Cancer Res. 2013, 19, 4079–4091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chevrier, S.; Levine, J.H.; Zanotelli, V.R.T.; Silina, K.; Schulz, D.; Bacac, M.; Ries, C.H.; Ailles, L.; Jewett, M.A.S.; Moch, H.; et al. An Immune Atlas of Clear Cell Renal Cell Carcinoma. Cell 2017, 169, 736–749 e718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varn, F.S.; Wang, Y.; Mullins, D.W.; Fiering, S.; Cheng, C. Systematic Pan-Cancer Analysis Reveals Immune Cell Interactions in the Tumor Microenvironment. Cancer Res. 2017, 77, 1271–1282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawashima, A.; Kanazawa, T.; Kidani, Y.; Yoshida, T.; Hirata, M.; Nishida, K.; Nojima, S.; Yamamoto, Y.; Kato, T.; Hatano, K.; et al. Tumour grade significantly correlates with total dysfunction of tumour tissue-infiltrating lymphocytes in renal cell carcinoma. Sci. Rep. 2020, 10, 6220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senbabaoglu, Y.; Gejman, R.S.; Winer, A.G.; Liu, M.; Van Allen, E.M.; de Velasco, G.; Miao, D.; Ostrovnaya, I.; Drill, E.; Luna, A.; et al. Tumor immune microenvironment characterization in clear cell renal cell carcinoma identifies prognostic and immunotherapeutically relevant messenger RNA signatures. Genome Biol. 2016, 17, 231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braun, D.A.; Bakouny, Z.; Hirsch, L.; Flippot, R.; Van Allen, E.M.; Wu, C.J.; Choueiri, T.K. Beyond conventional immune-checkpoint inhibition - novel immunotherapies for renal cell carcinoma. Nat Rev Clin Oncol 2021, 18, 199–214. [Google Scholar] [CrossRef]
- Giraldo, N.A.; Becht, E.; Pagès, F.; Skliris, G.; Verkarre, V.; Vano, Y.; Mejean, A.; Saint-Aubert, N.; Lacroix, L.; Natario, I.; et al. Orchestration and Prognostic Significance of Immune Checkpoints in the Microenvironment of Primary and Metastatic Renal Cell Cancer. Clin. Cancer Res. 2015, 21, 3031–3040. [Google Scholar] [CrossRef] [Green Version]
- Thompson, R.H.; Dong, H.; Lohse, C.M.; Leibovich, B.C.; Blute, M.L.; Cheville, J.C.; Kwon, E.D. PD-1 is expressed by tumor-infiltrating immune cells and is associated with poor outcome for patients with renal cell carcinoma. Clin. Cancer Res. 2007, 13, 1757–1761. [Google Scholar] [CrossRef] [Green Version]
- Beuselinck, B.; Job, S.; Becht, E.; Karadimou, A.; Verkarre, V.; Couchy, G.; Giraldo, N.; Rioux-Leclercq, N.; Molinie, V.; Sibony, M.; et al. Molecular subtypes of clear cell renal cell carcinoma are associated with sunitinib response in the metastatic setting. Clin. Cancer Res. 2015, 21, 1329–1339. [Google Scholar] [CrossRef] [Green Version]
- Giraldo, N.A.; Becht, E.; Vano, Y.; Petitprez, F.; Lacroix, L.; Validire, P.; Sanchez-Salas, R.; Ingels, A.; Oudard, S.; Moatti, A.; et al. Tumor-Infiltrating and Peripheral Blood T-cell Immunophenotypes Predict Early Relapse in Localized Clear Cell Renal Cell Carcinoma. Clin. Cancer Res. 2017, 23, 4416–4428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piedra-Quintero, Z.L.; Wilson, Z.; Nava, P.; Guerau-de-Arellano, M. CD38: An Immunomodulatory Molecule in Inflammation and Autoimmunity. Front. Immunol. 2020, 11, 597959. [Google Scholar] [CrossRef] [PubMed]
- Obradovic, A.; Chowdhury, N.; Haake, S.M.; Ager, C.; Wang, V.; Vlahos, L.; Guo, X.V.; Aggen, D.H.; Rathmell, W.K.; Jonasch, E.; et al. Single-cell protein activity analysis identifies recurrence-associated renal tumor macrophages. Cell 2021, 184, 2988–3005.e2916. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Xu, J.; Lan, H. Tumor-associated macrophages in tumor metastasis: Biological roles and clinical therapeutic applications. J. Hematol. Oncol. 2019, 12, 76. [Google Scholar] [CrossRef] [PubMed]
- Italiani, P.; Boraschi, D. From Monocytes to M1/M2 Macrophages: Phenotypical vs. Functional Differentiation. Front. Immunol. 2014, 5, 514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bi, K.; He, M.X.; Bakouny, Z.; Kanodia, A.; Napolitano, S.; Wu, J.; Grimaldi, G.; Braun, D.A.; Cuoco, M.S.; Mayorga, A.; et al. Tumor and immune reprogramming during immunotherapy in advanced renal cell carcinoma. Cancer Cell 2021, 39, 649–661 e645. [Google Scholar] [CrossRef] [PubMed]
- Braun, D.A.; Street, K.; Burke, K.P.; Cookmeyer, D.L.; Denize, T.; Pedersen, C.B.; Gohil, S.H.; Schindler, N.; Pomerance, L.; Hirsch, L.; et al. Progressive immune dysfunction with advancing disease stage in renal cell carcinoma. Cancer Cell 2021, 39, 632–648.e638. [Google Scholar] [CrossRef]
- Gocher, A.M.; Workman, C.J.; Vignali, D.A.A. Interferon-γ: Teammate or opponent in the tumour microenvironment? Nat. Rev. Immunol. 2022, 22, 158–172. [Google Scholar] [CrossRef]
- Mo, X.; Zhang, H.; Preston, S.; Martin, K.; Zhou, B.; Vadalia, N.; Gamero, A.M.; Soboloff, J.; Tempera, I.; Zaidi, M.R. Interferon-γ Signaling in Melanocytes and Melanoma Cells Regulates Expression of CTLA-4. Cancer Res. 2018, 78, 436–450. [Google Scholar] [CrossRef] [Green Version]
- Hänze, J.; Wegner, M.; Noessner, E.; Hofmann, R.; Hegele, A. Co-Regulation of Immune Checkpoint PD-L1 with Interferon-Gamma Signaling is Associated with a Survival Benefit in Renal Cell Cancer. Target. Oncol. 2020, 15, 377–390. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Tao, J.; Zhu, W.; Liu, W.; Anwaier, A.; Tian, X.; Su, J.; Shi, G.; Huang, H.; Wei, G.; et al. Comprehensive Multi-Omics Identification of Interferon-gamma Response Characteristics Reveals That RBCK1 Regulates the Immunosuppressive Microenvironment of Renal Cell Carcinoma. Front. Immunol. 2021, 12, 734646. [Google Scholar] [CrossRef] [PubMed]
- Balachandran, S.; Adams, G.P. Interferon-γ-induced necrosis: An antitumor biotherapeutic perspective. J. Interferon Cytokine Res. 2013, 33, 171–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, P.; Nogusa, S.; Thapa, R.J.; Shaller, C.; Simmons, H.; Peri, S.; Adams, G.P.; Balachandran, S. Anti-CD70 Immunocytokines for Exploitation of Interferon-γ-Induced RIP1-Dependent Necrosis in Renal Cell Carcinoma. PLoS ONE 2013, 8, e61446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsimafeyeu, I.; Demidov, L.; Stepanova, E.; Wynn, N.; Ta, H. Overexpression of fibroblast growth factor receptors FGFR1 and FGFR2 in renal cell carcinoma. Scand. J. Urol. Nephrol. 2011, 45, 190–195. [Google Scholar] [CrossRef] [PubMed]
- Babina, I.S.; Turner, N.C. Advances and challenges in targeting FGFR signalling in cancer. Nat. Rev. Cancer 2017, 17, 318–332. [Google Scholar] [CrossRef] [PubMed]
- Adachi, Y.; Kamiyama, H.; Ichikawa, K.; Fukushima, S.; Ozawa, Y.; Yamaguchi, S.; Goda, S.; Kimura, T.; Kodama, K.; Matsuki, M.; et al. Inhibition of FGFR Reactivates IFNγ Signaling in Tumor Cells to Enhance the Combined Antitumor Activity of Lenvatinib with Anti-PD-1 Antibodies. Cancer Res. 2022, 82, 292–306. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Matsui, J.; Matsushima, T.; Obaishi, H.; Miyazaki, K.; Nakamura, K.; Tohyama, O.; Semba, T.; Yamaguchi, A.; Hoshi, S.S.; et al. Lenvatinib, an angiogenesis inhibitor targeting VEGFR/FGFR, shows broad antitumor activity in human tumor xenograft models associated with microvessel density and pericyte coverage. Vasc. Cell 2014, 6, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González, A.; Varo, N.; Alegre, E.; Díaz, A.; Melero, I. Immunosuppression routed via the kynurenine pathway: A biochemical and pathophysiologic approach. Adv. Clin. Chem. 2008, 45, 155–197. [Google Scholar] [CrossRef] [PubMed]
- Trott, J.F.; Kim, J.; Aboud, O.A.; Wettersten, H.; Stewart, B.; Berryhill, G.; Uzal, F.; Hovey, R.C.; Chen, C.-H.; Anderson, K.; et al. Inhibiting tryptophan metabolism enhances interferon therapy in kidney cancer. Oncotarget 2016, 7, 66540–66557. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.; Taylor, S.L.; Ganti, S.; Guo, L.; Osier, M.V.; Weiss, R.H. Urine metabolomic analysis identifies potential biomarkers and pathogenic pathways in kidney cancer. Omics 2011, 15, 293–303. [Google Scholar] [CrossRef] [Green Version]
- Lo, U.G.; Bao, J.; Cen, J.; Yeh, H.C.; Luo, J.; Tan, W.; Hsieh, J.T. Interferon-induced IFIT5 promotes epithelial-to-mesenchymal transition leading to renal cancer invasion. Am. J. Clin. Exp. Urol. 2019, 7, 31–45. [Google Scholar] [PubMed]
- Au-Yeung, N.; Mandhana, R.; Horvath, C.M. Transcriptional regulation by STAT1 and STAT2 in the interferon JAK-STAT pathway. Jakstat 2013, 2, e23931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lo, U.G.; Pong, R.-C.; Yang, D.; Gandee, L.; Hernandez, E.; Dang, A.; Lin, C.-J.; Santoyo, J.; Ma, S.; Sonavane, R.; et al. IFNγ-Induced IFIT5 Promotes Epithelial-to-Mesenchymal Transition in Prostate Cancer via miRNA Processing. Cancer Res. 2019, 79, 1098–1112. [Google Scholar] [CrossRef] [PubMed]
- Charych, D.H.; Hoch, U.; Langowski, J.L.; Lee, S.R.; Addepalli, M.K.; Kirk, P.B.; Sheng, D.; Liu, X.; Sims, P.W.; VanderVeen, L.A.; et al. NKTR-214, an Engineered Cytokine with Biased IL2 Receptor Binding, Increased Tumor Exposure, and Marked Efficacy in Mouse Tumor Models. Clin. Cancer Res. 2016, 22, 680–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.C.; Pyo, K.H.; Song, Y.J.; Lee, W.J.; Kim, J.; Ji, H.N.; Park, S.S.; Koh, Y.J.; Lee, K.; Cho, B.; et al. GI101, a novel triple-targeting bispecific CD80-IgG4-IL2variant fusion protein, elicits synergistic anti-tumour effects in preclinical models. Ann. Oncol. 2019, 30, v500. [Google Scholar] [CrossRef]
- Pyo, K.-H.; Koh, Y.J.; Synn, C.-B.; Kim, J.H.; Byeon, Y.; Jo, H.N.; Kim, Y.S.; Lee, W.; Kim, D.H.; Lee, S.; et al. Abstract 1826: Comprehensive preclinical study on GI-101, a novel CD80-IgG4-IL2 variant protein, as a therapeutic antibody candidate with bispecific immuno-oncology target. Cancer Res. 2021, 81, 1826. [Google Scholar] [CrossRef]
- Grohmann, U.; Belladonna, M.L.; Bianchi, R.; Orabona, C.; Ayroldi, E.; Fioretti, M.C.; Puccetti, P. IL-12 acts directly on DC to promote nuclear localization of NF-κB and primes DC for IL-12 production. Immunity 1998, 9, 315–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aste-Amezaga, M.; D’Andrea, A.; Kubin, M.; Trinchieri, G. Cooperation of natural killer cell stimulatory factor/interleukin-12 with other stimuli in the induction of cytokines and cytotoxic cell-associated molecules in human T and NK cells. Cell. Immunol. 1994, 156, 480–492. [Google Scholar] [CrossRef]
- Cheever, M.A. Twelve immunotherapy drugs that could cure cancers. Immunol. Rev. 2008, 222, 357–368. [Google Scholar] [CrossRef]
- Lasek, W.; Zagożdżon, R.; Jakobisiak, M. Interleukin 12: Still a promising candidate for tumor immunotherapy? Cancer Immunol. Immunother. 2014, 63, 419–435. [Google Scholar] [CrossRef] [Green Version]
- Greiner, J.W.; Morillon, Y.M., 2nd; Schlom, J. NHS-IL12, a Tumor-Targeting Immunocytokine. Immunotargets Ther. 2021, 10, 155–169. [Google Scholar] [CrossRef] [PubMed]
- Rudman, S.M.; Jameson, M.B.; McKeage, M.J.; Savage, P.; Jodrell, D.I.; Harries, M.; Acton, G.; Erlandsson, F.; Spicer, J.F. A Phase 1 Study of AS1409, a Novel Antibody-Cytokine Fusion Protein, in Patients with Malignant Melanoma or Renal Cell Carcinoma. Clin. Cancer Res. 2011, 17, 1998–2005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, P.; Li, X.; Wang, J.; Gao, D.; Li, Y.; Li, H.; Chu, Y.; Zhang, Z.; Liu, H.; Jiang, G.; et al. Re-designing Interleukin-12 to enhance its safety and potential as an anti-tumor immunotherapeutic agent. Nat. Commun. 2017, 8, 1395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaminska, K.; Czarnecka, A.M.; Escudier, B.; Lian, F.; Szczylik, C. Interleukin-6 as an emerging regulator of renal cell cancer. Urol. Oncol. 2015, 33, 476–485. [Google Scholar] [CrossRef] [PubMed]
- Mihara, M.; Hashizume, M.; Yoshida, H.; Suzuki, M.; Shiina, M. IL-6/IL-6 receptor system and its role in physiological and pathological conditions. Clin. Sci. 2012, 122, 143–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, Q.; Bournazou, E.; Sansone, P.; Berishaj, M.; Gao, S.P.; Daly, L.; Wels, J.; Theilen, T.; Granitto, S.; Zhang, X.; et al. The IL-6/JAK/Stat3 feed-forward loop drives tumorigenesis and metastasis. Neoplasia 2013, 15, 848–862. [Google Scholar] [CrossRef] [Green Version]
- Johnson, D.E.; O’Keefe, R.A.; Grandis, J.R. Targeting the IL-6/JAK/STAT3 signalling axis in cancer. Nat. Rev. Clin. Oncol. 2018, 15, 234–248. [Google Scholar] [CrossRef] [PubMed]
- Kumari, N.; Dwarakanath, B.S.; Das, A.; Bhatt, A.N. Role of interleukin-6 in cancer progression and therapeutic resistance. Tumour Biol. 2016, 37, 11553–11572. [Google Scholar] [CrossRef]
- Horiguchi, A.; Oya, M.; Marumo, K.; Murai, M. STAT3, but not ERKs, mediates the IL-6-induced proliferation of renal cancer cells, ACHN and 769P. Kidney Int. 2002, 61, 926–938. [Google Scholar] [CrossRef] [Green Version]
- Chang, W.H.; Lai, A.G. An immunoevasive strategy through clinically-relevant pan-cancer genomic and transcriptomic alterations of JAK-STAT signaling components. Mol. Med. 2019, 25, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linker-Israeli, M.; Deans, R.J.; Wallace, D.J.; Prehn, J.; Ozeri-Chen, T.; Klinenberg, J.R. Elevated levels of endogenous IL-6 in systemic lupus erythematosus. A putative role in pathogenesis. J. Immunol. 1991, 147, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Su, H.; Lei, C.T.; Zhang, C. Interleukin-6 Signaling Pathway and Its Role in Kidney Disease: An Update. Front. Immunol. 2017, 8, 405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pecoits-Filho, R.; Heimburger, O.; Barany, P.; Suliman, M.; Fehrman-Ekholm, I.; Lindholm, B.; Stenvinkel, P. Associations between circulating inflammatory markers and residual renal function in CRF patients. Am. J. Kidney Dis. 2003, 41, 1212–1218. [Google Scholar] [CrossRef] [PubMed]
- Dalla Vestra, M.; Mussap, M.; Gallina, P.; Bruseghin, M.; Cernigoi, A.M.; Saller, A.; Plebani, M.; Fioretto, P. Acute-phase markers of inflammation and glomerular structure in patients with type 2 diabetes. J. Am. Soc. Nephrol. 2005, 16 (Suppl. S1), S78–S82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arakawa, T.; Masaki, T.; Hirai, T.; Doi, S.; Kuratsune, M.; Arihiro, K.; Kohno, N.; Yorioka, N. Activation of signal transducer and activator of transcription 3 correlates with cell proliferation and renal injury in human glomerulonephritis. Nephrol. Dial. Transplant. 2008, 23, 3418–3426. [Google Scholar] [CrossRef] [PubMed]
- Horii, Y.; Iwano, M.; Hirata, E.; Shiiki, M.; Fujii, Y.; Dohi, K.; Ishikawa, H. Role of interleukin-6 in the progression of mesangial proliferative glomerulonephritis. Kidney Int. Suppl. 1993, 39, S71–S75. [Google Scholar] [PubMed]
- Zhang, J.; Li, Y.; Shan, K.; Wang, L.; Qiu, W.; Lu, Y.; Zhao, D.; Zhu, G.; He, F.; Wang, Y. Sublytic C5b-9 induces IL-6 and TGF-beta1 production by glomerular mesangial cells in rat Thy-1 nephritis through p300-mediated C/EBPbeta acetylation. FASEB J. 2014, 28, 1511–1525. [Google Scholar] [CrossRef] [PubMed]
- Roofeh, D.; Lin, C.J.F.; Goldin, J.; Kim, G.H.; Furst, D.E.; Denton, C.P.; Huang, S.; Khanna, D.; the focuSSced Investigators. Tocilizumab Prevents Progression of Early Systemic Sclerosis-Associated Interstitial Lung Disease. Arthritis Rheumatol. 2021, 73, 1301–1310. [Google Scholar] [CrossRef]
- Gomez-Guerrero, C.; Lopez-Armada, M.J.; Gonzalez, E.; Egido, J. Soluble IgA and IgG aggregates are catabolized by cultured rat mesangial cells and induce production of TNF-alpha and IL-6, and proliferation. J. Immunol. 1994, 153, 5247–5255. [Google Scholar] [CrossRef]
- Kielar, M.L.; John, R.; Bennett, M.; Richardson, J.A.; Shelton, J.M.; Chen, L.; Jeyarajah, D.R.; Zhou, X.J.; Zhou, H.; Chiquett, B.; et al. Maladaptive role of IL-6 in ischemic acute renal failure. J. Am. Soc. Nephrol. 2005, 16, 3315–3325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranganathan, P.; Jayakumar, C.; Ramesh, G. Proximal tubule-specific overexpression of netrin-1 suppresses acute kidney injury-induced interstitial fibrosis and glomerulosclerosis through suppression of IL-6/STAT3 signaling. Am. J. Physiol. Renal Physiol. 2013, 304, F1054–F1065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blay, J.Y.; Negrier, S.; Combaret, V.; Attali, S.; Goillot, E.; Merrouche, Y.; Mercatello, A.; Ravault, A.; Tourani, J.M.; Moskovtchenko, J.F.; et al. Serum level of interleukin 6 as a prognosis factor in metastatic renal cell carcinoma. Cancer Res. 1992, 52, 3317–3322. [Google Scholar] [PubMed]
- Ljungberg, B.; Grankvist, K.; Rasmuson, T. Serum interleukin-6 in relation to acute-phase reactants and survival in patients with renal cell carcinoma. Eur. J. Cancer 1997, 33, 1794–1798. [Google Scholar] [CrossRef] [PubMed]
- Negrier, S.; Perol, D.; Menetrier-Caux, C.; Escudier, B.; Pallardy, M.; Ravaud, A.; Douillard, J.Y.; Chevreau, C.; Lasset, C.; Blay, J.Y.; et al. Interleukin-6, interleukin-10, and vascular endothelial growth factor in metastatic renal cell carcinoma: Prognostic value of interleukin-6--from the Groupe Francais d’Immunotherapie. J. Clin. Oncol. 2004, 22, 2371–2378. [Google Scholar] [CrossRef]
- Hrab, M.; Olek-Hrab, K.; Antczak, A.; Kwias, Z.; Milecki, T. Interleukin-6 (IL-6) and C-reactive protein (CRP) concentration prior to total nephrectomy are prognostic factors in localized renal cell carcinoma (RCC). Rep. Pract. Oncol. Radiother. 2013, 18, 304–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gudbrandsdottir, G.; Aarstad, H.H.; Bostad, L.; Hjelle, K.M.; Aarstad, H.J.; Bruserud, O.; Tvedt, T.H.A.; Beisland, C. Serum levels of the IL-6 family of cytokines predict prognosis in renal cell carcinoma (RCC). Cancer Immunol. Immunother. 2021, 70, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Polimeno, M.; Napolitano, M.; Costantini, S.; Portella, L.; Esposito, A.; Capone, F.; Guerriero, E.; Trotta, A.; Zanotta, S.; Pucci, L.; et al. Regulatory T cells, interleukin (IL)-6, IL-8, vascular endothelial growth factor (VEGF), CXCL10, CXCL11, epidermal growth factor (EGF) and hepatocyte growth factor (HGF) as surrogate markers of host immunity in patients with renal cell carcinoma. BJU Int. 2013, 112, 686–696. [Google Scholar] [CrossRef] [PubMed]
- Porta, C.; Paglino, C.; Imarisio, I.; Ganini, C.; Sacchi, L.; Quaglini, S.; Giunta, V.; De Amici, M. Changes in circulating pro-angiogenic cytokines, other than VEGF, before progression to sunitinib therapy in advanced renal cell carcinoma patients. Oncology 2013, 84, 115–122. [Google Scholar] [CrossRef]
- Chen, Q.; Yang, D.; Zong, H.; Zhu, L.; Wang, L.; Wang, X.; Zhu, X.; Song, X.; Wang, J. Growth-induced stress enhances epithelial-mesenchymal transition induced by IL-6 in clear cell renal cell carcinoma via the Akt/GSK-3beta/beta-catenin signaling pathway. Oncogenesis 2017, 6, e375. [Google Scholar] [CrossRef] [Green Version]
- Esteban, E.; Exposito, F.; Crespo, G.; Lambea, J.; Pinto, A.; Puente, J.; Arranz, J.A.; Redrado, M.; Rodriguez-Antona, C.; de Andrea, C.; et al. Circulating Levels of the Interferon-gamma-Regulated Chemokines CXCL10/CXCL11, IL-6 and HGF Predict Outcome in Metastatic Renal Cell Carcinoma Patients Treated with Antiangiogenic Therapy. Cancers 2021, 13, 2849. [Google Scholar] [CrossRef]
- Ishibashi, K.; Haber, T.; Breuksch, I.; Gebhard, S.; Sugino, T.; Kubo, H.; Hata, J.; Koguchi, T.; Yabe, M.; Kataoka, M.; et al. Overriding TKI resistance of renal cell carcinoma by combination therapy with IL-6 receptor blockade. Oncotarget 2017, 8, 55230–55245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Liu, H.; Xu, L.; An, H.; Liu, W.; Liu, Y.; Lin, Z.; Xu, J. p21-activated kinase 1 determines stem-like phenotype and sunitinib resistance via NF-kappaB/IL-6 activation in renal cell carcinoma. Cell Death Dis. 2015, 6, e1637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quan, Z.; He, Y.; Luo, C.; Xia, Y.; Zhao, Y.; Liu, N.; Wu, X. Interleukin 6 induces cell proliferation of clear cell renal cell carcinoma by suppressing hepaCAM via the STAT3-dependent up-regulation of DNMT1 or DNMT3b. Cell Signal. 2017, 32, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Yang, R.; Wei, R.; Du, Y.; He, P.; Liu, X. Pan-cancer analysis reveals RIPK2 predicts prognosis and promotes immune therapy resistance via triggering cytotoxic T lymphocytes dysfunction. Mol. Med. 2022, 28, 47. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Zhou, B.; Qian, C.; Vasquez, A.; Kamra, M.; Chatterjee, A.; Lee, Y.J.; Yuan, X.; Ellis, L.; Di Vizio, D.; et al. Receptor-interacting protein kinase 2 (RIPK2) stabilizes c-Myc and is a therapeutic target in prostate cancer metastasis. Nat. Commun. 2022, 13, 669. [Google Scholar] [CrossRef] [PubMed]
- Rossi, J.F.; Negrier, S.; James, N.D.; Kocak, I.; Hawkins, R.; Davis, H.; Prabhakar, U.; Qin, X.; Mulders, P.; Berns, B. A phase I/II study of siltuximab (CNTO 328), an anti-interleukin-6 monoclonal antibody, in metastatic renal cell cancer. Br. J. Cancer 2010, 103, 1154–1162. [Google Scholar] [CrossRef] [Green Version]
- Schafer, J.A.; Kjesbo, N.K.; Gleason, P.P. Formulary review of 2 new biologic agents: Tocilizumab for rheumatoid arthritis and ustekinumab for plaque psoriasis. J. Manag. Care Pharm. 2010, 16, 402–416. [Google Scholar] [CrossRef] [Green Version]
- Ruperto, N.; Brunner, H.I.; Ramanan, A.V.; Horneff, G.; Cuttica, R.; Henrickson, M.; Anton, J.; Boteanu, A.L.; Penades, I.C.; Minden, K.; et al. Subcutaneous dosing regimens of tocilizumab in children with systemic or polyarticular juvenile idiopathic arthritis. Rheumatology 2021, 60, 4568–4580. [Google Scholar] [CrossRef] [PubMed]
- Brunner, H.I.; Ruperto, N.; Zuber, Z.; Cuttica, R.; Keltsev, V.; Xavier, R.M.; Burgos-Vargas, R.; Penades, I.C.; Silverman, E.D.; Espada, G.; et al. Efficacy and Safety of Tocilizumab for Polyarticular-Course Juvenile Idiopathic Arthritis in the Open-Label Two-Year Extension of a Phase III Trial. Arthritis Rheumatol. 2021, 73, 530–541. [Google Scholar] [CrossRef]
- De Benedetti, F.; Brunner, H.I.; Ruperto, N.; Kenwright, A.; Wright, S.; Calvo, I.; Cuttica, R.; Ravelli, A.; Schneider, R.; Woo, P.; et al. Randomized trial of tocilizumab in systemic juvenile idiopathic arthritis. N. Engl. J. Med. 2012, 367, 2385–2395. [Google Scholar] [CrossRef] [Green Version]
- Stone, J.H.; Klearman, M.; Collinson, N. Trial of Tocilizumab in Giant-Cell Arteritis. N. Engl. J. Med. 2017, 377, 1494–1495. [Google Scholar] [CrossRef] [PubMed]
- Le, R.Q.; Li, L.; Yuan, W.; Shord, S.S.; Nie, L.; Habtemariam, B.A.; Przepiorka, D.; Farrell, A.T.; Pazdur, R. FDA Approval Summary: Tocilizumab for Treatment of Chimeric Antigen Receptor T Cell-Induced Severe or Life-Threatening Cytokine Release Syndrome. Oncologist 2018, 23, 943–947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khanna, D.; Denton, C.P.; Jahreis, A.; van Laar, J.M.; Frech, T.M.; Anderson, M.E.; Baron, M.; Chung, L.; Fierlbeck, G.; Lakshminarayanan, S.; et al. Safety and efficacy of subcutaneous tocilizumab in adults with systemic sclerosis (faSScinate): A phase 2, randomised, controlled trial. Lancet 2016, 387, 2630–2640. [Google Scholar] [CrossRef] [PubMed]
- Khanna, D.; Lin, C.J.F.; Furst, D.E.; Goldin, J.; Kim, G.; Kuwana, M.; Allanore, Y.; Matucci-Cerinic, M.; Distler, O.; Shima, Y.; et al. Tocilizumab in systemic sclerosis: A randomised, double-blind, placebo-controlled, phase 3 trial. Lancet Respir. Med. 2020, 8, 963–974. [Google Scholar] [CrossRef]
- Angus, D.C.; Berry, S.; Lewis, R.J.; Al-Beidh, F.; Arabi, Y.; van Bentum-Puijk, W.; Bhimani, Z.; Bonten, M.; Broglio, K.; Brunkhorst, F.; et al. The REMAP-CAP (Randomized Embedded Multifactorial Adaptive Platform for Community-acquired Pneumonia) Study. Rationale and Design. Ann. Am. Thorac. Soc. 2020, 17, 879–891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Group, R.C. Tocilizumab in patients admitted to hospital with COVID-19 (RECOVERY): A randomised, controlled, open-label, platform trial. Lancet 2021, 397, 1637–1645. [Google Scholar] [CrossRef]
- Ishibashi, K.; Koguchi, T.; Matsuoka, K.; Onagi, A.; Tanji, R.; Takinami-Honda, R.; Hoshi, S.; Onoda, M.; Kurimura, Y.; Hata, J.; et al. Interleukin-6 induces drug resistance in renal cell carcinoma. Fukushima J. Med. Sci. 2018, 64, 103–110. [Google Scholar] [CrossRef] [Green Version]
- Oguro, T.; Ishibashi, K.; Sugino, T.; Hashimoto, K.; Tomita, S.; Takahashi, N.; Yanagida, T.; Haga, N.; Aikawa, K.; Suzutani, T.; et al. Humanised antihuman IL-6R antibody with interferon inhibits renal cell carcinoma cell growth in vitro and in vivo through suppressed SOCS3 expression. Eur. J. Cancer 2013, 49, 1715–1724. [Google Scholar] [CrossRef]
- Anders, H.J. Of Inflammasomes and Alarmins: IL-1beta and IL-1alpha in Kidney Disease. J. Am. Soc. Nephrol. 2016, 27, 2564–2575. [Google Scholar] [CrossRef] [Green Version]
- Chittezhath, M.; Dhillon, M.K.; Lim, J.Y.; Laoui, D.; Shalova, I.N.; Teo, Y.L.; Chen, J.; Kamaraj, R.; Raman, L.; Lum, J.; et al. Molecular profiling reveals a tumor-promoting phenotype of monocytes and macrophages in human cancer progression. Immunity 2014, 41, 815–829. [Google Scholar] [CrossRef] [Green Version]
- Krelin, Y.; Voronov, E.; Dotan, S.; Elkabets, M.; Reich, E.; Fogel, M.; Huszar, M.; Iwakura, Y.; Segal, S.; Dinarello, C.A.; et al. Interleukin-1beta-driven inflammation promotes the development and invasiveness of chemical carcinogen-induced tumors. Cancer Res. 2007, 67, 1062–1071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voronov, E.; Shouval, D.S.; Krelin, Y.; Cagnano, E.; Benharroch, D.; Iwakura, Y.; Dinarello, C.A.; Apte, R.N. IL-1 is required for tumor invasiveness and angiogenesis. Proc. Natl. Acad. Sci. USA 2003, 100, 2645–2650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaplanov, I.; Carmi, Y.; Kornetsky, R.; Shemesh, A.; Shurin, G.V.; Shurin, M.R.; Dinarello, C.A.; Voronov, E.; Apte, R.N. Blocking IL-1β reverses the immunosuppression in mouse breast cancer and synergizes with anti-PD-1 for tumor abrogation. Proc. Natl. Acad. Sci. USA 2019, 116, 1361–1369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Najjar, Y.G.; Rayman, P.; Jia, X.; Pavicic, P.G., Jr.; Rini, B.I.; Tannenbaum, C.; Ko, J.; Haywood, S.; Cohen, P.; Hamilton, T.; et al. Myeloid-Derived Suppressor Cell Subset Accumulation in Renal Cell Carcinoma Parenchyma Is Associated with Intratumoral Expression of IL1β, IL8, CXCL5, and Mip-1α. Clin. Cancer Res. 2017, 23, 2346–2355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aggen, D.H.; Ager, C.R.; Obradovic, A.Z.; Chowdhury, N.; Ghasemzadeh, A.; Mao, W.; Chaimowitz, M.G.; Lopez-Bujanda, Z.A.; Spina, C.S.; Hawley, J.E.; et al. Blocking IL1 Beta Promotes Tumor Regression and Remodeling of the Myeloid Compartment in a Renal Cell Carcinoma Model: Multidimensional Analyses. Clin. Cancer Res. 2021, 27, 608–621. [Google Scholar] [CrossRef] [PubMed]
- Balkwill, F. Tumour necrosis factor and cancer. Nat. Rev. Cancer 2009, 9, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Sethi, G.; Sung, B.; Aggarwal, B.B. TNF: A master switch for inflammation to cancer. Front. Biosci. Landmark 2008, 13, 5094–5107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, N.; Ikemoto, S.; Narita, K.; Sugimura, K.; Wada, S.; Yasumoto, R.; Kishimoto, T.; Nakatani, T. Interleukin-6, tumour necrosis factor α and interleukin-1β in patients with renal cell carcinoma. Br. J. Cancer 2002, 86, 1396–1400. [Google Scholar] [CrossRef]
- Mikami, S.; Mizuno, R.; Kosaka, T.; Saya, H.; Oya, M.; Okada, Y. Expression of TNF-α and CD 44 is implicated in poor prognosis, cancer cell invasion, metastasis and resistance to the sunitinib treatment in clear cell renal cell carcinomas. Int. J. Cancer 2015, 136, 1504–1514. [Google Scholar] [CrossRef]
- Ho, M.-Y.; Tang, S.-J.; Chuang, M.-J.; Cha, T.-L.; Li, J.-Y.; Sun, G.-H.; Sun, K.-H. TNF-α Induces Epithelial–Mesenchymal Transition of Renal Cell Carcinoma Cells via a GSK3β-Dependent MechanismGSK-3β Inactivation Enhances Tumorigenicity of RCCs. Mol. Cancer Res. 2012, 10, 1109–1119. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.-T.; Sun, G.-H.; Hsu, C.-Y.; Huang, C.-S.; Wu, Y.-H.; Wang, H.-H.; Sun, K.-H. Tumor necrosis factor-α induces epithelial–mesenchymal transition of renal cell carcinoma cells via a nuclear factor kappa B-independent mechanism. Exp. Biol. Med. 2011, 236, 1022–1029. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Jiao, M.; Wu, K.; Li, L.; Zhu, G.; Wang, X.; He, D.; Wu, D. TNF-α induced epithelial mesenchymal transition increases stemness properties in renal cell carcinoma cells. Int. J. Clin. Exp. Med. 2014, 7, 4951. [Google Scholar] [PubMed]
- Chuang, M.J.; Sun, K.H.; Tang, S.J.; Deng, M.W.; Wu, Y.H.; Sung, J.S.; Cha, T.L.; Sun, G.H. Tumor-derived tumor necrosis factor-alpha promotes progression and epithelial-mesenchymal transition in renal cell carcinoma cells. Cancer Sci. 2008, 99, 905–913. [Google Scholar] [CrossRef] [PubMed]
- Sun, K.H.; Sun, G.H.; Wu, Y.C.; Ko, B.J.; Hsu, H.T.; Wu, S.T. TNF-α augments CXCR2 and CXCR3 to promote progression of renal cell carcinoma. J. Cell. Mol. Med. 2016, 20, 2020–2028. [Google Scholar] [CrossRef] [Green Version]
- Zhong, M.; Zhu, M.; Liu, Y.; Lin, Y.; Wang, L.; Ye, Y.; Chen, H.; Yang, Y.; Zhuang, G.; Huang, J. TNFAIP8 promotes the migration of clear cell renal cell carcinoma by regulating the EMT. J. Cancer 2020, 11, 3061–3071. [Google Scholar] [CrossRef] [PubMed]
- Perez-Gracia, J.L.; Prior, C.; Guillén-Grima, F.; Segura, V.; Gonzalez, A.; Panizo, A.; Melero, I.; Grande-Pulido, E.; Gurpide, A.; Gil-Bazo, I.; et al. Identification of TNF-alpha and MMP-9 as potential baseline predictive serum markers of sunitinib activity in patients with renal cell carcinoma using a human cytokine array. Br. J. Cancer 2009, 101, 1876–1883. [Google Scholar] [CrossRef] [Green Version]
- Raval, G.; Biswas, S.; Rayman, P.; Biswas, K.; Sa, G.; Ghosh, S.; Thornton, M.; Hilston, C.; Das, T.; Bukowski, R.; et al. TNF-α Induction of GM2 Expression on Renal Cell Carcinomas Promotes T Cell Dysfunction1. J. Immunol. 2007, 178, 6642–6652. [Google Scholar] [CrossRef] [Green Version]
- Hillman, G.G.; Puri, R.K.; Kukuruga, M.A.; Pontes, J.E.; Haas, G.P. Growth and major histocompatibility antigen expression regulation by IL-4, interferon-gamma (IFN-gamma) and tumour necrosis factor-alpha (TNF-alpha) on human renal cell carcinoma. Clin. Exp. Immunol. 1994, 96, 476–483. [Google Scholar] [CrossRef] [PubMed]
- Falkensammer, C.; Jöhrer, K.; Gander, H.; Ramoner, R.; Putz, T.; Rahm, A.; Greil, R.; Bartsch, G.; Thurnher, M. IL-4 inhibits the TNF-alpha induced proliferation of renal cell carcinoma (RCC) and cooperates with TNF-alpha to induce apoptotic and cytokine responses by RCC: Implications for antitumor immune responses. Cancer Immunol. Immunother. 2006, 55, 1228–1237. [Google Scholar] [CrossRef]
- Bauer, S.; Oosterwijk-Wakka, J.C.; Adrian, N.; Oosterwijk, E.; Fischer, E.; Wüest, T.; Stenner, F.; Perani, A.; Cohen, L.S.; Knuth, A.K.; et al. Targeted therapy of renal cell carcinoma: Synergistic activity of cG250-TNF and IFNg. Int. J. Cancer 2009, 125, 115–123. [Google Scholar] [CrossRef]
- Harrison, M.L.; Obermueller, E.; Maisey, N.R.; Hoare, S.; Edmonds, K.; Li, N.F.; Chao, D.; Hall, K.; Lee, C.; Timotheadou, E.; et al. Tumor necrosis factor alpha as a new target for renal cell carcinoma: Two sequential phase II trials of infliximab at standard and high dose. J. Clin. Oncol. 2007, 25, 4542–4549. [Google Scholar] [CrossRef]
- Larkin, J.M.; Ferguson, T.R.; Pickering, L.M.; Edmonds, K.; James, M.G.; Thomas, K.; Banerji, U.; Berns, B.; de Boer, C.; Gore, M.E. A phase I/II trial of sorafenib and infliximab in advanced renal cell carcinoma. Br. J. Cancer 2010, 103, 1149–1153. [Google Scholar] [CrossRef]
- Gordon, K.J.; Blobe, G.C. Role of transforming growth factor-beta superfamily signaling pathways in human disease. Biochim. Biophys. Acta 2008, 1782, 197–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moustakas, A.; Heldin, C.H. The regulation of TGFbeta signal transduction. Development 2009, 136, 3699–3714. [Google Scholar] [CrossRef] [Green Version]
- Bostrom, A.K.; Lindgren, D.; Johansson, M.E.; Axelson, H. Effects of TGF-beta signaling in clear cell renal cell carcinoma cells. Biochem. Biophys. Res. Commun. 2013, 435, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Massague, J. TGFbeta in Cancer. Cell 2008, 134, 215–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mu, Y.; Gudey, S.K.; Landstrom, M. Non-Smad signaling pathways. Cell Tissue Res. 2012, 347, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.Y.; Liu, X.S.; Huang, X.R.; Yu, X.Q.; Lan, H.Y. Diverse Role of TGF-beta in Kidney Disease. Front. Cell Dev. Biol. 2020, 8, 123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshioka, K.; Takemura, T.; Murakami, K.; Okada, M.; Hino, S.; Miyamoto, H.; Maki, S. Transforming growth factor-beta protein and mRNA in glomeruli in normal and diseased human kidneys. Lab. Investig. 1993, 68, 154–163. [Google Scholar]
- Yamamoto, T.; Watanabe, T.; Ikegaya, N.; Fujigaki, Y.; Matsui, K.; Masaoka, H.; Nagase, M.; Hishida, A. Expression of types I, II, and III TGF-beta receptors in human glomerulonephritis. J. Am. Soc. Nephrol. 1998, 9, 2253–2261. [Google Scholar] [CrossRef]
- Yamamoto, T.; Nakamura, T.; Noble, N.A.; Ruoslahti, E.; Border, W.A. Expression of transforming growth factor beta is elevated in human and experimental diabetic nephropathy. Proc. Natl. Acad. Sci. USA 1993, 90, 1814–1818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, K.; Ziyadeh, F.N.; Alzahabi, B.; McGowan, T.A.; Kapoor, S.; Kurnik, B.R.; Kurnik, P.B.; Weisberg, L.S. Increased renal production of transforming growth factor-beta1 in patients with type II diabetes. Diabetes 1997, 46, 854–859. [Google Scholar] [CrossRef]
- Meng, X.M.; Nikolic-Paterson, D.J.; Lan, H.Y. TGF-beta: The master regulator of fibrosis. Nat. Rev. Nephrol. 2016, 12, 325–338. [Google Scholar] [CrossRef] [PubMed]
- Okuda, S.; Languino, L.R.; Ruoslahti, E.; Border, W.A. Elevated expression of transforming growth factor-beta and proteoglycan production in experimental glomerulonephritis. Possible role in expansion of the mesangial extracellular matrix. J. Clin. Investig. 1990, 86, 453–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isaka, Y. Targeting TGF-beta Signaling in Kidney Fibrosis. Int. J. Mol. Sci. 2018, 19, 2532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomooka, S.; Border, W.A.; Marshall, B.C.; Noble, N.A. Glomerular matrix accumulation is linked to inhibition of the plasmin protease system. Kidney Int. 1992, 42, 1462–1469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, X.M.; Huang, X.R.; Xiao, J.; Chen, H.Y.; Zhong, X.; Chung, A.C.; Lan, H.Y. Diverse roles of TGF-beta receptor II in renal fibrosis and inflammation in vivo and in vitro. J. Pathol. 2012, 227, 175–188. [Google Scholar] [CrossRef]
- Hegele, A.; Varga, Z.; von Knobloch, R.; Heidenreich, A.; Kropf, J.; Hofmann, R. TGF-beta1 in patients with renal cell carcinoma. Urol. Res. 2002, 30, 126–129. [Google Scholar] [CrossRef]
- Mitropoulos, D.; Kiroudi, A.; Christelli, E.; Serafetinidis, E.; Zervas, A.; Anastasiou, I.; Dimopoulos, C. Expression of transforming growth factor beta in renal cell carcinoma and matched non-involved renal tissue. Urol. Res. 2004, 32, 317–322. [Google Scholar] [CrossRef]
- Sitaram, R.T.; Mallikarjuna, P.; Landstrom, M.; Ljungberg, B. Transforming growth factor-beta promotes aggressiveness and invasion of clear cell renal cell carcinoma. Oncotarget 2016, 7, 35917–35931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Copland, J.A.; Luxon, B.A.; Ajani, L.; Maity, T.; Campagnaro, E.; Guo, H.; LeGrand, S.N.; Tamboli, P.; Wood, C.G. Genomic profiling identifies alterations in TGFbeta signaling through loss of TGFbeta receptor expression in human renal cell carcinogenesis and progression. Oncogene 2003, 22, 8053–8062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishida, J.; Miyazono, K.; Ehata, S. Decreased TGFBR3/betaglycan expression enhances the metastatic abilities of renal cell carcinoma cells through TGF-beta-dependent and -independent mechanisms. Oncogene 2018, 37, 2197–2212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukasawa, H.; Yamamoto, T.; Fujigaki, Y.; Misaki, T.; Ohashi, N.; Takayama, T.; Suzuki, S.; Mugiya, S.; Oda, T.; Uchida, C.; et al. Reduction of transforming growth factor-beta type II receptor is caused by the enhanced ubiquitin-dependent degradation in human renal cell carcinoma. Int. J. Cancer 2010, 127, 1517–1525. [Google Scholar] [CrossRef] [PubMed]
- Margulis, V.; Maity, T.; Zhang, X.Y.; Cooper, S.J.; Copland, J.A.; Wood, C.G. Type III transforming growth factor-beta (TGF-beta) receptor mediates apoptosis in renal cell carcinoma independent of the canonical TGF-beta signaling pathway. Clin. Cancer Res. 2008, 14, 5722–5730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tretbar, S.; Krausbeck, P.; Muller, A.; Friedrich, M.; Vaxevanis, C.; Bukur, J.; Jasinski-Bergner, S.; Seliger, B. TGF-beta inducible epithelial-to-mesenchymal transition in renal cell carcinoma. Oncotarget 2019, 10, 1507–1524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, D.; Feng, H.; Yang, J.; Yin, A.; Qian, A.; Sugiyama, H. Landscape of immune cell infiltration in clear cell renal cell carcinoma to aid immunotherapy. Cancer Sci. 2021, 112, 2126–2139. [Google Scholar] [CrossRef] [PubMed]
- Kaur, G.; Li, C.G.; Chantry, A.; Stayner, C.; Horsfield, J.; Eccles, M.R. SMAD proteins directly suppress PAX2 transcription downstream of transforming growth factor-beta 1 (TGF-beta1) signalling in renal cell carcinoma. Oncotarget 2018, 9, 26852–26867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feldkoren, B.; Hutchinson, R.; Rapoport, Y.; Mahajan, A.; Margulis, V. Integrin signaling potentiates transforming growth factor-beta 1 (TGF-beta1) dependent down-regulation of E-Cadherin expression—Important implications for epithelial to mesenchymal transition (EMT) in renal cell carcinoma. Exp. Cell Res. 2017, 355, 57–66. [Google Scholar] [CrossRef]
- Tian, T.; Fu, X.; Hu, L.; Yang, X.; Sun, P.; Sun, F. FAST1 Predicts Poor Survival of Renal Carcinoma and Promotes Its Progression Through the TGF-beta/Smad Pathway. Onco Targets Ther. 2021, 14, 1487–1499. [Google Scholar] [CrossRef]
- Ananth, S.; Knebelmann, B.; Gruning, W.; Dhanabal, M.; Walz, G.; Stillman, I.E.; Sukhatme, V.P. Transforming growth factor beta1 is a target for the von Hippel-Lindau tumor suppressor and a critical growth factor for clear cell renal carcinoma. Cancer Res. 1999, 59, 2210–2216. [Google Scholar]
- Meng, F.; Li, Y.; Tian, X.; Fu, L.; Yin, Y.; Sui, C.; Ma, P.; Jiang, Y. Identification of TGF-beta-activated kinase 1 as a possible novel target for renal cell carcinoma intervention. Biochem. Biophys. Res. Commun. 2014, 453, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Busse, A.; Asemissen, A.; Nonnenmacher, A.; Ochsenreither, S.; Fusi, A.; Braun, F.; Stather, D.; Schmittel, A.; Miller, K.; Thiel, E.; et al. Systemic immune tuning in renal cell carcinoma: Favorable prognostic impact of TGF-beta1 mRNA expression in peripheral blood mononuclear cells. J. Immunother. 2011, 34, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Zhang, Q.; Zhen, Q.; Zhao, Y.; Liu, N.; Li, T.; Hao, Y.; Zhang, Y.; Luo, C.; Wu, X. Negative regulation of tumor-infiltrating NK cell in clear cell renal cell carcinoma patients through the exosomal pathway. Oncotarget 2017, 8, 37783–37795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Wen, W.; Yuan, J.; Helfand, B.; Li, Y.; Shi, C.; Tian, F.; Zheng, J.; Wang, F.; Chen, L.; et al. Immunotherapy for human renal cell carcinoma by adoptive transfer of autologous transforming growth factor beta-insensitive CD8+ T cells. Clin. Cancer Res. 2010, 16, 164–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voelker, J.; Berg, P.H.; Sheetz, M.; Duffin, K.; Shen, T.; Moser, B.; Greene, T.; Blumenthal, S.S.; Rychlik, I.; Yagil, Y.; et al. Anti-TGF-beta1 Antibody Therapy in Patients with Diabetic Nephropathy. J. Am. Soc. Nephrol. 2017, 28, 953–962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, M.E.; Smith, D.C.; Branton, M.H.; Penzak, S.R.; Kopp, J.B. Pirfenidone slows renal function decline in patients with focal segmental glomerulosclerosis. Clin. J. Am. Soc. Nephrol. 2007, 2, 906–913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, D.D.; Ding, M.; Tong, P.; Chong, Y.Y.; Gu, W.Y.; Li, Y.; Fang, X.J.; Li, N. Synergistic effects of low-dose chemotherapy and T cells in renal cell carcinoma. Oncol. Rep. 2020, 44, 897–908. [Google Scholar] [CrossRef]
- Liu, L.; Wang, Q.; Mao, J.; Qin, T.; Sun, Y.; Yang, J.; Han, Y.; Li, L.; Li, Q. Salinomycin suppresses cancer cell stemness and attenuates TGF-beta-induced epithelial-mesenchymal transition of renal cell carcinoma cells. Chem. Biol. Interact. 2018, 296, 145–153. [Google Scholar] [CrossRef]
- Wang, G.; Zhou, X.; Guo, Z.; Huang, N.; Li, J.; Lv, Y.; Han, L.; Zheng, W.; Xu, D.; Chai, D.; et al. The Anti-fibrosis drug Pirfenidone modifies the immunosuppressive tumor microenvironment and prevents the progression of renal cell carcinoma by inhibiting tumor autocrine TGF-β. Cancer Biol. Ther. 2022, 23, 150–162. [Google Scholar] [CrossRef]
- Heidegger, I.; Pircher, A.; Pichler, R. Targeting the Tumor Microenvironment in Renal Cell Cancer Biology and Therapy. Front Oncol 2019, 9, 490. [Google Scholar] [CrossRef] [Green Version]
- Song, W.; He, D.; Chen, Y.; Yeh, C.R.; Hsu, I.; Huang, Q.; Zhang, X.; Chang, L.S.; Zuo, L.; Chen, J.; et al. Targeting newly identified ERβ/TGF-β1/SMAD3 signals with the FDA-approved anti-estrogen Faslodex or an ERβ selective antagonist in renal cell carcinoma. Mol. Oncol. 2018, 12, 2055–2071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurujeyalakshmi, G.; Hollinger, M.A.; Giri, S.N. Pirfenidone inhibits PDGF isoforms in bleomycin hamster model of lung fibrosis at the translational level. Am. J. Physiol. 1999, 276, L311–L318. [Google Scholar] [CrossRef] [PubMed]
- Iyer, S.N.; Gurujeyalakshmi, G.; Giri, S.N. Effects of pirfenidone on transforming growth factor-beta gene expression at the transcriptional level in bleomycin hamster model of lung fibrosis. J. Pharmacol. Exp. Ther. 1999, 291, 367–373. [Google Scholar] [PubMed]
- Iyer, S.N.; Gurujeyalakshmi, G.; Giri, S.N. Effects of pirfenidone on procollagen gene expression at the transcriptional level in bleomycin hamster model of lung fibrosis. J. Pharmacol. Exp. Ther. 1999, 289, 211–218. [Google Scholar] [PubMed]
- Chan, M.K.; Chung, J.Y.; Tang, P.C.; Chan, A.S.; Ho, J.Y.; Lin, T.P.; Chen, J.; Leung, K.T.; To, K.F.; Lan, H.Y.; et al. TGF-beta signaling networks in the tumor microenvironment. Cancer Lett 2022, 550, 215925. [Google Scholar] [CrossRef] [PubMed]
- Vincenti, F.; Fervenza, F.C.; Campbell, K.N.; Diaz, M.; Gesualdo, L.; Nelson, P.; Praga, M.; Radhakrishnan, J.; Sellin, L.; Singh, A.; et al. A Phase 2, Double-Blind, Placebo-Controlled, Randomized Study of Fresolimumab in Patients with Steroid-Resistant Primary Focal Segmental Glomerulosclerosis. Kidney Int. Rep. 2017, 2, 800–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, J.C.; Tan, A.R.; Olencki, T.E.; Shapiro, G.I.; Dezube, B.J.; Reiss, M.; Hsu, F.J.; Berzofsky, J.A.; Lawrence, D.P. Phase I study of GC1008 (fresolimumab): A human anti-transforming growth factor-beta (TGFbeta) monoclonal antibody in patients with advanced malignant melanoma or renal cell carcinoma. PLoS ONE 2014, 9, e90353. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, A.F.; Ten Dijke, P.; Zhu, H.J. On-Target Anti-TGF-beta Therapies Are Not Succeeding in Clinical Cancer Treatments: What Are Remaining Challenges? Front. Cell Dev. Biol. 2020, 8, 605. [Google Scholar] [CrossRef]
- Dodagatta-Marri, E.; Meyer, D.S.; Reeves, M.Q.; Paniagua, R.; To, M.D.; Binnewies, M.; Broz, M.L.; Mori, H.; Wu, D.; Adoumie, M.; et al. α-PD-1 therapy elevates Treg/Th balance and increases tumor cell pSmad3 that are both targeted by α-TGFβ antibody to promote durable rejection and immunity in squamous cell carcinomas. J. Immunother. Cancer 2019, 7, 62. [Google Scholar] [CrossRef]
- Nagarsheth, N.; Wicha, M.S.; Zou, W. Chemokines in the cancer microenvironment and their relevance in cancer immunotherapy. Nat. Rev. Immunol. 2017, 17, 559–572. [Google Scholar] [CrossRef] [Green Version]
- Vandercappellen, J.; Van Damme, J.; Struyf, S. The role of CXC chemokines and their receptors in cancer. Cancer Lett. 2008, 267, 226–244. [Google Scholar] [CrossRef]
- Schott, A.F.; Goldstein, L.J.; Cristofanilli, M.; Ruffini, P.A.; McCanna, S.; Reuben, J.M.; Perez, R.P.; Kato, G.; Wicha, M. Phase Ib Pilot Study to Evaluate Reparixin in Combination with Weekly Paclitaxel in Patients with HER-2-Negative Metastatic Breast Cancer. Clin. Cancer Res. 2017, 23, 5358–5365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dufies, M.; Grytsai, O.; Ronco, C.; Camara, O.; Ambrosetti, D.; Hagege, A.; Parola, J.; Mateo, L.; Ayrault, M.; Giuliano, S.; et al. New CXCR1/CXCR2 inhibitors represent an effective treatment for kidney or head and neck cancers sensitive or refractory to reference treatments. Theranostics 2019, 9, 5332–5346. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, M.E.; Mezzapelle, R. The Chemokine Receptor CXCR4 in Cell Proliferation and Tissue Regeneration. Front. Immunol. 2020, 11, 2109. [Google Scholar] [CrossRef] [PubMed]
- Romoli, S.; Angelotti, M.L.; Antonelli, G.; Kumar Vr, S.; Mulay, S.R.; Desai, J.; Anguiano Gomez, L.; Thomasova, D.; Eulberg, D.; Klussmann, S.; et al. CXCL12 blockade preferentially regenerates lost podocytes in cortical nephrons by targeting an intrinsic podocyte-progenitor feedback mechanism. Kidney Int. 2018, 94, 1111–1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, Y.; Long, Q.; Guan, B.; Mu, L. Prognostic Value of High CXCR4 Expression in Renal Cell Carcinoma: A System Review and Meta-Analysis. Dis. Markers 2015, 2015, 568980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, D.; Gassenmaier, M.; Maruschke, M.; Riesenberg, R.; Pohla, H.; Stief, C.G.; Zimmermann, W.; Buchner, A. Expression and Prognostic Significance of a Comprehensive Epithelial-Mesenchymal Transition Gene Set in Renal Cell Carcinoma. J. Urol. 2014, 191, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.M.; Ji, S.; Li, Y.; Fu, L.Y.; Jiang, T.; Meng, F.D. β-Catenin promotes cell proliferation, migration, and invasion but induces apoptosis in renal cell carcinoma. Onco Targets Ther. 2017, 10, 711–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDermott, D.F.; Vaishampayan, U.; Matrana, M.; Rha, S.Y.; Saavedra, A.Z.; Ho, T.; Keam, B.; Lee, J.L.; Joseph, R.; Ali, S.; et al. 1186PD—Safety and efficacy of the oral CXCR4 inhibitor X4P-001 + axitinib in advanced renal cell carcinoma patients: An analysis of subgroup responses by prior treatment. Ann. Oncol. 2019, 30, v482–v483. [Google Scholar] [CrossRef]
- Choueiri, T.K.; Atkins, M.B.; Rose, T.L.; Alter, R.S.; Ju, Y.; Niland, K.; Wang, Y.; Arbeit, R.; Parasuraman, S.; Gan, L.; et al. A phase 1b trial of the CXCR4 inhibitor mavorixafor and nivolumab in advanced renal cell carcinoma patients with no prior response to nivolumab monotherapy. Investig. New Drugs 2021, 39, 1019–1027. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic overview of molecular mechanisms and potential targets of RCC-associated inflammation. Intricate crosstalk between renal cancer cells and TME, regulates pro-inflammatory mediator release, angiogenesis, tumor cell plasticity, survival and proliferation, immune evasion, thereby leading to drug resistance, tumor progression and metastasis.
Figure 1.
Schematic overview of molecular mechanisms and potential targets of RCC-associated inflammation. Intricate crosstalk between renal cancer cells and TME, regulates pro-inflammatory mediator release, angiogenesis, tumor cell plasticity, survival and proliferation, immune evasion, thereby leading to drug resistance, tumor progression and metastasis.

Figure 2.
(a) Macrophages express a variety of markers, including pro-inflammatory, immune-regulatory, and suppressive markers. While M1 macrophages are responsible for the eradication of pathogens and tumor cells, M2 macrophages have immune suppressive properties inducing an exhausted signature in CD8+ T cells as well as the tendency for tissue repair and tumor progression. (b) DCs with impaired maturation (DCLamp; DCCD83-; DCMHCII,Low) are located outside of tumor-associated tertiary lymphoid structures (NTLS) of RCC tumors and are associated with high levels of PD-L1/PD-L2 and poor prognosis. (c) T reg cells reduce the cytotoxicity of effector T cells, thereby inducing immunosuppressive phenotype in RCC tumors. (d) RCC tumors are abundant in immune cell infiltrates. In contrast to other solid tumor types, infiltration of CD8 + and CD4 + T cells into the TME is often associated with a poor prognosis and advanced tumor grade. Furthermore, inflammatory TME contains an elevated number of polyclonal exhausted CD8+Tim-3+/Lag-3+ TIL, associated with strong infiltration of dysfunctional DCs. PSP: pseudoprogression.
Figure 2.
(a) Macrophages express a variety of markers, including pro-inflammatory, immune-regulatory, and suppressive markers. While M1 macrophages are responsible for the eradication of pathogens and tumor cells, M2 macrophages have immune suppressive properties inducing an exhausted signature in CD8+ T cells as well as the tendency for tissue repair and tumor progression. (b) DCs with impaired maturation (DCLamp; DCCD83-; DCMHCII,Low) are located outside of tumor-associated tertiary lymphoid structures (NTLS) of RCC tumors and are associated with high levels of PD-L1/PD-L2 and poor prognosis. (c) T reg cells reduce the cytotoxicity of effector T cells, thereby inducing immunosuppressive phenotype in RCC tumors. (d) RCC tumors are abundant in immune cell infiltrates. In contrast to other solid tumor types, infiltration of CD8 + and CD4 + T cells into the TME is often associated with a poor prognosis and advanced tumor grade. Furthermore, inflammatory TME contains an elevated number of polyclonal exhausted CD8+Tim-3+/Lag-3+ TIL, associated with strong infiltration of dysfunctional DCs. PSP: pseudoprogression.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kruk, L.; Mamtimin, M.; Braun, A.; Anders, H.-J.; Andrassy, J.; Gudermann, T.; Mammadova-Bach, E. Inflammatory Networks in Renal Cell Carcinoma. Cancers 2023, 15, 2212. https://doi.org/10.3390/cancers15082212
AMA Style
Kruk L, Mamtimin M, Braun A, Anders H-J, Andrassy J, Gudermann T, Mammadova-Bach E. Inflammatory Networks in Renal Cell Carcinoma. Cancers. 2023; 15(8):2212. https://doi.org/10.3390/cancers15082212
Chicago/Turabian StyleKruk, Linus, Medina Mamtimin, Attila Braun, Hans-Joachim Anders, Joachim Andrassy, Thomas Gudermann, and Elmina Mammadova-Bach. 2023. "Inflammatory Networks in Renal Cell Carcinoma" Cancers 15, no. 8: 2212. https://doi.org/10.3390/cancers15082212
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.