
Virtual Screening and Quantitative Structure–Activity Relationship of Moringa oleifera with Melanoma Antigen A (MAGE-A) Genes against the Therapeutics of Non-Small Cell Lung Cancers (NSCLCs)

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Compilation of the Dataset and Ligand Preparation
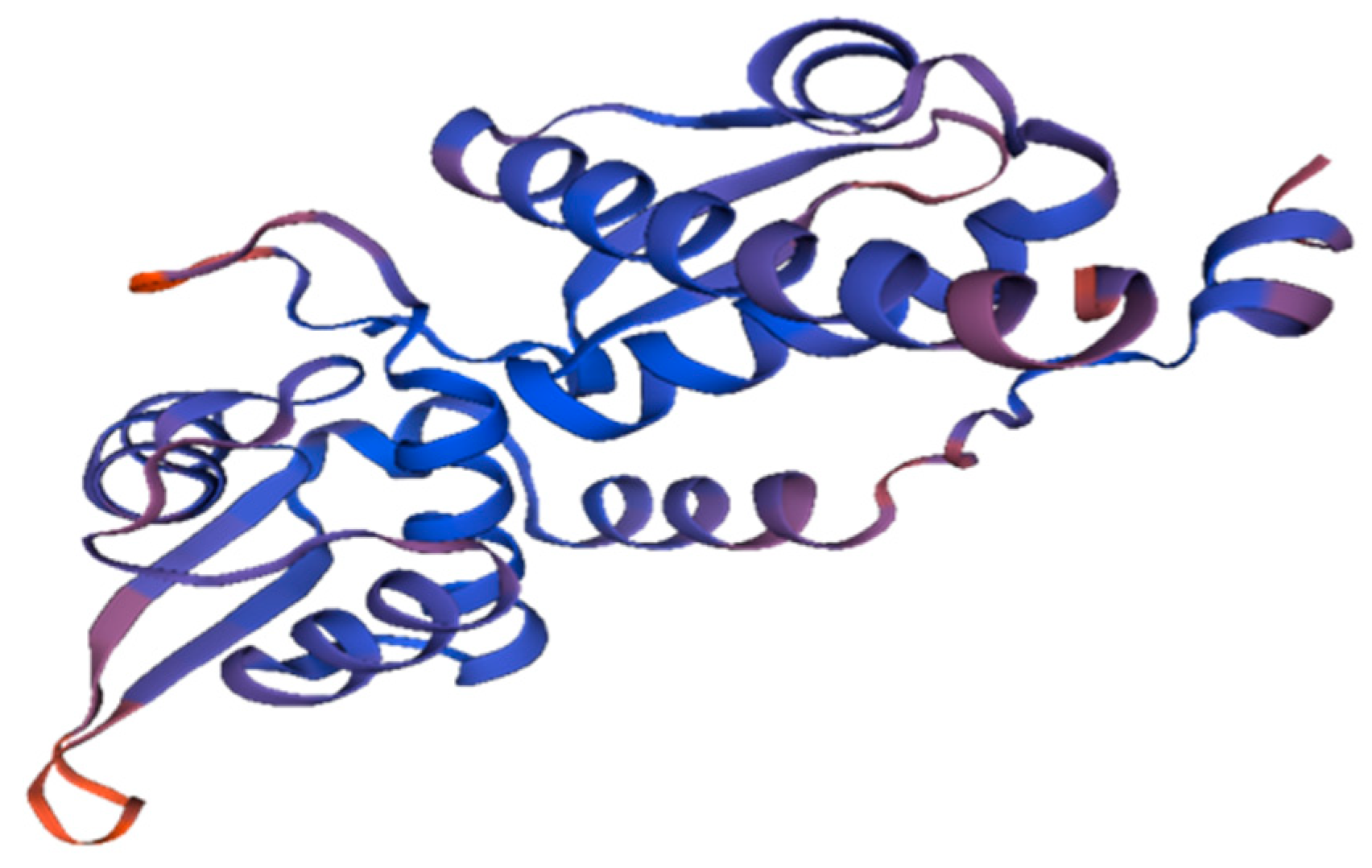
2.2. Homology Modeling of MAGE-A
2.3. Refinement of Validation of the Model
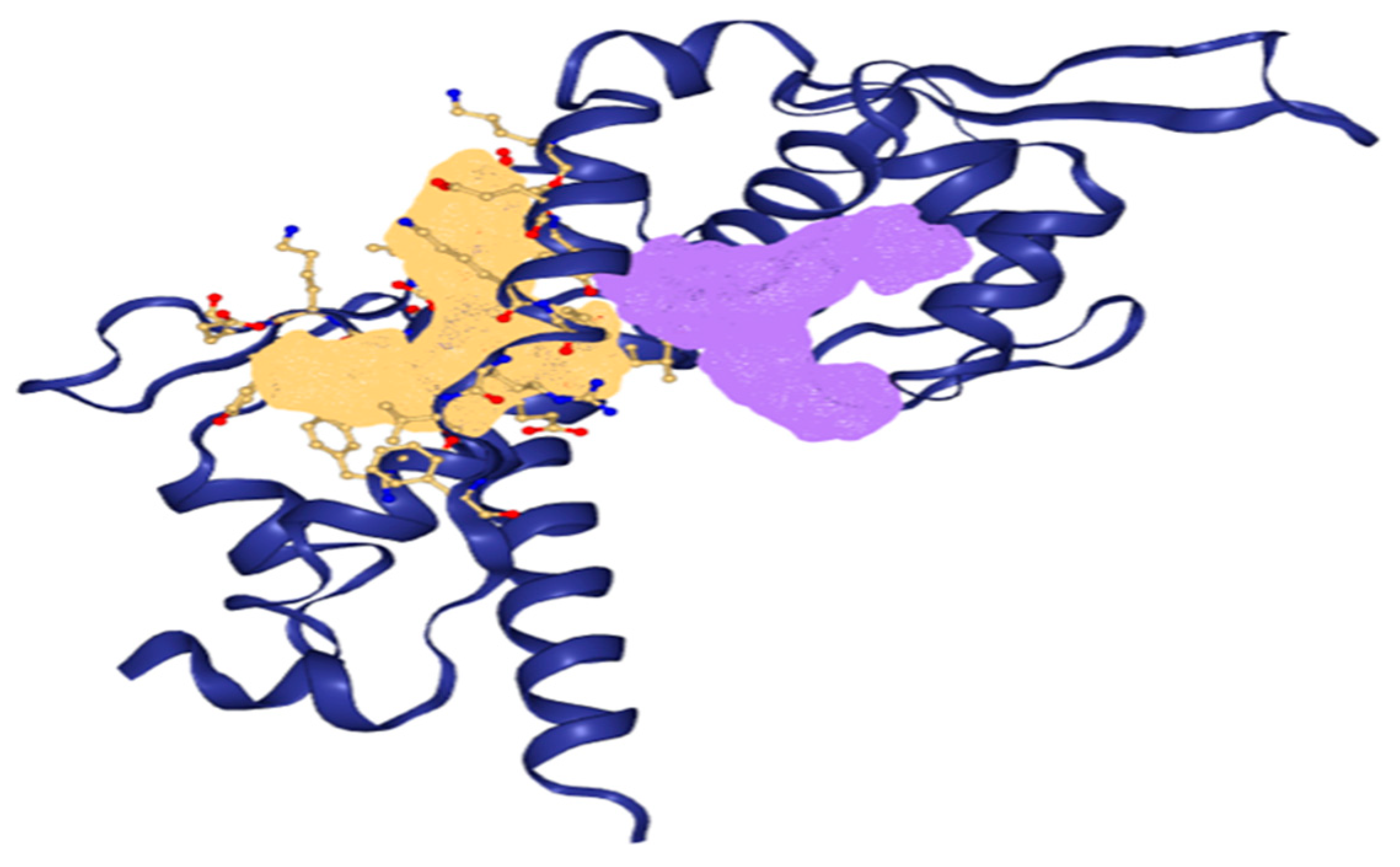
2.4. Prediction of Active Binding Site
2.5. In Silico Preclinical Testing of Phytocompounds
2.5.1. Drug Likeness, ADMET Analysis, and Prediction of Toxicity
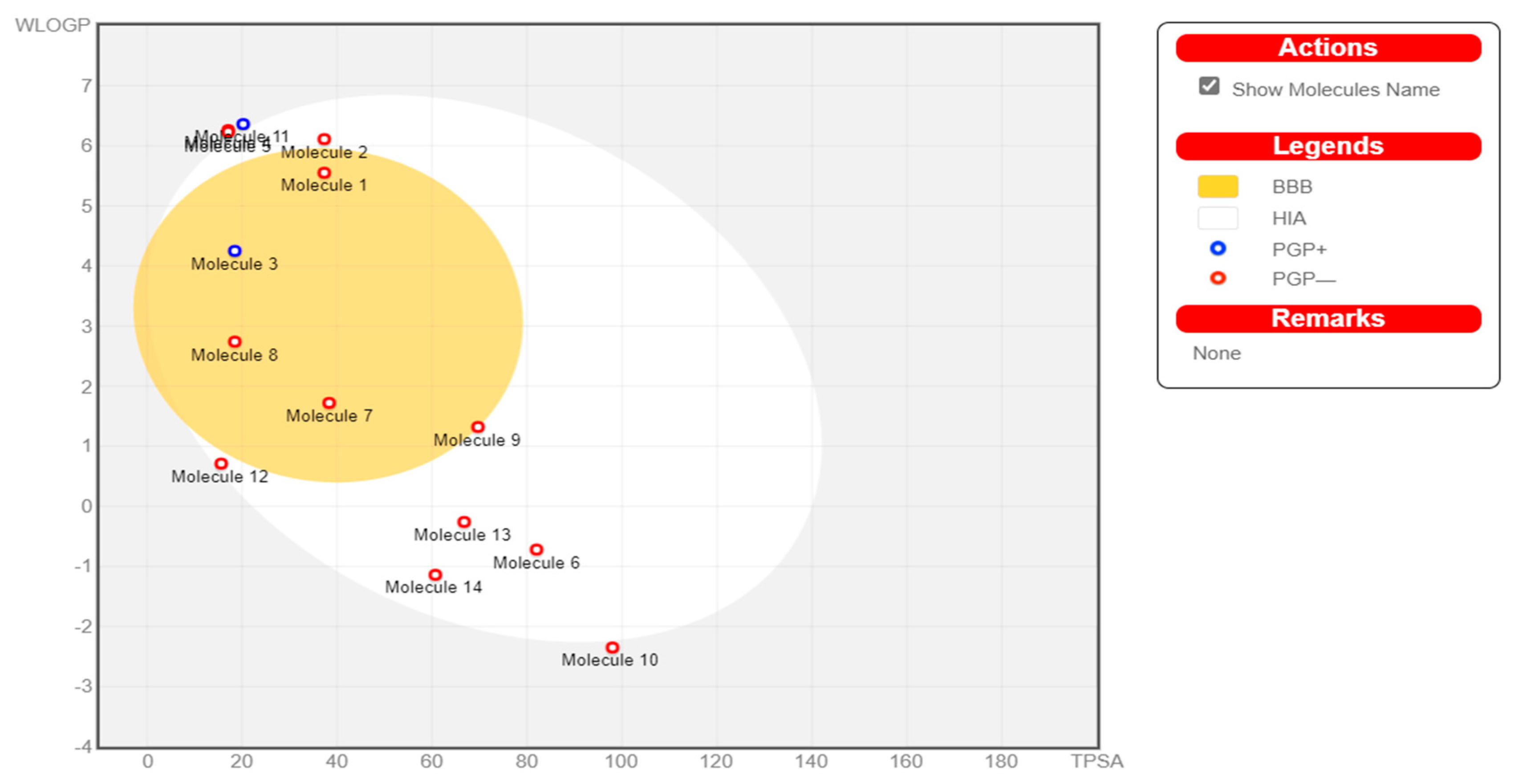
2.5.2. BOILED-Egg Analysis
2.6. Quantitative Structure–Activity Relationship Analysis
2.6.1. Collection of Datasets
2.6.2. Optimization of the Geometry
2.6.3. Molecular Descriptor Calculation
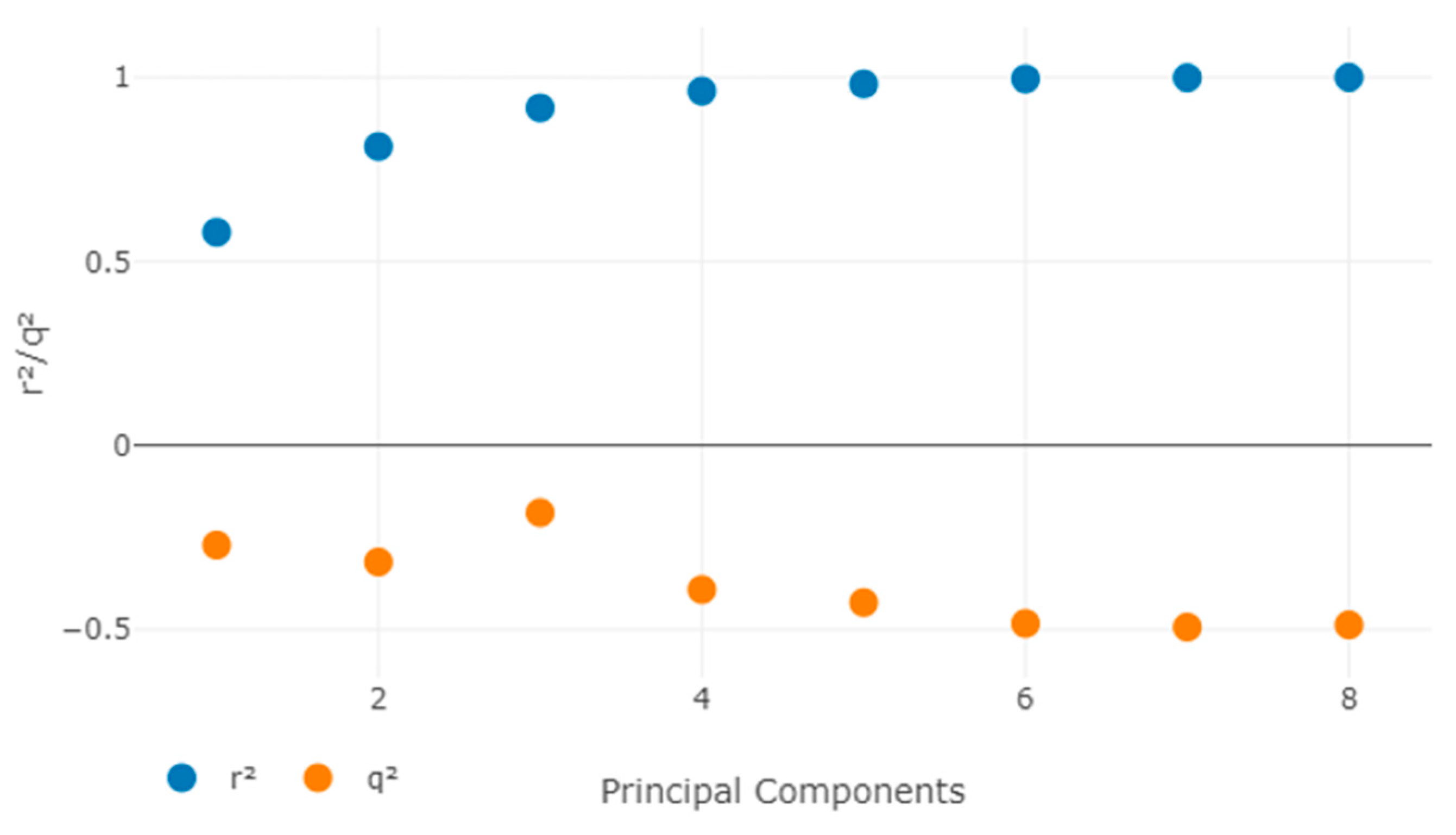
2.6.4. QSAR Model Generation
2.7. Virtual Screening
2.8. Molecular Dynamic Simulations Analysis (MD Simulations)
3. Results
3.1. Ligand Dataset
3.2. Homology Modeling
3.3. Prediction of Active Binding Site
3.4. In Silico Preclinical Testing of Phytocompounds
3.4.1. Drug Likeness, ADMET Analysis, and Prediction of Toxicity
3.4.2. Boiled-Egg Analysis
3.5. QSAR Analysis
3.5.1. Biological Activity
3.5.2. Alignment
3.5.3. C.Y Randomization Test
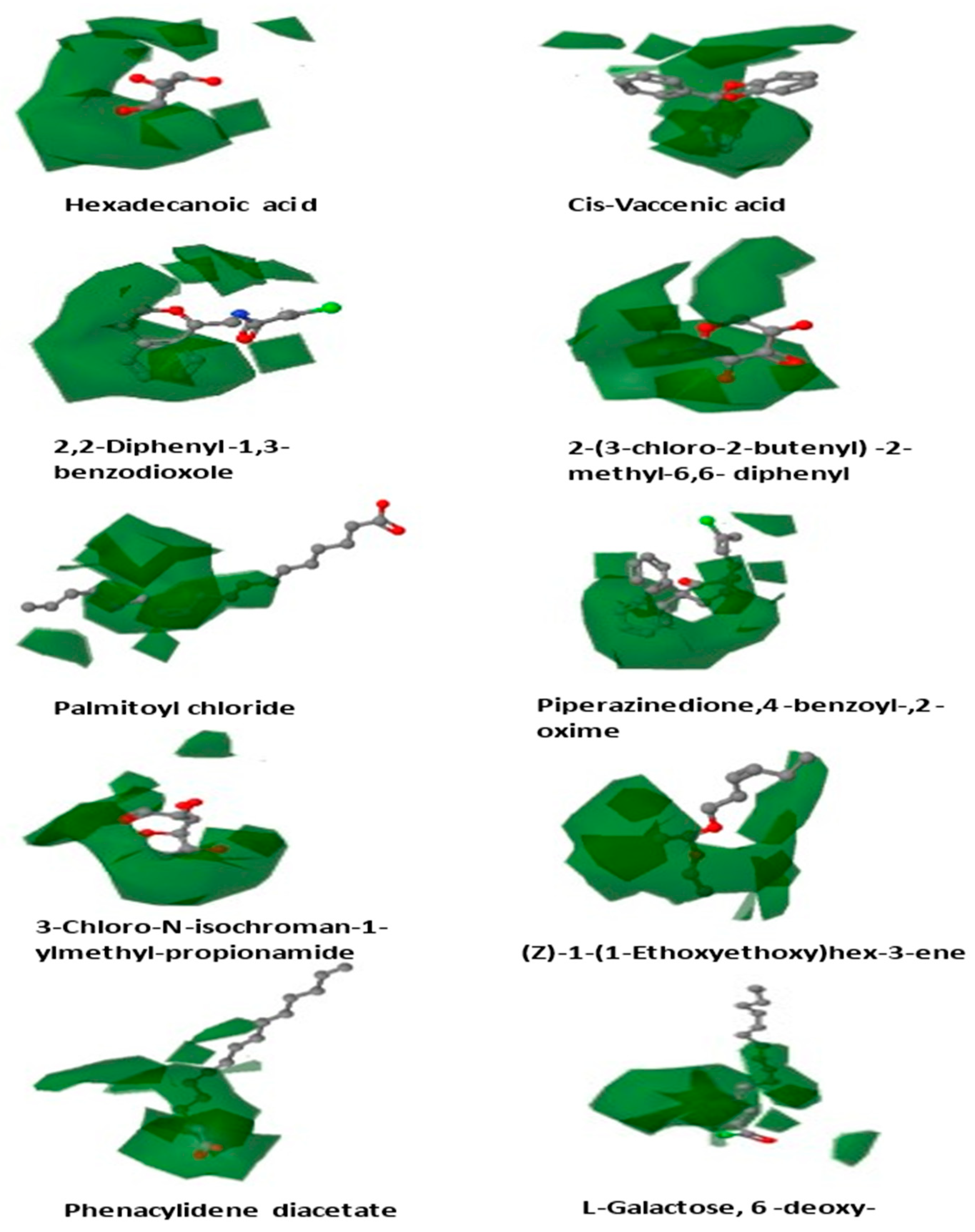
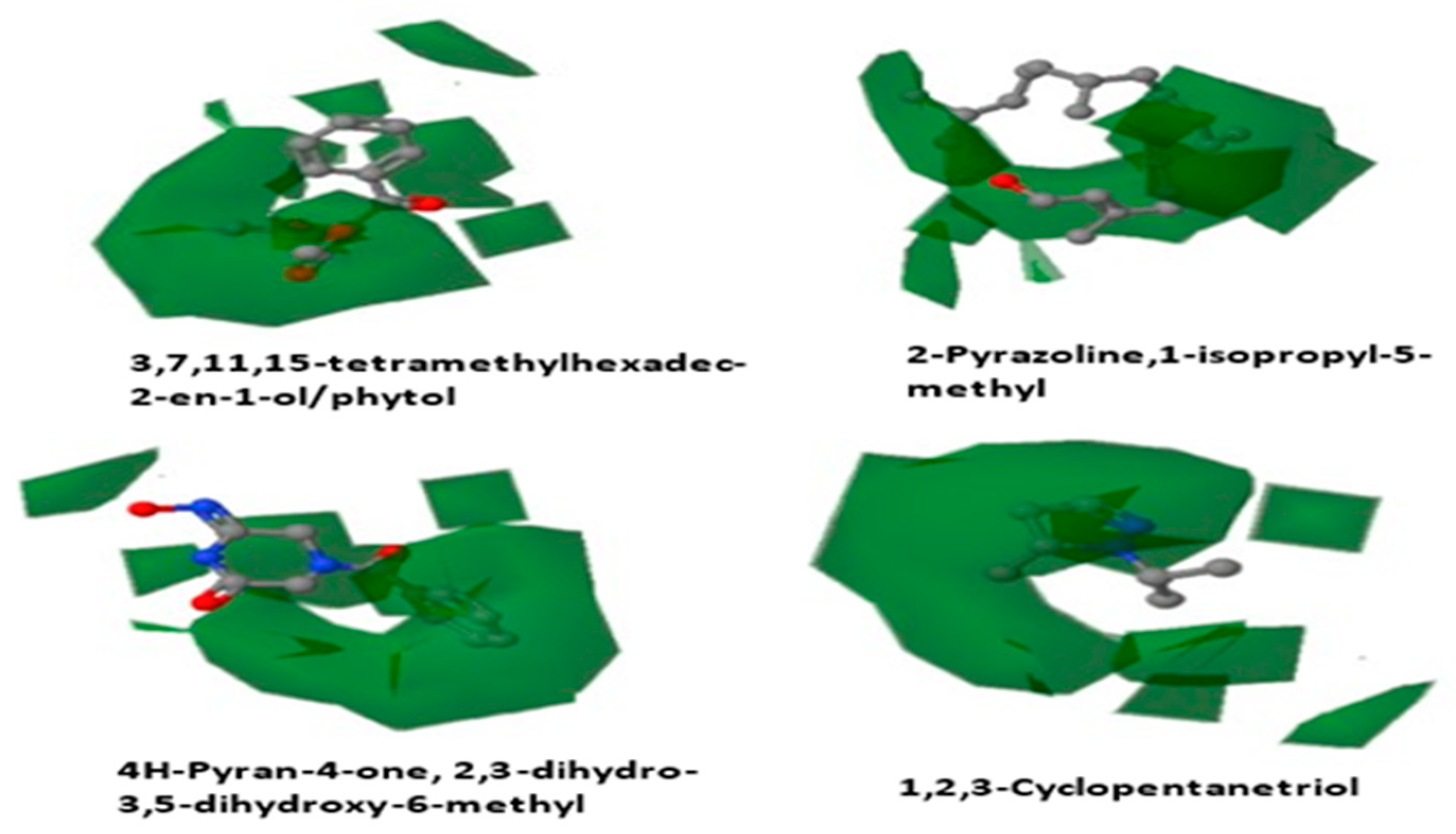
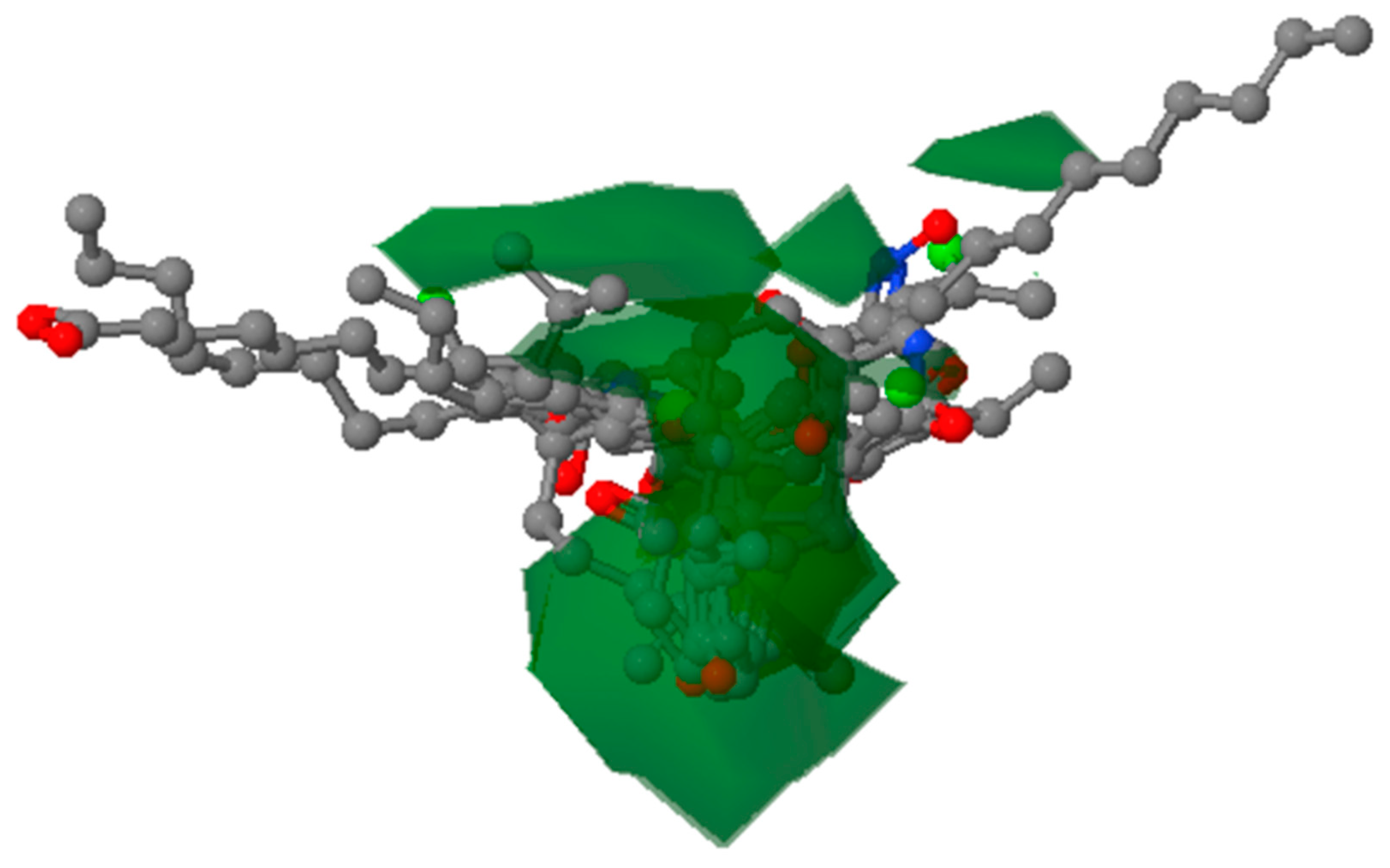
3.5.4. Contour Map Analysis
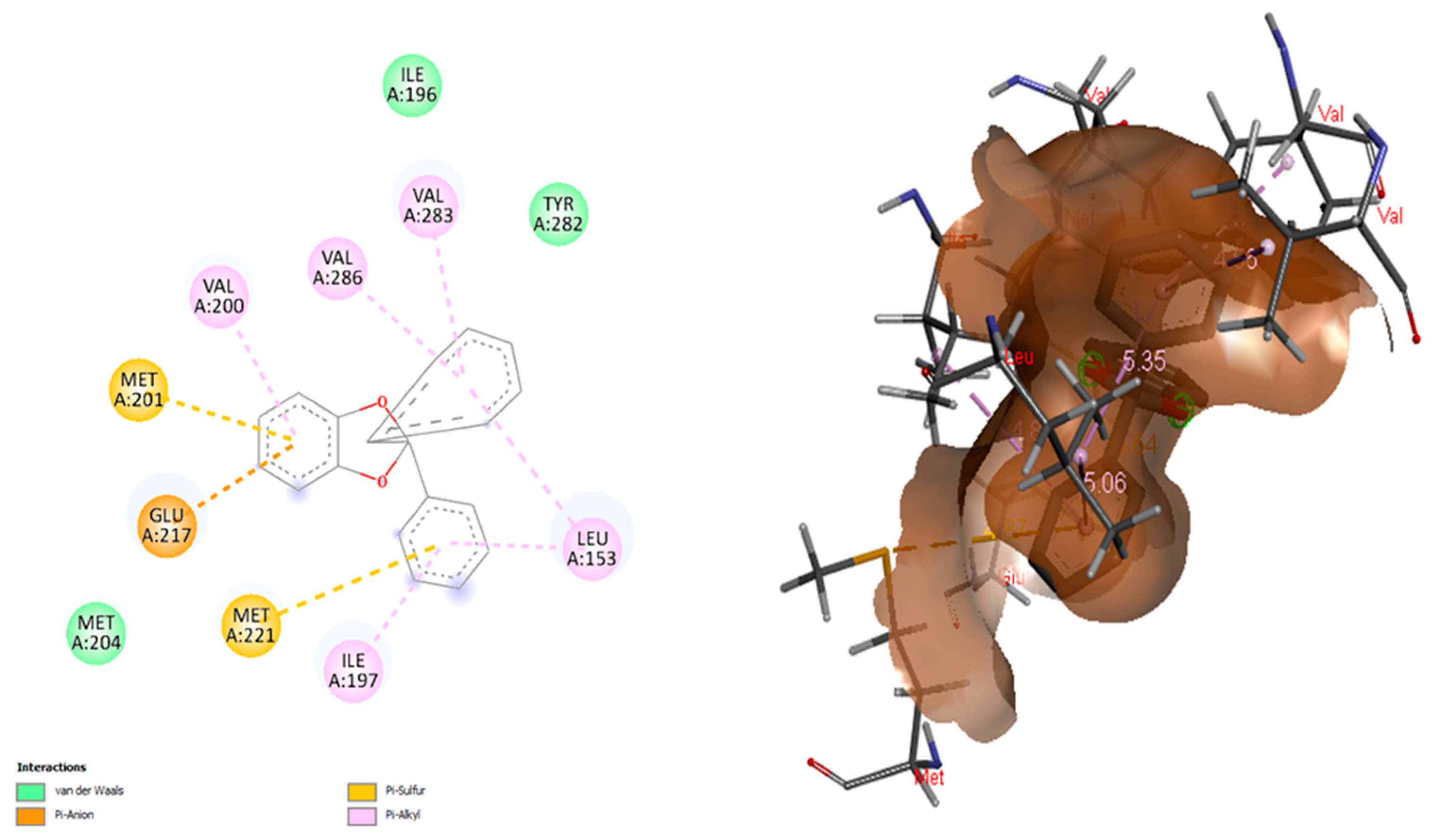
3.6. Molecular Docking and Visualization
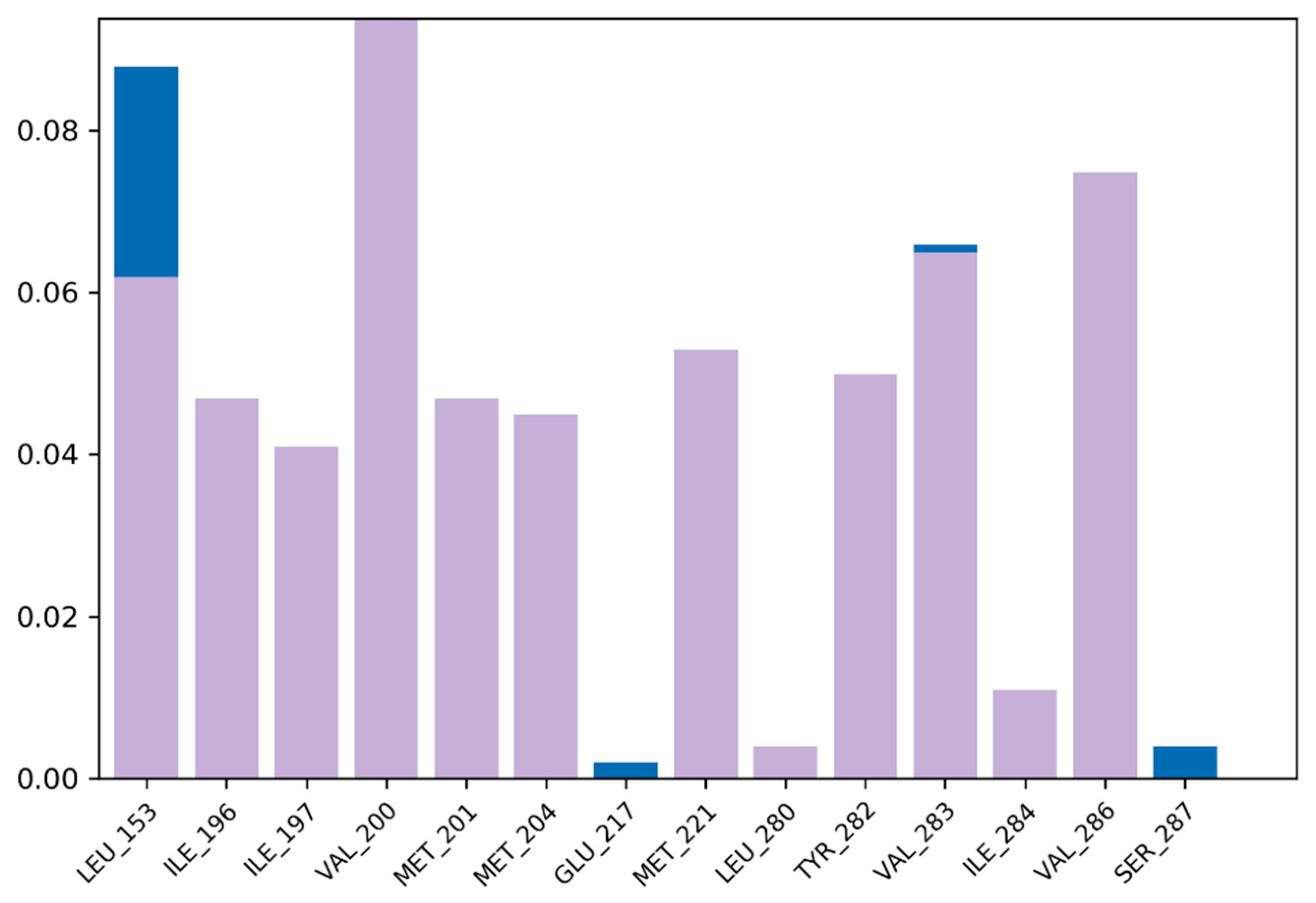
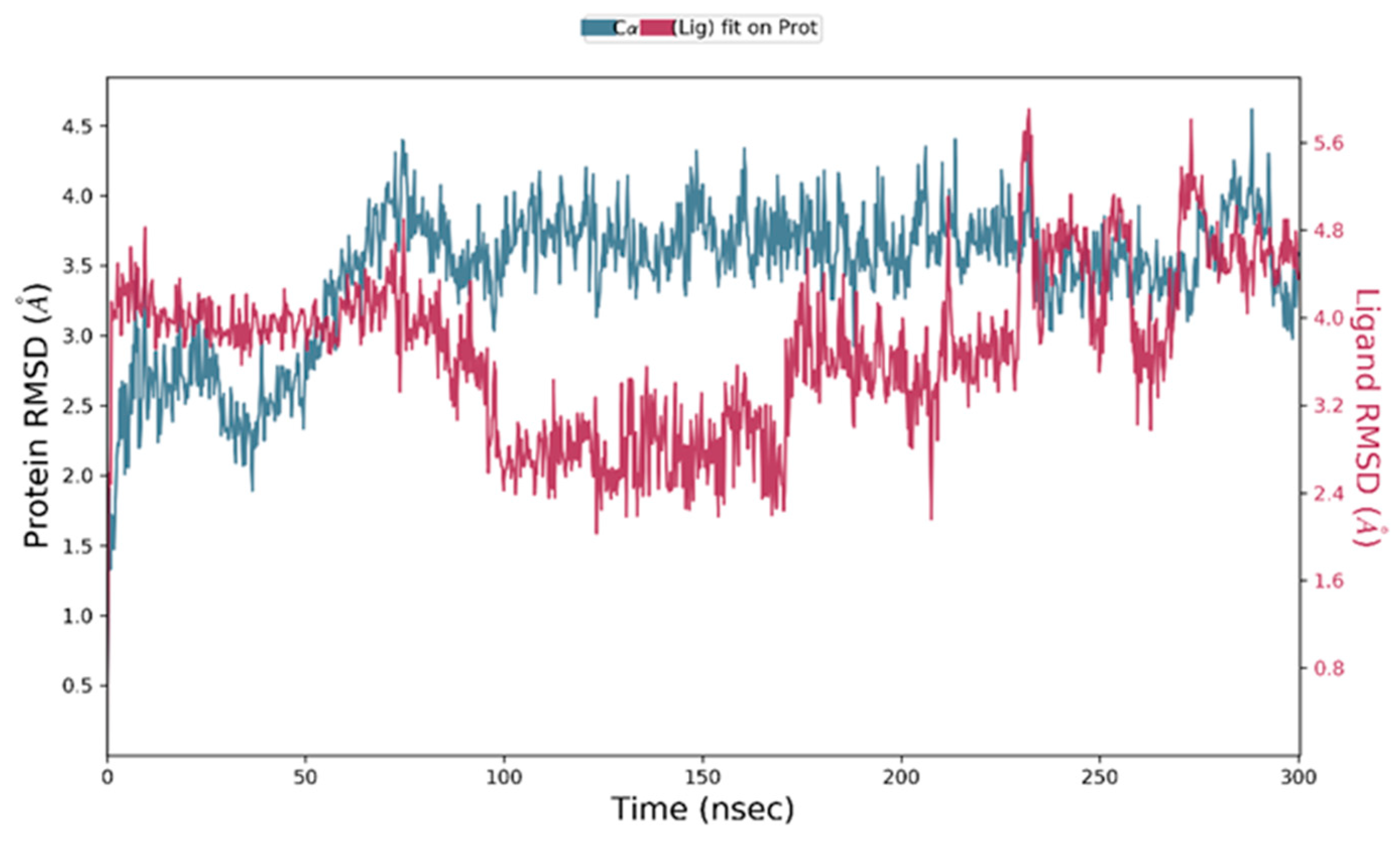
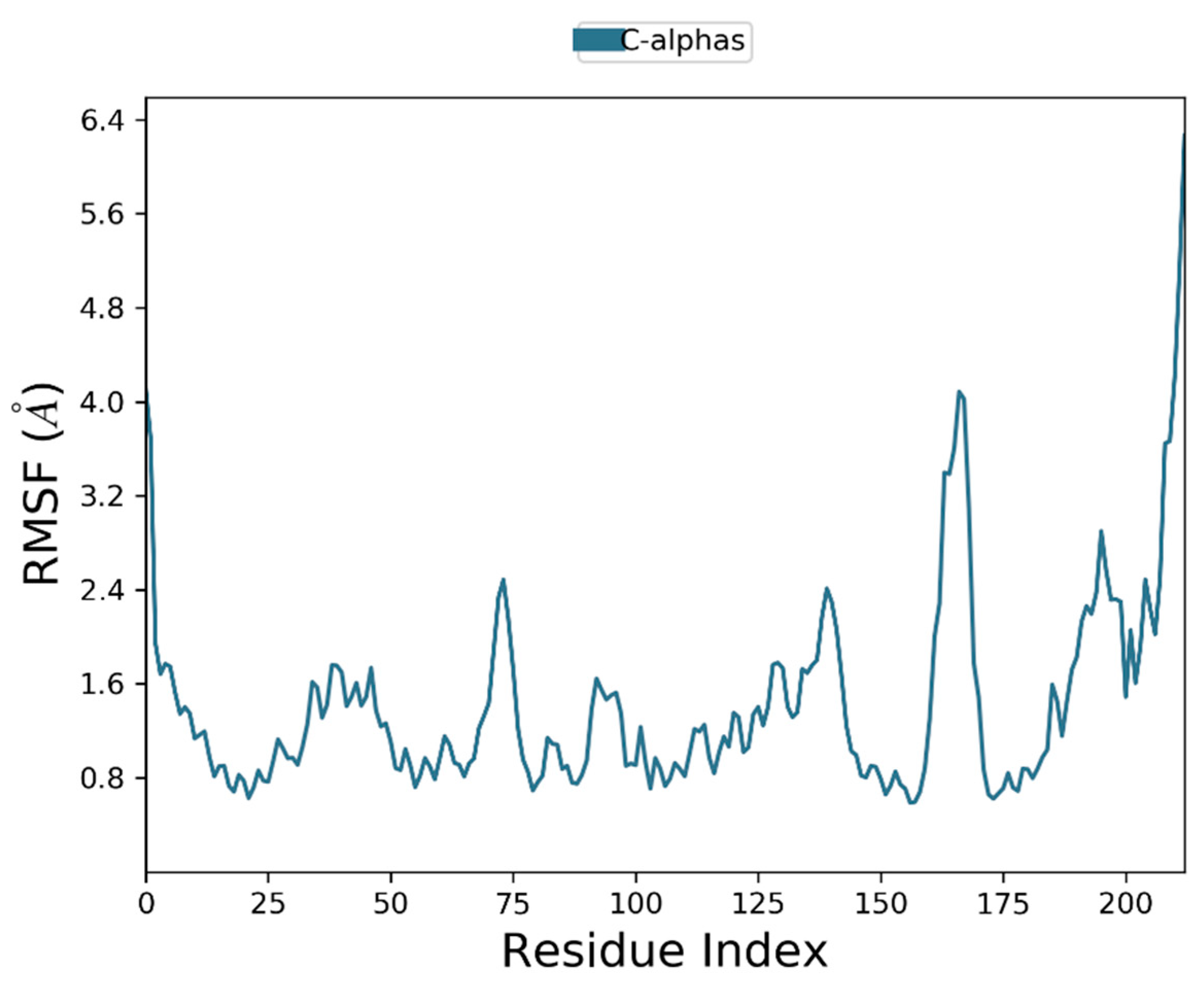
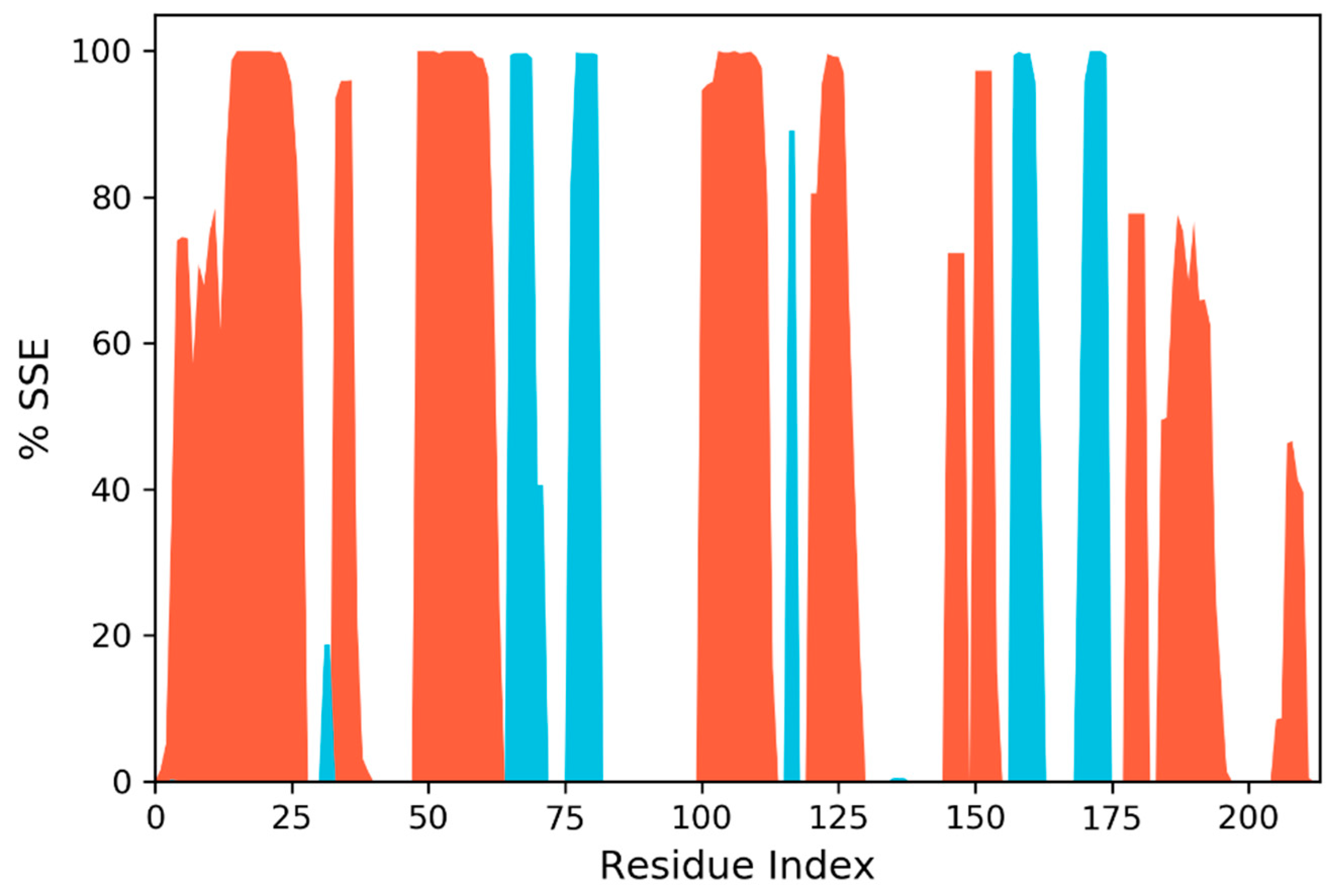
3.7. Molecular Dynamic Simulation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bade, B.C.; Cruz, C.S.D. Lung Cancer 2020: Epidemiology, Etiology, and Prevention. Clin. Chest Med. 2020, 41, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Jones, G.S.; Baldwin, D.R. Recent advances in the management of lung cancer. Clin. Med. 2018, 18, s41–s46. [Google Scholar] [CrossRef] [PubMed]
- Collins, L.G.; Haines, C.; Perkel, R.; Enck, R.E. Lung Cancer: Diagnosis and Management. Am. Fam. Physician 2007, 75, 56–63. [Google Scholar]
- Mao, Y.; Yang, D.; He, J.; Krasna, M.J. Epidemiology of Lung Cancer. Surg. Oncol. Clin. N. Am. 2016, 25, 439–445. [Google Scholar] [CrossRef] [PubMed]
- De Sousa, V.M.L.; Carvalho, L. Heterogeneity in Lung Cancer. Pathobiology 2018, 85, 96–107. [Google Scholar] [CrossRef] [PubMed]
- Relli, V.; Trerotola, M.; Guerra, E.; Alberti, S. Abandoning the Notion of Non-Small Cell Lung Cancer. Trends Mol. Med. 2019, 25, 585–594. [Google Scholar] [CrossRef] [PubMed]
- Suster, D.I.; Mino-Kenudson, M. Molecular Pathology of Primary Non-Small Cell Lung Cancer. Arch. Med. Res. 2020, 51, 784–798. [Google Scholar] [CrossRef] [PubMed]
- Alexander, M.; Kim, S.Y.; Cheng, H. Update 2020: Management of Non-Small Cell Lung Cancer. Lung 2020, 198, 897–907. [Google Scholar] [CrossRef]
- Jonna, S.; Subramaniam, D.S. Molecular Diagnostics and Targeted Therapies in Non-Small Cell Lung Cancer (NSCLC): An Update. Discov. Med. 2019, 27, 167–170. [Google Scholar]
- Schooten, E.; Di Maggio, A.; van Bergen En Henegouwen, P.M.P.; Kijanka, M.M. MAGE-A Antigens as Targets for Cancer Immunotherapy. Cancer Treat. Rev. 2018, 67, 54–62. [Google Scholar] [CrossRef]
- Sang, M.; Lian, Y.; Zhou, X.; Shan, B. MAGE-A Family: Attractive Targets for Cancer Immunotherapy. Vaccine 2011, 29, 8496–8500. [Google Scholar] [CrossRef]
- Yi, E.; Chang, J.-E.; Leem, C.; Jeon, C.-H.; Jheon, S. Association of MAGE A1-6 Expression with Lung Cancer Progression. J. Cancer 2017, 8, 1324–1329. [Google Scholar] [CrossRef] [PubMed]
- Barker, P.A.; Salehi, A. The MAGE Proteins: Emerging Roles in Cell Cycle Progression, Apoptosis, and Neurogenetic Disease. J. Neurosci. Res. 2002, 67, 705–712. [Google Scholar] [CrossRef] [PubMed]
- Simpson, A.J.G.; Caballero, O.L.; Jungbluth, A.; Chen, Y.-T.; Old, L.J. Cancer/Testis Antigens, Gametogenesis and Cancer. Nat. Rev. Cancer 2005, 5, 615–625. [Google Scholar] [CrossRef] [PubMed]
- Doyle, J.M.; Gao, J.; Wang, J.; Yang, M.; Potts, P.R. MAGE-RING Protein Complexes Comprise a Family of E3 Ubiquitin Ligases. Mol. Cell 2010, 39, 963–974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gure, A.O.; Chua, R.; Williamson, B.; Gonen, M.; Ferrera, C.A.; Gnjatic, S.; Ritter, G.; Simpson, A.J.; Chen, Y.-T.; Old, L.J.; et al. Cancer-Testis Genes Are Coordinately Expressed and Are Markers of Poor Outcome in Non–Small Cell Lung Cancer. Clin. Cancer Res. 2005, 11, 8055–8062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Zhai, X.; Wang, G.; Feng, J.; Zhu, H.; Xu, L.; Mao, G.; Huang, J. High expression of MAGE-A9 in tumor and stromal cells of non-small cell lung cancer was correlated with patient poor survival. Int. J. Clin. Exp. Pathol. 2015, 8, 541–550. [Google Scholar] [PubMed]
- Chen, X.; Wang, L.; Liu, J.; Huang, L.; Yang, L.; Gao, Q.; Shi, X.; Li, J.; Li, F.; Zhang, Z.; et al. Expression and prognostic relevance of MAGE-A3 and MAGE-C2 in non-small cell lung cancer. Oncol. Lett. 2017, 13, 1609–1618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyagi, P.; Mirakhur, B. MAGRIT: The Largest-Ever Phase III Lung Cancer Trial Aims to Establish a Novel Tumor-Specific Approach to Therapy. Clin. Lung Cancer 2009, 10, 371–374. [Google Scholar] [CrossRef] [PubMed]
- MAGE-A4c1o32T for Multi-Tumor. Available online: https://clinicaltrials.gov/ct2/show/NCT03132922 (accessed on 17 August 2022).
- Zhang, X.-W.; Liu, W.; Jiang, H.-L.; Mao, B. Chinese Herbal Medicine for Advanced Non-Small-Cell Lung Cancer: A Systematic Review and Meta-Analysis. Am. J. Chin. Med. 2018, 46, 923–952. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Guo, P.; Guo, P.; Dong, K.; Liu, F.; Wu, Y.; Li, J.; Shan, B.; Sang, M. 5-aza-2’-deoxycytidine (DAC) treatment induces the MAGE-A10 expression and improves the cytotoxicity of MAGE-A10-specific CTLs in lung cancer cells. Transl. Cancer Res. 2020, 9, 1235–1245. [Google Scholar] [CrossRef] [PubMed]
- Luo, B.; Mao, G.; Ma, H.; Chen, S. The role of seven autoantibodies in lung cancer diagnosis. J. Thorac. Dis. 2021, 13, 3660–3668. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Song, P.; Zhang, B. Expression and significance of MAGE genes in human lung cancer. Zhongguo Fei Ai Za Zhi 2013, 16, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Al-Asmari, A.K.; AlBalawi, S.M.; Athar, T.; Khan, A.Q.; Al-Shahrani, H.; Islam, M. Moringa oleifera as an Anti-Cancer Agent against Breast and Colorectal Cancer Cell Lines. PLoS ONE 2015, 10, e0135814. [Google Scholar] [CrossRef]
- Tiloke, C.; Phulukdaree, A.; A Chuturgoon, A. The antiproliferative effect of Moringa oleifera crude aqueous leaf extract on cancerous human alveolar epithelial cells. BMC Complement. Altern. Med. 2013, 13, 226. [Google Scholar] [CrossRef] [Green Version]
- Barhoi, D.; Upadhaya, P.; Barbhuiya, S.N.; Giri, A.; Giri, S. Aqueous Extract of Moringa Oleifera Exhibit Potential Anticancer Activity and Can Be Used as a Possible Cancer Therapeutic Agent: A Study Involving In Vitro and In Vivo Approach. J. Am. Coll. Nutr. 2021, 40, 70–85. [Google Scholar] [CrossRef]
- Jung, I.L.; Lee, J.H.; Kang, S.C. A Potential Oral Anticancer Drug Candidate, Moringa Oleifera Leaf Extract, Induces the Apoptosis of Human Hepatocellular Carcinoma Cells. Oncol. Lett. 2015, 10, 1597–1604. [Google Scholar] [CrossRef]
- Sadek, K.M.; Abouzed, T.K.; Abouelkhair, R.; Nasr, S. The Chemo-Prophylactic Efficacy of an Ethanol Moringa Oleifera Leaf Extract against Hepatocellular Carcinoma in Rats. Pharm. Biol. 2017, 55, 1458–1466. [Google Scholar] [CrossRef] [Green Version]
- Mumtaz, M.Z.; Kausar, F.; Hassan, M.; Javaid, S.; Malik, A. Anticancer activities of phenolic compounds from Moringa oleifera leaves: In vitro and in silico mechanistic study. Beni-Suef Univ. J. Basic Appl. Sci. 2021, 10, 1–11. [Google Scholar] [CrossRef]
- Apeh, V.O.; Asogwa, E.; Chukwuma, F.I.; Okonkwo, O.F.; Nwora, F.; Uke, R. Chemical analysis and in silico anticancer and anti-inflammatory potentials of bioactive compounds from Moringaoleifera seed oil. Adv. Tradit. Med. 2020, 22, 59–74. [Google Scholar] [CrossRef]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.A.; Yu, B.; et al. PubChem in 2021: New data content and improved web interfaces. Nucleic Acids Res. 2021, 49, D1388–D1395. [Google Scholar] [CrossRef] [PubMed]
- Kemmish, H.; Fasnacht, M.; Yan, L. Fully automated antibody structure prediction using BIOVIA tools: Validation study. PLoS ONE 2017, 12, e0177923. [Google Scholar] [CrossRef]
- O’Boyle, N.M.; Banck, M.; James, C.A.; Morley, C.; Vandermeersch, T.; Hutchison, G.R. Open Babel: An Open Chemical Toolbox. J. Cheminform. 2011, 3, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; De Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [Green Version]
- Guex, N.; Peitsch, M.C. SWISS-MODEL and the Swiss-Pdb Viewer: An Environment for Comparative Protein Modeling. Electrophoresis 1997, 18, 2714–2723. [Google Scholar] [CrossRef] [PubMed]
- Tian, W.; Chen, C.; Lei, X.; Zhao, J.; Liang, J. CASTp 3.0: Computed Atlas of Surface Topography of Proteins. Nucleic Acids Res. 2018, 46, W363–W367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sympli, H.D. Estimation of drug-likeness properties of GC–MS separated bioactive compounds in rare medicinal Pleione maculata using molecular docking technique and SwissADME in silico tools. Netw. Model. Anal. Heal. Informatics Bioinform. 2021, 10, 1–36. [Google Scholar] [CrossRef] [PubMed]
- Verma, J.; Khedkar, V.M.; Coutinho, E.C. 3D-QSAR in Drug Design—A Review. Curr. Top. Med. Chem. 2010, 10, 95–115. [Google Scholar] [CrossRef] [PubMed]
- Garro Martinez, J.C.; Vega-Hissi, E.G.; Andrada, M.F.; Estrada, M.R. QSAR and 3D-QSAR Studies Applied to Compounds with Anticonvulsant Activity. Expert Opin. Drug Discov. 2015, 10, 37–51. [Google Scholar] [CrossRef] [PubMed]
- Ruan, Z.-X.; Huangfu, D.-S.; Xu, X.-J.; Sun, P.-H.; Chen, W.-M. 3D-QSAR and Molecular Docking for the Discovery of Ketolide Derivatives. Expert Opin. Drug Discov. 2013, 8, 427–444. [Google Scholar] [CrossRef] [PubMed]
- Dallakyan, S.; Olson, A.J. Small-Molecule Library Screening by Docking with PyRx. Methods Mol. Biol. 2015, 1263, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Research, D.E.S. Schrödinger Release 2021-4: Desmond Molecular Dynamics System; Maestro-Desmond Interoperability Tools: New York, NY, USA; Schrödinger: New York, NY, USA, 2021. [Google Scholar]
- Karpf, A.R.; Bai, S.; James, S.R.; Mohler, J.L.; Wilson, E.M. Increased Expression of Androgen Receptor Coregulator MAGE-11 in Prostate Cancer by DNA Hypomethylation and Cyclic AMP. Mol. Cancer Res. 2009, 7, 523–535. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Zhou, X.; Miao, X.; Zhang, T.; Hang, X.; Tie, R.; Liu, N.; Tian, F.; Wang, F.; Yuan, J. MAGEC2, an Epithelial-Mesenchymal Transition Inducer, Is Associated with Breast Cancer Metastasis. Breast Cancer Res. Treat. 2014, 145, 23–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, Y.-H.; Doyle, J.M.; Ramanathan, S.; Gomez, T.S.; Jia, D.; Xu, M.; Chen, Z.J.; Billadeau, D.D.; Rosen, M.K.; Potts, P.R. Regulation of WASH-Dependent Actin Polymerization and Protein Trafficking by Ubiquitination. Cell 2013, 152, 1051–1064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhat, S.S.; R, S.; Prasad, S.K. In silico Screening of Violacein as an epidermal growth factor receptor inhibitor. Int. J. Heal. Allied Sci. 2022, 11, 6. [Google Scholar] [CrossRef]
- Karakoti, H.; Mahawer, S.K.; Tewari, M.; Kumar, R.; Prakash, O.; de Oliveira, M.S.; Rawat, D.S. Phytochemical Profile, In Vitro Bioactivity Evaluation, In Silico Molecular Docking and ADMET Study of Essential Oils of Three Vitex Species Grown in Tarai Region of Uttarakhand. Antioxidants 2022, 11, 1911. [Google Scholar] [CrossRef]
- Tagashira, M.; Ohtake, Y. A New Antioxidative 1,3-Benzodioxole from Melissa Officinalis. Planta Med. 1998, 64, 555–558. [Google Scholar] [CrossRef]
- Sun, Y.; Yang, Z.; Liu, Q.; Sun, X.; Chen, L.; Sun, L.; Gu, W. Design, Synthesis, and Fungicidal Evaluation of Novel 1,3-Benzodioxole-Pyrimidine Derivatives as Potential Succinate Dehydrogenase Inhibitors. J. Agric. Food Chem. 2022, 70, 7360–7374. [Google Scholar] [CrossRef] [PubMed]
- Hassan, R.; Abdallah, W.; Salman, A.; el Azzouny, A.; Aboul-Enein, M. Design, Synthesis and Anticancer Evaluation of Novel 1,3-Benzodioxoles and 1,4-Benzodioxines. Eur. J. Pharm. Sci. 2019, 139, 105045. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound No. | Compound | PubChem CID | Canonical Smiles |
|---|---|---|---|
| 1 | Hexadecanoic acid | 985 | CCCCCCCCCCCCCCCC(=O)O |
| 2 | Cis-Vaccenic acid | 5282761 | CCCCCCC=CCCCCCCCCCC(=O)O |
| 3 | 2,2-Diphenyl-1,3-benzodioxole | 343959 | C1=CC=C(C=C1)C2(OC3=CC=CC=C3O2)C4=CC=CC=C4 |
| 4 | Cyclohexanone,2-(3-chloro-2-butenyl) -2-methyl-6,6-diphenyl | 57116063 | CC(=CCC1(CCCC(C1=O)(C2=CC=CC=C2)C3=CC=CC=C3)C)Cl |
| 5 | Palmitoyl chloride | 8206 | CCCCCCCCCCCCCCCC(=O)Cl |
| 6 | Piperazinedione,4-benzoyl-,2-oxime | 135594388 | C1C(=NO)NC(=O)CN1C(=O)C2=CC=CC=C2 |
| 7 | 3-Chloro-N-isochroman-1-ylmethyl-propionamide | 583868 | C1COC(C2=CC=CC=C21)CNC(=O)CCCl |
| 8 | (Z)-1-(1-Ethoxyethoxy)hex-3-ene | 108504 | CCC=CCCOC(C)OCC |
| 9 | Phenacylidene diacetate | 569561 | CC(=O)OC(C(=O)C1=CC=CC=C1)OC(=O)C |
| 10 | L-Galactose, 6-deoxy-2-O-methyl- | 169586 | CC(C(C(C(C=O)O)O)O)O |
| 11 | 3,7,11,15-tetramethylhexadec-2-en-1-ol/phytol | 145386 | CC(C)CCCC(C)CCCC(C)CCCC(=CCO)C |
| 12 | 2-Pyrazoline,1-isopropyl-5-methyl | 573933 | CC1CC=NN1C(C)C |
| 13 | 4H-Pyran-4-one, 2,3-dihydro-3,5-dihydroxy-6-methyl | 119838 | CC1=C(C(=O)C(CO1)O)O |
| 14 | 1,2,3-Cyclopentanetriol | 92530 | C1CC(C(C1O)O)O |
| Pocket Name | Volume Å3 | Surface Å2 | Drug Score | H-Bond Acceptors | Hydrophobic Interactions | Pocket Atoms | Apolar Amino Acid Ratio | Polar Amino Acid Ratio |
|---|---|---|---|---|---|---|---|---|
| Pocket 1 | 561.86 | 872.29 | 0.77 | 40 | 47 | 124 | 0.33 | 0.33 |
| Pocket 2 | 517.12 | 764.62 | 0.77 | 37 | 49 | 125 | 0.56 | 0.28 |
| Compound | Molecular Weight | Heavy Atoms | Aromatic Heavy Atoms | TPSA |
|---|---|---|---|---|
| Hexadecanoic acid | 256.42 | 18 | 0 | 37.3 |
| Cis-Vaccenic acid | 282.46 | 20 | 0 | 37.3 |
| 2,2-Diphenyl-1,3-benzodioxole | 274.31 | 21 | 18 | 18.46 |
| Cyclohexanone,2-(3-chloro-2-butenyl) 2-methyl-6,6-diphenyl | 352.9 | 25 | 12 | 17.07 |
| Palmitoyl chloride | 274.87 | 18 | 0 | 17.07 |
| Piperazinedione,4-benzoyl-,2-oxime | 233.22 | 17 | 6 | 82 |
| 3-Chloro-N-isochroman-1-ylmethyl-propionamide | 253.72 | 17 | 6 | 38.33 |
| (Z)-1-(1-Ethoxyethoxy)hex-3-ene | 172.26 | 12 | 0 | 18.46 |
| Phenacylidene diacetate | 236.22 | 17 | 6 | 69.67 |
| L-Galactose, 6-deoxy | 164.16 | 11 | 0 | 97.99 |
| 3,7,11,15-tetramethylhexadec-2-en-1-ol/phytol | 296.53 | 21 | 0 | 20.23 |
| 2-Pyrazoline, 1-isopropyl-5-methyl | 126.2 | 9 | 0 | 15.6 |
| 4H-Pyran-4-one, 2,3-dihydro-3,5-dihydroxy-6-methyl | 144.13 | 10 | 0 | 66.76 |
| 1,2,3-Cyclopentanetriol | 118.13 | 8 | 0 | 60.69 |
| Compound | Molecular Weight | H-Bond Acceptors | H-Bond Donors | Molar Refractivity | Consensus Log P |
|---|---|---|---|---|---|
| Hexadecanoic acid | 256.42 | 2 | 1 | 80.8 | 5.2 |
| Cis-Vaccenic acid | 282.46 | 2 | 1 | 89.94 | 5.68 |
| 2,2-Diphenyl-1,3-benzodioxole | 274.31 | 2 | 0 | 81.36 | 4.25 |
| Cyclohexanone,2-(3-chloro-2-butenyl) -2-methyl-6,6-diphenyl | 352.9 | 1 | 0 | 106 | 5.62 |
| Palmitoyl chloride | 274.87 | 1 | 0 | 84.02 | 5.96 |
| Piperazinedione,4-benzoyl-,2-oxime | 233.22 | 4 | 2 | 67.09 | 0.45 |
| 3-Chloro-N-isochroman-1-ylmethyl-propionamide | 253.72 | 2 | 1 | 67.18 | 2.06 |
| (Z)-1-(1-Ethoxyethoxy)hex-3-ene | 172.26 | 2 | 0 | 51.88 | 2.6 |
| Phenacylidene diacetate | 236.22 | 5 | 0 | 58.43 | 1.62 |
| L-Galactose, 6-deoxy- | 164.16 | 5 | 4 | 35.8 | –1.49 |
| 3,7,11,15-tetramethylhexadec-2-en-1-ol/phytol | 296.53 | 1 | 1 | 98.94 | 6.22 |
| 2-Pyrazoline,1-isopropyl-5-methyl | 126.2 | 1 | 0 | 47.47 | 1.29 |
| 4H-Pyran-4-one, 2,3-dihydro-3,5-dihydroxy-6-methyl | 144.13 | 4 | 2 | 32.39 | –0.22 |
| 1,2,3-Cyclopentanetriol | 118.13 | 3 | 3 | 27.52 | –0.58 |
| Molecule Number | Compound | GI Absorption | BBB Permeability | Pgp Substrate | Silicos-IT LogSw | Silicos-IT Class |
|---|---|---|---|---|---|---|
| 1 | Hexadecanoic acid | High | Yes | No | –5.31 | Moderately soluble |
| 2 | Cis-Vaccenic acid | High | No | No | –5.39 | Moderately soluble |
| 3 | 2,2-Diphenyl-1,3-benzodioxole | High | Yes | Yes | –7.17 | Poorly soluble |
| 4 | Cyclohexanone,2-(3-chloro-2-butenyl) -2-methyl-6,6-diphenyl | Low | No | No | –8.29 | Poorly soluble |
| 5 | Palmitoyl chloride | Low | No | No | –6.5 | Poorly soluble |
| 6 | Piperazinedione,4-benzoyl-,2-oxime | High | No | No | –2.2 | Soluble |
| 7 | 3-Chloro-N-isochroman-1-ylmethyl-propionamide | High | Yes | No | –4.57 | Moderately soluble |
| 8 | (Z)-1-(1-Ethoxyethoxy)hex-3-ene | High | Yes | No | –2.27 | Soluble |
| 9 | Phenacylidene diacetate | High | Yes | No | –2.56 | Soluble |
| 10 | L-Galactose, 6-deoxy- | Low | No | No | 1.9 | Soluble |
| 11 | 3,7,11,15-tetramethylhexadec-2-en-1-ol/phytol | Low | No | Yes | –5.51 | Moderately soluble |
| 12 | 2-Pyrazoline,1-isopropyl-5-methyl | High | No | No | –0.8 | Soluble |
| 13 | 4H-Pyran-4-one, 2,3-dihydro-3,5-dihydroxy-6-methyl | High | No | No | 0.15 | Soluble |
| 14 | 1,2,3-Cyclopentanetriol | High | No | No | 1.2 | Soluble |
| Compound | Toxic Dose (LD50 Values in mg/kg Body Weight) | Toxicity Class | Hepatotoxicity |
|---|---|---|---|
| Hexadecanoic acid | 900 | Class IV | No |
| Cis-Vaccenic acid | 48 | Class II | No |
| 2,2-Diphenyl-1,3-benzodioxole | 720 | Class IV | No |
| Cyclohexanone,2-(3-chloro-2-butenyl) -2-methyl-6,6-diphenyl | 750 | Class IV | No |
| Palmitoyl chloride | 400 | Class IV | No |
| Piperazinedione,4-benzoyl-,2-oxime | 6800 | Class VI | No |
| 3-Chloro-N-isochroman-1-ylmethyl-propionamide | 380 | Class IV | No |
| (Z)-1-(1-Ethoxyethoxy)hex-3-ene | 5000 | Class V | No |
| Phenacylidene diacetate | 5000 | Class V | No |
| L-Galactose, 6-deoxy | 23,000 | Class VI | No |
| 3,7,11,15-tetramethylhexadec-2-en-1-ol/phytol | 5000 | Class V | No |
| 2-Pyrazoline,1-isopropyl-5-methyl | 800 | Class IV | No |
| 4H-Pyran-4-one, 2,3-dihydro-3,5-dihydroxy-6-methyl | 595 | Class IV | No |
| 1,2,3-Cyclopentanetriol | 12,500 | Class VI | No |
| Property | GM Label | GM ID | GM Conf | Longest Label | Longest ID | Longest Conf |
|---|---|---|---|---|---|---|
| Least Active | 2,2-Diphenyl-1,3-benzodioxole | 3623329 | 0 | 2,2-Diphenyl-1,3-benzodioxole | 3623329 | 2 |
| Most Active | Cyclohexanone,2-(3-chloro-2-butenyl) -2-methyl-6,6-diphenyl | 3623333 | 0 | Cyclohexanone,2-(3-chloro-2-butenyl) -2-methyl-6,6-diphenyl | 3623333 | 6 |
| Heaviest | Cyclohexanone,2-(3-chloro-2-butenyl) -2-methyl-6,6-diphenyl | 3623333 | 0 | Cyclohexanone,2-(3-chloro-2-butenyl) -2-methyl-6,6-diphenyl | 3623333 | 6 |
| Longest | Palmitoyl chloride | 3623337 | 0 | Cis-Vaccenic acid | 3623332 | 12 |
| Most Flexible | Phytol | 3623339 | 0 | Phytol | 3623339 | 12 |
| Most Rigid | 2,2-Diphenyl-1,3-benzodioxole | 3623329 | 0 | 2,2-Diphenyl-1,3-benzodioxole | 3623329 | 2 |
| Least Polar | Pyrazoline | 3623341 | 0 | Pyrazoline | 3623341 | 0 |
| Most Polar | Fucose | 3623334 | 0 | Fucose | 3623334 | 0 |
| PC | r2 | SDEC | q2 | SDEP |
|---|---|---|---|---|
| 1 | 0.579 | 0.203 | –0.271 | 0.354 |
| 2 | 0.812 | 0.136 | –0.317 | 0.36 |
| 3 | 0.917 | 0.091 | –0.183 | 0.341 |
| 4 | 0.963 | 0.06 | –0.392 | 0.37 |
| 5 | 0.982 | 0.042 | –0.427 | 0.375 |
| 6 | 0.996 | 0.02 | –0.484 | 0.382 |
| 7 | 0.999 | 0.009 | –0.494 | 0.383 |
| 8 | 1 | 0.005 | –0.488 | 0.383 |
| Sl No | Name | Experimental Activity |
|---|---|---|
| 1 | 1,2,3-Cyclopentanetriol | 7.63827 |
| 2 | 2,2-Diphenyl-1,3-benzodioxole | 7.63827 |
| 3 | 3-Chloro-N-isochroman-1-ylmethyl-propionamide | 7.33724 |
| 4 | 4H-Pyran-4-one, 2,3-dihydro-3,5-dihydroxy-6-methyl | 8.22185 |
| 5 | Cis-Vaccenic acid | 8.22185 |
| 6 | Cyclohexanone,2-(3-chloro-2-butenyl) -2-methyl-6,6-diphenyl | 7.92082 |
| 7 | L-Galactose, 6-deoxy- | 8.22185 |
| 8 | (Z)-1-(1-Ethoxyethoxy)hex-3-ene | 8.22185 |
| 9 | hexadecanoic acid | 8.22185 |
| 10 | Palmitoyl chloride | 7.49485 |
| Compound | Binding Affinity |
|---|---|
| 3-Chloro-N-isochroman-1-ylmethyl-propionamide | –5.5 |
| (Z)-1-(1-Ethoxyethoxy)hex-3-ene | –3.3 |
| 4H-Pyran-4-one, 2,3-dihydro-3,5-dihydroxy-6-methyl | –3.8 |
| Cis-Vaccenic acid | –4 |
| 2-(3-chloro-2-butenyl) -2-methyl-6,6-diphenyl | –6.3 |
| 1,2,3-Cyclopentanetriol | –3.6 |
| 2,2-Diphenyl-1,3-benzodioxole | –7.3 |
| L-Galactose, 6-deoxy- | –3.4 |
| Hexadecanoic acid | –4.1 |
| Palmitoyl chloride | –4.1 |
| Phenacylidene diacetate | –4.9 |
| 3,7,11,15-tetramethylhexadec-2-en-1-ol | –4.3 |
| 2-Pyrazoline,1-isopropyl-5-methyl | –3.6 |
| Piperazinedione,4-benzoyl-,2-oxime | –4.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhat, S.S.; Mahapatra, S.D.; R, S.; Sommano, S.R.; Prasad, S.K. Virtual Screening and Quantitative Structure–Activity Relationship of Moringa oleifera with Melanoma Antigen A (MAGE-A) Genes against the Therapeutics of Non-Small Cell Lung Cancers (NSCLCs). Cancers 2022, 14, 5052. https://doi.org/10.3390/cancers14205052
Bhat SS, Mahapatra SD, R S, Sommano SR, Prasad SK. Virtual Screening and Quantitative Structure–Activity Relationship of Moringa oleifera with Melanoma Antigen A (MAGE-A) Genes against the Therapeutics of Non-Small Cell Lung Cancers (NSCLCs). Cancers. 2022; 14(20):5052. https://doi.org/10.3390/cancers14205052
Chicago/Turabian StyleBhat, Smitha S., Shreya Das Mahapatra, Sindhu R, Sarana Rose Sommano, and Shashanka K. Prasad. 2022. "Virtual Screening and Quantitative Structure–Activity Relationship of Moringa oleifera with Melanoma Antigen A (MAGE-A) Genes against the Therapeutics of Non-Small Cell Lung Cancers (NSCLCs)" Cancers 14, no. 20: 5052. https://doi.org/10.3390/cancers14205052